
Abstract
An eradication program conducted on Rangitoto and Motutapu islands in New Zealand successfully removed stoats (Mustela erminea), cats (Felis catus), hedgehogs (Erinaceus europaeus occidentalis), rabbits (Oryctolagus cuniculus), mice (Musmusculus) and three species of rat (Rattus, R. exulans and R. norvegicus) from an area of 3,842 ha. The project was significant because it was completed so close to Auckland, New Zealand’s largest city, but also, in contrast to many eradication projects, it targeted a suite of invasive mammals in a single operation. To achieve success and avoid conflict in the allocation of resources, target species were prioritized by likelihood of eradication success with resources allocated preferentially to species posing the greatest risk of failure and methods applied in a sequence that allowed each technique to capitalize on its predecessor. Consequences of applying this approach were increased operational efficiency, a shorter operation than planned and reduced project cost. When compared to other projects that targeted the same species but individually, we estimate the Rangitoto and Motutapu project to have cost less than 50 % of the total potential cost if each species had been removed in a discrete operation. Logistical efficiencies created by condensing several operations into one and the use of eradication and detection techniques that targeted multiple species are credited as having the greatest influence on the increased efficiencies observed.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background
Invasive alien species (IAS) are a critical and escalating threat to global biodiversity (McGeoch et al. 2010; Vitousek et al. 1997) and island ecosystems continue to suffer disproportionately from their impacts (Blackburn et al. 2004). Combating the threat of IAS is an enormous challenge and will require the adoption of a range of strategies including biosecurity, control and eradication (Genovesi 2000). Eradication, or the complete removal of an IAS, has proven a particularly effective approach in the battle to preserve island biodiversity (Genovesi 2005) and successful eradication projects have resulted in species and ecosystem recovery (Bellingham et al. 2010). Although generally more cost effective than long term sustained IAS control (Pascal et al. 2008), eradications are capitally intensive (Simberloff 2010) and funding for their implementation scarce (Genovesi 2005). Consequently, new and innovative approaches are required to ensure that available resources are used as efficiently as possible.
Targeting multiple species simultaneously or sequentially over a defined time period is one means by which costs can be reduced (Griffiths 2011). However, planning for an eradication operation that focuses on more than one species requires more extensive consideration of techniques, the impact they may have and the sequence and timing of their deployment. For instance interactions between IAS can affect the efficacy of techniques e.g. competitive exclusion of mice by Rattus spp. was one factor identified by Mackay et al. (2007) as a potential contributor to the higher failure rate observed for mouse eradications. Similarly the presence of one species may hinder the capacity to target another e.g. dogs trained for pig (Sus scrofa) eradication were distracted by goats (Capra hircus) on Santiago Island (Cruz et al. 2005). However, if these factors are taken into account in the project planning process, then the prospects for delivering a successful outcome in a reduced timeframe and cost is likely (Griffiths 2011). Risks posed to endangered species populations by prey switching or the increase in abundance of one invasive species following the removal of another may also be averted (Griffiths 2011).
Building on prior eradication successes on Rangitoto and MotutapuFootnote 1 and experience gained in recent decades including multi-species eradication operations undertaken within predator fenced sanctuaries on the New Zealand (NZ) mainland (e.g. Speedy et al. 2007), eight invasive vertebrate species were successfully removed from Rangitoto and Motutapu (3,842 ha) in a single operation that spanned 2 years. The project, originally envisaged by Miller et al. (1994) and undertaken by the NZ Department of Conservation (DOC), targeted stoats (Mustela erminea), cats (Felis catus), hedgehogs (Erinaceus europaeus occidentalis), rabbits (Oryctolagus cuniculus), mice (Mus musculus) and three species of rat (Rattus norvegicus Footnote 2 R. rattus and R. exulans Footnote 3). The project was structured to maximize the efficacy of each technique and ensure subsequent methods capitalized on their predecessors and in doing so, highlighted the efficiency gains and cost savings that are possible when multiple species are targeted within the same operation. In this paper the methods utilized on Rangitoto and Motutapu and the sequence of their deployment are described along with the rationale for their use. The results of the project are presented and lessons from the project highlighted.
Study site
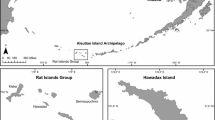
Formed just 600 years ago, the volcanic island of Rangitoto (2,311 ha) lies just 9 km from downtown Auckland, NZ’s largest city of nearly 1.4 million inhabitants (Fig. 1). The island is roughly circular in shape, 259 m high and comprised of gently sloping basaltic lava and an ash base with a central cinder cone. Rangitoto is of international significance for its succession from barren rock and lava to forest which now covers much of the island. Although much older and geologically distinct, Motutapu (1,509 ha) is linked to Rangitoto via a causeway and an extensive area of sand flats exposed at low tide (Fig. 1). Motutapu is a relatively low lying island with a gentle and rolling topography and a coastline dominated by steep coastal cliffs, rocky shore platforms and swampy, steeply backed beaches. Habitat on Motutapu is dominated by pastoral farmland dissected by areas of regenerating native forest, forest remnants and wetlands. The nearest sources of reinvasion for the invasive species targeted are Waiheke Island (4.3 km) and the north shore of Auckland (3 km) (Fig. 1). Both Rangitoto and Motutapu are managed together as an open sanctuary and are freely accessible to the public. The range of activities that occur on the islands is described in Griffiths et al. (2012). Aside from farm dogs (Canis familiaris) and livestock (cattle Bos taurus and sheep Ovis aries) on Motutapu, no other non-native mammals are now present on the islands.

Location of Rangitoto and Motutapu islands
Project feasibility and planning
Before committing resources toward planning and implementation, a feasibility study was commissioned to better understand the likelihood of project success and appraise the project’s biological, social and economic costs and benefits (Griffiths and Towns 2008). Griffiths and Towns (2008) proposed an eradication strategy that would span 3 years and encompass the application of rodent bait to target rats, mice and rabbits followed by the deployment of standard trapping and hunting techniques to remove surviving rabbits, cats, stoats and hedgehogs. This approach was then tested for its technical feasibility, social acceptability and legality. Specific questions regarding technical feasibility were addressed for mice, hedgehogs and rabbits because these species were perceived as being particularly challenging. All three species had been targeted in two recently completed mainland sanctuary projects, Tawharanui (Wedding 2007) and Maungatautari (Speedy et al. 2007), but both operations had failed to remove mice and rabbits, and hedgehogs were still present at Tawharanui. Because of the proximity of the islands to the mainland and the high numbers of visitors, a significant focus for the feasibility study was an assessment of the risks of reinvasion and how these might be mitigated (Griffiths et al. 2012).
