
Abstract
The prevalence of Amblyomma geoemydae infesting yellow-margined box turtle (Cuora flavomarginata) was determined for the first time in northern Taiwan. In total, 352 ticks (140 adults, 61 nymphs and 151 larvae) were collected from 56 yellow-margined box turtles. The overall infestation was observed with an average density of 6.3 ticks per turtle and the highest seasonal prevalence was observed on March with an average tick density of 14.3 ticks per turtle. The higher seasonal prevalence was observed on April, May, July and October with an average density of 4.8, 8.3, 5.7 and 8.5 ticks per turtle, respectively. The seasonal activity of these turtles may contribute to the variation in the number of collected ticks. The genetic identity was determined by comparing the sequences of ITS2 genes obtained from seven Taiwan species of A. geoemydae and nine other species representing eight Amblyomma species and Ixodes persulcatus as outgroup. Our results provide the first zootiological survey of A. geoemydae infesting wild turtles in northern Taiwan and highlight the importance of this tick species in the study of parasite dynamics on testudines in Taiwan. Further study focused on the seasonality pattern of turtle ticks will help reveal the ecology of this host-parasite system and its impact on animal/human health.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Ticks are obligate blood-sucking arthropods that parasitize terrestrial vertebrate groups around the world (Guglielmone et al. 2014). Systematics and nomenclature studies of ticks have shown that the genus Amblyomma is the third largest genus of hard ticks (Guglielmone et al. 2020). Although ticks have been known to be associated with both humans and animals for hundreds of years (Balashov 1972), the global impact of ticks has been recognized by their ability to transmit a variety of tick-borne pathogens and by direct injury to livestock because of their feeding behavior (Jongejan and Uilenberg 2004). Their detrimental impact on humans and animals has become a major concern in medical and veterinary health.
Amblyomma geoemydae is a tick species first described from the tortoises of Bengal (Cantor 1847) and is widely distributed throughout Asia, ranging from South India to the Philippines, Vietnam, Myanmar, Japan China and Taiwan (Yamaguti et al. 1971; Robbins et al. 2006; Robbins and Platt 2011; Sun et al. 2016; Takahashi et al. 2017; Chao et al 2022; Amarga et al. 2022). It has been reported as a common ectoparasite of several species of reptiles, including elongated tortoise (Indotestudo elongata), Asian yellow pond turtles (Mauremys mutica kami) and yellow-margined box turtles (Cuora flavomarginata) (Takano et al. 2012; Bilbija et al. 2019; Kaenkan et al. 2020; Qiu et al. 2021; Chao et al 2022; Amarga et al. 2022). Although this reptilian tick has been recognized as a potential vector for the natural transmission of Rickettsia, Ehrlichia and Borrelia pathogens in wild environment (Takano et al. 2012; Kaenkan et al. 2020; Qiu et al. 2021), very little is known about its biology or life cycle in Taiwan.
The yellow-margined box turtle (Cuora flavomarginata) was first described as Cistoclemmys flavormarginata from material collected from China and Taiwan (listed as Formosa) (Gray 1863) and is found in various provinces along with the Yangtze drainage of Central China (Hsu 1930). Ernst and Lovich (1990) reported this species as Cuora evelynae, a new species of Cuora from the Ryukyu Islands of Japan, but this name was later moved as synonym of C. flavomarginata. This species is an ectothermic terrestrial reptile that commonly habitats the low-elevation forested hills (Zhao 1998; Lin et al. 2010). In Taiwan, C. flavomarginata mainly feeds on earthworms, snails, slugs and mealworms (Chen and Lue 1999, 2002). The adult yellow-margined box turtle has a highly domed shell, the carapace and plastron of which are dark brown with a cream-yellow strip on the vertebral keel. The edge of the plastron is lightly pigmented due to the marginal and plastral scutes near their edges. The external difference between male and female C. flavomarginata is slight, males have a broader tail, whereas females have a thinner tail that is almost triangular in shape. This species is recognized as endangered and protected species in Taiwan as well as listed as endangered under the criteria of the IUCN (https://www.iucnredlist.org).
