
Abstract
Despite advances in understanding the ecology and consequences of pine invasions, information on the patterns of structure, dynamics, and growth needed to manage these invasive populations are still poorly known. Here we used tree ring analysis to elucidate growth dynamics, age distribution and fire effects, by sampling three populations of slash pine invading Cerrado vegetation in southeastern Brazil. We found that the invasion occurred in waves, every 5–7 years. These pulses are likely related to the time needed by founder trees to reach maturity, and to climatic events. We found distinct growth patterns among the three study areas. Individual trees in the open sites, under low competition for light, showed an increasing trend of diameter and of the basal area growth rates with age. In the dense stands, however, the diameter increment rates gradually decreased with age, as the competition for light was intensified, and the basal area growth rate increased at lower rates. Growth rates were markedly reduced after fire, likely due to fire impact on the canopies, reducing photosynthesis. Fire scars in the wood allowed us to confirm that the formation of the tree rings occurs annually. When competition for light intensifies as the canopy of the stand closes, the invasive populations experience reduced individual growth of adult trees and constraints to the recruitment of young individuals. We did not find, however, evidence of self-thinning due to competition that could reduce the population of adult trees, open gaps and allow colonization by shade-tolerant native species. Therefore, a monodominant pine stand—the “pine desert” feature of the invaded sites—tends to be persistent over time.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Pinus elliottii Engelm. (slash pine) is a native tree from North America that became one of the most aggressive invaders of savannas and grasslands in the southern hemisphere, after being introduced for timber production (Richardson et al. 1994; Richardson and Higgins 1998; Richardson 2006). Species of the genus Pinus L. have significant economic and environmental importance (Nuñez et al. 2017). In Brazil, tax breaks were given to forestry of Pinus species after the 1960s, and they became a significant part of forest plantations, reaching 1.8 million hectares in 2008. Today, Pinus species are used in several segments of forestry and wood sectors, mainly for pulp, paper, charcoal, panels and lumber production (Vasquez et al. 2007; Sociedade Brasileira de Silvicultura 2008). P. elliottii has been reported as invasive in Argentina, Australia (NSW), Brazil, Hawaii, and South Africa (Richardson 2006), promoting structural, floristic and functional changes in plant communities, reducing the biodiversity, changing hydrological regimes, and altering soil nutrients (Vasquez et al. 2007; Simberloff et al. 2009; Abreu and Durigan 2011; Abreu 2013; Fischer et al. 2014). The native range of the species are the floodplains from the Southeast USA, were slash pine experiences a natural fire regime of about 25 fires per century (Carey 1992). The species grows faster in Brazil than in the native habitat, forming invasive stands with twice the density and a litter depth 14 times thicker than in native pine forests of Mississippi (Brewer et al. 2018). Pinus elliottii is able to survive light fires due to the thick bark and bark structures that dissipate the heat. Small individuals can die due to fire, but after 10–12 years slash pine trees are high enough to resist to fire that does not crown (Carey 1992). From its introduction to Brazil in the 1940s, P. elliottii has become one of the most planted pine species in Brazil, mostly in temperate climates (Kronka et al. 2005; Vasquez et al. 2007).
Slash pine has been recognized in the last two decades as highly invasive in the Brazilian grasslands and savannas of the Cerrado (Zanchetta and Diniz 2006; Vasquez et al. 2007; Abreu and Durigan 2011; Abreu 2013; Bechara et al. 2013). The Brazilian savanna (Cerrado) is a biome which covers around 2 million km2 and has 12,000 plant species, 4400 of which are endemic (Oliveira-Filho and Ratter 2002), and it has been the region most heavily invaded by P. elliottii (Zanchetta and Diniz 2006; Abreu and Durigan 2011; Abreu 2013). The invasion process in southeastern Brazil is fast; 22 years after the first pine trees arrived at one site, a grassland savanna was converted into a dense Pinus forest with 12,455 individuals and 26.44 m2 of basal area per hectare. The species richness of the understory was reduced from 52 to 16 species, and 90% of native individuals > 50 cm tall was lost, along with the entirety of the herbaceous layer. Pinus invasion thus produces profound changes in the vegetation structure, causing significant losses of plant diversity (Abreu and Durigan 2011).
Despite the recent advances in the geography and ecology of pine invasion, aspects such as the structure, dynamics and growth patterns of the invasive populations are still poorly known. Simberloff (2014) argues that the problem of biological invasions could be better addressed by moving from a focus on invasive species to addressing invasive populations, because reducing the population size of an invader could be a desired goal for invasive species management. The potential for dendrochronology to provide the needed information and to inform management has been demonstrated by several studies in tropical regions, some encompassing Pinus species (Biondi 1999; Worbes 1999; Worbes et al. 2003; Couralet et al. 2005; Brienen and Zuidema 2006; Schöngart 2008; Zimmer and Baker 2009; Rozendaal and Zuidema 2011; Locosssselli et al. 2017; Tomazello-Filho et al. 2017; Vlam et al. 2017). Pinus species have distinct tree-rings which can be analyzed by dendrochronology. Schulman (1944) developed the first tree-ring chronologies with Pinus species in the tropical regions of Mexico. Since then, dendrochronological studies with Pinus were developed in many tropical countries, including Honduras, Guatemala, El Salvador, Dominican Republic, Laos, Thailand, Vietnam and Brazil (Johnson 1980; Sarutanon et al. 1995; D’Arrigo et al. 1997; Biondi 2001; Speer et al. 2004; Buckley et al. 2007; Moya and Filho 2009; Zimmer and Baker 2009; Sigal 2011; Zuidema et al. 2011; Venegas-González et al. 2015). These studies provided evidence of annual growth rings and proposed tree-ring chronologies and inferences about growth-climate relationships. Specifically for P. elliottii, tree-ring chronologies and dendroclimatological studies were developed in temperate and subtropical climates (Harley et al. 2011). In Brazil, tree-ring analysis in Pinus has been used to provide information about age and to quantify wood growth, including for forestry production and height growth evaluation, pruning and thinning effects, soil nutrition effects and wood density variations (Trovati and Ferraz 1984; Brito et al. 1986; Pereira and Ahrens 2003; Ferreira and Filho 2009; Coelho and Hosokawa 2010; Elesbão and Schneider 2011; Schneider et al. 2013; Ortega Rodriguez et al. 2018). Dendrochronological analyses have also been used to evaluate height growth, to differentiate juvenile and adult wood, to assess thinning effects on wood quality and to explain wood density variations (Palermo et al. 2003, 2013; Pereira and Tomaselli 2004; Schneider et al. 2013). Three studies developed in Brazil so far have used tree-ring analysis to investigate the invasion process and to evaluate the effects on community level of Pinus invasion (Abreu and Durigan 2011; Fischer et al. 2014; Tomazello-Filho et al. 2017).