Rodents
Other projects had failed to remove mice for unknown reasons (Mackay 2011) and the challenge of removing a highly fecund IAS with a small home range size from an area the size of Rangitoto and Motutapu was widely recognized. For this reason and to dispel concerns about the possible presence of bait shyness or anticoagulant resistance, a laboratory trial was undertaken with 40 mice collected from Rangitoto and Motutapu. Rodenticides, including anticoagulants, had been used in the past for rodent control on Rangitoto and Motutapu. The trial confirmed that Pestoff 20R™ the proposed rodent bait, was highly palatable and found no evidence for anticoagulant resistance. In the trial, three mice survived for between 18 and 21 days after ingesting a lethal dose (Morriss 2007) and this, together with the results of other laboratory trials (e.g. Cleghorn and Griffiths 2002), led to a decision to increase the interval between successive bait applications beyond the normally recommended 7–10 days (Broome et al. 2009) as a means of extending the period of bait availability. An increased application rate for the first application and the addition of a third application of bait was also prescribed because of the potential for inter-specific competition to limit the amount of bait available to mice. By tailoring the baiting strategy to mice the operation was considered over engineered for rats so no specific trials were undertaken for the other rodent species present. Mice were also considered a reliable proxy for anticoagulant resistance because the species’ susceptibility to brodifacoum, as defined by LD50 values, lies midway between that of ship rats and Norway rats (Dubock and Kaukeinen 1978).
Rodents were proposed to be targeted first as it was known from previous projects (e.g. Alterio 1996; Speedy et al. 2007; Torr 2002) that rabbits, stoats, cats and hedgehogs would be affected by primary or secondary poisoning. As with other rodent eradications undertaken in temperate climates (Howald et al. 2007), the operation was planned for winter when rodent population abundance and reproductive activity in New Zealand is at its lowest (King 1990). Prior research had also confirmed that rodent populations on Rangitoto were food limited during this season (Miller and Miller 1995). Aerial bait application was planned as opposed to hand spreading or deploying bait in bait stations, primarily because parts of the islands could not be safely accessed but also because the scale and logistics of a ground based operation targeting both rats and mice would have been prohibitively expensive and, based on the historical eradication record (Howald et al. 2007), posed a greater risk of failure. Permission to target rodents in the presence of both native and non-native species was obtained from DOC (Section 54 of the Wildlife Act 1953), the Auckland Regional Council and the Auckland City Council (Resource Management Act 1991) (Griffiths and Towns 2008). To meet a national Code of Practice (Anonymous 2006), all livestock were removed from the island prior to the application of rodent bait and were not returned until after the specified minimum withholding period of 60 days after bait application.
Rabbits
Rabbits are known to consume rodent bait (Torr 2002), and were considered the species most likely to influence the availability of bait to rodents. Because of this, the density of rabbits was evaluated and a gross estimate of the population made using spotlight counts after Fletcher et al. (1999). This estimate was then used to inform the decision on bait application rates and scale the level of resourcing allocated to the detection and control of survivors. After rodents, rabbits were considered the next highest priority for implementation because of the species’ propensity to breed rapidly (Twigg et al. 1998) and change their behavior when threatened (Jaksic and Soriguer 1981; Lombardi et al. 2003). Based on the results of previous operations (Micol and Jouventin 2002; Torr 2002) most rabbits but not all were expected to die as a result of primary poisoning. To minimize the likelihood of behavioral changes by survivors, the operational strategy specified the initial use of passive methods (those considered least likely to educate surviving animals e.g. other bait types and toxicants) followed by increasingly assertive techniques such as trapping and hunting (Griffiths 2009). Targeting individual survivors with silenced. 22 caliber rifles and where more than one rabbit was found, carrot coated with pindone (‘Pin-25’) (Nelson and Hickling 1994) was planned as was the gassing of burrows known to contain surviving rabbits with magnesium phosphide (Magtoxin® granules). However, these techniques were never used.
Stoats
A decline in the islands’ stoat population as a consequence of the rodent eradication was anticipated, but not assumed (Griffiths and Towns 2008) and the establishment of an island wide 100 m × 400 m network of DOC 200™ traps set into wooden tunnels was proposed. The proposed grid was more intensive than those used in past successful stoat eradication attempts (Elliott et al. 2010; McMurtrie et al. 2011) because of the need to also target hedgehogs.
Hedgehogs
Prior to the Rangitoto and Motutapu project only six attempts to eradicate hedgehogs had been implemented (unpubl. data). Four of these were successful, one (Tawharanui, NZ) had failed, and one (Uists, Scotland, UK) was ongoing (Bremner and Park 2007). Evidence from two of the successful projects suggested that hedgehogs were vulnerable to both primary and secondary poisoning when exposed to Pestoff 20R. However, the level of survivorship that might be encountered during the Rangitoto and Motutapu project was unknown because, as indicated by other studies (e.g. Parkes and Brockie 1977), many individuals within the population were expected to be inactive over the period of rodent bait application. To gain an insight into these factors and guide plans to detect and remove survivors, the over wintering activity and movements of 20 hedgehogs were assessed on Motutapu using radio telemetry. Although periods of inactivity of up to 34 days were recorded, all hedgehogs were active for intervals during the study suggesting that exposure of all individuals within the population to primary and secondary poisoning was possible. Minimum convex polygons generated from fixes of radio tagged hedgehogs (mean 11.3 ha, range 1.4–70.0 ha) and home ranges estimated from other studies (Berry 1999; Moss 1999) indicated that other methods in addition to the trapping grid described above would be required to remove all survivors and ensure eradication success.
Cats
Although not intentionally targeted, it was predicted that the islands’ cat population would be affected but not eliminated by secondary poisoning as a result of the rodent eradication. To target survivors, a trapping program that focused on the islands’ track and road networks, edge habitats and coastline was proposed. The vulnerability of cats to trapping was expected to increase following the removal of rabbits and rodents as their primary prey.
Project implementation
Bait application
Three aerial applications of Pestoff 20R™ rodent bait (10 mm cereal baits containing the second generation anticoagulant, brodifacoum at 20 ppm) were completed, the first over 2 days on 19 and 20 June, the second on 9 July and the third on 6 August 2009 to target rodents and rabbits (Table 1). Bait application rates were 22.1, 9.5 and 6.6 kg/ha respectively and a total of 147,000 kg of bait was used. At the same time as the first bait application, 996 bait stations (15 cm diameter foil dishes) containing Pestoff 20R were deployed in all enclosed manmade structures that exceeded 1 m2 in area. Bait stations were replenished at the same time as the second and third applications and finally removed on 18 September 2009. Approximately 150 kg of bait was used for this purpose.
