
Abstract
Throughout Europe the blue mussel Mytilus edulis is dominant in the north while the Mediterranean mussel Mytilus galloprovincialis prevails in the south. Studies from the 1970s to the late 2000′s documented the northward range expansion of M. galloprovincialis in Europe and predicted this trend to continue with climate change. The objectives of this study were to sample predominantly wild mussels (n = 1459) at twenty-four Irish intertidal sites over a seven year period and at three Welsh sites to investigate (a) the abundance and distribution of Mytilus spp., identified molecularly by polymerase chain reaction (PCR), and (b) compare with historical observations, at Irish sites where M. galloprovincialis was observed to be more abundant than M. edulis. Mussels were sampled more than once at certain sites to investigate if there was a temporal effect. The findings of this study indicated that M. edulis was consistently the most abundant species, followed by hybrids and M. galloprovincialis. At certain sites, hybrids were detected while M. galloprovincialis was absent. This finding may indicate transient M. galloprovinicialis populations or input from individuals subtidally. One factor that may be of importance was the anomalous cold winter “polar snaps” (2010 and 2011) that occurred during this study. In addition, heavy precipitation events and subsequent increased freshwater loading in bays and estuaries also occurred during this time period. Future warming climate scenarios have been predicted to facilitate the northwards establishment of M. galloprovincialis, however, nearshore meteorological extremes may have an impact in its larval settlement, establishment and subsequent reproductive output.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Due to their widespread distribution, ecological importance and commercial value, Mytilus spp. are one of the most studied bivalves (Dailianis 2010). Throughout Europe the dominant species of mussels present are the blue mussel Mytilus edulis to the north and the Mediterranean mussel Mytilus galloprovincialis to the south. M. edulis are boreo-temperate in their distribution on both coastlines of the Atlantic Ocean and are found intertidally, with a distribution in the eastern Atlantic occurring from Norway (Christiansen 1965) to the border of France and Spain (Sanjuan et al. 1994). M. galloprovincialis is native to the Mediterranean, Black and Adriatic Seas and has expanded its range on the Atlantic coast from Ireland to Morocco (Branch and Stephanni 2004). It is listed in the “World’s worst 100 invasive species” where it has outcompeted certain native mussel species and other bivalve species worldwide (Branch and Steffani 2004; Ozer and Guneydag 2014) and is categorized as a non-indigenous species (NIS) in the British Isles (GISD 2017), having first being reported in England in the 1950s (Hepper 1957; Lewis and Seed 1969). M. galloprovincialis was first reported to have expanded its range into Ireland in the 1970s (Gosling and Wilkins 1977).
Mytilus galloprovincialis outcompetes the native mussel species due to its faster growth rates, being more tolerant to air exposure and having a higher reproductive output (between 20 and 200% higher output) then the native mussel species (Branch and Steffani 2004). In populations found in the subtidal zones, M. galloprovincialis can grow to full size in a short period of time with the growth rate being roughly 50 mm shell length within approximately six to eight months (Page and Hubbard 1987) and is categorized as an invasive species in the British Isles (GISD 2017). Unlike M. edulis, which can tolerate a range of temperature changes (5–20 °C) (Zagata et al. 2008), M. galloprovincialis has a more constrained temperature range from 10–20 °C (FAO 2017). Lenz et al. (2018) investigated the effect of heat stress in five mussel species, including M. edulis, that would occur during transportation in regions with high sea surface temperatures when those species were on ship hulls or in ballast water tanks. That study observed that thermal tolerance was significantly enhanced in M. edulis when this species was exposed to a previous heat stress experience. Many studies have shown that M. edulis can tolerate wide fluctuations in salinity, desiccation and temperature, which has led to their wide distributional pattern (Tyler-Walters and Seed 2006; FAO 2017). M. galloprovincialis is generally cultured at higher salinities (< 34) and mussel production can be affected by low salinity, which kills mussels on the upper 0.5 to 1 m of the ropes (FAO 2017). In Ireland, M. edulis is predominantly found at sheltered sites (Gosling and Wilkins 1981; Gosling et al. 2008) and on wave-exposed Irish shores it intermixes with M. galloprovincialis in varying proportions and this may also be seen worldwide (Gosling and Wilkins 1981; Gosling 2004). Gosling et al. (2008) observed that for the sixteen Atlantic sites on the south, west and north Irish coasts M. galloprovincialis and hybrids were detected while M. edulis was exclusive to east coast sites in the Irish Sea. In that study, it was concluded that M. galloprovincialis was increasing, not just at exposed sites but more strikingly at sheltered wave protected locations.