As in other grassy ecosystems, natural and anthropogenic fires are common in the Cerrado (Klink and Machado 2005; Miranda et al. 2009), including in areas invaded by P. elliottii. Fire is an essential factor for maintaining natural communities in the Cerrado (Miranda et al. 2009) and has been used in experiments to manage invasive species (Abreu 2013). Fire events can be recorded by fire scars marked in the wood and dated by the tree ring analysis (Brandes et al. 2018), and the formation of fire scars has been verified in Pinus species (Speer et al. 2004; Harley et al. 2011). Fire-scar dating of P. elliottii could provide information about fire events and the effects of fire on the structure of the invasive populations.
In this study, we used tree ring analysis to elucidate growth dynamics, wood production, and age distribution in three populations of slash pine invading Cerrado vegetation in southeastern Brazil. Among the questions we address are: (1) Can the fire scars demonstrate the annual nature of the growth rings? (2) Does the pine population age structure and growth dynamics differ among open and closed invaded habitats? We searched for distinct patterns in population structure and growth dynamics depending on the type of habitat invaded, namely open grasslands and riparian forests.
Materials and methods
Study areas
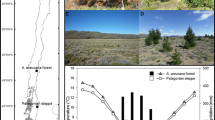
The study was carried out in two reserves located in São Paulo state, Southeast Brazil, within the Cerrado biome: Itirapina Ecological Station (IEcS) and Santa Barbara Ecological Station (SBEcS) (Fig. 1). Both reserves protect typical savanna vegetation (cerrado sensu lato), ranging from grasslands (campo cerrado, campo sujo, campo limpo) to woodlands (cerradão). The climate is classified as Cwa following Köppen’s system (humid subtropical with dry winter and hot summer). Biological invasion is amongst the major challenges for management and conservation of the reserves. Pinus species planted for forestry purposes in the 1960s in the vicinity promote constant propagule pressure over the Cerrado remnants, with large areas already invaded (Silva et al. 2006; Melo and Durigan 2009; Miashike 2015).

Map of South America and detail of São Paulo State, Brazil. Gray = Cerrado biome; circle = Santa Barbara Ecological Station (SBEcS—Area 3); triangle = Itirapina Ecological Station (IEcS—Areas 1 and 2)
The IEcS (22°11′S–22°15′S, 47°51′W–48°00′W—Fig. 1) covers 2300 ha, with altitude ranging from 710 to 830 m a.s.l. (Silva et al. 2006). Mean annual temperature is 21.9 °C, the hottest months were January (24.9 °C) and February (24.8 °C), and the coldest months were June (17.8 °C) and July (17.9 °C). The average annual precipitation is 1459 mm, with means ranging from 1128 mm (77%) in the rainy season (Oct–Mar) and 331 mm (23%) in the dry season (Apr–Sep). The monthly average precipitation in the driest months is 24 mm in July and 42 mm in August (Silva et al. 2006). At Itirapina we sampled two distinct sites with invaded patches of similar size, around 0.03 ha. Area 1 (or IEcS1) (22°11′52.15″S, 47°53′19.94″W) was in a grassland savanna recently invaded, in well drained soils despite being close to a water dam. Pine trees were at low density, with sunlight able to penetrate the canopies and reach the ground. Area 2 (or IEcS2) (22°14′42.24″S, 47°51′28.15″W) was in the riparian zone of a stream, likely invaded for longer than Area 1. Previous vegetation was a low-biomass riparian forest, characterized by a shaded environment with high competition for light in the understory, with no sunlight reaching the ground. Temporal series of climate data for these areas (1 and 2) were available from São Carlos meteorological station, placed 25 km far from the study sites (INMET 2018). In Areas 1 and 2 all individuals of P. elliottii within the invaded patches were sampled.
The third study site (Area 3) was located at SBEcS, a reserve covering 2715 ha (22°46′33″S–22°50′33″S, 49°10′27″W–49°15′36″W, Fig. 1). Monthly temperature ranges from 16 to 24 °C and annual precipitation from 1100 to 1300 mm. The grassland physiognomies (campo cerrado and campo sujo), where trees are scarce, occupy 225 ha. Of these grasslands, approximately 136 ha were lost due to a massive Pinus invasion, especially in the riparian zones (Melo and Durigan 2009). Area 3 (22°47′7.92″S and 49°14′44.21″W) corresponded to a large invaded site composed by 5 blocks (with 10 plots of 10 m × 10 m each) covering around 0.5 ha where the effects of slash pine invasion were assessed in 2010 (Abreu 2013). At this site, we sampled all pine individuals within the plots. In 2011, a wildfire reached the established sample units, and 2 years later we collected the basal cross-sections of all surviving individuals of P. elliottii within the 50 plots. Despite observing many dead saplings in the understory or burned after the wildfire in 2011, we must highlight that we did not observe dead adult trees standing in any of the study sites. A time series of meteorological data for Area 3 was obtained from a meteorological station in Avaré, 45 km from the sample area (INMET 2018).
Growth dynamics, age structure, and fire record
In December 2009, 169 trees were felled in Area 1, and 192 in Area 2. In September 2013, 168 trees were felled in Area 3. In each area, all P. elliottii trees with a basal diameter ≥ 1.0 cm were collected. The total of sampled trees was 529. The samples (wood discs) were obtained with a chainsaw operating close to the tree base (~ 10 cm above ground). The wood discs were polished using an orbital sander. Four radii were delineated, then tree rings were cross-dated within trees to detect wedging and false rings. Cross-dating between trees was not performed because of complacent and short tree-ring width series. Leica® Stereomicroscope was used for tree ring observations. The tree rings dates followed Schulman’s (1956) convention for tree ring studies in the South Hemisphere, where growth ring formation starts in the spring (September–December) and the rings are labeled with the year that they started to develop. We scanned each disc with an HP 2400 scanner and measured ring width with Image Pro Plus software, version 3.0 for Windows (Media Cybernetics Inc). Each individual establishment year was inferred by the year of the first ring, and the tree ages were inferred from the total number of annual rings. Some trees may be older than the age estimated because we did not sample in the exact root-stem boundary. The age structures of the three populations were analyzed by age classes with 2 years range.
