
Abstract
The effects of five diets consisting of two natural prey (Acanthococcus abaii Danzig and Aganoscena pistaciae Burckhardt et Lauterer) and three factitious foods (frozen eggs of Ephestia kuehniella Zeller, a mixture of frozen corn pollen with eggs of E. kuehniella and a diet consisting of eggs of Sitotroga cerealella Olivier) on the development, survival and reproduction of Chilocorus bipustulatus L. were investigated. The results revealed that this predator successfully completed immature developmental procedures on the offered foods, but developmental duration was affected. Total larval survival of C. bipustulatus was higher on natural diets than factitious diets. Fresh weights of the newly emerged female and male coccinellids were also affected by the diet offered to this predator during their larval stage. The highest intrinsic rate of increase (r), the net reproductive rate (R 0) and the finite rate of increase (λ) were obtained on A. pistaciae, and these values were significantly different from A. abaii.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Chilocorus bipustulatus L. (Coleoptera: Coccinellidae) is a predatory coccinellid that is widely distributed in Europe, the Eastern Palaearctic region, Near East and North Africa. This coccinellid is a polyphagous predator that preys on aphids, armored scales and other scale families such as Eriococcidae, Coccidae, Asterolecaniidae and Pseudococcidae (Stathas et al. 2003, 2009; Eliopoulos et al. 2010) and more rarely on whiteflies (Yigit et al. 2003). This predator has been considered for augmentative release because of its wide host range (Gaillot 1967; Laudeho et al. 1970). However, its polyphagous nature may also include environmental risks when the insect is used in areas where it is not native (De Clercq et al. 2002; van Lenteren et al. 2003). The effects of different diets and temperatures on the development and reproduction of C. bipustulatus have received attention in several studies (Rosen and Gerson 1965; Hattingh and Samways 1993; Uygun and Elekcioglu 1998). However, information on the qualitative and quantitative effects of nutrition on the life table parameters of C. bipustulatus remains scarce. Acanthococcus abaii Danzig (Hemiptera: Eriococcidae) is a key pest of the saxaul plants Haloxylon aphyllum L., which is the main plant in desert areas of the Kerman, Sistan and Balouchestan provinces of Iran (Miller and Gimpel 2000; Moghadam and Tvakoli 2010). The common pistachio psylla, Agonoscena pistaciae Burckhardt et Lauterer (Hemiptera: Psyllidae), is an important pest of pistachio trees that damages pistachio orchards every year in Iran (Mehrnejad and Copland 2006; Panahi et al. 2013).
It is necessary to know the effects of different diets on the growth and fecundity of the biocontrol agents to manage a successful biological control program. The nutritional value or food quality of prey or artificial diets is critical for the fitness of most predators. Parameters such as immature development, consumption capacity and egg production in response to feeding are used to define the food quality of prey. Therefore, a good understanding of the feeding habits and nutritional needs of a natural enemy is elementary for its implementation in biological control strategies (Hattingh and Samways 1993; Mahdian et al. 2005). There is much interest in food studies and their relationships with coccinellids as natural enemies of aphids, scales and mites among specialists of biological controls (Lucas et al. 2002; Villanueva et al. 2004; Evans 2009; Hodek and Honek 2009). The use of alternative prey may make the production of natural enemies for biological control more cost effective and reduce problems with space for the mass rearing of predators (De Clercq et al. 2005). Adult reproduction depends closely on prey suitability for larval development, but larvae and adults may differ in nutritional requirements (Michaud 2005). Eggs of the Ephestia kuehniella Zeller (Lepidoptera: Pyralidae) and Sitotroga cerealella (Olivier) (Lepidoptera: Gelechiidae) are commonly used as alternative diets for the laboratory production of several coccinellids (Specty et al. 2003; Hamasaki and Matsui 2006; Berkvens et al. 2008b). A mixture of frozen, moist bee pollen and E. kuehniella eggs is a suitable alternative diet for the development and reproduction of lady beetles (Jalali et al. 2010). Evaluations of demographic parameters may be a useful tool for analyzing and understanding the effect of external factors such as food on the development, survival, reproduction, and intrinsic rate of increase of insect populations to provide a comprehensive description of the fecundity of predator populations (Chi and Su 2006; Farhadi et al. 2011). The present study evaluated the effects of different natural prey and factitious food on survival, development, longevity, reproduction and the intrinsic growth rate of C. bipustulatus. The results may improve rearing methods of C. bipustulatus as a biocontrol agent.