Larval dispersal of both mytilid species is influenced by the speed and direction of surface currents generated by the wind, and M. galloprovincialis has successfully established populations worldwide in temperate regions, in particular, where shipping ports are present (Branch and Stephanni 2004; Braby and Somero 2006). Mussel larvae are planktotrophic and can remain in the water column for up to several months before settling out (Bayne 1964; Seed 1969). A “regime shift” caused by climate change during the last century in the north Atlantic (Jones et al. 2001) has resulted in the northward migration of cold-water species of phytoplankon and zooplankton and warm-water species, including the larvae that feed on them, following them. In the North Sea, several demersal fish species have shown northward distributional shifts as a response to global warming (Perry et al. 2005). Lima et al. (2007) evaluated the direction of range shifts of macroalgae spp. based on historical data along the Portuguese coast and observed that cold-water species showed no particular shifting trend while all warm-water species, with shifting distributions, expanded their range northwards. More recently, poleward advances of subtropical species into the European theatre referred to as "African Creep" are occurring in the Eastern Atlantic (Canning-Clode and Carlton 2017), a "mirror" phenomenon of “Caribbean Creep” (Canning-Clode et al. 2011), however, the former study also highlighted that atypical cooling periods may provide a “reset mechanism” that will result in temporary setbacks or rate reductions in range expansions of non-indigenous species.
Hybridization in species that use external fertilization is common and marine hybrid zones are often referred to as extensive and heterogenous due to their large population size, high fecundity and larval dispersal (Gardner 1997). Mytilus spp. populations show extensive hybridisation worldwide (Gosling 1992; Gardner 1994; Saavedra et al. 1996; Toro et al. 2002; Riginos and Cunningham 2005). The European Mytilus hybrid zone is spatially complex containing a mixture of pure, hybrid and introgressed individuals spanning over a 1400 km of coastline from France to the north of Scotland but is absent from the Irish Sea (Gosling et al. 1992; Gosling et al. 2008). Evidence of hybridisation and hybrid zones of M. edulis and M. galloprovincialis in the south west of England and Ireland were first recorded in the 1970s and subsequent studies have further documented this phenomenon (Gosling and Wilkins 1977; Skibinski et al. 1983; Gosling et al. 2008). Gosling et al. (2008) examined the population genetic structure of mussels at twenty sites from 2003 to 2006 on the Irish coast to determine the current range of M. edulis and M. galloprovincialis. In that study, a higher frequency of M. galloprovincialis and hybrids were observed on the Irish Atlantic coast compared to M. edulis.
The objectives of this current study were to determine mytilid species composition and abundance at a number of sites around the coast of Ireland (2010–2017) and at several Welsh sites (2011). In this study six Irish sites were revisited more than once and a comparison with the Gosling et al. (2008) was carried out using molecular markers at four sites to determine if any variation has occurred in distribution since that study was carried out, possibly due to changing weather patterns, habitat suitability, mussel fitness, recruitment success and or climate change.
Materials and methods
-
(1)
Mussels sampled in current study
In this study, 1549 mussels were screened from 27 sites around Ireland and Wales with single sample sizes varying from 19 to 106 individuals. Screening took place from 2010 to 2017 with sampling occurring once at most sites, apart from six Irish sites that were sampled more than once (Table 1).
Table 1 Mussel sample sites with description of sample month, year and size at each site -
(2)
Comparison with previous distribution study
The distribution and abundance of M. edulis, M. galloprovincialis and hybrids observed in this study were compared with a previous study by Gosling et al. (2008), which was carried out with the sampling of 780 mussels at twenty sites from all Irish coasts from May 2003 to February 2006. Four sites were common to both studies: Carlingford Lough (Site #19), Carnsore Point (Site #1), Garrettstown (Site #13) and Lough Hyne (Site #14) (Table 1) and for additional comparison between this study and the Gosling et al. (2008) study, all sites were categorised by coast (north, west, south and east) with overall abundance of each Mytilus species and hybrids being compared.
-
(3)
Molecular analysis to identify mussel species
Gill tissue (5 mm2) was excised from each mussel (n = 1549) and was stored in 96% ethanol if DNA extraction was not conducted on the same day of sampling. DNA was extracted using the chelex-100 extraction method (Walsh et al. 1991).