Accuracy of bait coverage was ensured by a differential global positioning system (GPS). All helicopters were equipped with GPS hardware (AgGPS® TrimFlight 3, Trimble Navigation Limited, Sunnyvale, CA) and a fixed GPS base station positioned at the summit of Rangitoto enabled helicopters to be tracked to within 5 m. The boundary for bait spread was defined as Mean High Water Spring and this boundary was flown and logged before bait was applied. All bait spreading buckets used were calibrated prior to the operation and application rate and the area covered checked each time a helicopter returned to refuel throughout the operation. Actual application rates were assessed using GIS software ESRI® Arc-Map™ (v. 9.3) to map helicopter flight paths and 10 m × 10 m quadrats on the ground.
Targeting survivors
Commencement of detection and control work for surviving rabbits, stoats, hedgehogs and cats aimed to coincide with the anticipated peak of mortality generated by primary or secondary poisoning as a consequence of rodent bait application. Consequently, follow up effort for surviving rabbits began 2 weeks after the second application of rodent bait, and for cats, stoats and hedgehogs, considered more likely to be affected by secondary poisoning, immediately after the last application of rodent bait.
Search effort
Locating surviving rabbits was the initial focus and this was undertaken by searching with spotlights at night and looking for fresh sign during the day. Spotlight searches were undertaken with a helmet mounted or hand held 100 W spotlight (Lightforce Scope Mounted 170 mm Striker 7″ Spotlight) most commonly from four wheeled motorbikes (ATVs) or on foot, but four wheel drive vehicles (4WDs) with an observer on the back were also occasionally used. Within 3 months, spotlighting and searching for sign soon became a less efficient means of detection on Motutapu because of rapid grass growth that occurred after removal of livestock. Although both methods continued to be used, other techniques (outlined below) were instigated earlier than anticipated (Table 1). Livestock were re-established on Motutapu on 20 October 2009 and their numbers were gradually increased over subsequent months but grass length did not return to its pre-operational state until June 2010. Spotlight searches for rabbits and day searches for sign were reinstated after this date. Hedgehogs and cats were opportunistically targeted during spotlight searches conducted for rabbits and later this technique was used specifically to target hedgehogs. Spotlight searches specific for hedgehogs utilized the same equipment, but searching was conducted on foot rather than from ATV and 4WD vehicles.
Trapping
The trap and tunnel design used to target stoats and hedgehogs mirrored the layout described in Waddington and Peters (2012) except that openings in the outer mesh and inner baffle were enlarged to 90 mm (wide) and 110 mm (high) so as not to exclude large hedgehogs. In line with previous stoat eradications (Elliott et al. 2010; McMurtrie et al. 2011), the trap network was pre-baited (baited but traps not set) twice prior to the rodent eradication to minimize any potential for neophobia. Traps were initially baited with fresh rabbit meat and a single hen’s egg to target stoats, but from December 2009 were baited with a variety of different bait types for hedgehogs. All traps were initially checked once every 2 weeks but this was scaled back on Rangitoto from November 2009 to an average of once every 4 months because of the lack of captures there. In contrast 2 weekly trap checks on Motutapu were maintained.
Intensification of trapping grids, repositioning of traps, the use of different trap types, baits and lures were required to complete the eradication of hedgehogs. In January 2010 a more intensive grid of DOC 200™ traps was trialed in three separate locations on Motutapu to confirm the presence of hedgehogs between existing trap lines. Traps were established on a 100 m × 100 m grid in three different habitats, un-grazed pasture, un-grazed regenerating forest and open pasture. Following the capture of two hedgehogs, one in each of the un-grazed sites, a 100 m × 100 m grid consisting of DOC 200™ or DOC 150™ traps set into wooden tunnels and Philproof® ferret tunnels (309 trap sets in total) each containing two Mk VI Fenn™ traps was established across all un-grazed areas of Motutapu (ca. 309 ha). Additional traps were not established in grazed areas because hedgehogs were not captured in this habitat during the trial and these areas were readily searched with indicator dogs. In February 2011, every second line of traps on Rangitoto was removed (310 traps) and used, together with all DOC 200™ traps on Motutapu (411 traps), to form an intensive buffer of traps around all ungrazed areas on Motutapu. These traps were spaced 100 m apart along two parallel lines in grazed pasture, one at 0 m and the other at 100 m from the fenced boundary of all targeted areas.
Several different baits and lures were used to attract hedgehogs to traps. Whiskas® canned fish or chicken based cat food along with fresh rabbit meat, were used most often, but dog biscuits soaked in fish oil were also tried. Peanut butter, used to bait mouse traps, is also attractive to hedgehogs (Thomsen et al. 2000) and, although only used in very small amounts, may have also served as a lure. A blend of corn oil and sardines, and liquid fish fertilizer (Nitrosol® Oceanic) were at times spread outside the trap entrance as a means of luring hedgehogs into trap tunnels. Hedgehog urine was collected but never used.
Cage traps were used initially to catch cats living around some of the islands’ residences but within 2 weeks these were replaced by Oneida Victor®#1.5 Soft Catch™ leg hold traps deployed in ‘cubby sets’ (Wood et al. 2002) along the islands’ track and road networks, edge habitats and coastline. Large pieces of fresh rabbit with fur attached, fish frames and pilchards were used to bait traps. Fish fertilizer was also occasionally used as a scent lure. Cat urine was collected from cats that were shot or trapped but never used. Leg-hold traps (Oneida Victor®#1.5 Soft Catch®) were also used to catch the last surviving rabbit. To target this individual, traps were set into the ground beside a rolled over piece of turf and lured with male rabbit urine. Sand traps, where a section of sand on a beach or trail was regularly raked and checked for footprints, were used to detect cats, stoats, hedgehogs and rabbits.