Materials and methods
Predator rearing
A laboratory colony of C. bipustulatus was established in May–June 2014 using individuals originally collected from pistachio orchards and Haloxylon shrubs near Rafsanjan, Kerman province, Iran. The food of the stock colony of predators primarily consisted of nymphs and adult A. abaii. Insects were reared in a growth chamber at 25 ± 1 °C, 60 ± 5 % RH, and a photoperiod of 16:8 (L:D) h. The coccinellids were maintained in transparent plastic containers (25 × 20 × 10 cm) covered by a lid with a fine mesh for ventilation. A piece of moist cotton in a 1-cm-diameter plastic dish served as a source of water for the ladybirds.
Natural diets
Two natural diets including A. abaii and A. pistaciae were used as natural foods, which were collected daily from saxaul plants and pistachio orchards, respectively, and used in the experiments. Both prey were collected from a place with no insecticide treatment. The predators were supplied ad libitum with some branches of saxaul plants and two leaves of Pistaciae vera infested with A. abaii and A. pistaciae, respectively.
Factitious food
Mediterranean flour moth larvae were reared on wheat flour under laboratory conditions. The eggs of E. kuehniella were collected daily and stored in a deep freeze at −18 °C for no longer than one month. Frozen eggs of S. cerealella were obtained from the Iranian Pistachio Research Institute, Rafsanjan, Iran, and were stored in a deep freeze at −18 °C. All experiments were performed under laboratory conditions at 25 ± 1 °C, 60 ± 5 % RH, and a photoperiod of 16:8 (L:D) h.
Effect of diets on larval development and survival of C. bipustulatus
Immature development of C. bipustulatus was monitored on each of five diets: female adults of A. abaii, fifth-instar nymphs and adults of A. pistaciae, frozen eggs of S. cerealella, frozen eggs of the Mediterranean flour moth E. kuehniella and a mixture of frozen corn pollen with eggs of E. kuehniella. Monitoring of larval development began with first instar larvae. Batches of C. bipustulatus eggs (<24 h old) were collected from the stock colony for the experiments on developmental time. Thirty-five newly molted first instars of C. bipustulatus were randomly taken from laboratory cultures for each treatment and placed individually in Petri dishes (9 cm in diameter), covered with a fine mesh lid for ventilation and furnished with absorbent papers. A moist paper plug in a small cup (1 cm in diameter) supplied water. Each factitious diet was supplied ad libitum in a plastic cup (2.5 cm in diameter). Infested foliage of H. aphyllum and P. vera L. with A. abaii and A. pistaciae, respectively, was supplied ad libitum for the natural diets. The developmental time and survival of each C. bipustulatus larval instar were recorded, and adults were sexed and weighed on a Sartorius digital balance (±0.01 mg) upon emergence.
Effects of diets on C. bipustulatus reproduction
Reproductive capacities of the predators were evaluated on all diets that yielded development to the adult stage. Adults obtained from developmental tests upon emergence were placed together in rearing containers and were given the same food as in the larval stage. Twelve pairs of adults from each diet were collected from rearing containers after the first mating was observed. Mating pairs were isolated in Petri dishes (12 cm in diameter) furnished with an absorbent paper. A moist paper plug in a small cup provided water. Natural and factitious foods were presented in excess and replaced daily. Oviposition and survival were monitored daily until 41 days after the adult females began oviposition. Males that died before females were replaced by males reared under the same conditions, and females that died within five days were omitted from analyses. The experiments were performed in growth chambers at 25 ± 1 °C with a RH of 60 ± 5 % and a photoperiod of 16:8 (L:D) h.