PCR was carried out to detect the nuclear DNA markers Me15/Me16 (Inoue et al. 1995). The PCR mastermix was modified slightly to include 5 × green buffer. Amplification was conducted in 25 µl of the reaction mixture containing 14 µl ddH2O, 5 µl 5 × green buffer (Promega), 2.5 µl of each of the four deoxyribonucleotide triphosphates (dATP, dCTP, dGTP, dTTP), 1.5 µl MgCl2, 0.5 µl of the primer Me15 (5′-CCAGTATACAAACCTGTGAAGA-3′), 0.5 µl of the primer Me16 (5′-TGTTGTCTTAATGGTTTGTAAGA-3′), 1 µl of Taq polymerases and 1 µl of total DNA. The following conditions were used for the PCR in a thermocycler: 94 °C for 30 s, 55 °C for 45 s and 70 °C for 90 s (40 cycles). Electrophoresis of the amplification products was conducted in a 2% agarose gel. The expected product size for M. galloprovincialis is 126 bp while in M. edulis it is 180 bp. In hybrids, both bands occur simultaneously at 126 bp and 180 bp.
-
(4)
Meteorological data
Irish sea surface temperature data and graphs were sourced from Casal and Lavender (2017) while surface temperature data was sourced from the national meteorological agency Met Eireann (https://www.met.ie/climate/what-we-measure/temperature).
A long time series of annual rainfall from twenty-five stations in Ireland was sourced from Met Eireann (https://www.met.ie/climate/what-we-measure/rainfall) and Noone et al. (2015).
-
(5)
Statistical analysis
Chi-square (χ2) analysis was carried out using Excel to calculate significant difference in the abundance of each mussel species between (a) sites with varying water quality and (c) sites with varying exposures, with significance determined at p < 0.05. Chi-square (χ2) analysis was also carried between this current study and the Gosling et al. (2008) study, comparing the abundance of each mussel species and their overall abundance on each coast.
Results
-
(1)
Mytilus edulis, Mytilus galloprovincialis and hybrids of both parent species distribution and abundance at Irish sample sites
In total, 1413 mussels (96.9%) were identified to species by PCR from the Irish study sites. Of this, 898 (63.6%) individuals were M. edulis, 235 (16.6%) M. galloprovincialis and 280 (19.8%) hybrids were detected along the coast (Table 2 and Fig. 1). Irish sites (#19, #20 and #21) in the Irish Sea were the only sites that were found to have 100% M. edulis while all of the Welsh mussels (n = 90) were identified as M. edulis in the PCR.
Table 2 PCR analysis of mussel samples (No. of individuals/Sample total) from twenty-four Irish and three Welsh sites to determine relative abundance of Mytilus edulis, Mytilus galloprovincialis and hybrids spatially and temporally Fig. 1 
Map of Ireland and Wales showing the wild Mytilus edulis (black), Mytilus galloprovincialis (white) and hybrid (grey) abundance and distribution at study sites (study site number in parentheses)
Excluding the Irish Sea samples on the east coast, M. edulis was the dominant species in eighteen samples on the south, west and north coasts, M. galloprovincialis was the dominant species in two samples (Sites #5 and #15) on the south and west coast respectively and hybrids dominated nine samples at sites on the south coast (Table 2). Of the five Irish sites that were sampled more than once and with a time difference of two (Site #8), four (Site #14) and seven (Site #12) years between sampling time points, the status of the dominant species remained the same except for one of the locations Roches Point within Cork Harbour (Site #12), where there was a decrease in M. edulis and an increase in hybrids in November 2017. Hybrids were consistently the dominant species at the marine nature reserve at Lough Hyne (Site #14).
-
(2)
Comparison of mussel species distribution and abundance in this study with historical study
Comparing the results of this study and the Gosling et al. (2008) study by coast abundance and distribution for each of the Mytilus spp. and hybrids, a significant difference in M. edulis abundance (p < 0.05) was observed (Figs. 2 and 3). M. edulis was reported to be the dominant species on all coasts in this study, while M. galloprovincialis and hybrids were the most abundant in the Gosling et al. (2008) study. A similar abundance in hybrids on the south coast was observed in both studies while no significant difference in M. galloprovincilis abundance in both studies was recorded on the west coast.
Fig. 2 
Comparison of Mytilus spp. and hybrid distribution and abundance on each Irish coast in the Gosling et al. (2008) study (block colour fill sites) and in this study (pattern fill)
Fig. 3 
Comparison of mussel species abundance at Irish sites sampled in this study and in the Gosling et al. (2008) study* including sites that were revisited more than once in this study
Over the time points of the two Irish studies there has been an increase in M. edulis abundance at certain sites (Dungarvan Site #8, Garrettstown* Site #13 and Carnsore Point* Site #1), while its abundance has remained stable at other sites such as Ringaskiddy (Site #12) and Carlingford Lough* (Site #19). A decrease in M. edulis abundance has been observed in more recent years at Roches Point (Site #12 from 2015 to 2017) and at Lough Hyne* (from 2013 to 2017) (Fig. 3). M. galloprovincialis abundance has been showing a decreasing trend at Dungarvan, Ringaskiddy, Garrettstown*, (significantly so p < 0.05) and at Carnsore Point* (significantly so ( p < 0.05) while its abundance has started to increase at Lough Hyne*. Hybrid abundance has significantly decreased at Carnsore Point* along with a decrease at Dungarvan and Garrettstown, however, the latter site has shown a slight increase from 2013 to 2017 (+ 8%). Hybrid abundance is consistently increasing over the study time points at Lough Hyne and has significantly increased at Roches Point from 2015 to 2017 (+ 30%).