Little confidence existed that rodents surviving beyond the period of aerial bait application could be effectively detected and removed before re-establishing on the island. However, monitoring was considered important to help identify potential causes of operational failure if survivors were detected. Consequently, in contrast to many other NZ operations, monitoring for surviving rodents on Rangitoto and Motutapu began immediately after bait application. With an island wide 100 m × 400 m network of DOC 200™ traps in place for stoats and hedgehogs, little additional effort was required. DOC 200™ traps were considered an effective means to detect rats but for mice, Victor mouse traps (Easy Set Mouse Trap Model M035, Woodstream Corporation, Lititz, PA), baited with a peanut butter and rolled oat mixture and paired with every DOC 200™ trap, were deployed. Monitoring for rodents across the trapping grid continued until the conclusion of the operation in July 2011 (Table 1). Monitoring for rodents was also undertaken via a permanent surveillance network established in October 2009. This network consisted of 121 tracking tunnels (Russell et al. 2009) placed at all key boat landing sites and dwellings on both islands. Mouse traps were also installed in all inhabited dwellings on both islands.
Indicator dogs
Due to the apparent absence of surviving rabbits, the ‘careful use’ of specifically trained dogs (one dog kept close to the handler at all times) to detect fresh sign was deployed earlier than anticipated, within 1 week of searching for survivors. Similarly, rapid grass growth and the loss of efficacy of previously mentioned search methods meant that the less cautious use of dogs (i.e. multiple dogs ranging further from the handler) was also utilized earlier than anticipated, from September 2009, less than 2 months after work began. Four complete searches of both islands with indicator dogs were subsequently completed. In contrast to the dog searches completed for rabbits, indicator dogs were used at night to search for hedgehogs when animals were active. Just one or two dogs were used at a time and dogs wore a harness with flashing LED lights to facilitate location by the handler. Dogs were trained to stop beside a hedgehog until the handler arrived to euthanize the animal. Although off site trials suggested dogs could detect hedgehogs over much greater distances, for the purposes of estimating search effort, 50 m to either side of the handler’s path was used as the limit of detection. Early on during the operation, tracks were widely spaced (ca. 200 m), but this was reduced to just 50 m later during the operation.
Two complete sweeps of both islands with dogs were undertaken, one in December 2009 and the other in February 2010 to search for stoat sign and four extensive searches for cats were undertaken with dogs between 30 November 2009 and the end of March 2011. To keep dogs stimulated and focused, dogs that were stationed on the islands for the duration of the operation were given regular opportunities (at least once every 2 weeks) to spend time off the island in areas with the target species.
Information management
All search effort was logged using handheld Garmin® GPS 62CSX units and tracks mapped using GIS. Maps were then used to identify gaps in search effort and to guide subsequent work. Aging of rabbits was completed using the methods outlined in Wheeler and King (1980). Any cat, stoat, hedgehog and rabbit found, caught or shot was dissected and if possible its stomach contents examined for signs of rodent bait consumption. Internal hemorrhaging if present was also noted and used as an indication of brodifacoum poisoning. Mesenteric fat deposits for cats were scored as per (Windberg et al. 1991) to provide an index of body condition.
Detecting reinvasion
Prior to the operation DNA samples from all target species, except for Norway rats that could not be trapped, were collected to determine the provenance of individuals found on the islands following completion of the operation. Except for stoats which were the subject of another study (Veale et al. 2012), DNA samples were not obtained from mainland or other island populations of the target species. Analyses completed therefore made the assumption that an invading individual was identifiable by the presence of alleles not apparent within the Rangitoto and Motutapu genotype.
Resource allocation and confirmation of success
Because of the small number of surviving rabbits, the four person rabbit team was scaled back to three staff including one dog handler by September 2009 and then to two staff in December 2009. The discovery of a surviving rabbit in April 2010 led to a temporary increase to three staff but this was reduced to just one part time dog handler from July 2010. Search effort for rabbits ended on 8 February 2011. The decision to end search effort was based on a combination of a lack of sign seen over the preceding 10 months, the age and reproductive condition of the two survivors and the apparent behavioral changes that led to their detection. A team of four staff including the hedgehog dog handler was employed to run both stoat and hedgehog components of the operation and although team composition changed during the project, the size of this team was maintained until work ceased in July 2011. The decision to conclude search effort for stoats and hedgehogs was based on the lack of sign or captures recorded over the preceding 6 months despite the intensification of trapping and search effort. All project team members contributed to this decision. Four staff were initially employed to target cats but the team was reduced to three after 2 months and then to two after 6 months. Cat trapping and search effort was discontinued after February 2011. Confidence that all cats had been removed was provided by the absence of sign and captures over a 17 month period in the face of intensive trapping effort and search effort with indicator dogs.
Project cost
The operational cost of the project was estimated based on expenditure records for past projects completed by DOC and budgeted for and documented in DOC’s SAP software system. Project overheads (all expenses not directly related to labor, materials or third-party expenses) were estimated based on figures presented in DOC’s Annual Report (http://www.doc.govt.nz/publications/about-doc/archive/annual-reports/annual-report-for-year-ended-30-June-2010/14-financial-statements/statement-of-accounting-policies/). The cost efficiency of the project relative to other projects was assessed by comparing the total cost of the Rangitoto and Motutapu project with data for projects that eradicated rodents, cats and rabbits but individually (unpubl. data).
Eradication success
Rodents
Rodent monitoring subsequent to the application of rodent bait revealed evidence of only one possible survivor. Mouse scat was discovered inside a dwelling on Motutapu on 18 April 2010 but no mouse was trapped or further sign observed. Although there was an element of doubt about the age of the droppings and whether they were an artefact left from before the operation, the location where they were discovered had been cleaned shortly before their discovery, suggesting a mouse survived the operation but died before it could be caught. All 15 alleles assessed within the DNA extracted from the feces were present in the Rangitoto and Motutapu genotype. Although it is possible the mouse was a recent immigrant of the same genotype, we consider it more likely to have been a survivor. A Norway rat was detected near the main wharf on Rangitoto, 23 months after the application of rodent bait. DNA comparison was not possible for this species because Norway rats were not trapped prior to the operation but this individual was presumed to be a new arrival because of the intensity of trapping and monitoring that had been completed prior to its detection. Following application of Pestoff 20R in bait stations at the site of this detection, no further rodent sign was recorded.
Rabbits
Rabbit population estimates made prior to project implementation suggested a population size of between 100 and 300 individuals. Following the rodent eradication and the commencement of follow up work on rabbits, just two individuals, both juveniles, are known to have survived (Table 2). During initial spotlight searches, a juvenile was seen twice but was not re-sighted. Three months later, on the 26 October 2009, a juvenile female (ca. 125 days old) that had never bred was observed and then captured in a leg hold trap set for cats. This trap had been reset and baited with a frozen rabbit head the day before. No further detections were made until March 2010 when sign was discovered and 3 weeks later a sub-adult male (ca. 320 days old) was caught in a leg hold trap. A large number of fecal pellets found within 100 m of the trap location indicated that this individual had become at least partially territorial, but sign discovered up to 1.6 km away, suggested that it had also ranged widely prior to the time of its capture. No further sign was found after this time. Aging confirmed the two surviving rabbits had been born prior to the final application of rodent bait and indicated that no breeding occurred after this point.