Statistical analysis
The effect of different diets on developmental times and adult weights of C. bipustulatus was analyzed using one-way analysis of variance (ANOVA) followed by Duncan’s multiple range test (P = 0.05). No reproduction was observed with factitious diets. Therefore, reproductive parameters of C. bipustulatus were compared between two natural diets using Student’s t-test. Data were initially tested for normality (Kolmogorov–Smirnov test) and homoscedasticity (Levene’s test) prior to ANOVA. Sex ratios were analyzed using a χ2 test. The raw life history data for C. bipustulatus were analyzed based on the theory of the age-stage, two-sex life table using TWOSEX-MSChart (Chi 2013). The standard errors of the life table parameters were estimated using the bootstrap technique (Ebrahimi et al. 2013; Akca et al. 2015). The age-stage specific survival rate (s xj , the survivorship of the newborn eggs to age x and stage j), age-stage specific fecundity (f xj , the mean fecundity of individuals of age x and stage j), age-stage total fecundity (Ft xj , the total fecundity of all individuals of age x and stage j), age-specific survival rate (l x , the probability of a newly laid egg survives to age x), age-specific fecundity (m x , the mean fecundity of individuals at age x), age-stage life expectancy (e xj , the individuals of age x and stage j that are expected to be alive), age-specific maternity (l x m x ) and the life history parameters (r, the intrinsic rate of increase, λ, the finite rate of increase, R 0, the net reproductive rate and GRR, gross reproductive rate) were calculated. Student’s t-test was used to compare the life history parameters of C. bipustulatus between the two different natural diets.
Results
Effect of diets on developmental time, sex ratio and adult weight
The results revealed significant effects of all tested diets on developmental duration in C. bipustulatus (F = 37.37; df = 4,126; P < 0.05). All instars successfully completed their life cycle on the foods offered. The mean total developmental time of C. bipustulatus from first instar to adult was 22.92 ± 0.4–29.00 ± 0.3 days (Table 1). The longest and shortest total developmental times of C. bipustulatus were on eggs of S. cerealella and the diet of frozen corn pollen mixed with eggs of E. kuehniella, respectively (22.92 ± 0.4 vs. 29.0 ± 0.3 days) (Table 1). Significant differences (F = 7.37; df = 4,157; P < 0.05) in total developmental time of C. bipustulatus larvae were detected when fed on different natural and factitious diets. The results demonstrated that the diets had a significant effect on the development time of all different instar larvae from first to fourth instar. There was no significant difference in the developmental time of the prepupal stage (F = 0.981; df = 4,170; P > 0.05), but the diets affected the duration of the pupal stage (F = 4.68; df = 4,127; P < 0.05). The different diets did not affect the sex ratios of C. bipustulatus adults (χ2 = 2.21; df = 4; P > 0.05) (Table 1). The fresh weights of newly emerged female and male coccinellids were affected by the diet offered to the predator during their larval stage (Male: F = 7.21; df = 4,58; P < 0.05 and Female: F = 22.25; df = 4,68; P < 0.05). Student’s t-tests revealed significant differences (t = 4.78; df = 107; P < 0.05) between the weight of adult males and females for all diets, and males exhibited lower weights than females (Fig. 1).

Means of adult weights (mg) of Chilocorus bipustulatus fed different natural and factitious diets. Bars (±SE) with the same letter are not significantly different according to Duncan’s multiple range test (for diets with lower case) (P > 0.05) and Student’s t-test (for sex with uppercase) (P > 0.05)
Effect of diets on larval, prepupal and pupal survival and longevity
The highest percentage of mortality was observed in the first instar larvae of C. bipustulatus. Total larval survival of C. bipustulatus was higher on natural diets than on factitious diets. Survival of first instar of C. bipustulatus fed on A. pistaciae (88.57 %) was significantly lower than first instar fed on A. abaii (91.43 %), factitious diets consisting of E. kuehniella eggs, a mixture of frozen corn pollen or E. kuehniella eggs (91.43 %) and S. cerealella eggs (97.14 %) (F = 43.56; df = 4,10; P < 0.05). No mortality was observed on any diet from pre-pupa to adult emergence. There were no significant differences in survival of C. bipustulatus on live prey used in our study. The survival rate of pre-pupa, pupa and adults was 100 %, and the lowest survival rate was observed in first instar larva on all diets. The significantly highest and lowest total survivals of C. bipustulatus were observed on A. pistaciae (77.14 %) and the diet of eggs of S. cerealella (62.86 %), respectively (F = 95.43; df = 4,10; P < 0.05) (Table 2).