For each tree, we calculated the annual diameter increment as the average of the four measured ring widths, multiplied by two. We calculated the cumulative diameter increment by summing the annual diameter increment as the age of the tree increased. For the cumulative basal area increment we used the squared radius (sum of the ring widths per tree) multiplied by π. The annual basal area increment was calculated subtracting the previous year cumulative basal area from the subsequent year. For each site, we calculated the mean annual diameter and the basal area growth rate, the standard deviation, the standard error, and the maximum and minimum values. When comparing growth rates among the study areas, we analyzed the mean annual diameter and basal area increments at different age and size classes to avoid ontogenetic bias. The analysis was performed in ranges of 5 years, from 1 to 20 years (1–5, 6–10, 11–15, 16–20) and six diameter classes (every 10 cm). The non-parametric Kruskal–Wallis test was used in the comparison and the results presented in boxplots. The analyses were performed using the software Statistica 6.0 (StatSoft Inc). To evaluate the range of growth rates in each population, we plotted individual cumulative growth curves by study site. An average cumulative curve was built for each population using the mean annual diameter and the basal area increment values (Stahle et al. 1999; Worbes et al. 2003; Brienen and Zuidema 2006; Schöngart 2008). To evaluate the lifetime growth trends of the trees in each site, we plotted the annual diameter and the basal area increment by the age of the trees.
The fire record was evaluated by the presence of fire scars in the samples collected at Area 3, where a wildfire occurred in the winter of 2011 (17th July). By matching fire scars with the year of the growth rings, we evaluated growth ring periodicity formation (Worbes 1995; Harley et al. 2011). We calculated the mean diameter and the basal area increments in 2009, 2010, 2011 and 2012 to evaluate growth before and after the fire event. To eliminate the influence of size and/or age we standardized the data dividing the annual increment by the mean growth (Speer 2010) from 2009 to 2012. A paired t test with standardized data was performed to evaluate differences in diameter and basal area growth from 2 years before (2009–2010) and 2 two years after (2011–2012) the wildfire event.
The sample processing was done in Wood Anatomy and Dendrochronology Laboratory (LAMAD—Laboratório de Anatomia da Madeira e Dendrocronologia) in Fluminense Federal University (Universidade Federal Fluminense). Samples were deposited in the Rio de Janeiro Botanical Garden Wood Collection (RBw—Xiloteca do Instituto de Pesquisas Jardim Botânico do Rio de Janeiro) and Niterói Herbarium Wood Collection (NITw—Xiloteca do Herbário de Niterói).
Results
Fire records
In Area 3, 48 trees showed fire scars before the formation of early wood of the year 2011 (Fig. 2). The wildfire event that reached the sample site happened in 17th July 2011, before the early wood formation. The fire scars allow us to confirm the annual nature of the growth ring formation due to the matches between the date of the wildfire and the growth ring date. Tree growth was faster in the 2 years before fire than in the 2 years after fire either for diameter increment (t = − 11.252, df = 335, P < 0.0001) or for basal area increment (t = − 6.0853, df = 335, P < 0.0001). Before fire, the average diameter increment was 45–60% and the basal area increment was 22–48% higher than after fire. Diameter increment before fire was 14.7 mm/year (± 0.6 SE) in 2009 and 13.2 mm/year (± 0.7 SE) in 2010. After fire, diameter increment was 8.8 mm/year (± 0.6 SE) in 2011 and 9.1 mm/year (± 0.5 SE) in 2012. The average basal area increment before fire was 3572.1 mm2/year (± 280.0 SE) in 2009 and 3390.7 mm2/year (± 299.4 SE) in 2010. After fire, basal area increased 2409.0 mm2/year (± 207.3 SE) in 2011 and 2780.9 mm2/year (± 255.0 SE) in 2012.

Transverse stereomicroscope section of P. elliottii. The black arrow highlights a fire scar in the year of 2011. Scale bar = 1 cm
Age structures
Area 1
Tree ages ranged from 2 to 19 years. The greatest number of individuals was in the class of 4–5 years with 84 trees, followed by 6–7 years with 53 trees. Trees were absent in two classes 12–13 years and 16–17 years. Young trees were found at low frequency (2–3 years). One founder arrived in the year 1990, and 5 years later (1995) two other founders. After another 5 years (year 2000) the colonization of pine trees started to accelerate. An invasion pulse happened between 2002 and 2005, with the establishment of 137 trees, 7 years after the establishment of the first founders, in 1995 (Fig. 3).

Barplots of the age structure by study site according to the year of establishment of each individual. Number of individuals by age classes of 2 years. The years of establishment in Area 1 and 2 (Itirapina Ecological Station) were from 1988 to 2007, and the years of establishment in Area 3 (Santa Barbara Ecological Station) were from 1992 to 2011
Area 2
Tree ages ranged from 4 to 20 years. The greatest number of individuals was in the class of 18–19 years with 40 trees, followed by 8–9 years with 30 trees. The class with the lowest density was the 20-year class, with seven trees. Young trees were in low abundance (4–7 years). In this area, a continuous colonization started with the arrival of founders in 1989 and with the establishment of 40 trees between 1990 and 1991. Seven years later, an invasion wave occurred between 1998 and 2001, with the establishment of 59 trees (Fig. 3).
Area 3
Tree ages ranged from 4 to 19 years. The highest abundance was found in the class of 12–13 years with 46 trees, followed by 10–11 years with 33 trees. The class with the lowest density was 4–5 years. The founders arrived in the year of 1994 and the first invasion wave happened between 1998 and 2003, 7 years after the first founders, with the establishment of 107 trees (Fig. 3).
Growth dynamics
Area 1
In this area, 3714 annual rings were measured in 169 trees. The analysis period ranged from the year 1990 to the year 2008, covering a 19-year timespan. The mean annual diameter increment was 10.5 mm/year (± 0.3 SE) and basal area increment was 1101.0 mm2/year (± 100.1 SE) (Table 1). Despite the wide range of the annual increments for diameter (from 0.7 to 57.2 mm/year) and for basal area (from 0.7 to 29,359.1 mm2/year), the individual cumulative diameters and basal area curves had similar shapes (Fig. 4).

From the top to the bottom—samples of the growth rings in each study site. Cumulative diameter growth curves of invasive trees of P. elliottii. Diameter growth trends, cumulative basal area growth, basal area growth trends. Gray lines = individual cumulative curves; black line = average curve; gray dots = individual annual increment. Area 1 (IEcS1) and Area 2 (IEcS2) from Itirapina Ecological Station; Area 3 (SBEcS) from Santa Barbara Ecological Station
We observed a pattern of constant increase in diameter growth rate until the tree reaches the age of 10 years (Fig. 4). After this age, we observed an oscillation on the growth rate, but only a few individuals reached more than 10 years of age in this area. During the first 5 years, the diameter growth rate was higher than that observed in Area 2 and lower than that observed in Area 3 (Fig. 5). Considering trees from 6 to 10 years old, the growth rate was higher than observed in Area 2 and similar to that observed in Area 3. We could not analyze the last age classes (the youngest) due to the small number of individuals. Considering the diameter size class 1–10 cm, diameter growth rate in Area 1 was higher than observed in Area 2 and lower than observed in Area 3 (Fig. 6). For all other size classes (11–20, 21–30, 31–40, 41–50 cm), diameter growth rate was higher in Area 1 than observed in Areas 2 and 3.