Stoats
Over the course of the operation a large number of trap nights targeting both stoats and hedgehogs were completed (Table 2). One stoat was found dead and presumed to have died of secondary poisoning after the aerial application of bait but, no surviving stoats were detected or trapped on the island’s trap network. Searching with indicator dogs also failed to discover any evidence of stoats on the islands and we conclude that the island’s stoat population was eliminated as a consequence of the rodent eradication. An individual trapped on Rangitoto in July 2010 was confirmed to be a recent invader and not part of the original population (Veale et al. 2012).
Hedgehogs
Thirty-two hedgehogs were found dead and later confirmed as having being poisoned (Tearne 2010) during ad hoc searches across Motutapu following bait application. Subsequently, a total of 298 hedgehogs were trapped, detected by indicator dogs or found during spotlight searches on Motutapu. The trap nights and search effort expended on hedgehogs are presented in Table 2. Despite past observations of hedgehogs on Rangitoto, no carcasses were recovered or hedgehogs detected there. Based on the recovery of one of 28 marked hedgehogs, Tearne (2010) extrapolated that Motutapu supported a population of 8,246 individuals prior to the eradication. Although the data used by Tearne (2010) were limited, the estimate falls within density estimates made elsewhere in NZ (Jones and Sanders 2005). Based on Tearne’s (2010) figure, we estimate up to 96 % of the population succumbed to either primary or secondary poisoning following the application of rodent bait (Table 2). Brodifacoum residues were found in all five of the hedgehogs found dead after the application of rodent bait and submitted for assay and all 35 of the liver samples collected from surviving hedgehogs suggesting that 100 % of the population was exposed (Tearne 2010).
Seventy-five hedgehogs were trapped in the pre-established network of DOC 200™ traps and a further 49 were caught in Oneida Victor®#1.5 Soft Catch™ traps set for cats. Forty-nine hedgehogs were located during spotlighting and 113 were found at night by dogs. A further 12 hedgehogs were caught in either DOC 150™ traps, DOC 200™ or double set Mk VI Fenn™ traps following intensification of the trap network and the targeting of ungrazed habitat on Motutapu. The first hedgehog juvenile was trapped in January 2010 and 12 more were trapped subsequently. The last hedgehog caught was trapped in a Mk VI Fenn™ trap baited with chicken based cat food on 21 January 2011.
Cats
Two cats were found dead following the application of rodent bait and based on anecdotal reports of the number of cats present prior to the eradication, at least 50 % of the population is thought to have succumbed to primary or secondary poisoning. Fourteen cats were shot by the rabbit team while undertaking spotlight searches from ATVs for rabbits and the last three cats were trapped in leg hold and cage traps in a total of 130,800 trap nights (Table 2). The last three cats were all adults and no evidence of breeding after the rodent eradication was observed.
None of the cats shot or trapped showed signs of anticoagulant poisoning and all appeared to be in excellent condition as indicated by mesentery fat scores (\( \bar{x} = 3,\) n = 17). Observations of cat stomach contents and scavenged non-target species carcasses such as pukeko (Porphyrio melanotus melanotus) and paradise shelduck (Tadorna variegata), suggested that cats were consuming muscle tissue only (Griffiths 2011). Brodifacoum concentrates in the muscle tissue at lower concentrations than in the internal organs (Eason et al. 1996) so cats may have been at lower risk to secondary poisoning through this pathway (Dowding et al. 1999). No cats were trapped or detected on Rangitoto despite records of their presence on the island. Trapping, searches with indicator dogs and the use of passive detection tools failed to reveal any sign of cats after September 2009.
Cost efficiency
The final total operating cost of the Rangitoto/Motutapu eradication was NZ$3.2 M or NZ$4.5 M if overheads are included. This was approximately 6 % less than the NZ$3.4 M forecast for the project’s completion (Griffiths and Towns 2008). Approximately 12 % of total project cost was spent on project planning and environmental compliance with the balance spent on implementation. Forty-six percent of implementation costs were spent on removing rodents with the remainder covering the costs of detecting, controlling and confirming eradication success for the other four target species. Based on data from other projects that targeted rodents, cats and rabbits (unpubl. data), if a unique operation had been conducted independently for rodents, cats, stoats, hedgehogs and rabbits, and if rodents had been targeted last, we estimate the total cost for removing the eight IAS present on Rangitoto and Motutapu to have been approximately NZ$9.7 M. Consequently, by targeting eight IAS in the same operation NZ$5.2 M was saved or 54 % of the estimated total project cost.
Discussion
The Rangitoto and Motutapu IAS eradication was declared successful in August 2011. With its completion, the Rangitoto and Motutapu project set a new benchmark with respect to island size for the removal of mice, ship rats, Pacific rats, hedgehogs, rabbits and stoats although for mice, ship rats and rabbits this has recently been superseded by the Macquarie Island (12,700 ha) project (Springer 2012). Confidence in the project’s success was built up over a period of time during which search effort failed to reveal any sign of the target species. The decision to terminate search effort for each of the target species was based on a combination of factors that included the project team’s knowledge and experience of the target species, the species’ behavior and detectability, the extent and intensity of the search effort expended and the period of time over which sign was not detected. A probability detection model as used by Ramsey et al. (2011) would have provided a more robust framework for making these decisions and could have reduced project costs further. However, the utility of this approach was not appreciated at the time the Rangitoto and Motutapu project was planned.
The possible survival of a mouse past the point at which all rodents should have been eliminated is cause for concern and highlights the importance of learning why individuals of some species are less susceptible to eradication techniques such as the aerial application of rodent bait. Other projects have failed to remove mice for unknown reasons and the challenge of removing an IAS with such high fecundity and small home range size has been noted (Mackay 2011). In our case, an eradication operation, that applied rodent bait at a comparatively high density (cumulatively nearly two baits per m2) and the addition of a third application that extended bait availability out to 7 weeks, successfully reduced the island’s mouse population to a point where despite the survival of at least one individual, it could not recover. A better understanding of the factors influencing survivorship could reduce the need for future eradication projects to be as over engineered as the Rangitoto and Motutapu project was.