There were significant differences in adult longevity of C. bipustulatus fed different diets (male: F = 397.82; df = 4,51; P < 0.05 and female: F = 589.21; df = 4,68; P < 0.05), but no significant differences were observed between the different sexes. Longevity was significantly prolonged with factitious foods (F = 20.24; df = 2,71; P < 0.05) when a mixture of frozen corn pollen and eggs of E. kuehniella were provided as food (Fig. 2). The longest and shortest longevities of male and female adults occurred in C. bipustulatus fed A. abaii and eggs of S. cerealella, respectively.

Means of adult longevity (day) of Chilocorus bipustulatus fed different diets at 25 ± 1 °C. Bars (±SE) with the same letter are not significantly different according to Duncan’s multiple range test (for diets with lower case) (P > 0.05) and Student’s t-test (for sex with uppercase) (P > 0.05)
Effect of diets on egg hatching and oviposition
No reproduction was observed in C. bipustulatus fed factitious diets, but C. bipustulatus fed natural prey, including A. abaii and A. pistaciae, reproduced. The egg incubation period for C. bipustulatus fed A. abaii and A. pistaciae was significantly different (t = 4.45; df = 68; P < 0.05) and ranged from 5.06 ± 0.09 to 3.40 ± 0.08 days, respectively (Table 1). The percentage of egg hatching of C. bipustulatus fed A. pistaciae was 76.75 ± 5.51 %, which was significantly less than C. bipustulatus fed A. abaii (94.72 ± 1.59 %) (t = 17.05; df = 68; P > 0.05). Student’s t-tests revealed that the mean adult preoviposition period APOP of C. bipustulatus was significantly different in the A. abaii and A. pistaciae group (t = 7.20; df = 26; P < 0.05). There was no significant difference between the means of total preoviposition period TPOP of C. bipustulatus reared on natural diets (t = 0.735; df = 26; P > 0.05) (Table 3). The fecundity of C. bipustulatus fed A. abaii (473.07 ± 13.26 eggs) was higher (t = 19.09; df = 26; P < 0.05) than the fecundity on A. pistaciae (425.98 ± 10.4 eggs) (Table 3). Daily oviposition of C. bipustulatus revealed that the maximum oviposition of females fed A. abaii occurred on day 15 with 17.07 eggs and day 17 with 19.06 eggs for A. pistaciae (Fig. 3).

Daily oviposition of Chilocorus bipustulatus fed two natural diets at 25 ± 1 °C
Effect of diets on life table parameters
Table 4 shows the life table parameters of C. bipustulatus reared on two natural prey, A. abaii and A. pistaciae. The results indicated that diet affected all parameters (Student’s t-test, P = 0.05) except GRR. Net reproduction rate (R 0), intrinsic rate of increase (r) and finite rate of increase (λ) of C. bipustulatus on A. pistaciae were higher compared to predators fed A. abaii (R 0: t = 52.75; df = 22; P < 0.05) (r: t = 45.00; df = 22; P < 0.05) (λ: t = 427.00; df = 22; P < 0.05). The age-stage fecundity (f xj ) and age-stage total fecundity (Ft xj ) of C. bipustulatus fed A. pistaciae began earlier (at 30 days) than A. abaii (33 days). The highest female fecundity of C. bipustulatus occurred on the 43rd and 45th day on A. abaii and A. pistaciae, respectively (Fig. 4).

Age-stage fecundity (f xj ) and age-stage total fecundity (Ft xj ) of Chilocorus bipustulatus fed two natural diets at 25 ± 1 °C, Acanthococcus abaii (a) and Aganoscena pistaciae (b)
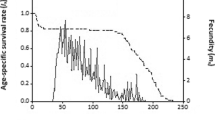
The variability of developmental rate between individuals was clearly observed in the overlaps of age-stage survival rates (S xj ). The survival rate on both diets decreased with increasing age of C. bipustulatus. Age-stage survival rates (S xj ) of C. bipustulatus were 0.485 and 0.428 when feeding on A. abaii and A. pistaciae, respectively (Fig. 5). The highest age-specific fecundities (m x ) of C. bipustulatus were 7.4 and 13 offspring on A. abaii and A. pistaciae, respectively. The highest fecundity was observed in l x m x and m x curves in females reared on A. pistaciae (Fig. 6).