-
(3)
Meterological data
Irish waters present a warming trend of 0.3 °C decade−1 with warming being more intense in autumn (Fig. 4, Casal and Lavender 2017). A significant latitudinal gradient is present in Irish waters with waters to the south being a lot warmer than waters to the north and a seasonal effect is also present with coldest SST values in March and highest SST values in August.
Fig. 4 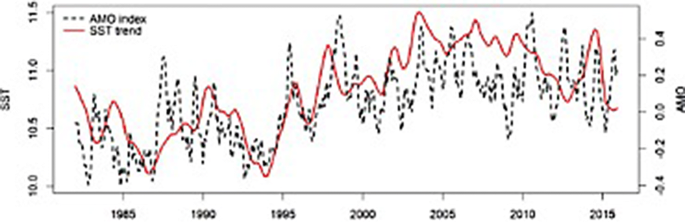
Sea surface temperature (°C) (SST) in Irish waters showing SST trend and Atlantic Multi-decadal Oscillation (AMO) index sourced and modified from Casal and Lavender (2017)




Mean air temperatures have varied considerably from year to year, however, warming periods have occurred from the late 1980s to the present time (https://www.met.ie/climate/what-we-measure/temperature). There has been an increase in the number of days with temperatures over 20 °C, and a decrease in the number of days with temperatures below 0 °C, however, extreme cold winters did occur in 2010 and in 2011 with air temperatures of -2.5 °C from the long term average (LTA).
Rainfall shows great year to year variability, however, from the 1970s (M. galloprovinicalis first recorded to be present in Ireland) to 2005, when the Gosling et al. (2008) study was primarily conducted, maximum national annual rainfall on average did not exceed 1290 mm of rainfall (Fig. 5A). Since 2005 national annual rainfall in Ireland has been increasing with up to 1400 mm of rainfall being recorded. Winter 2015/16 was the wettest winter on record, followed by winter 2013/14, and are the only winters where rainfall totals in excess of 500 mm were recorded (Noone et al. 2015) (Fig. 5B). A 30 year running mean of the national annual rainfall has indicated that an increase in average national rainfall of approximately 70 mm over the last two decades has occurred with all seasons showing a small increase in totals over the last few decades (Noon et al. 2015).

modified from Noone et al. (2015)
A Mean national annual rainfall for Ireland (25 stations) from the 1960s to 2015 with B winter rainfall highlighted. Both graphs sourced and
Discussion
Mytilus edulis, M. galloprovincialis and hybrid forms of both parental species were detected around the coast of Ireland, while Mytilus trossolus or hybrids of this species were not detected. M. edulis was the dominant species found at most Irish sites with a lower abundance of M. galloprovincialis and hybrids. These findings are in stark contrast to the study of Gosling et al. (2008) carried out seven years prior to this study. In the intervening time period (fourteen years) the relative abundance of the parental forms of both mussel species and hybrids around the coast of Ireland has changed with M. edulis, the native species, being once again the most frequently observed and abundant species. The slightly higher abundance of hybrids over M. galloprovincialis may indicate that these are more suited to survival under Irish conditions and a changing Irish climate than the Mediterranean parent species or it is possible that they are first generation transient hybrids. In the Irish Sea area, M. edulis continues to be the only species present between Carlingford Lough and Poulshone Strand along the Irish coast and Gallows Point and Swansea along the Welsh coast. It has been suggested that a thermal front develops during late spring at the boundary between the Celtic Sea and the southern Irish Sea (Brown et al. 2003), which prevents the movement of M. galloprovincialis into the Irish Sea region.