Because of climatic and geographical factors, the tick species infesting wild hosts may not be the same around the world (Guglielmone et al. 2014). Although the broad geographical distribution of A. geoemydae has been known to exist from southeastern to eastern Asia, the prevalence of this tick species feeding on specific host has never been investigated in Taiwan. Thus, the objectives of this study intend to demonstrate the seasonal prevalence of A. geoemydae infesting C. flavomarginata in northern Taiwan and identify the collected turtle ticks using the internal transcribed spacer gene (ITS2).
Materials and methods
Collection of tick specimens from turtles
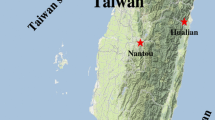
Ticks infesting yellow-margined box turtle were collected every month from April 2021 to March 2022 at the reservation area around the Fei-tsui Reservoir located in the Shiding District of New Taipei City in northern Taiwan (Fig. 1). In general, ticks were collected from the legs, head, body and shell of the turtles and cleaned by sonication in 75% ethanol solution for 5–10 min and then washed twice in sterile distilled water. All cleaned ticks were stored at −20 °C until further analysis (Chao et al. 2022).

Map of Taiwan showing the tick collection site of the Fei-tsui Reservoir in northern Taiwan
Species identification of ticks
Each tick developmental stage was placed on a glass slide and photographed using a SMZ 1500 Nikon stereomicroscope equipped with a fiber lamp (Chao et al. 2013). The external features of the Ambloymma ticks were recorded for species identification, as previously described (Yamaguti et al. 1971; Teng and Jiang 1991; Chao et al. 2022). In addition, molecular identification of collected ticks was also performed by targeting the ITS2 gene, as previously used by Chao et al. (2011).
DNA extraction from tick specimens
Genomic DNA was extracted from individual ticks. Briefly, the individual tick was homogenized in a microcentrifuge tube filled with 180-μl lysing buffer solution (DNeasy Blood &Tissue Kit; Qiagen, Taipei, Taiwan) and then homogenized with a TissueLyser II apparatus (Qiagen, Hilden, Germany), as instructed by the manufacturer. The homogenate was centrifuged at room temperature and the supernatant fluid was further processed by a DNeasy Tissue Kit. After filtration, the filtrated fluid was collected and the DNA concentration was determined spectrophotometrically with a DNA calculator (Epoch, microplate spectrophotometer; BioTek-Agilent, Santa Clara, CA, USA) and the extracted DNA is stored at −80 °C for further investigations.
DNA amplification by polymerase chain reaction (PCR)
DNA samples extracted from the tick specimens were used as a template for PCR amplification. A specific primer set of 3SA (50-CTAAGCGGTGGATCACTCGG-30) corresponding to the 3ʹ end of the 5.8S rDNA and JB9A (50-GCACTATCAAGCAACACGACTC-30) corresponding to the 5ʹ end of the 28S rDNA weas designed to target the ITS2 rDNA gene, as described previously (Barker 1998). All PCR reagents and Taq polymerase were obtained and used as recommended by the supplier (Takara Shuzo, Kyoto, Japan). Briefly, each 25-μl reaction mixture contained 3-μl DNA template, 1.5-μl forward and reverse primers, 2.5-μl 10 × PCR buffer (Mg2+), 2-μl dNTP mixture (10 mM each), 1 unit of Taq DNA polymerase and filled-up with adequate volume of ddH2O. In contrast, adequate amounts of sterile distilled water were added for serving as a negative control. PCR amplification was performed with a Veriti thermocycler (Applied Biosystems, Taipei, Taiwan) and was amplified for 35 cycles with the conditions of denaturation at 95 °C for 30 s, annealing at 50 °C for 30 s, and extension at 68 °C for 2 min., as described previously (Chao et al. 2011). Thereafter, amplified DNA products were electrophoresed on 2% agarose gels in Tris–Borate-EDTA (TBE) buffer and visualized under ultraviolet (UV) light after staining with ethidium bromide. A 100-bp GeneRuler DNA ladder (Thermo Fisher Scientific, Taipei, Taiwan) was used as the standard marker for comparison. A negative control of distilled water was included in parallel with each amplification.