Age-stage survival rate (S xj ) of Chilocorus bipustulatus fed two natural diets at 25 ± 1 °C, Acanthococcus abaii (a) and Aganoscena pistaciae (b)

Age-specific survival rate (l x ), age-specific fecundity (m x ) and the age-specific maternity (l x m x ) of Chilocorus bipustulatus fed two natural diets at 25 ± 1 °C, Acanthococcus abaii (a) and Aganoscena pistaciae (b)
The life expectancy of a newly laid egg in the A. abaii and A. pistaciae groups was 64.11 and 57.51 days, respectively, and these expectancies decreased with increasing age. The life expectancy of C. bipustulatus was higher for females compared to males in both diets. The peak of reproductive values for both diets (A. pistaciae and A. abaii) occurred on days 39 and 40, respectively, and females of C. bipustulatus made the greatest contribution to the population.
Discussion
The predatory coccinellid C. bipustulatus may be reared on live prey in the laboratory. Nymphs and adults of many diaspidids and pseudococcids were successfully used as prey for the laboratory rearing of C. bipustulatus (Uygun and Elekcioglu 1998; Eliopoulos et al. 2010). Prey types for the Coccinellidae are classified as “essential” if the predators complete their larval development and reproduce on it or “alternative” if they provide only a source of energy for adult survival (Hodek and Honek 1996). A. pistaciae and A. abaii were essential prey for C. bipustulatus in the present study. Michaud (2005) suggested a difference between the nutritional requirements of larvae and adults. A diet that is suitable for the development of larval stages of insects may not be suitable for reproduction. C. bipustulatus completed its immature development on the alternative and natural prey diets at 25 ± 1 °C in the current study. Our results demonstrated that different diets significantly influenced the developmental time. Uygun and Elekcioglu (1998) reported that the developmental time of C. bipustulatus fed Aonidiella aurantii (Maskell) (Hemiptera: Diaspididae) and Aspidiotus nerii (Bouche) (Hemiptera: Diaspididae) was 27.8 ± 0.40 and 26 ± 0.41 days, respectively. The development of C. bipustulatus in the current study was slower on A. abaii and faster on A. pistaciae. The developmental times of C. bipustulatus on live prey in this study were close to the times reported by Uygun and Elekcioglu (1998), but they were shorter than C. bipustulatus reared on A. nerii and Chrysomphalus aonidum L. (Hemiptera: Diaspididae) from some previously published studies (Nadel and Biron 1964; Eliopoulos et al. 2010).
Eggs of E. kuehniella and a mixture of eggs of E. kuehniella and pollen were successfully used as factitious food for the laboratory rearing of several insect predators, including coccinellids (Berkvens et al. 2008a; Maes et al. 2014). C. bipustulatus fed a mixture of eggs of E. kuehniella and pollen developed faster than C. bipustulatus fed E. kuehniella eggs. C. bipustulatus reared on other diets did not successfully produce eggs. However, eggs of S. cerealella supported C. bipustulatus growth but not reproduction. This result may be explained by the fact that adults of C. bipustulatus have difficulty handling this type of factitious food, likely because of the small egg size, and cannot extract nutrients at an appropriate rate to achieve reproduction. Females of C. bipustulatus produced more egg numbers on A. abaii than A. pistaciae. The current study revealed that the total developmental time of immature stages reared on eggs of E. kuehniella was significantly lower than C. bipustulatus reared on a mixture of frozen corn pollen and eggs of E. kuehniella. These findings are consistent with Bonte et al. (2010), who found that the developmental time of the immature stage of Adalia bipunctata L. (Coleoptera: Coccinellidae) on eggs of E. kuehniella was lower than A. bipunctata L. fed a mixture of pollen and eggs of E. kuehniella. The results of the present study of the effect of eggs of E. kuehniella and a mixture of pollen and eggs of E. kuehniella on adult weight and developmental time of C. bipustulatus are consistent with earlier observations of De Clercq et al. (2005), who showed that A. bipunctata developmental time and adult weight were affected by these two diets. The authors indicated that the developmental time of A. bipunctata on eggs of E. kuehniella was significantly longer than A. bipunctata on a mixture of frozen corn pollen and eggs of E. kuehniella. They also demonstrated that the weight of adults of A. bipunctata was higher on a mixture frozen corn pollen and eggs of E. kuehniella than A. bipunctata fed eggs of E. kuehniella. It is encouraging to compare the adult body weight and survival rate of C. bipustulatus in this study with the values found by Hattingh and Samways (1993), which were 6.7 ± 0.2 mg and 69 % when C. bipustulatus fed on A. nerii and Aspidiotus miliaris L. (Hemiptera Diaspididae), respectively. Comparisons of the survival of C. bipustulatus that fed on A. pistaciae (77 %) and A. abaii (74 %) in the current study with survival in a previous study (Hattingh and Samways 1993) revealed that the survival of C. bipustulatus was higher. However, the mean weights of adults of C. bipustulatus reared on A. pistaciae (5.49 ± 0.149 mg) and A. abaii (4.86 ± 0.729 mg) were lower than adults that fed on A. nerii and higher than adults fed A. miliaris.
