
Abstract
Researchers have long theorized that genetic influence on mental health may differ as a function of environmental risk factors. One likely moderator of genetic and environmental influences on psychopathological symptoms is parenting behavior, as phenotypic research shows that negative aspects of parent–child relationships are associated with greater likelihood of mental illness in adulthood. The current study examined whether levels of reported parental discipline and affection experienced in childhood act as a trigger, or buffer, for adult mental health problems. Results from a nationwide twin sample suggest level of father’s discipline and affection, as reported by now-adult twins, moderated genetic and environmental influences on internalizing symptoms in adulthood, such that heritability was greatest at the highest levels of discipline and affection. Father’s affection also moderated the etiological influences on alcohol use problems, with greater heritability at the lowest levels of affection. No moderating effect was found for mothers. Findings suggest relationships with fathers in childhood can have long-lasting effects on the etiological influences on adult mental health outcomes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
All common forms of psychopathology have a significant heritable component (Plomin et al. 2012), and certain risk or protective factors may moderate genetic influences on mental health outcomes (e.g., Cadoret et al. 1995). For more than 10 years, researchers have utilized twin data to examine whether heritability estimates for different types of psychopathology vary as a function of “moderator” variables (Purcell 2002). Consistent with this idea, socioeconomic status (SES) moderates genetic influences on antisocial behavior (Tuvblad et al. 2006) and internalizing psychopathology (South and Krueger 2011). In both cases, greater genetic influences are found at the highest ends of SES, evidence of what Bronfenbrenner and Ceci (1994) referred to as a bioecological model or what others (Raine 2002) have called a social push theory, suggesting that heritability is greatest in the most enriched environment and lowest in the most disadvantaged environment. In other work, researchers have found that genetic influences are highest in the most distressed environments (South and Krueger 2008), evidence of a diathesis-stress model in which latent risk for psychopathology is triggered by stressful events (Monroe and Simons 1991). Finally, it is also possible for biometric moderation models to show that genetic influences are elevated at the extreme levels of the moderator (South and Krueger 2013), evidence of differential susceptibility (Belsky and Pluess 2009) or the “orchid” model (Ellis and Boyce 2008), the idea being that some individuals, like dandelions, will flourish anywhere, while some, like an orchid, need certain conditions to thrive. The quantitative models that test gene X environment interaction have the advantage of also modeling changes in environmental influences on psychopathology; going back to the example of SES and mental health, the greatest influence of the unique environment on internalizing psychopathology was at the extreme low end of SES (South and Kruger 2011).
One likely moderator of genetic and environmental influences on symptoms of psychopathology is the parent–child relationship, because parenting styles can place children at risk for psychopathology. In particular, parenting that is rejecting or overcontrolling has been linked to later mental illness (e.g., Parker et al. 1995). Retrospective reports of authoritarian parenting have been linked to later well-being and depressive symptoms but not substance abuse (Rothrauff et al. 2009). In addition to having phenotypic, correlational relationships with psychopathology, this type of parenting may work on a different level, moderating the etiological influences on mental illness.
A handful of studies have examined parent–child relationships as a moderator of the genetic and environmental influences on mental health outcomes. For example, Hicks et al. (2009b) reported that genetic variance of externalizing psychopathology was greater for adolescents who reported the most problems in relationships with their parents. This would support a diathesis-stress model of externalizing problems, with environmental adversity triggering expression of genetic influences. However, the findings across studies are inconsistent, possibly because researchers have not distinguished, first, the impact of relationships with mothers and fathers (e.g., Feinberg et al. 2007), and second, the two different dimensions of discipline and affection (e.g., Hicks et al. 2009a, b).
Even when separate studies use putatively similar parenting variables, results can be conflicting. In two separate studies, maternal discipline had opposite effects on internalizing and externalizing behavior. One study found the greatest genetic influences on internalizing problems (i.e., depressive symptoms) at the highest levels of punitive discipline (Lau and Eley 2008). This would also support a diathesis-stress model of psychopathology if the high punitive discipline environment is interpreted as the more “at-risk” environment. Another study using the same sample, however, found greater influence of genetics in the low maternal punitive discipline environment for externalizing behavior (Button et al. 2008), supportive of the social-push theory of psychopathology. Still other work has reported that maternal and paternal discipline had opposite effects on the genetic variance in externalizing behavior (Button et al. 2008), with greater genetic variance found at low levels of maternal discipline and high levels of paternal discipline.
The conflicting findings in the literature to date may result from a variety of method-related factors, including different ages of twin participants, assessment of psychopathology, and even the type of moderation model used. Indeed, recent work on biometric moderation suggests that failure to consider nonlinear effects may result in spurious evidence of GxE (Van Hulle et al. 2013). It is possible, however, that parenting may be a significant biometric moderator of psychopathology, and the mixed findings from studies conducted so far may reflect that parent–child relationships moderate genetic influence at the level of specific combinations of parent (mother, father), dimension of parental behavior (affection, discipline), and psychopathology (internalizing symptoms, alcohol use problems). These levels have not been examined in previous work. This is a key limitation that prevents us from understanding when and under what circumstances the parent–child relationship can serve as a trigger or a buffer for the expression of genetic and environmental influences on psychopathology.