Sequence alignments and phylogenetic analysis
Approximately 10-μl of each selected samples with clear bands on the agarose gel was submitted for DNA sequencing (Mission Biotech, Taipei, Taiwan). After purification (QIAquick PCR Purification Kit), sequencing reaction was performed with 25 cycles under the same conditions and same primer set of nested amplification by dye-deoxy terminator reaction method using the Big Dye Terminator Cycle Sequencing Kit in an ABI Prism 377-96 DNA Sequencer (Applied Biosystems, Foster City, CA, USA). The resulting sequences were initially edited by BioEdit software (v5.3) and aligned with the CLUSTAL W software (Thompson et al. 1994). Thereafter, the aligned sequences of ITS2 genes from seven tick species of Taiwan were analyzed by comparing with nine other tick species from different biological and geographical origins available from GenBank. Phylogenetic analysis was performed by neighbour-joining method to estimate the phylogeny of the entire alignment using MEGA X software package (Kumar et al. 2018). The phylogenetic tree was constructed and performed with 1000 bootstrap replications to evaluate the reliability of the construction, as described previously (Felsenstein 1985).
Results
Collection of various life-stages of Amblyomma geoemydae from turtles
In total, 352 tick specimens representing 99 females, 41 males, 61 nymphs and 151 larvae were collected from 56 turtles (Table 1). Initially, yellow-margined box turtles were captured from the wet-bottomed forest (Fig. 2A) at the reservation area around the Fei-tsui Reservoir in northern Taiwan. The attached ticks were found on the turtles’ carapace (Fig. 2B), neck (Fig. 2C), edge of the plastron (Fig. 2D) and leg (Fig. 2E) of the turtle. A magnified view of a female tick revealed a body length of 8–9 mm with the scutum covering less than half of the body (Fig. 2F).

Field micrographs of Amblyomma geoemydae ticks observed on the yellow-margined box turtle, Cuora flavomarginata. A A turtle in the wet-bottomed forest at the Fei-tsui Reservoir. B Adult ticks attached to the turtle’s carapace. C A partially engorged adult female tick on the neck of a box turtle. D Adult ticks attached to the edge of the plastron, E Adult tick attached to a leg. F An attached female with the scutum covering less than half of the body
Monthly prevalence and infestation of Amblyomma geoemydae on turtles
The monthly prevalence of A. geoemydae infesting turtles was recorded from April 2021 to March 2022 (Fig. 3). The total monthly number of collected ticks ranged from 114 (March; 59 larvae, 24 nymphs, and 31 adults) to two (December; 2 adults) (Table 2; Fig. 3). No ticks were collected in the winter months (January and February) to prevent disturbance of the turtles (Fig. 3). The overall average infestation was 6.29 ticks per turtle (all stages included), ranging from 14.25 ticks per turtle (March) to two ticks per turtle (December, but just a single turtle was examined this month) (Table 2). Higher seasonal mean abundance was observed in April, May, July and October with an average density of 4.82, 8.33, 5.67 and 8.50 ticks per turtle, respectively (Table 2).

Monthly prevalence of various life-stages of Amblyomma geoemydae ticks infesting wild Cuora flavomarginata turtles in northern Taiwan
Genetic and phylogenetic analysis of tick specimens
The sequences of ITS2 rDNA fragments of seven representative samples used in the study were aligned and compared with the downloaded sequences of eight Amblyomma species and one Ixodes outgroup species (Ixodes persulcatus) from GenBank. The aligned nucleotide sequences of 789 bp long of A. geoemydae were highly conserved between the seven Taiwan samples with only a few point mutations/substitutions. All these A. geoemydae of Taiwan were genetically affiliated to a monophyletic group with highly homogeneous sequences (99.2–100% similarity), and can be distinguished from the other lineage group of Amblyomma spp. and the outgroup Ixodes persulcatus (Fig. 4).