Just two rabbits (both juveniles) were caught subsequent to the rodent eradication and it is estimated that upwards of 95 % of all rabbits and 100 % of the adult population succumbed to poisoning as a consequence of eating rodent bait. The survival of individuals was anticipated and the results observed on Rangitoto and Motutapu mirror those of other eradication projects (e.g. Torr 2002). Juvenile rabbits are known to behave differently to adults (Simonetti and Fuentes 1982) and we believe it was these behavioral differences that allowed the two surviving rabbits on Motutapu to evade detection by spotlighting and other methods for such a long period of time. Paradoxically, we suggest that it was other, perhaps also age related, changes in behavior that eventually resulted in the two surviving individuals being detected and caught.
The complete elimination of stoats as a consequence of rodent eradication has now been achieved three times (Griffiths 2011) and highlights the potential of targeting IAS indirectly through their prey. For future projects that aim to eradicate stoats in the presence of invasive rodents, we recommend that stoats are targeted along with rodents through the application of rodent bait containing a second generation anticoagulant. However, although the viability of this approach has been demonstrated multiple times, the survival of hedgehogs and cats after bait application highlights that complete elimination of an IAS via indirect methods is not guaranteed and follow up detection and removal work should always be factored into project planning.
Of the IAS targeted on Rangitoto and Motutapu, hedgehogs were the least well understood as few eradications of this species have been attempted. Consequently, an adaptive approach, encompassing actions such as intensifying trapping grids, changing trap sets and varying lures, was a crucial element in ensuring eradication success. The use of indicator dogs on Motutapu was particularly effective, with more than a third of all hedgehogs being removed by this technique. The strong scent of hedgehogs (Thomsen et al. 2000) allowed them to be readily located by dogs when active at night. However, the use of dogs during the day was abandoned early on during the project because of the difficulties associated with detecting and accessing hedgehogs in underground burrows or thick cover.
Several cats were found dead, presumed poisoned, after the application of rodent bait, and upwards of 50 % of the cat population is estimated to have died as a consequence of the application of rodent bait but, a number of individuals survived and had to be shot or trapped. The presence of surviving cats on Rangitoto and Motutapu is consistent with most projects that have targeted rodents and cats and reinforces the view that elimination of this species solely through direct or indirect poisoning is not guaranteed (Parkes et al. 2014). Surviving cats on Rangitoto and Motutapu appeared to be in good health and did not appear to be affected by anticoagulant poisoning. Griffiths (2011) attributed the health of surviving cats to their feeding on muscle tissue of poisoned birds which contained lower residues of brodifacoum than other body parts.
Although more complex than many IAS eradication projects, the Rangitoto and Motutapu project was completed in a shorter time frame and at less cost than anticipated, highlighting the effectiveness of targeting multiple IAS in the same operation. Like other multi-species eradication projects (Griffiths 2011) the Rangitoto and Motutapu project achieved success in a shorter period of time and at less cost, in this case potentially upwards of 50 % of total project cost, than if an independent operation had been undertaken for each of the IAS present and rodents had been targeted last. The two factors we consider contributed most significantly to this outcome were the logistical efficiencies gained by combining multiple eradication operations into one and using eradication and detection techniques that targeted multiple IAS simultaneously. Logistical efficiencies reduced both time and cost because transport, equipment, personnel and infrastructure were able to be shared within the project. Conducting a single operation on Rangitoto and Motutapu rather than a discrete operation for each of the five groups of IAS also minimized the impact and subsequent cost on external stakeholders such as tourism, farming and education camp operators (Griffiths et al. 2012).
The technique that had the greatest impact on the suite of IAS present was the application of rodent bait containing brodifacoum to target rats and mice. This action alone eradicated four rodent species, eliminated the islands’ stoat population, reduced rabbit and hedgehog abundance by more than 96 % and removed at least 50 % of the island’s estimated cat population. As seen in other projects (e.g. Torr 2002; Towns and Broome 2003) rabbits succumbed to primary poisoning and based on observations, liver residue levels (Tearne 2010) and evidence from other projects (e.g. Speedy et al. 2007) most hedgehogs may have been similarly affected. Stoats may also have been affected by primary poisoning but we consider secondary poisoning to have been the most likely mechanism for the population level impact observed. Secondary poisoning also likely contributed to cat mortality but circumstances as described above permitted others to survive.
As a consequence of primary and secondary poisoning, the follow up effort required for rabbits and cats was reduced and trapping resources established for stoats were refocused on hedgehogs earlier than planned. Deployment of eradication techniques shortly after the anticipated peak of mortality ensured that resources were not expended unnecessarily. As a consequence, trap nights per hectare and the time taken to eradicate cats and stoats were less than for operations where these species were targeted individually (Griffiths 2011). Further cost efficiencies were gained because other methods successfully targeted multiple species. Cats and hedgehogs were both targeted while spotlight searching for rabbits. Stoats and hedgehogs, susceptible to the same trap types, were targeted with the same equipment and hedgehogs were caught in traps set and baited for cats. One of the last two surviving rabbits was also caught in a trap set for cats, a surprising but valuable outcome. Monitoring techniques such as sign searches and sand traps provided presence/absence information for several species and having skilled team members able to recognize the sign of multiple species resulted in more eyes on the ground and increased cumulative search effort.
Prioritization between target species and sequencing of methods created efficiencies and avoided potential conflicts over shared resources. Although no breeding eventuated, giving priority to rabbits in project planning and implementation pre-empted any possibility of rabbit breeding and rapidly provided the confidence necessary to scale back efforts and refocus resources on other species. Similarly, a lack of captures despite baiting traps specifically for stoats provided the impetus to change the focus of trapping to hedgehogs. It is important to note that operational planning for the Rangitoto and Motutapu project did not assume the outcomes observed. Resources were in place at the commencement of the project to manage much higher levels of survivorship and sufficient flexibility was built into the operation to ensure that unexpected scenarios such as the survival of hedgehogs between trap lines could be dealt with. Initial efforts may have thus been over-engineered, but they rapidly provided confidence that the operation was proceeding in line with expectations and allowed informed decisions to be made on when to scale efforts up or down.
Notes
Norway rats were not detected in trapping completed on Rangitoto or Motutapu prior to the eradication but are considered to have been present because of the existence of museum specimens collected from the islands (the most recent in 1977). The detection of a Norway rat (presumed newly arrived) on Rangitoto 23 months after the application of rodent bait lends further weight to the assumption that Norway rats were present.
Rat DNA samples genotyped after completion of the eradication unexpectedly confirmed the presence R. exulans. This species was not known to be present prior to the eradication proceeding.