The aim of the current research was to examine components of the parent–child relationship that moderate genetic and environmental influence on adult symptoms of psychopathology. Previous research has exclusively examined childhood and adolescent samples, leaving unanswered the question of whether parenting can have a lasting impact on the etiology of mental health into adulthood. The current study used an exploratory approach to try and untangle mixed findings from previous work. Retrospective reports of parental discipline and affection were obtained, separately for mothers and fathers, from monozygotic (MZ) and dizygotic (DZ) twins. Each combination of parent and parental dimension (e.g., mother’s discipline) was examined as a potential moderator for the genetic and environmental influences on an internalizing factor score and a sum score of alcohol use problems.
Given the conflicting results in the literature, and the exploratory nature of the current study, predictions about the direction of effects were not hypothesized but it was expected that moderation would be found for some, but not all, combinations of mental health problems, parent, and dimension of parenting. If genetic influences on symptoms of psychopathology were greatest in the most “enriched” environment (i.e., one marked by high affection and low levels of strict discipline) a social push theory of psychopathology would be supported, whereas if genetic influences were greatest in the most “risky” environment (i.e., with lowest levels of affection and strictest discipline) it would be evidence in favor of a diathesis-stress model. It is also possible that genetic influences might be elevated at extreme ends of the moderator variable, evidence of a differential susceptibility/orchid model. If differential susceptibility was found for discipline, for instance, it might suggest that either too little or too much discipline leads to a greater expression of genetic influences.
Method
Sample
Participants were drawn from a nationwide twin sample recruited for the MacArthur Foundation Survey of Midlife Development in the United States (MIDUS). The larger MIDUS sample is a nationally representative cohort of individuals aged 25–74 years drawn from the non-institutionalized civilian population of the continental United States. The MIDUS study twin subsample (Kessler et al. 2004) was ascertained through a telephone survey of approximately 50,000 households (Kessler et al. 2004). The final response rate (i.e., both members of the twin pair were contacted by an interviewer, agreed to participate, and completed a short zygosity screening questionnaire) was 26 %. The twin subsample completed the same MIDUS battery as the full sample, consisting of a computer-assisted telephone interview and two mailed questionnaire booklets (Kendler et al. 2000).
A total of 1996 people were recruited for and enrolled in the MIDUS twin sample. Twins completed a brief twin screen to determine zygosity, a technique that is generally more than 90 % accurate (cf. Lykken et al. 1990). Twins with missing or indeterminate zygosity information, twins who only completed the twin screener and no other measures, and opposite-sex twins were eliminated. This left a total of 1386 twins. The biometric models used here require data from both twins on the moderator variable; some twins were missing all data for a paired co-twin, leaving a total of 1344 twins from 672 twin pairs used in the biometric models: 164 monozygotic (MZ) male pairs, 186 MZ female pairs, 124 dizygotic (DZ) male pairs, and 198 DZ female pairs. The average age of the sample of complete twin pairs was 45 (SD = 12.17, range = 25–74).
Measures
Parent–child relationship quality The MIDUS questionnaire obtained retrospective reports of parental affection and discipline received during childhood. Seven items each retrospectively assessed maternal and paternal affection. One of the items (How would you rate your relationship with your mother/father during the years you were growing up?) was rated on a 5-point scale (1 = Excellent, 5 = Poor). The other six items (i.e., How much did s/he understand your problems and worries?, How much could you confide in him/her about things that were bothering you?, How much love and affection did s/he give you?, How much time and attention did s/he give you when you needed it?, How much effort did s/he put into watching over you and making sure you had a good upbringing?, How much did s/he teach you about life?) were rated on a 4-point scale (1 = A lot, 4 = Not at all). A total scale score was calculated by taking the mean of the 7 items (the first item was multiplied by .75 to maintain continuity with the other six variables). Four items were included to assess maternal and paternal discipline by examining how strict, consistent, and harsh parents were during childhood (Rossi 2001). These items (i.e., How strict was s/he with her rules for you?, How consistent was s/he about the rules?, How harsh was s/he when s/he punished you?, How much did s/he stop you from doing things that other kids your age were allowed to do?) were rated on a 4-point scale (1 = A lot, 4 = Not at all), and total scores were calculated as means of the 4 items.