Life table analysis for estimating the demographic parameters is the most common basic procedure for investigations of insect population dynamics and predicting population growth (Carey 2001; Stark et al. 2007). Adult and total pre-oviposition periods of C. bipustulatus females in the current study were reduced when fed eriococcid A. abaii. This finding corroborates the results of Uygun and Elekcioglu (1998), who demonstrated that the pre-oviposition period of C. bipustulatus was the lowest (9.6 ± 0.26 days) when fed A. nerii compared to other diets (A. aurantii: 12.8 ± 0.43 days and Pseudaulacaspis pentagona (Targioni Tozzetti) (Hemiptera: Diaspididae): 12.2 ± 0.43 days). The curves of age-stage specific survival rates (S xj ) of C. bipustulatus revealed a survivorship differentiation and variable developmental rates. For example, the probability that a newborn egg of C. bipustulatus will survive to the adult stage was 0.428 for females on A. pistaciae and 0.485 for females on A. abaii. The intrinsic rate of increase (r) is a useful indicator for comparing populations of different species or the same population on different diets (Gabre et al. 2005). The intrinsic rate of C. bipustulatus on A. pistaciae in the present study was higher than C. bipustulatus reared on A. abaii. The GRR is a simple summation of m x at the beginning of reproduction, and m x is calculated based on the fecundity of all surviving females. However, m x at older ages is generally calculated based on the fecundity of a few surviving females, sometimes even a single female. Therefore, m x at older ages contributes significantly less to the population. No significant difference was observed in the GRR of C. bipustulatus on both natural diets (t = 0.819; df = 22; P > 0.05).
Nutrients and food preference play important roles in the developmental rate and fecundity of most species of Coccinellidae. However, generalist predators usually exhibit some degree of nutritional stringency and may display distinct food preferences in choice situations (Matthews and Matthews 1978; De Clercq 2002). Knowledge of life table parameters is necessary to provide a better understanding of predator population dynamics. The data of the current study suggest that all instar larvae of C. bipustulatus successfully completed their life cycle on the foods offered. However, the biological parameters of C. bipustulatus on A. pistaciae were inferior to A. abaii, and survival of C. bipustulatus on A. pistaciae was higher. Survival rate, developmental rate and fecundity describe the life history, and differences between stages are used to predict growth rate. The intrinsic rate of increase (r) is a practical parameter to compare the potential of insect populations under different situations in pest management. In conclusion, our results demonstrate that the lady beetle, C. bipustulatus, is an effective biocontrol agent of A. pistaciae and A. abaii, and these beetles developed successfully to the adult stage and reproduced. This study aids the development of pest control management, and this type of laboratory study provides better insights into the management of the population dynamics of predators.