Boxplots of the average diameter growth rate of trees growing in the study sites by different age classes. Itirapina Ecological Station—Area 1 (IEcS1) and Area 2 (IEcS2); and Santa Barbara Ecological Station—Area 3 (SBEcS). Different letters means difference by Kruskal–Wallis test

Boxplots of the average diameter growth rates of trees by different size classes. Itirapina Ecological Station—Area 1 (IEcS1) and Area 2 (IEcS2); and Santa Barbara Ecological Station—Area 3 (SBEcS). Different letters means difference by Kruskal–Wallis test
Basal Area growth rate exponentially increased until trees reached the age of 10 years (Fig. 4). After this age, an oscillation on basal area increment was detected, but few individuals compound this data. Considering 1–5 years and 6–10 years age classes, basal area increment rate in Area 1 was higher than observed in Area 2 and lower than observed in Area 3 (Fig. 7). In the diameter size class 1–10 cm, basal area growth rate in Area 1 was higher than observed in Area 2 and lower than observed in Area 3 (Fig. 8). For all other size classes (11–20, 21–30, 31–40, 41–50 cm), basal area growth rate was higher in Area 1 than observed in Areas 2 and 3.

Boxplots of the average basal area growth rates of trees by different age classes. Itirapina Ecological Station—Area 1 (IEcS1) and Area 2 (IEcS2); and Santa Barbara Ecological Station—Area 3 (SBEcS). Different letters means difference by Kruskal–Wallis test

Boxplots of the average basal area growth rate of trees by different size classes. Itirapina Ecological Station—Area 1 (IEcS1) and Area 2 (IEcS2); and Santa Barbara Ecological Station—Area 3 (SBEcS). Different letters means statistical difference by Kruskal–Wallis test
Area 2
We measured 9570 tree-rings in 192 trees. The analysis period ranged from the year 1989 to the year 2008, covering 20 years of growth. Annual diameter and basal area increments were 4.5 mm/year (± 0.1 SE) and 336.5 mm2/year (± 9.6 SE), respectively (Table 1). Annual diameter increment ranged from 0.3 to 24.3 mm/year and basal area increment ranged from 0.3 to 3166.5 mm2/year. Both cumulative curves—diameter and basal area—had similar shapes (Fig. 4).
We observed a growth pattern with variations and slow growth rate during all the lifetime of the trees (Fig. 4). Diameter and basal area growth rates showed increase in the first 5 years. After this age, diameter growth rate decreased and basal area growth rate had a small increase. Compared to the other sites, the diameter and the basal area growth rates were the lowest in Area 2 for all age and size classes (Figs. 5, 6, 7, 8).
Area 3
We measured 7726 tree-rings in 168 trees with the analysis period ranging from 1994 to 2012 (covering 19 years). The mean annual diameter and the basal area increments were 15.1 mm/year (± 0.2 SE) and 2683.4 mm2/year (± 70.6 SE), respectively (Table 1). This area had the greatest variation in annual diameter increment (from 0.0 to 58.2 mm/year) and basal area increment (from 0.0 to 32,596.0 mm2/year). The individual cumulative diameter and basal area curves had similar shapes (Fig. 4).
We observed a pattern of increasing growth rate in the first 5 years, followed by a constant decrease in the diameter growth rate and a small increase in the basal area growth rate as the trees got older (Fig. 4). During the first 5 years, the diameter growth rate was higher than observed in the other sites (Fig. 5). When observing trees from 6 to 10 years old, the growth rate was similar to the observed in Area 1 and higher than the observed in Area 2. In the classes of 11–15 and 16–20 years, the growth rate was still higher than that observed in Area 2, but there was a noticeable reduction in the growth rate in the 16–20 years age-class. Considering the size classes, diameter growth rate was higher in Area 3 than observed in the other sites in the size class 1–10 cm (Fig. 6). In the other size classes, basal area growth rate in Area 3 was higher than the observed in Area 2 and lowest than the observed in Area 1.
Basal area growth rate in Area 3 was higher than observed in the other sites for all age classes, and in the first size class (1–10 cm; Figs. 7, 8). However, it was lower than Area 1 and higher than Area 2 in the other size classes.
Discussion
Fire record and effect
We detected a reduction in the increment for both diameter and basal area after the wildfire in most trees, even in trees without fire scars. Diameter and basal area increments after fire (2011 and 2012) were lower than increments recorded before fire (2009 and 2010). In Brazil, many studies demonstrating the presence of annual growth rings in Pinus species, including P. elliottii, were developed in plantations of known age (Trovati and Ferraz 1984; Brito et al. 1986; Palermo et al. 2003, 2013; Pereira and Ahrens 2003; Pereira and Tomaselli 2004; Ferreira and Filho 2009; Coelho and Hosokawa 2010; Elesbão and Schneider 2011; Schneider et al. 2013). However, in the Brazilian savanna, the periodicity of growth rings formation for P. elliottii has not been tested yet.
Fire scars have previously been used to indicate the annual tree-ring formation in P. elliottii in Florida savannas (Harley et al. 2011). We used fire scars in this study to demonstrate that rings formation of P. elliottii was also annual in the invaded Cerrado sites. The wildfire that burned into Area 3 in 2011 was recorded in the wood of 48 P. elliottii trees by fire scars. The fire scars were detected before the early wood of the year 2011. This is expected because the fire occurred in 17th July, in the middle of the winter, historically the coldest and driest month of the year in that region (Melo and Durigan 2009).
It is expected that the Pinus trees present a reduction in growth in the years of fire occurrence (Seifert et al. 2017). During the period after fire, we observed that most adult pine trees were not killed by fire, but had their canopies severely reduced by fire, reducing their photosynthetic activity and growth rates. A similar phenomenon has been observed for native trees in Australian savannas (Murphy et al. 2010). The absence of fire scars in some trees can be explained by the intensity of fire that reached each individual, by the fire type (surface fire, crown fire or ground fire), and by other environmental conditions (Taylor and Skinner 2003; Speer 2010; Brandes et al. 2018) whereby the fire was not able to damage the vascular cambium and produce a fire scar, but still affected the cambial activity, reducing wood production.