There was missing data for reports of maternal and paternal discipline/affection (maternal discipline = 7.5 % missing, maternal affection = 7.4 % missing, paternal discipline = 12.4 % missing, paternal affection = 12.0 % missing). Because the biometric moderation model used in the current analyses requires that both twins have data on the moderator variable, mean substitution techniques were used to account for the missing data and maintain the sample size. Higher scores indicate more discipline and more affection, respectively. The average score for maternal discipline was 2.95 (SD = .61, range = 1–4, alpha = .76). The mean for maternal affection was 3.17 (SD = .66, range = .96–3.96, alpha = .91). Paternal discipline and affection had average scores of 2.98 (SD = .71, range = 1–4, alpha = .84) and 2.73 (SD = .74, range = .75–3.96, alpha = .92), respectively.Footnote 1
Internalizing symptoms Internalizing scores were taken from three DSM-III-R disorders, major depressive episode (MD), generalized anxiety disorder (GAD), and panic disorder (PD), as well as a rating of neuroticism. The 12-month prevalence for symptoms of the three disorders were measured during the phone interview using Composite International Diagnostic Interview Short Form scales (CIDI-SF; Kessler et al. 1998b). Research demonstrates good total classification accuracy (percentage of respondents whose CIDI-SF classification is the same as their classification of the full CIDI; Kessler et al. 1998a) and agreement with clinical diagnoses (Kessler et al. 1998b; Wittchen 1994). Neuroticism was assessed on the MIDUS self-administered questionnaire, using a personality measure based on the Five Factor Model (FFM; Lachman and Weaver 1997). The Neuroticism scale includes four adjectives rated on a 1–4 scale (1 = A lot, 4 = Not at all): ‘moody,’ ‘worrying,’ ‘nervous,’ and ‘calm’ (reverse scored). Items were averaged to create a total neuroticism score.
As detailed elsewhere (see South and Krueger 2011), symptom counts for GAD, MD, and PD, and the scale score for neuroticism, were entered into a one-factor confirmatory factor analysis conducted in Mplus (Muthén and Muthén 1998–2012). Factor scores were extracted for use in the biometric moderation analyses and reversed to be positively correlated with the parenting variables (e.g., higher parental affection was positively correlated with less internalizing).
Alcohol use problems Alcohol use problems were assessed for the previous 12 months using the Alcohol Screening Test (AST; Selzer 1971) and two other questions (i.e., “using larger amounts longer than intended,” and “suffering the effects or after-effects of alcohol at home or work”). The AST consists of five dichotomous (Yes/No) items asking if alcohol use heightened chances of getting hurt, if emotional or psychological problems resulted from alcohol use, if there was a strong desire or urge to use alcohol, if a great deal of time was spent using or recovering from the effects of alcohol, and if more alcohol than usual had to be used to get the same effect. The additional alcohol use questions were rated on a 6-point scale (1 = never, 2 = once or twice, 3 = 3 to 5 times, 4 = 6 to 10 times, 5 = 11 to 20 times, 6 = more than 20 times) but dichotomized (Never/Occurred) and summed with the other items (see Jarnecke and South 2014). Raw scores for alcohol use problems ranged from 0 to 7 and participants were required to respond to at least five of the seven items examined for analysis. Higher scores reflected more alcohol use problems. The average score for alcohol problems was .50 (SD = 1.08, range = 0–7, alpha = .73). Scores were inversely transformed (and thus reversed) for use in the biometric analyses to adjust for negative skew.
Data analysis
The current analyses utilized biometric models designed for twin data. In general, biometric modeling with twin data uses the differences in MZ twins, who share 100 % of their genes, and DZ twins, who share 50 % of their independently segregating genes, to decompose the variance in a phenotype, like mental illness, into the variance due to genetic effects (A), common environmental influences (C), and unique environmental influences (E).
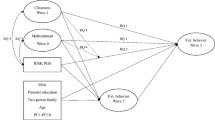
To determine the degree to which the ACE estimates on internalizing symptoms and alcohol use problems were shared with those contributing to parenting behavior, bivariate (Cholesky) decompositions were tested first. This bivariate decomposition includes latent factors representing the additive genetic, shared environmental, and nonshared environmental variance in the phenotype of interest (here, internalizing or alcohol problems) as well as latent factors representing the variance shared between the parenting behavior variable and the outcome psychopathology variable. To allow for the possibility that ACE estimates of internalizing symptoms and alcohol use problems vary as a function of parental behavior (abbreviated here for simplicity as GxM), moderation terms were included in the second set of models (Van Hulle et al. 2013).Footnote 2 For each of the paths influencing the outcome variable, this bivariate Cholesky with GxM includes an overall coefficient separate from the moderator variable (i.e., parenting behavior) that indicates the magnitude of each effect of A, C, or E on the outcome, and the product of a coefficient that indexes the moderation of the outcome by parental behavior multiplied by the level of the moderator (see Fig. 1a). Parameter estimates from the model are then used to plot the model-predicted genetic and environmental components of variance for psychopathological symptoms at different levels of parental behavior.