References
Akca I, Tamer A, Eda Y, Cecil LS, Chi H (2015) Demography and population projection of Aphis fabae (Hem. Aphididae) with additional comments on life table research criteria. J Econ Entomol 108(4):1466–1478
Berkvens N, Bonte J, Berkvens D, Deforce K, Tirry L, De Clercq P (2008a) Pollen as an alternative food for Harmonia axyridis. BioControl 53:201–210
Berkvens N, Bonte J, Berkvens D, Tirry L, De Clercq P (2008b) Influence of diet and photoperiod on development and reproduction of European populations of Harmonia axyridis (Pallas) (Col. Coccinellidae). BioControl 53:211–221
Bonte M, Samih MA, De Clercq P (2010) Development and reproduction of Adalia bipunctata on factitious and artificial foods. BioControl 55:485–491
Carey JR (2001) Insect biodemography. Annu Rev Entomol 46:79–110
Chi H, Su HY (2006) Age-stage, two-sex life tables of Aphidius gifuensis (Ashmed) (Hym. Braconidae) and its host Myzus persicae (Sulzer) (Hom. Aphidaidae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ Entomol 35:10–21
Chi H (2013) TWOSEX-MSChart. In: Computer program for the age-stage, two-sex life table analysis. National Chung Hsing University. Available via DIALOG. http://140.120.197.173/Ecology/prod02.htm
De Clercq P (2002) Dark clouds and their silver linings: exotic generalist predators in augmentative biological control. Neotrop Entomol 31(2):169–176
De Clercq P, Wyckhuys K, De Oliveira HN, Klapwijk J (2002) Predation by Podisus maculiventris on different life stages of Nezara viridula. Fla Entomol 85:197–202
De Clercq P, Bonte M, Van Speybroeck K, Bolckmans K, Deforce K (2005) Development and reproduction of Adalia bipunctata (Col. Coccinellidae) on eggs of Ephestia kuehniella (Lep. Phycitidae) and pollen. Pest Manag Sci 61:1129–1132
Ebrahimi M, Sahragard A, Talaei-Hassanloui R, Kavousi A, Chi H (2013) The Life table and parasitism rate of Diadegma insulare (Hym. Ichneumonidae) reared on larvae of Plutella xylostella (Lep. Plutellidae), with special reference to the variable sex ratio of the offspring and comparison of jackknife and bootstrap techniques. Ann Ent Soc Am 106:279–287
Eliopoulos PA, Kontodimas DC, Stathas GJ (2010) Temperature-dependent development of Chilocorus bipustulatus (Col. Coccinellidae). Environ Entomol 39:1352–1358
Evans EW (2009) Ladybeetles as predators of insects other than Hemiptera. Biol Control 51:255–267
Farhadi R, Allahyari H, Chi H (2011) Life table and predation capacity of Hippodamia variegate (Col. Coccinellidae) feeding on Aphis fabae (Hem. Aphididae). Biol Control 59:83–89
Gabre RM, Adham FK, Chi H (2005) Life table of Chrysomya megacephala (Fabricius) (Dipt. Calliphoridae). Acta Oecol 27(3):179–183
Gaillot P (1967) Contribution à la lutte biologique contre Parlatoria blanchardi. L’entomologiste 23:5–6
Hamasaki K, Matsui M (2006) Development and reproduction of an aphidophagous coccinellid, Propylea japonica (Thunberg) (Col. Coccinellidae), reared on an alternative diet, Ephestia kuehniella Zeller (Lep. Pyralidae) eggs. Appl Entomol Zool 41:233–237
Hattingh V, Samways MJ (1993) Evaluation of artificial diets and two species of natural prey as laboratory food for Chilocorus spp. Entomol Exp Appl 69:13–20
Hodek I, Honek A (1996) Ecology of coccinellidae. Kluwer Academic Publishers, Dordrecht
Hodek I, Honek A (2009) Scale insects, mealy bugs, whiteflies and psyllids (Hemiptera, Sternorrhyncha) as prey of ladybirds. Biol Control 51:232–243
Jalali MA, Tirry L, Arbab A, De Clercq P (2010) Temperature-dependent development of the two-spotted ladybeetle Adalia bipunctata on the green peach aphid Myzus persicae and a factitious food under constant temperatures. J Insect Sci 10:1536–1552
Laudeho Y, De Janvry EC, Iperti C, Brun J (1970) Intervention bio-écologique contre la cochenille blanche du palmier-dattier Parlatoria blanchardi (Coccoidea: Diaspididae) en Adrar Mauritanien. Fruits 25:147–160