Mosaicism or spatial genetic patchiness in distribution is a common feature of Mytilus hybrid zones (Comesaña and Sanjuan 1997; Rawson and Hilbish 1998) and was observed at most Irish sites in this study. In the Gosling et al. (2008) study, this phenomenon was not observed, however, hybrids and M. galloprovincialis only were present at a single site on the west coast of Ireland, with the hybrids outnumbering the parent species possibly indicating hybrid vigour. Hybrids may have lower fitness due to genetic discontinuities (Mayr 1963; Barton and Hewitt 1985) or a lack of “hybrid” habitat (Howard 1986) and mytilid hybrids can be more susceptible to disease (Fuentes et al. 2002), however, certain studies have shown “hybrid vigour” (heterosis) in the first hybrid generation (F1 hybrid) which may allow hybrids to function over a wider range of environmental conditions than the two parental species (Moore 1977; Arnold 1997). Hybrids were consistently the dominant group at the marine reserve Lough Hyne and this may possibly be due to a number of reasons. One is that fertile hybrids may be reproducing earlier than Mytilus spp. and their larvae are settling earlier within the lough. This phenomenon of Mytilus hybrids being fertile and reproducing at a faster rate has been observed (Dias et al. 2009). Another reason could be that hybrids grow at a faster rate than either of their parent species, which would mean that there are outcompeting the Mytilus spp. for food and space. Additionally, M. galloprovincialis are showing signs of recovery in the lough after decreasing in abundance from 58% in 2005 to 14% in 2013 with an increase to 20% in 2017.
Irish air temperature recorded by Met Eireann (https://www.met.ie/climate/what-we-measure/temperature) during this study and the Gosling et al. (2008) study indicates that two prolonged cold winters occurred in 2010 and in 2011. Although surface seawater temperatures did not deviate dramatically from average temperatures during and shortly after this time period, air temperature, snowfall and ground frosts did. Severe winters have been associated with the complete or almost complete extinction of intertidal populations (Blegvad 1929; Smidt 1944; Kristensen 1957; Beukema 1985), however, M. edulis can withstand extreme cold and freezing, surviving when its tissue temperature drops to −10 to −30 °C (Williams 1970; Seed and Suchanek 1992; Loomis 1995). M. edulis was relatively unaffected by the severe winter of 1962/63 on the south-east coast of England, with 30% mortality reported and 2% in south Wales (Crisp 1964). Crisp (1964) noted that most mortality was caused by predation on individuals weakened or moribund due to the low temperatures rather than the temperature itself. It is unclear what the minimum temperature tolerance limit is in M. galloprovincialis but this mussel species is known to tolerate 7 °C, during upwelling events on the South Africa coastline (Erkom and Van Griffiths 1992). Jansen et al. (2007) noted that M. galloprovincialis populations were more sensitive to cold shock than M. edulis populations.
Northward seawater currents make contact with Ireland initially on the south coast before being transported in coastal currents that move around the Irish coast in a clockwise direction (Steele et al. 2013). This would support the higher abundance of M. galloprovinicalis observed on the south and southwest coasts in this study, as the warmer seawater temperatures experienced in southern Irish waters (Casal and Lavender 2017) would allow a more stable population to remain and possibly self-recruit. Previous studies of mytilid spp. population distribution have focussed on the limiting effects of water temperature on dispersal (Barry et al. 1995; Suchanek et al. 1997; Sagarin et al. 1999). M. galloprovincialis prefers warmer waters (Sarver and Foltz 1993; Suchanek et al. 1997) and a range expansion of M. galloprovincialis is thought to have occurred from the 1930′s to the early 1990′s due to increased water temperatures (Geller 1999; Jones et al. 2001; Schneider 2008). However, air temperature and prolonged frosts associated with more frequently occurring “cold snaps” during winter months may be have a limiting effect on populations sustaining themselves and outcompeting the native mussel populations.
Of interest, national annual rainfall in Ireland has been increasing with up to 1400 mm of rainfall being recorded since 2005 and with winters 2013/14 and 2015/16 being the wettest winters on record (Noone et al. 2015). Heavy precipitation events and subsequent increased freshwater loading in bays and estuaries may result in sudden fluctuating and reduced salinity events, which would have an impact on M. galloprovincialis. An earlier Irish study carried out by Seed (1974) stated that when salinity was not reduced the invasive M. galloprovincialis was the dominant mussel, out competing the native species. M. galloprovincialis made up 40% of the population in those locations (Seed 1974). Shurova (2001) observed that the lowering of salinity resulted in an increase in mortality rate and a decrease in numbers, biomass, juvenile recruitment, growth rates and annual production of M. galloprovincialis in the Black Sea.