a Bivariate Choleksy with GxM model with the parent–child relationship variable moderating genetic and environmental effects on the mental health variable (model displayed for single member of a twin pair). A additive genetic effects, C shared environmental variance, E nonshared environmental influences. aM, cM, and eM represent genetic, shared environmental, and nonshared environmental parameter estimates, respectively, for the moderator variable. ac, cc, and ec signify variance shared between the moderator variable and the outcome variable. au, cu, and eu represent residual variance unique to the outcome variable. b Nonlinear main effects with GxM model with the parent–child relationship variable moderating genetic and environmental effects on the mental health variable (displayed for one member of a twin pair). A additive genetic effects, C shared environmental variance, E nonshared environmental influences. aM, cM, and eM represent genetic, shared environmental, and nonshared environmental estimates for the moderator variable. au, cu, and eu denote residual variance unique to the outcome variable. αu, κu, and εu denote interactions between the moderator (M) and the genetic and environmental influences on the outcome variable. β1 and β2 represent the linear and non-linear main effects of the moderator variable on the outcome variable
Because the biometric moderation model described above may be prone to false positives (van der Sluis et al. 2012), several alternative models were tested as well. The first of these alternative models was a nonlinear main effects model with GxM (Van Hulle et al. 2013; see Fig. 1b). This model removes moderation paths common to the moderator and outcome variables but retains the unique moderation paths, while including the linear and nonlinear main effects (β1 and β2) of the moderator on the outcome. This allows for the linear and nonlinear effect of the moderator on the phenotype to be modeled directly rather than through common moderation paths. Next, a more restrictive, nonlinear main effects model was fit, which constrained the unique moderation paths to zero. Finally, a linear main effects model was tested that examined the direct effect of the moderator on the outcome variable, allowing for the ACE decomposition of both the moderator and outcome variables.
Parenting behavior and psychopathology (i.e., internalizing symptoms and alcohol use problems) variables were regressed on age, age2, age × gender, and age2 × gender (McGue and Bouchard 1984) to correct for potential biases in model fitting due to gender and age differences. The residuals were then used to fit models in Mplus (Muthén and Muthén 1998-2012) using full-information maximum-likelihood to account for missing data in the outcome variable. Model fit was evaluated using the Akaike Information Criterion (AIC; Akaike 1987), Bayesian Information Criterion (BIC), and the likelihood ratio test (LRT). The difference in the −2 log-likelihood values for two separate models, the LRT is distributed as Chi square; a statistically significant difference in LRT between two models can indicate improvement in the model’s fit as a result of adding or removing parameters. The AIC and BIC are information theoretic fit statistics that balances fit and number of parameters when choosing the best-fitting model, with lower values indicating better fitting models (Markon and Krueger 2004).
Results
Parenting behavior and internalizing symptoms
The raw internalizing factor score (not reversed) was significantly correlated with raw scores for maternal affection (r = −.18, p < .001) but not maternal discipline (r = .00, p = 1.00). The internalizing score was correlated with paternal affection (r = −.15, p < .001) but not paternal discipline (r = .03, p = .25). The five models described above (and listed in Table 1) were fit to the data for each combination of parenting variable and internalizing: bivariate Cholesky (BivCholesky), bivariate Cholesky with GxM (BivCholesky GxM), nonlinear main effects model with GxM (NL Main GxM), nonlinear main effects model (NL Main), and linear main effects model (Linear Main).
When maternal discipline and internalizing were entered into the models, the NL Main model provided the best fit to data according to AIC (7397) and BIC (7442; see Table 1). The NL Main model resulted in a nonsignificant loss of fit from the NL Main GxM model (\(\chi^{ 2}_{\text{diff}}\) = 2.85, 3 df, p = .42), but moving to the Linear Main model significantly worsened model fit (\(\chi^{ 2}_{\text{diff}}\) = 124.27, 1 df, p = .00). When the models were fit to the maternal affection and internalizing variables, the NL Main model again provided the best fit according to LRT and BIC (\(\chi^{ 2}_{\text{diff}}\) = 5.04, 3 df, p = .17; BIC = 7299). The AIC was lowest for the BivCholesky GxM (7252) but close to the NL Main model (7254). These results indicated that the genetic and environmental components of internalizing symptoms did not vary by level of perceived maternal discipline and affection, after accounting for the linear and nonlinear effects of maternal variables on internalizing.
Because there was no support for moderation, the ACE estimates for these models were interpreted at the average levels of maternal discipline and affection, respectively (see Table 2). For the maternal discipline NL Main model, genetic influences accounted for 21 % of the variance in internalizing, shared environmental influences accounted for 14 %, and nonshared environmental influences accounted for 64 % of the variance. Forty-four percent of the variance in maternal discipline was due to genetic effects, 11 % was due to shared environmental effects, and 45 % was due to nonshared environmental effects. Estimates for internalizing from the maternal affection model suggested that 14, 18, and 64 % of the variance in internalizing was accounted for by genetic, shared environmental, and nonshared environmental influences, respectively. Forty-four, 23, and 34 % of the variance in maternal affection was attributed to genetic, shared environmental, and nonshared environmental influences, respectively.
Next, the models were fit to paternal variables and internalizing. For paternal discipline and internalizing, the NL Main GxM provided the best fit to data according to AIC (7360) and BIC (7418). Moving from the NL Main GxM model to the NL Main model also resulted in a significant decrement in fit (\(\chi^{ 2}_{\text{diff}}\) = 15.17, 3 df, p = .00). This suggests that paternal discipline moderated the genetic and environmental influences unique to internalizing symptoms. When the parameter estimates from the NL Main GxM model were plotted, heritability estimates increased from low (a2 = 2 %) to high (a2 = 48 %) levels of paternal discipline (see Table 2; Fig. 2a). Influences from the shared environment decreased from 53 % at low levels of discipline to 1 % at +1 SD of discipline before increasing to 8 % at high levels (+2SD) of discipline. Finally, the proportion of variance from the unique environment was curvilinear, increasing from low (e2 = 45 %) to high (e2 = 66 %) levels of paternal discipline before decreasing at higher levels of paternal discipline (e2 = 44 %). Confidence intervals around the moderation parameter estimates suggest that much of the moderation seems to be occurring on the shared environmental path (see Table 3). This model also provided ACE estimates for paternal discipline. The model showed that 19 % of the variance in paternal discipline was explained by genetic influences, 31 % by shared environmental influences, and 48 % by nonshared environmental influences.