Phylogenetic relationships based on ITS2 gene sequences between seven Taiwan strains (samples) of Amblyomma geoemydae ticks from northern Taiwan and nine other species belonging to (eight Amblyomma spp. and one Ixodes species as outgroup). The tree was constructed and analyzed by neighbor-joining method using 1000 bootstraps replicates. Numbers at the nodes indicate the reliability (%) of each branch of the tree. Branch lengths are drawn in proportion to the estimated sequence divergence
Nucleotide sequence accession numbers
The nucleotide sequences of PCR-amplified ITS2 genes of seven strains of A. geoemydae of Taiwan determined in this study have been registered and assigned the following GenBank accession numbers: Amb-L810R11-F1 (ON920915), Amb-L210R8-M1 (ON920870), Amb-L210R8-F2 (ON920919), Amb-L89R12-M1 (ON920857), Amb-L38R11-M2 (ON920858), Amb-L39R12-PEF1 (ON920859), and Amb-L9R212-PEF1 (ON920860). The GenBank accession numbers of the eight other Amblyomma species and the outgroup Ixodes are shown in Fig. 4.
Discussion
This study described the first year-round survey of A. geoemydae infesting C. flavomarginata in northern Taiwan. All life-stages of this tick could be collected almost every month from this turtle host. The highest collection of larval, nymph, and adult ticks was observed in March and April, which is the spring season in northern Taiwan. This observation may be attributed to the availability of turtle hosts in the spring which is highly favorable for the tick feeding. Reduced activity of these turtles in the field was observed during the winter season (December to February), which probably explains the low/no collection of A. geoemydae in this survey in December. Previous studies indicated that one family of snake and three families of tortoises are parasitized by A. geoemydae (Takano et al. 2012; Takahashi et al. 2017; Bilbija et al. 2019; Kaenkan et al. 2020; Qiu et al. 2021). Thus, further investigation on the diverse hosts for A. geoemydae in the field would help to reveal the natural life cycle of A. geoemydae in northern Taiwan.
Although A. geoemydae is known as a naturally inhabited species in Taiwan, the ecology and distribution of this tick species need to be further identified. In previous reports, the yellow-margined box turtle has been recognized as a suitable host for A. geoemydae in various countries (Yamaguti et al 1971; Robbins et al 2006; Takano et al 2012; Tsai et al 2012; Bilbija et al 2019; Qiu et al 2021; Chao et al 2022; Amarga et al 2022). However, this turtle is restricted to low-elevation forest hills in northern Taiwan, and the populations in lowland areas may have been extirpated by land development and habitat fragmentation (Lin et al 2010). Because of extensive habitat loss and commercial exploitation (such as burgeoning demand in the food and pet markets), C. flavomarginata is categorized as endangered species on the ICUN Red List. Further study focused on the seasonal dynamics of A. geoemydae on reptilian hosts will help reveal the ecology of this host-parasite system in Taiwan.
Although morphological features of the adult stage of hard ticks have traditionally been recognized in pictorial keys for species identification (Keirans and Clifford 1978; Sonenshine 1991), a DNA-based investigation provides the opportunity to discriminate the genetic variance at the individual base-pair level and gives much more direct measures of the genetic diversity between and within species of Ixodidae (Black and Roehrdanz 1998; Crosbie et al. 1998; Chigagure et al. 2000). Indeed, previous phylogenetic studies based on rDNA sequences of ITS2 region also demonstrated the heterogeneity among the closely related tick species (Zahler et al. 1997; Rich et al. 1997; Barker 1998; Fukunaga et al. 2000). Through genetic analysis, the turtle ticks collected in this study were identified as A. geoemydae with highly homogeneous sequences (99.2–100% similarity) associated with A. geoemydae from Germany (GenBank no. MH130071), and could be distinguished from the other Amblyomma spp. and the outgroup Ixodes sp. (Fig. 4). Thus, molecular analysis based on the genetic analysis of ITS2 region has made particular success for the identification and discrimination of taxonomically similar tick species.