Lucas E, Gagne I, Coderre D (2002) Impact of the arrival of Harmonia axyridis on adults of Coccinella septempunctata and Coleomegilla maculata (Coleoptera: Coccinellidae). Eur J Entomol 99:457–463
Maes S, Gregoire JC, De Clercq P (2014) Prey range of the predatory ladybird Cryptolaemus montrouzieri. BioControl 59(6):729–738
Mahdian K, Kerckhove J, Tirry L, De Clercq P (2005) Effects of diet on development and reproduction of the predatory pentatomids Picromerus bidens and Podisus maculiventris. BioControl 51:725–739
Matthews RW, Matthews JR (1978) Insect behavior. Wiley, New York
Mehrnejad MR, Copland MJW (2006) Behavioral responses of the parasitoid Psyllaephagus pistaciae (Hym. Encyrtidae) to host plant volatiles and honeydew. Entomol Sci 9:31–37
Michaud JP (2005) On the assessment of prey suitability in aphidophagous Coccinellidae. Eur J Entomol 102:385–390
Miller DR, Gimpel ME (2000) A systemic catalogue of the Eriococcidae (felt scales) (Hem. Coccidae) of the world. Intercept Ltd, Andover
Moghadam M, Tvakoli M (2010) Scale insects of the central Zagros region in Iran (Hem. Coccidae). Appl Entomol Phytopathol 77(2):25–46
Nadel DJ, Biron S (1964) Laboratory studies and controlled mass rearing of Chilocorus bipustulatus (Col. Coccinellidae) a citrus scale predator in Israel. Rev de Parasit 3:162–206
Panahi B, Basirat M, Hosseinifard SJ (2013) The effect of liquid detergent on common pistachio psylla, Agonoscena Pistaciae, soil and plant. Int J Agric Crop Sci 6(12):794–807
Rosen D, Gerson U (1965) Field studies of Chilocorus bipustulatus on citrus in Israel. Ann de Epiphyt 16:71–76
Specty O, Febvay G, Grenier S, Delobel B, Piotte C, Pageaux J, Ferran A, Guillaud J (2003) Nutritional plasticity of the predatory lady beetle Harmonia axyridis (Col. Coccinellidae) comparison between natural and substitution prey. Arch Insect Biochem 52:81–91
Stark JD, Sugayama RL, Kovaleski A (2007) Why demographic and modeling approaches should be adopted for estimating the effects of pesticides on biocontrol agents. Biol Control 52:365–374
Stathas GJ, Eliopoulos PA, Bouras SL, Economou LP, Kontodimas DC (2003) The scale Parthenolecanium persicae (Fabricius) on grapes in Greece. Bull OILB/SROP 26:253–257
Stathas GJ, Eliopoulos PA, Japoshvili G, Kontodimas DC (2009) Phenological and ecological aspects of Protopulvinaria pyriformis (Cockerell) (Hem. Coccidae) in Greece. J Pest Sci 82:33–39
Uygun N, Elekcioglu NZ (1998) Effect of three Diaspididae prey species on development and fecundity of the ladybeetle Chilocorus bipustulatus in the laboratory. BioControl 43:153–162
van Lenteren J, Babendreier D, Bigler F, Burgio G, Hokkanen HMT, Kuske S, Loomans AJM, Menzler-Hokkanen I, Van Rijn PCJ, Thomas MB, Tommasini MG, Zeng QQ (2003) Environmental risk assessment of exotic natural enemies used in inundative biological control. BioControl 48:3–38
Villanueva R, Michaud JP, Childers CC (2004) Ladybeetles as predators of pest and predacious mites in citrus. J Entomol Sci 39:23–29
Yigit A, Canhilal R, Ekmekci U (2003) Seasonal population fluctuations of Serangium parcesetosum (Col. Coccinellidae), a predator of citrus whitefly, Dialeurodes citri (Hom. Aleyrodidae) in Turkey’s Eastern Mediterranean citrus groves. Environ Entomol 32(5):1105–1114
Acknowledgments
The authors are grateful to the Vali-e-Asr University of Rafsanjan for financial support to H. Atrchian.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Marta Montserrat.
Rights and permissions
About this article

Cite this article
Atrchian, H., Mahdian, K. & Shahidi, S. Effect of different natural and factitious diets on development and life history parameters of Chilocorus bipustulatus . BioControl 61, 703–715 (2016). https://doi.org/10.1007/s10526-016-9754-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10526-016-9754-z