a Proportion of variance in internalizing as a function of paternal discipline. b Proportion of variance in internalizing as a function of paternal affection. c Proportion of variance in alcohol use problems as a function of paternal affection. A additive genetic, C shared environment, E nonshared environment
For paternal affection and internalizing, the NL Main GxM model provided the best fit to the data according to AIC (7215). However, moving to the NL Main model resulted in a nonsigificant loss of fit (\(\chi^{ 2}_{\text{diff}}\) = 5.45, 3 df, p = .14). BIC was better for the NL Main model (7265) than the NL Main GxM model (7274) but by less than the 10 points commonly accepted as “very strong evidence” in favor of the model with the more negative value (Raftery 1995). Given that recent simulation studies with the models tested showed that differences in BIC values from −10 to 10 suggest that the alternative models fit “equally well” (Van Hulle et al. 2013), we chose to interpret the NL Main GxM model; however, the results from this model were interpreted with caution. As presented in Table 2 and Fig. 2b, the heritability of internalizing increased from 9 % at the lowest level of affection to 32 % at the highest level of affection. Shared environmental influences decreased from low (c2 = 34 %) to high (c2 = 0 %) levels of paternal affection. Nonshared environmental effects were weaker for low (e2 = 57 %) levels of paternal affection, but increased at high levels of affection (e2 = 68 %). This model also provide estimates for paternal affection: 36 % of the variance was due to genetic influences and 32 and 31 % of the variance was due to shared and nonshared environmental influences, respectively.
Parenting behavior and alcohol use problems
The raw alcohol use problems score was not significantly correlated with raw scores for maternal discipline (r = −.04, p = .20), maternal affection (r = .00, p = .98), or paternal discipline (r = .00, p = 1.00). It was significantly correlated with paternal affection, though the correlation was small (r = −.07, p = .01).
For maternal discipline and maternal affection, the NL Main model provided the best fit to data according to AIC, BIC, and LRT (see Table 1). ACE estimates for these models were interpreted (see Table 2). For the NL Main model with maternal discipline, 38 % of the variance in alcohol problems was accounted for by genetic influences, 0 % of the variance was accounted for by shared environmental influences, and 62 % of the variance was accounted for by nonshared environmental influences. The variance estimates for maternal discipline were identical to those found for the model with internalizing: 44 % genetic, 11 % shared environment, and 45 % nonshared environment. For the NL Main model with maternal affection, genetic influences accounted for 37 % of the variance in alcohol use problems and the remainder of the variance (63 %) was accounted for by nonshared environmental influences; variance components for maternal affection were almost identical to those from the model with internalizing (43 % genetic, 23 % shared environment, 34 % nonshared environment).
For paternal discipline and alcohol use, again, the NL Main model best fit the data. This model resulted in the lowest AIC (7110) and BIC (7155) and resulted in a nonsignificant loss of fit from the NL Main GxM model (\(\chi^{ 2}_{\text{diff}}\) = 0.47, 3 df, p = .92). The ACE estimates for the NL Main model indicate that 38 % of the variance in alcohol problems is accounted for by genetic effects and the remainder (62 %) of the variance is accounted for by nonshared environmental effects. Variance estimates for paternal discipline were identical to those found in the model with internalizing: 19 % genetic, 31 % shared environment, 48 % nonshared environment.
Finally, for paternal affection and alcohol problems, there was evidence suggesting that the NL Main GxM provided the best fit to data. It resulted in the lowest AIC (6950) and had a BIC value (7008) that was less than 10 points from the lowest BIC value (7000 for NL Main), although going from the NL Main GxM to the NL Main model resulted in a nonsignificant loss of fit (\(\chi^{ 2}_{\text{diff}}\) = 5.70, 3 df, p = .13). Therefore, as with paternal affection and internalizing, we chose to interpret the NL Main GxM model, but with caution. Genetic influences on alcohol problems decreased from low (a2 = 40 %) to high (a2 = 32 %) levels of paternal affection (see Table 2; Fig. 2c). Shared environmental influences were negligible across level of paternal affection. Unique environmental influences increased slightly from low (e2 = 60 %) to high (e2 = 68 %) levels of paternal affection. The estimates of genetic and environmental influences on paternal affection were identical to those found in the model with internalizing: 36 % genetic, 32 % shared environment, and 31 % nonshared environment.