Age structure and invasion
Some species of pine are aggressive invaders in the Southern Hemisphere, allowing their inclusion in the category of “transformers” (Simberloff and Rejmánek 2011), due to the radical changes they cause to the native vegetation. Their invasion potential is associated with species attributes like small seed mass, which facilitates wind dispersal, the short juvenile phase allowing them to quickly reach reproductive size (e.g. in less than 10 years), and high seed production (Richardson and Rejmánek 2004). They can change the structure of the native vegetation, promote losses of plant diversity (Zanchetta and Diniz 2006; Abreu and Durigan 2011), and rapidly convert grasslands and savannas into pine forests (Langdon et al. 2010). In the three study areas we verified that the invasion pulses occurred every 5–7 years after the establishment of the first invaders (hereafter founders). This same pattern was found in a previous study that used time-series of aerial photographs and satellite images for historical evaluation (Abreu and Durigan 2011). The invasion of grasslands in pulses was also noticed for other Pinus species in the Chilean Patagonia (Langdon et al. 2010). Those pulses, also called waves of invasion, could be related to climatic conditions like temperature and precipitation, or to founder populations reaching mature size and increasing propagule pressure (Abreu and Durigan 2011; Bourscheid and Reis 2011; Diez et al. 2012; Bechara et al. 2013).
In the year of 1994, the study sites experienced a severe winter including several rare frost episodes (Brando and Durigan 2004; INMET 2018), at the same time that the first founders were established in Area 1 and Area 3. Another severe winter with frost episodes happened in the year 2000 (Brando and Durigan 2004; INMET 2018) and it coincided with a wave of seedling establishment in Areas 2 and 3, as well as an increase in colonization in Area 1. These particularly cold winters were also wetter than average (INMET 2018). Severe winters with frosts and above-average precipitation can play an important role in the pine invasion process, perhaps by lessening the potential for native species to act as a biotic filter (Colautti and MacIsaac 2004). Most Cerrado species are not frost resistant (Brando and Durigan 2004; Hoffmann et al. 2019), and frost may temporarily reduce the ground cover by vegetation and open gaps of bare soil, potentially facilitating the invasion by providing germination microsites (Richardson et al. 1994; Diez et al. 2012). Frost, combined with rainy winters, can promote ideal conditions for pine seed germination as P. elliottii invades preferably places with high water availability (Zanchetta and Diniz 2006; Melo and Durigan 2009; Almeida et al. 2010; Abreu 2013). Pine seed dispersal occurs mostly in autumn and winter (Bechara et al. 2013), resulting in large amounts of viable propagules when frosts occur. Frost and high rainfall, together, could enhance colonization by the pine invaders.
Another important factor related to the invasiveness of the pine trees is the distance from the seed sources (Zenni and Simberloff 2013; Miashike 2015; Pauchard et al. 2016). Both reserves have Pinus plantations in the surrounding areas that act as propagule sources (Zanchetta and Diniz 2006; Melo and Durigan 2009), but despite these nearby propagule sources, we suggest that dispersal limitation restrains population growth rates until the maturation of founder individuals. P. elliottii needs 5–7 years to reach maturity (Zanchetta and Diniz 2006; Zanchetta and Pinheiro 2007). In all study sites, dendrochronology showed that invasion waves happened every 5–7 years after the arrival of the founders. Thus, the founders are likely responsible for the rapid population growth. After they reach maturity, these founders become a new dispersal source in the area (Langdon et al. 2010; Abreu and Durigan 2011), increasing the propagule pressure at the site (Richardson et al. 1994). This period of 7 years since the arrival of the founders and the invasion wave, as the invasion pulse in Area 3 in the year 2000, was also noticed in a previous study at the same reserve of Area 3 (Abreu and Durigan 2011). Based on these findings, the suppression of founders before they reach maturity could hinder the Pinus invasion waves.
The low density of young trees in the sample area indicates low recruitment in the population and limitations for establishment of seedlings in the understory of the pine stands (Abreu and Durigan 2011; Bechara et al. 2013). In the SBEcS, another explanation for this age structure could be related to the wildfire event occurred in 2011 that caused the death of small trees. The absence of young trees after fires reinforces that management using prescribed fire can easily prevent the onset of intense colonization (Pyke et al. 2010; Abreu 2013). In fire-prone savanna ecosystems, where the native plant species are resistant to fire (Lawes et al. 2011), prescribed burning can be a management option. However, as our results showed and as demonstrated by Sah et al. (2010), old P. elliottii trees can survive fire, so other techniques to eradicate them are necessary to limit seed dispersal and reduce new recruitment of the invaders (Abreu 2013).
Growth dynamics
We found different patterns of individual growth among the study areas. At Area 1, the trees showed an increasing trend in diameter and basal area growth rate as they got older, while at Area 2, the trees showed narrow variation in growth among years. The diameter growth rate decreased over the lifetime of trees and the basal area increment rate had a small increase. The highest values of growth rates were recorded in Area 3 for all age classes, and the trees presented a unimodal pattern of growth rates, with diameter growth rates increasing at young ages and declining later, and basal area growth rate increasing at lower rates after 5 years old. This last pattern is also found in pine plantations, where even-aged stands grow quickly until the onset of canopy closure (Worbes 1999). Variation in growth rates was also described for P. montana populations where trees with the highest early growth continually decreased the growth rates as the age increased (Bigler 2016), a pattern similar to that observed in Area 3. Trees with the lowest early growth had slow growth with little variation until they reach the age of 50–150 years, when they experienced abrupt growth releases (Bigler 2016). We found this trend in Area 2, except the growth release, likely because the trees did not reach that age. In Area 1, trees were too young to have the long-term trend assessed. The diameter growth rates found for P. elliottii invaders in our study were similar to the values reported for cultivated P. caribaea in Venezuela (8.5 mm/year) (Worbes 1999), P. taeda (11.3 and 26.8 mm/year) (Inoue et al. 2011) and P. elliottii (10.8 mm/year) (Figueiredo Filho et al. 1994) in Paraná state, Brazil.
Several environmental factors can modulate the radial growth of the invasive P. elliottii trees in the Cerrado. In the native habitat, the southeast USA, the wood production and growth dynamics of P. elliottii are influenced by the water availability (Harley et al. 2011). However, another important factor regulating growth is the light environment and the age of the tree. Reduction in growth rates over time, known as “age trend”, may happen due to physiological and/or environmental conditions (Fritts 1976; Cook 1985; Weiner and Thomas 2001; Coelho and Hosokawa 2010). The age trend was detected in the individuals of Area 3, and also described in an invasive population of P. taeda in southern Brazil (Tomazello-Filho et al. 2017). This growing pattern was also found in the Cerrado native species Hymenaea courbaril L. and H. stignocarpa Hayne (Locosssselli et al. 2017) where the trees presented a fast growth in the first 50 years of life followed by a substantial reduction in the growth rate after that period.