The results of this study would indicate that although M. galloprovincialis and hybrids of both parent species are present on the south, west and north coasts of Ireland, the native M. edulis is dominant at most sites. Although a warming trend has been observed in sea surface temperatures over several decades, several anomalous cold winters compounded by an increase in rainfall and subsequent run off of freshwater loadings into coastal locations may be having an effect on nearshore habitat suitability for M. galloprovincialis. Future predictions of colder winters over Northern Europe, due to climate change and cold polar air streaming down in to Europe (Tang et al. 2013), may be limiting for this Mediterranean species as well as predicted increased precipitation (Noone et al. 2015). Hughes et al. (2015) observed that the eastern limit of distribution of M. galloprovincialis in South Africa was also set by unfavourable abiotic conditions. Further sampling of mussels at the Irish sites in the future, nearshore and subtidally, would be of interest to determine if the Mytilus spp. population structure is remaining stable over time or if localised factors and meteorological influences impact its composition.
References
Arnold ML (1997) Natural hybridization and evolution. Oxford University Press, Oxford
Barry JP, Baxter CH, Sagarin RD, Gilman SE (1995) Climate-related, long-term faunal changes in a California rocky intertidal community. Science 267:672–675
Barton NH (1979) The dynamics of hybrid zones. Heredity 43:341–359
Barton NH, Hewitt GM (1985) Analysis of hybrid zones. Ann Rev Ecol Syst 16:113–148
Barton NH, Hewitt GM (1989) Adaptation, speciation and hybrid zones. Nature 431:497–503
Bayne BL (1964) Primary and secondary settlement in Mytilus edulis L. (Mollusca). J Animal Ecol 33:513–523
Beukema JJ (1985) Zoobenthos survival during severe winters on high and low tidal flats in the Dutch Wadden Sea. In: Gray JS, Christiansen ME (eds) Marine biology of polar regions and effects of stress on marine organisms. Wiley, Chichester, pp 351–361
Blegvad H (1929) Mortality among animals of the littoral region in ice winters. Rep Dan Biol Stn 35:49–62
Braby CE, Somero GN (2006) Following the heart: temperature and salinity effects on heart rate in native and invasive species of blue mussels (genus Mytilus). J Exp Biol 209:2554–2566
Branch GM, Steffani CN (2004) Can we predict the effects of alien species? A case-history of the invasion of South Africa by Mytilus galloprovincialis (Lamarck). J Exp Mar Biol Ecol 300:189–215
Brown J, Carrillo L, Fernand L, Horsburgh KJ, Hill AE, Young EF, Medler KJ (2003) Observations of the physical structure and seasonal jet-like circulation of the Celtic Sea and St. George's Channel of the Irish Sea. Cont Shelf Res 23:33–561
Canning-Clode J, Fowler AE, Byers JE, Carlton JT, Ruiz GM (2011) 'Caribbean Creep' chills out: climate change and marine invasive species. PLoS ONE 6(12):e29657
Canning-Clode J, Carlton JT (2017) Refining and expanding global climate change scenarios in the sea: poleward creep complexities, range termini, and setbacks and surges. Divers Distrib 23:463–473
Casal G, Lavendar SJ (2017) Spatio-temporal variability of sea surface temperature in Irish waters (1982–2015) using AVHRR sensor. J Sea Res 129:89–104
Christiansen BO (1965) Notes of the littoral fauna of Bear Island. Astarte 26:1–15
Comensañsa AS, Sanjuan A (1997) Microgeographic allozyme differentiation in the hybrid zone of Mytilus galloprovincialis and M. edulis on the continental European coast. Helg Meeresunters 51:107–124
Crisp DJ (1964) The effects of the severe winter of 1962–63 on marine life in Britain. J Anim Ecol 33:165–210
Dailianis S (2010) Environmental impact of anthropogenic activities: the use of mussels as a reliable tool for monitoring marine pollution. In: McGevin LE (ed) Mussels: anatomy, habitat and environmental impact. Nova Science Publishers Inc, New York, pp 43–72
Dias PJ, Dordor A, Tulett D, Piertney S, Davies IM, Snow M (2009) Survey of mussel (Mytilus) species at Scottish shellfish farms. Aqua Res 40:1715–1722
FAO (2017) Cultured aquatic species information programme: Mytilus edulis. www.fao.org/fishery/culturedspecies/Mytilus_edulis/en Accessed 16 Mar 2020