Discussion
Findings from a growing body of research have empirically established the presence of GxE for major domains of psychopathology, including internalizing symptoms and alcohol use problems. One likely moderator of adult mental health that has not been fully explored to this point is the parent–child relationship. Parents who use strict discipline or are lacking in affection may put their genetically predisposed children at risk for mental health problems in adulthood. In the current study, we examined retrospective reports of parenting behavior—discipline and affection measured separately for mothers and fathers—as moderators of the genetic and environmental influences on internalizing symptoms and alcohol use problems in a nationally representative sample of adult twins. As has been found previously (e.g., Rowe 1981), our results demonstrated that aspects of the parent–child relationship were moderately heritable. Our main findings indicated that father’s parenting behavior moderated the etiology of both internalizing symptoms and alcohol use problems. The genetic and environmental influences for internalizing symptoms and alcohol use problems differed depending on the level of affection received from a father during childhood, while genetic and environmental variance on internalizing symptoms also varied as a function of paternal discipline.
When paternal discipline was the moderator of the etiological influences on internalizing, there was a substantial increase in heritability (and genetic variance) from low to high levels of discipline. In line with other similar findings (Button et al. 2008; Lau and Eley 2008), our results support a diathesis-stress model, with greater genetic effects expressed in the “risky” environment—one where there is strict paternal discipline. To our knowledge, this is the first study to find evidence of paternal discipline moderating the genetic and environmental influences on internalizing psychopathology. Like extreme levels of maternal discipline (Lau and Eley 2008), paternal discipline may also have an effect on the etiology of internalizing symptoms by allowing for the expression of genetic influences.
When paternal affection was the moderator of genetic and environmental influences on internalizing, the genetic variance (and heritability) increased from low to high levels of affection. Given the mixed evidence in support of biometric moderation, it is recommended that these results be interpreted with caution. Indeed, a similar study failed to find a moderating effect of a summary parental affection score on the etiological influences on depressive symptoms (Feinberg et al. 2007). If replicated, the current findings may be suggestive of a social push model, with greater genetic influences in the most “enriched” environment (i.e., one marked by the highest levels of affection). Individuals who are immersed in an advantageous environment, such as an environment with high paternal affection, may exhibit internalizing symptoms despite a warm and caring father-child relationship because they are unable to counteract a genetic predisposition.
Of note, the proportion of variance in internalizing due to shared environmental influences was highest in the least affectionate and least disciplined environments. Any non-zero estimates of the shared environment on adult symptoms of psychopathology are notoriously difficult to find; thus, the fact that the proportion of variance in internalizing due to the shared environment was estimated at 53 % (from a model with paternal discipline) and at 34 % (from a model with paternal affection) at extremely low levels of the moderator is important. These substantial estimates can be interpreted as meaning that the type of father parenting experienced by both twins had a moderate to strong impact on whether each sibling later reported internalizing symptoms. This suggests that lack of father engagement per se, even when that engagement is controlling and strict, has an important role in the development of mental illness.
Paternal affection also significantly moderated the etiological influences on alcohol use problems. The drop in genetic variance from low to high levels of affection was paralleled by a concurrent drop in nonshared environmental variance, such that there was greater overall variance in alcohol use problems when paternal affection was lower. Our findings provide evidence for a diathesis-stress model of etiology, wherein genetic influences emerge in an environment where the father is less warm and loving. This is consistent with a majority of studies investigating biometric moderation of alcohol use problems and related phenotypes, which tend to find support for a diathesis-stress model of etiology (Young-Wolff et al. 2011).
No moderation of internalizing or alcohol use problems as a function of maternal affection or maternal discipline was found. Hicks et al. (2009a) reported moderation of the ACE components of internalizing as a function of mother–child relationship problems, but only for the nonshared environmental parameter. Differences could be due to several factors, including different measures of the parent–child relationship, different mean ages of the samples when assessed for psychopathological symptoms, or the cross-sectional nature of that study compared to the retrospective measurement of parent–child relationship utilized in the current study.
Even though this study failed to find moderation of the etiological influences on internalizing and alcohol use problems as a function of the mother–child relationship, this does not suggest that only the paternal relationship is important in the development of mental health problems. Our findings will certainly need to be replicated in other samples. The mother–child relationship may contribute to later psychopathological symptoms but may not have (or maintain) a moderating effect on the etiological influences in psychopathology when one is well into adulthood. Our findings are, however, consistent with research supporting the importance of the father in the home for predicting later offspring psychopathology (Blazei et al. 2008; Harold et al. 2012).
This study is not without limitations. Foremost, our measure of the parent–child relationship was ascertained during adulthood in our sample, which ranged in age from 25 to 74 at time of data collection. Thus, symptoms of adult mental illness may have colored participants’ recollection of the parent–child relationship and this bias may have been more similar between MZ twin pairs, compared to DZ pairs. If this is the case, estimates for genetic variance may be inflated and estimates of the nonshared environment may actually contain a large degree of measurement error. It is also possible that individuals from different generations might have had different interpretations of what “discipline” and “affection” entailed in regards to their relationship with their parents. Although there are limitations surrounding use of this measure, evidence suggests that adult retrospective reports of parenting behavior are consistent with childhood assessments (Brewin et al. 1993; Wilhelm et al. 2005), and retrospective reports of parenting predicts well-being across the life-course (Russek and Schwartz 1997). Nevertheless, future studies should endeavor to replicate these findings using prospective studies with more homogenous and recently studied samples. Second, our measures of mental health problems were all self-report and different results might have been obtained using clinician- or informant-report measures. Third, the size of the sample used in the current analyses was somewhat small compared to other twin studies that use biometric moderation models. Despite this relatively small sample size there was still enough power to find moderation of paternal relationship quality on the etiological influences on adult mental health. It is unknown if maternal relationship quality would have acted as a moderator if there was a larger sample size, however.