Changes in light availability can influence growth rates. A high light incidence can increase growth rates, while a low light incidence, such as under closed canopy conditions, can reduce growth rates (Worbes 1999). In our findings, the reduction of growth rates was remarkable in the invaded area with the densest population, where the trees experienced fierce competition for light (Area 2). For that reason, despite competition for light not killing the adult pines, we can assume that light availability is an important limiting factor in the growth of invasive pines in the Neotropical Savanna. Moreover, experiments with P. elliottii and P. taeda demonstrated that tree-ring width and diameter growth increased with increased light availability (Pereira and Tomaselli 2004; Coelho and Hosokawa 2010; Inoue et al. 2011). Although we did not evaluate the relationship between light intensity and tree growth in this study, we found a high average growth for most age and size classes in the area at the earliest stage of invasion (Area 1), where the canopy was not fully closed, large trees were less frequent, and sunlight was able to reach some portions of the ground.
Conclusions
The dendroecology of the invasive populations of Pinus elliottii in the Cerrado revealed that, as the trees aged, diameter growth decreased and basal area continued to increase, at lower rates. Fire scars confirmed that the tree rings were formed annually in invasive populations of P. elliottii in the Cerrado. The fire-scar analysis indicated that this species can potentially be used to reconstruct the fire record in invaded areas in the tropics. We also found that the invasion occurred in pulses (waves of invasion), with an intense establishment of individuals every 5–7 years, likely related to periods of seed production and dispersal. The low frequency of young trees indicated limitations to the establishment of seedlings and a reduction of recruitment in the understory. When competition for light intensifies as the canopy closes, the response of the invasive populations is indicated by lower individual diameter growth of adult trees, lower increase in basal area, and reduced recruitment of young individuals. We did not find, however, evidence of self-thinning due to competition that could reduce the population of adult trees, open gaps and allow colonization by shade-tolerant native species. Therefore, the pine monodominance feature of the invaded sites tends to be persistent over time. Further studies to investigate climate-growth relationships are suggested, as well as experimentation to disentangle the factors triggering the invasion waves.
References
Abreu RCR (2013) Ecologia e controle da invasão de Pinus elliottii no campo cerrado. São Carlos : Escola de Engenharia de São Carlos, Universidade de São Paulo. Tese de Doutorado em Ciências da Engenharia Ambiental. https://doi.org/10.11606/T.18.2013.tde-23092013-085713
Abreu RCR, Durigan G (2011) Changes in the plant community of a Brazilian grassland savannah after 22 years of invasion by Pinus elliottii Engelm. Plant Ecol Divers 4:269–278. https://doi.org/10.1080/17550874.2011.594101
Almeida RS, Cielo-Filho R, Souza SCPM, Aguiar OT et al (2010) Campo sujo úmido: fisionomia de Cerrado ameaçada pela contaminação de Pinus elliotti Engelm. na Estação Ecológica de Itapeva, Estado de São Paulo. Rev do Inst Florest, pp 71–91
Bechara FC, Reis A, Bourscheid K et al (2013) Reproductive biology and early establishment of Pinus elliottii var. elliottii in Brazilian sandy coastal plain vegetation: implications for biological invasion. Sci Agric 70:88–92. https://doi.org/10.1590/S0103-90162013000200005
Bigler C (2016) Trade-offs between growth rate, tree size and lifespan of mountain pine (Pinus montana) in the swiss national park. PLoS ONE 11:1–18. https://doi.org/10.1371/journal.pone.0150402
Biondi F (1999) Comparing tree-ring chronologies and repeated timber inventories as forest monitoring tools. Ecol Appl 9:216–227
Biondi F (2001) A 400-year tree-ring chronology from the tropical treeline of North America. Ambio 30:162–166
Bourscheid K, Reis A (2011) Dinâmica da invasão de Pinus elliottii Engelm. em restinga sob processo de restauração ambiental no Parque Florestal do Rio Vermelho, Florianópolis, SC. Biotemas 23:23–30. https://doi.org/10.5007/2175-7925.2010v23n2p23
Brandes AFN, Sánchez-Tapia A, Sansevero JBB et al (2018) Fire records in tree rings of Moquiniastrum polymorphum: potential for reconstructing fire history in the Brazilian Atlantic Forest. Acta Bot Bras. https://doi.org/10.1590/0102-33062018abb0282
Brando PM, Durigan G (2004) Changes in cerrado vegetation after disturbance by frost (São Paulo State, Brazil). Plant Ecol 175:205–215
Brewer JS, Souza FM, Callaway RM, Durigan G (2018) Impact of invasive slash pine (Pinus elliottii) on groundcover vegetation at home and abroad. Biol Invasions 20:2807–2820. https://doi.org/10.1007/s10530-018-1734-z
Brienen RJW, Zuidema PA (2006) The use of tree rings in tropical forest management: projecting timber yields of four Bolivian tree species. For Ecol Manage 226:256–267. https://doi.org/10.1016/j.foreco.2006.01.038
Brito JO, Ferraz ESB, Barrichelo LEC, do Couto HTZ (1986) A adubação mineral e seus efeitos sobre os anéis de crescimento da madeira de Pinus caribaea var. bahamensis. IPEF 32:5–17
Buckley BM, Duangsathaporn K, Palakit K et al (2007) Analyses of growth rings of Pinus merkusii from Lao P.D.R. For Ecol Manage 253:120–127. https://doi.org/10.1016/j.foreco.2007.07.018
Carey JH (1992) Pinus elliottii. In: Fire effects information system (Online). U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). https://www.fs.fed.us/database/feis/plants/tree/pinell/all.html. Accessed on 21 Aug 2019
Coelho VCM, Hosokawa RT (2010) Avaliação da reação de crescimento aos desbastes em Pinus taeda L. Rev Agro@ mbiente 4:42–48
Colautti RI, MacIsaac HJ (2004) A neutral terminology to define “invasive” species. Divers Distrib 10:135–141. https://doi.org/10.1111/j.1366-9516.2004.00061.x
Cook ER (1985) Time series analysis approach to tree ring standardization. Dissertation, University of Arizona
Couralet C, Sassklaassen U, Sterck F et al (2005) Combining dendrochronology and matrix modelling in demographic studies: an evaluation for in Ethiopia. For Ecol Manag 216:317–330. https://doi.org/10.1016/j.foreco.2005.05.065