References
Alterio N (1996) Secondary poisoning of stoats (Mustela erminea), feral ferrets (Mustela furo), and feral house cats (Felis catus) by the anticoagulant poison, brodifacoum. NZ J Zool 23:37
Anonymous (2006). Code of practice: aerial and hand broadcast application of Pestoff® Rodent Bait 20R for the intended eradication of rodents from specified areas of New Zealand. http://www.pestoff.co.nz/pdf/Code%20of%20Practice%2020R.pdf Accessed 21 March 2011
Bellingham PJ, Towns DR, Cameron EK, Davis JJ, Wardle DA, Wilmshurst JM, Mulder CPH (2010) New Zealand island restoration: seabirds, predators, and the importance of history. NZ J Ecol 34(1):115–136
Berry CJJ (1999) European hedgehogs (Erinaceus europaeus L.) and their significance to the ecological restoration of Boundary Stream Mainland Island, Hawkes Bay. M.Sc. Thesis, Victoria University, Wellington
Blackburn TM, Cassey P, Duncan RP, Evans KL, Gaston KJ (2004) Avian extinction and Mammalian Introductions on Oceanic Islands. Science 305:1955–1958
Bremner A, Park K (2007) Public attitudes to the management of invasive non-native species in Scotland. Biol Conserv 139(3):306–314
Broome KG, Brown D, Cox A, Cromarty P, McClelland P, Golding C, Griffiths R, Bell P (2009) Current agreed best practice for rodent eradication—aerial broadcasting poison bait (Version 1.1). New Zealand Department of Conservation Internal Document DOCDM-8390981. Department of Conservation, Wellington
Cleghorn M, Griffiths R (2002) Palatability and efficacy of Pestoff 20R bait on mice from Mokoia Island, Rotorua. DOC Science Internal Series 25. Department of Conservation, Wellington
Cruz F, Donlan CJ, Campbell K, Carrion V (2005) Conservation action in the Galapagos: feral pig (Sus scrofa) eradication from Santiago Island. Biol Conserv 121:473–478
Dowding JE, Murphy EC, Veitch CR (1999) Brodifacoum residues in target and non-target species following an aerial poisoning operation on Motuihe Island, Hauraki Gulf. NZ J Ecol 23(2):207–214
Dubock AC, Kaukeinen DE (1978). Brodifacoum (TALON Rodenticide), a novel concept. In: 8th vertebrate pest conference, pp 127–137
Eason CT, Wright GR, Batcheler D (1996) Anticoagulant effects and the persistence of brodifacoum in possums (Trichosurus vulpecula). NZ J Agric Res 39(3):397–400
Elliott G, Willans M, Edmonds H, Crouchley D (2010) Stoat invasion, eradication and re-invasion of islands in Fiordland. NZ J Zool 37(1):1–12
Fletcher DJ, Moller H, Clapperton BK (1999) Spotlight counts for assessing abundance of rabbits (Oryctolagus cuniculus L.). Wildl Res 26(5):609–620
Genovesi P (2000) Guidelines for eradication of terrestrial vertebrates: a European contribution to the invasive alien species issue. Council of Europe, Strasbourg
Genovesi P (2005) Eradications of invasive alien species in Europe: a review. Biol Invasions 7(1):127–133
Griffiths R (2009) The Rangitoto and Motutapu pest eradication operational plan. Department of Conservation, Auckland
Griffiths R (2011) Targeting multiple species—a more efficient approach to pest eradication. In: Veitch CR, Clout MN, Towns DR (eds) Island invasives: eradication and management. IUCN SSC Invasive Species Specialist Group, Gland, pp 172–176
Griffiths R , Towns DR (2008) The Rangitoto and Motutapu pest eradication—a feasibility study. ISBN 978-0-478-14495-6. Department of Conservation, Auckland
Griffiths R, Buchanan F, Broome K, Butland B (2012) Rangitoto and Motutapu—a starting point for future vertebrate pest eradications on inhabited islands. In Timm RM, Baldwin RA (eds) Proceedings of the 25th vertebrate pest conference. University of California Davis, pp 211–218
Howald G, Donlan CJ et al (2007) Invasive rodent eradication on islands. Conserv Biol 21(5):1258–1268
Jaksic FM, Soriguer RC (1981) Predation upon the European rabbit (Oryctolagus cuniculus) in mediterranean habitats of Chile and Spain: a comparative analysis. J Anim Ecol 50:269–281
Jones C, Sanders MD (2005) European hedgehog. In: Ed C, King M (eds) The handbook of New Zealand mammals, 2nd edn. Oxford University Press, Melbourne, pp 81–94
Julian A (1992) The vegetation pattern of Rangitoto. PhD Thesis, Auckland University, Auckland
King CM (ed) (1990) The handbook of New Zealand mammals. Oxford University Press, Oxford
Lombardi L, Fernández N, Moreno S, Villafuerte R (2003) Habitat related differences in rabbit (Oryctolagus cuniculus) abundance, distribution and activity. J Mammal 84(1):26–36
Mackay JWB (2011) Improving the success of mouse eradication attempts on islands. PhD Thesis, University of Auckland, Auckland
MacKay JW, Russell JC , Murphy EC (2007) Eradicating house mice from islands: successes, failures and the way forward. In: Witmer GW, Pitt W, Fagerstone KA (eds) Managing vertebrate invasive species: proceedings of an international symposium. USDA/APHIS/WS. Fort Collins. National Wildlife Research Center, pp 294–304
McGeoch MA, Butchart SHM, Spear D, Marais E, Kleynhans EJ, Symes A, Chanson J, Hoffmann M (2010) Global indicators of biological invasion: species numbers, biodiversity impact and policy responses. Divers Distrib 16:95–108
McMurtrie P, Edge K-A, Crouchley D, Gleeson D, Willans MJ, Veale A (2011) Eradication of feral stoats (Mustela erminea) from Secretary Island, New Zealand. In: Veitch CR, Clout MN, Towns DR (eds) Island invasives: eradication and management. IUCN SSC Invasive Species Specialist Group, Gland, pp 455–460
Micol T, Jouventin P (2002) Eradication of rats and rabbits from Saint-Paul Island, French Southern Territiories. In: Veitch CR, Clout MN (eds) Turning the tide: the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, Gland, pp 199–205
Miller CJ, Miller TK (1995) Population dynamics and diet of rodents on Rangitoto Island, New Zealand, including the effect of a 1080 poison operation. NZ J Ecol 19:19–27
Miller CJ, Craig JL, Mitchell ND (1994) Ark 2020: a conservation vision for Rangitoto and Motutapu Islands. NZ J R Soc 24(1):65–90