In summary, our results reveal the importance of the father-child relationship for understanding the etiology of symptoms of psychopathology later in life. Following a diathesis-stress model of psychopathology, greater genetic influences on internalizing were found at extremely high levels of paternal discipline. A diathesis-stress model was also supported for the effect of paternal affection on alcohol use problems. The effect of paternal affection on the etiological components of internalizing, however, was best explained by a social push model. Our findings continue to add to what is known about the gene-environment interplay between aspects of the parent–child relationship and later psychopathology. Affection and discipline, however, while important constructs, are only two dimensions of parenting; future work is needed to determine whether and in what way other aspects of the parent–child relationship may impact the etiology of the child’s future mental health.
Notes
Means were taken from the sample of 1386 participants, who had known zygosity, information from multiple measures, and a same-sex co-twin. These means were used in the mean substitution procedures. Analyses were also conducted using means from the sample of 1344 twins, where each twin had data for a co-twin for the mean substitution procedures. The latter did not influence the findings of the study.
Biometric moderation was also tested using alternative modeling procedures (see Purcell 2002) in Mx. Results were largely consistent with what is reported here, in terms of model fit and variance components. Full results are available from the first author. Note that the GxM model tested here is equivalent to the bivariate biometric moderation model described by Purcell (2002).
References
Akaike H (1987) Factor analysis and AIC. Psychometrika 52:317–332
Belsky J, Pluess M (2009) Beyond diathesis stress: differential susceptibility to environmental influences. Psychol Bull 135:885–908
Blazei RW, Iacono WG, McGues M (2008) Father-child transmission of antisocial behavior: the moderatingr role of father’s presence in the home. J Am Acad Child Adolesc Psychiatry 47(4):406–415
Brewin CR, Andrews B, Gotlib IH (1993) Psychopathology and early experience: a reappraisal of retrospective reports. Psychol Bull 113(1):82–98
Bronfenbrenner U, Ceci SJ (1994) Nature-nurture reconceptualized in developmental perspective: a bioecological model. Psychol Rev 101:568–586
Button TM, Lau JYF, Maughan B, Eley TC (2008) Parental punitive discipline, negative life events and gene-environment interplay in the development of externalizing behavior. Psychol Med 38:29–39
Cadoret RJ, Yates W, Troughton E, Woodworth G, Stewart MA (1995) Genetic-environmental interaction in the genesis of aggressivity and conduct disorders. Arch Gen Psychiatry 52:916–924
Ellis BJ, Boyce WT (2008) Biological sensitivity to context. Curr Dir Psychol Sci 17:183–187
Feinberg ME, Button TMM, Neiderhiser JM, Reiss D, Hetherington EM (2007) Parenting and adolescent antisocial behavior and depression: evidence of genotype x parenting environment interaction. Arch Gen Psychiatry 64(4):457–465
Harold GT, Elam KK, Lewis G, Rice F, Thapar A (2012) Interparental conflict, parent psychopathology, hostile parenting, and child antisocial behavior: examining the role of maternal versus paternal influences using a novel genetically sensitive research design. Dev Psychopathol 24(4):1283–1295
Hicks BM, DiRago AC, Iacono WG, McGue M (2009a) Gene-environment interplay in internalizing disorders: consistent findings across six environmental risk factors. J Child Psychol Psychiatry 50(10):1309–1317
Hicks BM, South SC, DiRago AC, Iacono WG, McGue M (2009b) Environmental adversity and increasing genetic risk for externalizing disorders. Arch Gen Psychiatry 66(6):640–648
Jarnecke AM, South SC (2014) Genetic and environmental influences on alcohol use problems: moderation by romantic partner support, but not family or friend support. Alcohol Clin Exp Res 38(2):367–375
Kendler KS, Thornton LM, Gilman SE, Kessler RC (2000) Sexual orientation in a national sample of twin and sibling pairs. Am J Psychiatry 157:1843–1846
Kessler RC, Andrews G, Mroczek D, Ustun B, Wittchen H-U (1998a) The World Health Organization Composite International Diagnostic Interview Short-Form (CIDI-SF). Int J Methods Psychiatric Res 7:171–185
Kessler RC, Wittchen H-U, Abelson JM, Kendler KS, Knauper B, McGonagle KA, Zhao S (1998b) Methodological studies of the composite international diagnostic interview (CIDI) in the US national comorbidity survey. Int J Method Psychiatr Res 7:33–55
Kessler RC, Gilman SE, Thornton LM, Kendler KS (2004) Health, wellbeing, and social responsibility in the MIDUS twin and sibling subsamples. In: Brim OG, Ryff CD, Kessler RC (eds) How healthy are we? A national study of wellbeing at midlife. University of Chicago Press, Chicago, pp 124–152