Fuentes J, López JL, Mosquera E, Vázquez J, Villalba A, Alvarez G (2002) Growth, mortality, pathological conditions and protein expression of Mytilus edulis and M. galloprovincialis crosses cultured in the Ría de Arousa (NW of Spain). Aquaculture 213:233–251
Gardner JPA (1994) The structure and dynamics of naturally occurring hybrid Mytilus edulis Linnaeus, 1758 and Mytilus galloprovincialis Lamarck, 1819 (Bivalvia: Mollusca) populations: review and interpretation. Arch Hydrobiol Suppl 99:37–71
Gardner JPA (1997) Hybridization in the sea. Adv Mar Biol 31:1–65
Geller JB (1999) Decline of a native mussel masked by sibling species invasion. Conserv Biol 13:661–664
GISD (Global Invasive Species Database). 2017. Species profile: Mytilus galloprovincialis. Downloaded from https://www.iucngisd.org/gisd/species.php?sc=102. Accessed 16 Mar 2020
Gosling EM, Wilkins NP (1981) Ecological genetics of the mussels Mytilus edulis and M. galloprovincialis on Irish Coasts. Mar Ecol Prog Ser 4:221–227
Gosling EM, Wilkins NP (1977) Phosphoglucoisomerase allele frequency data in Mytilus edulis from Irish coastal sites: its ecological significance. In: Keegan BF, O’Ceidigh P, Boaden PJS (eds) Biology of benthic organisms. Pergamon Press, London, pp 297–309
Gosling EM (1992) Systematics and geographic distribution of Mytilus. In: Gosling E (ed) The mussel Mytilus: ecology, physiology, genetics and culture. Elsevier, Amsterdam, pp 1–20
Gosling E (2004) Bivalve molluscs: biology, ecology and culture. Blackwell Publishing, New Jersey
Gosling E, Doherty S, Howley N (2008) Genetic characterization of hybrid mussel (Mytilus) populations on Irish coasts. J Mar Biol Assoc UK 88:341–346
Hepper BT (1957) Notes on Mytilus galloprovincialis Lamarck in Great Britain. J Mar Biol Assoc UK 36:33–40
Howard DJ (1986) A zone of overlap and hybridisation between two ground cricket species. Evolution 40:34–43
Hughes RN, Hughes DJ, Smith IP, Dale AC (2015) Oceanography and marine biology: an annual review. CRC Press, Florida
Inoue K, Waite HJ, Matsuoka M, Odo S, Harayama S (1995) Interspecific variations in adhesive proteion sequences of Mytilus edulis, M. galloprovincialis, and M. trossulus. Biol Bull 189:370–375
Jansen JM, Bonga SW, Hummel H (2007) Differential coldshock resistance among acclimated European mussel populations. Mar Freshw Behav Physiol 40:233–245
Jones PD, Osborn TJ, Briffa KR (2001) The evolution of climate over the last millennium. Science 292:662–667
Kristensen I (1957) Differences in density and growth in a cockle population in the Dutch Wadden Sea. Arch Neerl Zool 12:351–453
Lenz M, Ahmed Y et al (2018) Heat challenges can enhance population tolerance to thermal stress in mussels: a potential mechanism by which ship transport can increase species invasiveness. Biol Invasions 20(11):3107–3122
Lewis J, Seed R (1969) Morphological variations in Mytilus from southwest England in relation to the occurrence of M. galloprovincialis. Cah Bio Mar 10:231–253
Lima FP, Ribeiro PA, Queiroz N, Hawkins SJ, Santos AM (2007) Do distributional shifts of northern and southern species of algae match the warming pattern? Global Change Biol 13:2592–2604
Loomis SH (1995) Freezing tolerance of marine invertebrates. Oceanogr Mar Biol Annu Rev 33:337–350
Mayr E (1963) Animal species and evolution cambridge. Harvard University Press, Cambridge
Moore WS (1977) An evaluation of narrow hybrid zones in vertebrates. Q Rev Biol 52:263–277
Noone S, Murphy C, Coll J, Matthews T, Mullan M, Wilby RL, Walsh S (2015) Homogenisation and analysis of an expanded long-term monthly rainfall network for the Island of Ireland (1850–2010). Int J Climatol 36:2837–2853
Ozer A, Guneydag S (2014) First report of some parasites from Mediterranean mussel, Mytilus galloprovincialis Lamarck, 1819, collected from the Black Sea coast at Sinop. Turk J Zool 38:486–490
Page HM, Hubbard DM (1987) Temporal and spatial patterns of growth in mussels Mytilus edulis on an offshore platform: relationships to water temperature and food availability. J Exp Mar Biol Ecol 111:159–179
Perry AL, Low PJ, Ellis JR, Reynolds JD (2005) Climate change and distribution shifts in marine fishes. Science 308:1912–1915
Rawson PD, Hilbish TJ (1998) Asymmetric introgression of mitochondrial DNA among European populations of blue mussels (Mytilus spp.). Evolution 52:100–108