D’Arrigo R, Watanasak M, Boonchirdchoo S et al (1997) Progress in dendroclimatic studies of Mountain Pine in Northern Thailand. IAWA J 18:433–444. https://doi.org/10.1163/22941932-90001508
Diez JM, D’Antonio CM, Dukes JS et al (2012) Will extreme climatic events facilitate biological invasions? Front Ecol Environ 10:249–257. https://doi.org/10.1890/110137
Elesbão L, Schneider P (2011) Produção do Pinus taeda L. em povoamento desbastado na região dos campos de cima da serra. Rio Grande do Sul. Ciência Florest 21:119–124
Ferreira ATB, Filho MT (2009) Caracterização dos anéis de crescimento de árvores de Pinus caribaea var. hondurensis Barr. et Golf. por densitometria de raios X. Sci For 37:287–298
Figueiredo Filho A, do Machado SA, Figueiredo DJ DJ, Kikuti P (1994) Comparação do crescimento em diâmetro, altura e volume entre árvores resinadas e não resinadas de Pinus elliottii Engelm. var elliottii. Floresta 22:13–24
Fischer FM, Oliveira JM, Dresseno ALP, Pillar VD (2014) The role of invasive pine on changes of plant composition and functional traits in a coastal dune ecosystem. Nat Conserv 12:19–23. https://doi.org/10.4322/natcon.2014.004
Fritts HC (1976) Tree rings and climate. Academic Press, London
Harley G, Grissino-Mayer H, Horn S (2011) The dendrochronology of Pinus elliottii in the lower Florida Keys: chronology development and climate response. Tree Ring Res 67:39–50
Hoffmann WA, Flake SW, Abreu RCR et al (2019) Rare frost events reinforce tropical savanna–forest boundaries. J Ecol 107:468–477. https://doi.org/10.1111/1365-2745.13047
INMET (2018) BDMEP—Banco de Dados Meteorológicos para Ensino e Pesquisa. In: Instituto Nacional de Meteorologia
Inoue MT, Filho AF, Lima R (2011) Influence of vital growth space on height and diameter of Pinus taeda L. Sci For 39:377–385
Johnson WC (1980) Dendrochronological sampling of Pinus oocarpa Shiede near Copan, Honduras: a preliminary note. Biotropica 12:315. https://doi.org/10.2307/2387704
Klink CA, Machado RB (2005) A conservação do Cerrado brasileiro. Megadiversidade 1:147–155
Kronka FJN, Bertolani F, Ponce RH (2005) A cultura do pinus no Brasil. Sociedade Brasileira de Silvicultura, São Paulo
Langdon B, Pauchard A, Aguayo M (2010) Pinus contorta invasion in the Chilean Patagonia: local patterns in a global context. Biol Invasions 12:3961–3971. https://doi.org/10.1007/s10530-010-9817-5
Lawes MJ, Adie H, Russell-Smith J et al (2011) How do small savanna trees avoid stem mortality by fire? The roles of stem diameter, height and bark thickness. Ecosphere. https://doi.org/10.1890/es10-00204.1
Locosssselli GM, Krottenthaler S, Pitsch P et al (2017) Age and growth rate of congeneric tree species (Hymenaea Spp.—Leguminosae) inhabiting different tropical biomes. Erdkunde 71:45–57. https://doi.org/10.3112/erdkunde.2017.01.03
Melo ACG, Durigan G (2009) Plano de Manejo da Estação Ecológica de Santa Bárbara. Águas de Santa. Instituto Florestal/SEMA, São Paulo, 222 pp
Miashike RL (2015) Invasão por Pinus spp. em fisionomias campestres do Cerrado, no estado de São Paulo. Universidade de São Paulo
Miranda HS, Sato MN, Neto WN, Aires FS (2009) Fires in the cerrado, the Brazilian savanna. In: Tropical fire ecology, Springer, Berlin, pp 427–450
Moya R, Filho MT (2009) Wood density variation and tree ring demarcation in Gmelina arborea trees using X-ray densitometry. Cerne 15:92–100
Murphy BP, Russell-Smith J, Prior LD (2010) Frequent fires reduce tree growth in northern Australian savannas: implications for tree demography and carbon sequestration. Glob Chang Biol 16:331–343. https://doi.org/10.1111/j.1365-2486.2009.01933.x
Nuñez MA, Chiuffo MC, Torres A et al (2017) Ecology and management of invasive Pinaceae around the world: progress and challenges. Biol Invasions 19:3099–3120. https://doi.org/10.1007/s10530-017-1483-4
Oliveira-Filho A, Ratter J (2002) Vegetation Physiognomies and Woody Flora of the Cerrado Biome. In: Oliveira PS, Marquis RJ (eds) The cerrados of Brazil: ecology and natural history of a neotropical savanna. Columbia University Press, New York, pp 91–120
Ortega Rodriguez DR, de Andrade GC, Bellote AFJ, Tomazello-Filho M (2018) Effect of pulp and paper mill sludge on the development of 17-year-old loblolly pine (Pinus taeda L.) trees in Southern Brazil. For Ecol Manag 422:179–189. https://doi.org/10.1016/j.foreco.2018.04.016
Palermo GPM, Latorraca JVF, de Rezende MA et al (2003) Análise da densidade da madeira de Pinus elliottii Engelm por meio de radiação gama de acordo com as direções estruturais (longitudinal e radial) e a idade de crescimento. Floresta e Ambient 10:47–57
Palermo GP, Latorraca JVF, Severo ETD et al (2013) Delimitation the juvenile and mature wood of Pinus elliottii Engelm. Rev Árvore 37:191–200
Pauchard A, Escudero A, García RA et al (2016) Pine invasions in treeless environments: dispersal overruns microsite heterogeneity. Ecol Evol 6:447–459. https://doi.org/10.1002/ece3.1877
Pereira J, Ahrens S (2003) Efeito da Desrama sobre a Espessura ea Densidade da Madeira dos Anéis de Crescimento de Pinus taeda L. Bol Pesq Fl, Colombo 46:47–56
Pereira JCD, Tomaselli I (2004) A Influência do Desbaste na Qualidade da Madeira de Pinus elliottii Engelm. var. elliottii. Bol Pesq Fl 49:61–81
Pyke DA, Brooks ML, D’Antonio C (2010) Fire as a restoration tool: a decision framework for predicting the control or enhancement of plants using fire. Restor Ecol 18:274–284. https://doi.org/10.1111/j.1526-100X.2010.00658.x
Richardson DM (2006) Pinus: a model group for unlocking the secrets of alien plant invasions? Preslia 78:375–388
Richardson DM, Higgins SI (1998) Pines as invaders in the southern hemisphere. In: Richardson DM (ed) Ecology and biogeography of Pinus. Cambridge University Press, Cambridge, pp 450–473
Richardson DM, Rejmánek M (2004) Conifers as invasive aliens: a global survey and predictive framework. Divers Distrib 10:321–331. https://doi.org/10.1111/j.1366-9516.2004.00096.x
Richardson DM, Williams PA, Hobbs RJ (1994) Pine invasions in the southern hemisphere: determinants of spread and invadability. J Biogeogr 21:511–527. https://doi.org/10.2307/2845655