Morriss G (2007). Susceptibility of Rangitoto and Motutapu island house mice to 20R brodifacoum baits. Landcare Research Contract Report LC0607/155. Landcare Research, Lincoln
Moss KA (1999) Diet, nesting behaviour and home range size of the European hedgehog (Erinaceus europaeus) in the braided riverbeds of the Mackenzie Basin, New Zealand. M.Sc. Thesis, University of Canterbury, Christchurch, New Zealand
Mowbray SC (2002) Eradication of introduced Australian marsupials (brushtail possum and brushtailed rock wallaby) from Rangitoto and Motutapu Islands, New Zealand. In: Veitch CR, Clout M (eds) Turning the tide: the eradication of invasive species, pp 350–352
Nelson PC, Hickling GJ (1994) Pindone for rabbit control: efficacy, residue, and costs. In: Halverson WS, Crabb AC (eds) Proceedings of 16th vertebrate pest conference, vol 16. University of California, Davis, pp 217–222
Parkes JP, Brockie RE (1977) Sexual differences in hibernation of hedgehogs in New Zealand. Acta Theriol 22(29):384–386
Parkes J, Fisher P, Robinson S, Aguirre-Muñoz A (2014) Eradication of feral cats from large islands: an assessment of the effort required for success. NZ J Ecol 38(2):307–314
Pascal M, Lorvelec O, Bretagnolle V, Culioli J-M (2008) Improving the breeding success of a colonial seabird: a cost-benefit comparison of the eradication and control of its rat predator. Endanger Species Res 4:267–276
Ramsey DS, Parkes JP, Will D, Hanson CC, Campbell KJ (2011) Quantifying the success of feral cat eradication, San Nicolas Island. Cali. NZ J Ecol 35(2):163
Russell JC, Hasler N, Klette R, Rosenhahn B (2009) Automatic track recognition of footprints for identifying cryptic species. J Ecol 90:2007–2013
Simberloff D (2010) Today Tiritiri Matangi, tomorrow the world. Are we aiming too low in invasives control? In: Veitch CR, Clout MN, Towns DR (eds) Island invasives: eradication and management. IUCN SSC Invasive Species Specialist Group, Gland, pp 4–12
Simonetti JA, Fuentes ER (1982) Microhabitat use by European rabbits (Oryctolagus cuniculus) in central Chile: are adult and juvenile patterns the same? Oecologia 54(1):55–57
Speedy C, Day TD, Innes J (2007) Pest eradication technology—the critical partner to pest exclusion technology: the Maungatautari experience. In: Witmer GW, Pitt WC, Fagerstone KA (eds) Managing vertebrate invasive species: proceedings of an international symposium. U.S. Department of Agriculture/Animal and Plant Health Inspection ServiceNational Wildlife Research Center, Fort Collins, pp 115−126
Springer K (2012) Ecological restoration of Sub-antarctic Macquarie Island. University of California, Davis
Tearne DC (2010) Eradication and population Dynamics of the European hedgehog (Erinaceus europaeus) on Rangitoto and Motutapu Islands. MSc Thesis, University of Auckland, Auckland
Thomsen T, Bowie M , Hickling G. (2000) The potential for eradication of hedgehogs (Erinaceus europaeus) from Quail Island, Banks Peninsula. Wildlife Management Report 20. Lincoln University, Christchurch
Torr N (2002) Eradication of rabbits and mice from subantarctic Enderby and Rose Islands. In: Veitch CR, Clout MN (eds) Turning the tide: the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, Gland, pp 319–328
Towns DR, Broome KG (2003) From small Maria to massive Campbell: 40 years of rat eradications from New Zealand islands. NZ J Zool 30:387–398
Twigg LE, Lowe TJ, Martin GR, Wheeler AG, Gray GS, Griffin SL, O’Reilly CM, Butler TL, Robinson DJ, Hubach PH (1998) The ecology of the European rabbit (Oryctolagus cuniculus) in coastal southern Western Australia. Wildl Res 25(2):97–111
Veale A, Clout M, Gleeson D (2012) Genetic population assignment reveals a long-distance incursion to an island by a stoat (Mustela erminea). Biol Invasions 14(3):735–742
Vitousek PM, Mooney HA, Lubchenco J, Melillo JM (1997) Human domination of Earth’s ecosystems. Science 277:249–299
Waddington P, Peters D (2012) Predator Traps. http://www.predatortraps.com/traps.htmAccessed 2 March 2013
Wedding CJ (2007) Aspects of the impacts of mouse (Mus musculus) control on skinks in Auckland, New Zealand. M.S. Thesis, Massey University, Auckland
Wheeler SH, King DR (1980) The use of eye-lens weights for aging wild rabbits Oryctolagus cuniculus (L.) in Australia. Wildl Res 7(1):79–84
Windberg L, Engeman R, Bromaghin J (1991) Body size and condition of coyotes in southern Texas. J Wildl Dis 27(1):47–52
Wood B, Tershy BR, Hermosillo MA, Donlan CJ, Sanchez-Pacheco JA, Keitt B, Croll DA, Howald G, Biavaschi N (2002) Removing cats from islands in north-west Mexico. In: Veitch CR, Clout MN (eds) Turning the tide: the eradication of invasive species. IUCN SSC Invasive Species Specialist Group, Gland, pp 374–380
Acknowledgments
We would like to thank all of those people and organizations that supported the Rangitoto and Motutapu IAS eradication. These include DOC, the Island Eradication Advisory Group, Auckland Regional Council, Motutapu Restoration Trust, Rangitoto Island Historic Conservation Trust, Rangitoto Island Bach Community Association, Ngai Tai, Ngati Paoa, Kawerau a Maki, Ngati Whatua, Fullers, Motutapu Outdoor Education Camp, Motutapu Farms Ltd, Environment Canterbury, NZ Rotary Club of Newmarket, Auckland Regional Public Health Service, Animal Control Products, Skywork Helicopters Ltd, Ports of Auckland, SeaLink Travel Group NZ Ltd and others. Without the support of these organizations and the staff and individuals that work for them the project would not have been the success it has been.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Griffiths, R., Buchanan, F., Broome, K. et al. Successful eradication of invasive vertebrates on Rangitoto and Motutapu Islands, New Zealand. Biol Invasions 17, 1355–1369 (2015). https://doi.org/10.1007/s10530-014-0798-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10530-014-0798-7