Riginos C, Cunningham CW (2005) Local adaptation and species segregation in two mussel (Mytilus edulis x Mytilus trossulus) hybrid zones. Mol Ecol 14:381–400
Saavedra C, Donald TS, Stanwood RR, Zouros E (1996) Species-specific segregation of gender-associated mitochondrial DNA types in an area where two mussel species (Mytilus edulis and M. trossulus) hybridize. Genetics 143:1359–1367
Sanjuan A, Zapata C, Alvarez G (1994) Mytilus galloprovincialis and M. edulis on the coasts of the Iberian Peninsula. Mar Ecol Prog Ser 113:131–146
Sagarin RD, Barry JP, Gilman SE, Baxter CH (1999) Climate related change in an intertidal community over short and long time scales. Ecol Monogr 69:465–490
Sarver SK, Foltz DW (1993) Genetic population-structure of a species complex of blue mussels (Mytilus spp.). Mar Biol 117:105–112
Schneider KR (2008) Heat stress in the intertidal: comparing survival and growth of an invasive and native mussel under a variety of thermal conditions. Biol Bull 215:253–264
Seed R (1969) The ecology of Mytilus edulis L. (Lamellibranchiata) on exposed rocky shores I breeding and settlement. Oecologia 3:277–316
Seed R (1974) Morphological variations in Mytilus from the Irish Coasts in relation to the occurrence and distribution of M. galloprovincialis Lmk. Cah Biol Mar 15:1–25
Seed R, Suchanek TH (1992) Population and community ecology of Mytilus. In: Gosling EM (ed) The mussel Mytilus: ecology, physiology, genetics and culture. Elsevier, Amsterdam, pp 87–169
Shurova N (2001) Influence of salinity on the structure and the state of bivalve Mytilus galloprovincialis populations. Russ J Mar Biol 27:151–155
Skibinski DOF, Beardmore JA, Cross TF (1983) Aspects of the population genetics of Mytilus (Mytilidae; Mollusca) in the British Isles. Biol J Linn Soc 19:137–183
Smidt ELB (1944) The effects of ice winters on marine littoral faunas. Folia Geogr Dan 2:1–36
Steele CJ, Dorling SR, von Glasow R, Bacon J (2013) Idealized WRF model sensitivity simulations of sea breeze types and their effects on offshore windfields. Atmos Chem Phys 13:443–461
Suchanek TH, Geller J, Kreiser B, Mitton J (1997) Zoogeographic distributions of the sibling species Mytilus galloprovincialis and M. trossulus (Bivalvia: Mytilidae) and their hybrids in the north Pacific. Biol Bull 193:187–194
Tang Q, Zhang X, Yang X, Francis JA (2013) Cold winter extremes in northern continents linked to Arctic sea ice loss. Environ Res Lett 8:014036
Toro JE, Thompson RJ, Innes DJ (2002) Reproductive isolation and reproductive output in two sympatric mussel species (Mytilus edulis, M. trossulus) and their hybrids from Newfoundland. Mar Biol 141:897–909
Tyler-Walters H, Seed R (2006) "The Marine Life Information Network" (On-line). at https://www.marlin.ac.uk/species/detail/1421 Accessed 16 Mar 2020
Van Erkom SC, Griffiths CL (1992) Physiological energetics of four South African mussel species in relation to size, ration and temperature. Comp Biochem Physiol 101:779–789
Walsh PS, Metzger DA, Higuchi R (1991) Chelex 100 as a medium for simple extraction of DNA for PCR-based typing from forensic material. Biotechniques 10:506–513
Williams RJ (1970) Freezing tolerance in Mytilus edulis. Comp Biochem Physiol 35:145–161
Zagata C, Young C, Sountis J, Kuehl M (2008) ``Mytilus edulis`` (Online), Animal diversity web. https://animaldiversity.org/accounts/Mytilus_edulis/ Accessed 16 Mar 2020
Acknowledgements
This study was part-funded by the SUSFISH (Grant Agreement No. 244132) and BlueFish (Grant Agreement No. 80991) Projects under the European Regional Development Fund (ERDF) through the EU Ireland/Wales Interregional (INTERREG) Programme. A cross-border programme investing in the overall economic, environmental and social well-being of Ireland and Wales. The authors would like to thank the reviewers for their positive response to this study and for providing interesting comments, thoughts and very relevant references.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Lynch, S.A., Coghlan, A., Leary, B.O. et al. Northward establishment of the mediterranean mussel Mytilus galloprovincialis limited by changing climate. Biol Invasions 22, 2725–2736 (2020). https://doi.org/10.1007/s10530-020-02294-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-020-02294-6