Lachman ME, Weaver SL (1997) The Midlife Development Inventory (MIDI) personality scales: Scale construction and scoring. Waltham
Lau JYF, Eley TC (2008) Disentangling gene-environment correlations and interactions on adolescent depressive symptoms. J Child Psychol Psychiatry 49:142–150
Lykken DT, Bouchard TJ, McGue M, Tellegen A (1990) The Minnesota Twin Family Registry: some initial findings. Acta Genetica Medica Gemellol 39:35–70
Markon K, Krueger RF (2004) An empirical comparison of information-theoretic selection criteria for multivariate behavior genetic models. Behav Genet 34:593–610
McGue M, Bouchard TJ (1984) Quality of twin data for the effects of age and sex. Behav Genet 14:325–343
Monroe SM, Simons AD (1991) Diathesis-stress theories in the context of life stress research: implications for the depressive disorders. Psychol Bull 110:406–425
Muthén LK, Muthén BO (1998-2012) Mplus user’s guide, 7th edn. Los Angeles, CA, Muthen & Muthen
Parker G, Hadzi-Pavlovic D, Greenwald S, Weissman M (1995) Low parental care as a risk factor to lifetime depression in a community sample. J Affect Disord 33(3):173–180
Plomin R, DeFries JC, Knopik V, Neiderhiser JM (2012) Behavioral genetics, 6th edn. Worth Publishers, New York
Purcell S (2002) Variance components models for gene-environment interaction in twin analysis. Twin Res 5:554–571
Raftery AE (1995) Bayesian model selection in social research. Sociol Methodol 25:111–163
Raine A (2002) Biosocial studies of antisocial and violent behavior in children and adults: a review. J Abnorm Child Psychol 30:311–326
Rossi AS (2001) Caring and doing for others: social responsibility in the domains of family, work, and community. University of Chicago Press, Chicago
Rothrauff TC, Cooney TM, An JS (2009) Remembered parenting styles and adjustment in middle and late adulthood. J Gerontol B Psychol Sci Soc Sci 64B(1):137–146
Rowe DC (1981) Environmental and genetic influences on dimensions of perceived parenting: a twin study. Dev Psychol 17:203–208
Russek LG, Schwartz GE (1997) Feelings of parental caring predict health status in midlife: a 35-year follow-up of the Harvard Mastery of Stress Study. J Behav Med 20:1–13
Selzer ML (1971) The Michigan alcohol screening test: the quest for a new diagnostic instrument. Am J Psychiatry 127:89–94
South SC, Krueger RF (2008) Marital quality moderates genetic and environmental influences on the internalizing spectrum. J Abnorm Psychol 117:826–837
South SC, Krueger RF (2011) Genetic and environmental influences on internalizing psychopathology vary as a function of economic status. Psychol Med 41:107–118
South SC, Krueger RF (2013) Marital satisfaction and physical health: evidence for an Orchid effect. Psychol Sci 24(3):373–378
Tuvblad C, Grann M, Lichtenstein P (2006) Heritability for adolescent antisocial behavior differs with socioeconomic status: gene-environment interaction. J Child Psychol Psychiatry 47:734–743
van der Sluis S, Posthuma D, Dolan CV (2012) A note on false positives and power in G × E modelling of twin data. Behav Genet 42(1):170–186
Van Hulle CA, Lahey BB, Rathouz PJ (2013) Operating characteristics of alternative statistical methods for detecting gene-by-measured environment interaction in the presence of gene–environment correlation in twin and sibling studies. Behav Genet 43(1):71–84
Wilhelm K, Niven H, Parker G, Hadzi-Pavlovic D (2005) The stability of the parental bonding instrument over a 20-year period. Psychol Med 35(3):387–393
Wittchen HU (1994) Reliability and validity studies of the WHO composite international diagnostic interview (CIDI): a critical review. J Psychiatr Res 28:57–84
Young-Wolff KC, Enoch M, Prescott CA (2011) The influence of gene-environment interactions on alcohol consumption and alcohol use disorders: a comprehensive review. Clin Psychol Rev 31:800–816
Acknowledgments
This research was supported by the John D. and Catherine T. MacArthur Foundation Research Network on Successful Midlife Development. The authors wish to thank Rob Kail for his comments on this manuscript and Carol Van Hulle for her assistance with the statistical models used in the analyses.
Conflict of Interest
Susan C. South, Amber M. Jarnecke declare that they have no conflict of interest.
Human and Animal Rights and Informed Consent
This study was approved by the Institutional Review Board of Purdue University.
Author information
Authors and Affiliations
Corresponding author
Additional information
Edited by Deborah Finkel.
Rights and permissions
About this article

Cite this article
South, S.C., Jarnecke, A.M. Genetic and Environmental Influences on Adult Mental Health: Evidence for Gene-Environment Interplay as a Function of Maternal and Paternal Discipline and Affection. Behav Genet 45, 438–450 (2015). https://doi.org/10.1007/s10519-015-9716-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10519-015-9716-8