Rozendaal DMA, Zuidema PA (2011) Dendroecology in the tropics: a review. Trees 25:3–16. https://doi.org/10.1007/s00468-010-0480-3
Sah JP, Ross MS, Snyder JR, Ogurcak DE (2010) Tree mortality following prescribed fire and a storm surge event in slash pine (Pinus elliottii var. densa) forests in the Florida Keys, USA. Int J For Res 2010:1–13. https://doi.org/10.1155/2010/204795
Sarutanon S, Boonchirdchoo S, Arrigo RD et al (1995) Dendrochronological investigations in Thailand. IAWA J 16:393–409. https://doi.org/10.1163/22941932-90001429
Schneider PR, Elesbão LEG, Schneider PSP, Longhi RV (2013) Crescimento em altura dominante do Pinus elliottii e Pinus taeda em solos arenizados degradados no oeste do Rio Grande do Sul. Ciência Rural 43:1981–1986
Schöngart J (2008) Growth-Oriented Logging (GOL): a new concept towards sustainable forest management in Central Amazonian varzea floodplains. For Ecol Manag 256:46–58. https://doi.org/10.1016/j.foreco.2008.03.037
Schulman E (1944) Dendrochronology in Mexico. Tree-ring Bull 10:18–24
Schulman E (1956) Dendroclimatic changes in semiarid America. University of Arizona Press, Tucson
Seifert T, Meincken M, Odhiambo BO (2017) The effect of surface fire on tree ring growth of Pinus radiata trees. Ann For Sci 74:34. https://doi.org/10.1007/s13595-016-0608-8
Sigal PS (2011) Tropical dendrochronology: exploring tree-rings of Pinus oocarpa in eastern Guatemala. Georg-August-Universität Göttingen
Silva CEF, Reis CM, Zanchetta D et al (2006) Plano de Manejo Integrado. Estações Ecológica e Experimental de Itirapina/SP. Itirapina
Simberloff D (2014) Biological invasions: What’s worth fighting and what can be won? Ecol Eng 65:112–121
Simberloff D, Rejmánek M (2011) Encyclopedia of biological invasions. University of California Press, Berkley
Simberloff D, Nuñez MA, Ledgard NJ et al (2009) Spread and impact of introduced conifers in South America: lessons from other southern hemisphere regions. Aust Ecol 35:489–504. https://doi.org/10.1111/j.1442-9993.2009.02058.x
Sociedade Brasileira de Silvicultura (2008) Fatos e Números do Brasil Florestal. Sociedade Brasileira de Silvicultura, São Paulo
Speer JH (2010) Fundamentals of tree-ring research. The University of Arizona Press, Arizona
Speer JH, Orvis KH, Grissino-Mayer HD et al (2004) Assessing the dendrochronological potential of Pinus occidentalis Swartz in the Cordillera Central of the Dominican Republic. Holocene 14:563–569. https://doi.org/10.1191/0959683604hl732rp
Stahle D, Mushove P, Cleaveland M (1999) Management implications of annual growth rings in Pterocarpus angolensis from Zimbabwe. For Ecol Manage 124:217–229
Taylor AH, Skinner CN (2003) Spatial patterns and controls on historical fire regimes and forest structure in the Klamath Mountains. Ecol Appl 13:704–719. https://doi.org/10.1890/1051-0761(2003)013%5b0704:spacoh%5d2.0.co;2
Tomazello-Filho M, Latorraca JVF, Fischer FM et al (2017) Avaliação da dispersão de sementes de Pinus taeda L. pela análise dos anéis de crescimento de árvores de regeneração Natural. Floresta e Ambient. https://doi.org/10.1590/2179-8087.040913
Trovati LR, Ferraz ESB (1984) Influência da precipitação e da temperatura na densidade dos anéis de crescimento de Pinus oocarpa. IPEF 26:31–36
Vasquez AG, Nogueira AS, Kirchner FF, Berger R (2007) Uma síntese da contribuição do Gênero Pinus para o desenvolvimento sustentável no sul do Brasil. Floresta 37:445–450
Venegas-González A, Chagas MP, Anholetto Júnior CR et al (2015) Sensitivity of tree ring growth to local and large-scale climate variability in a region of Southeastern Brazil. Theor Appl Climatol. https://doi.org/10.1007/s00704-014-1351-4
Vlam M, van der Sleen P, Groenendijk P, Zuidema PA (2017) Tree age distributions reveal large-scale disturbance-recovery cycles in three tropical forests. Front Plant Sci 7:1–12. https://doi.org/10.3389/fpls.2016.01984
Weiner J, Thomas SC (2001) The nature of tree growth and the“age-related decline in forest productivity”. Oikos 94:374–376
Worbes M (1995) How to measure growth dynamics in tropical trees: a review. Iawa J 16:337–351
Worbes M (1999) Annual growth rings, rainfall dependent growth and long-term growth patterns of tropical trees from the Caparo Forest Reserve in Venezuela. J Ecol 87:391–403
Worbes M, Staschel R, Roloff A, Junk W (2003) Tree ring analysis reveals age structure, dynamics and wood production of a natural forest stand in Cameroon. For Ecol Manag 173:105–123
Zanchetta D, Diniz FV (2006) Estudo da contaminação biológica por Pinus spp. em três diferentes áreas na estação ecológica de Itirapina (SP, Brasil). Rev Inst Flor 18:1–14
Zanchetta D, Pinheiro LS (2007) Análise biofísica dos processos envolvidos na invasão biológica de sementes de Pinus elliottii na Estação Ecológica de Itirapina—SP e alternativas de. Climatol Estud Paisag 2:72. http://cecemca.rc.unesp.br/ojs/index.php/climatologia/article/view/230/643
Zenni RD, Simberloff D (2013) Number of source populations as a potential driver of pine invasions in Brazil. Biol Invasions 15:1623–1639. https://doi.org/10.1007/s10530-012-0397-4
Zimmer H, Baker P (2009) Climate and historical stand dynamics in the tropical pine forests of northern Thailand. For Ecol Manag 257:190–198. https://doi.org/10.1016/j.foreco.2008.08.027
Zuidema PA, Vlam M, Chien PD (2011) Ages and long-term growth patterns of four threatened Vietnamese tree species. Trees 25:29–38. https://doi.org/10.1007/s00468-010-0473-2
Acknowledgments
We thank the staff of SBEcS and the staff of IEcS for logistical support. GD and RCRA thank the National Council for Scientific and Technological Development (CNPq) for research grants to GD (#561771/2010 and #312292/2016-3) and Ph.D. grant for RCRA. RCRA also thanks CAPES/FULBRIGHT for a research grant. GAFD thanks PIBIC/UFF for a research grant. Research permit# 260108—013.650/2009 (SMA-SP/IF). The authors thank to Sam Flake for the language review and one anonymous reviewer for the suggestions that helped to improve the MS.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Brandes, A.F.N., Albuquerque, R.P., Domingues, G.A.F. et al. Dendroecology of Pinus elliottii Engelm. reveals waves of invasion in a neotropical savanna. Biol Invasions 22, 403–419 (2020). https://doi.org/10.1007/s10530-019-02099-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-019-02099-2