In conclusion, this year-round survey reveals the seasonality pattern of A. geoemydae infesting yellow-margined box turtle in northern Taiwan. Further investigations focused on various tick-borne pathogens in this tick species would help to illustrate the epidemiological and geographical impacts of A. geoemydae on human/animal health in northern Taiwan.
References
Amarga AKS, Supsup CE, Tseng HY et al (2022) The Asian turtle tick Amblyomma geoemydae Cantor, 1847 (Acari: Ixodidae) in the Philippines: first confirmed local host and locality with a complete host index. Ticks Tick-Borne Dis 13:101958
Balashov YS (1972) Bloodsucking ticks (Ixodoidea)-vectors of diseases of man and animals. Misc Publ Entomol Soc Am 8:268–305
Barker SC (1998) Distinguishing species and populations of Rhipicephalus ticks with ITS 2 ribosomal RNA. J Parasitol 84:887–892
Bilbija B, Auer M, Siroky P (2019) Long term persistence of introduced Amblyomma geoemydae tick population under indoor conditions in Austria. Med Vet Entomol 33:317–321
Black WC IV, Roehrdanz RL (1998) Mitochondrial gene order is not conserved in arthropods: prostriate and metastriate tick mitochondrial genomes. Mol Biol Evol 15:1772–1785
Chao LL, Wu WJ, Shih CM (2011) Species identification of Ixodes granulatus based on the ribosomal DNA spacer, internal transcribed spacer 2 (ITS-2), sequences. Exp App Acarol 54:51–63
Chao LL, Hsieh CK, Shih CM (2013) First report of Amblyomma helvolum (Acari: Ixodidae) from the Taiwan stink snake, Elaphe carinata (Reptilia: Colubridae), collected in southern Taiwan. Ticks Tick Dis 4:246–250
Chao LL, Chen TH, Lien W et al (2022) Molecular and morphological identification of a reptile-associated tick, Amblyomma geoemydae (Acari: Ixodidae), infesting wild yellow-margined box turtles (Cuora flavomarginata) in northern Taiwan. Ticks Tick-Borne Dis 13(2):101901
Chen TH, Lue KY (1999) Population characteristics and egg production of the yellow-margined box turtle, Cuora flavomarginata, in northern Taiwan. Herpetologica 55:487–498
Chen TH, Lue KY (2002) Growth patterns of the yellow-margined box turtle (Cuora flavomarginata) in northern Taiwan. J Herpetol 36(2):201–208
Chigagure NN, Baxter GD, Barker SC (2000) Microsatellite loci of the cattle tick Boophilus microplus (Acari: Ixodidae). Exp Appl Acarol 24:951–956
Crosbie PR, Boyce WM, Rodwell TC (1998) DNA sequence variation in Dermacentor hunteri and estimated phylogenies of Dermacentor spp. (Acari: Ixodidae) in the new world. J Med Entomol 35:277–288
Ernst CH, Lovich JE (1990) A new species of Cuora (Reptilia: Testudines: Emydidae) from the Ryukyu Islands. Proc Biol Soc Wash 103:26–34
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 52:1119–1134
Fukunaga M, Yabuki M, Hamase A et al (2000) Molecular phylogenetic analysis of ixodid ticks based on the ribosomal DNA spacer, internal transcribed spacer 2, sequences. J Parasitol 86:38–42
Gray JE (1863) Observations on the box tortoises, with the descriptions of three new Asiatic species. Proc Zool Soc Lond 1863:173–179
Guglielmone AA, Robbins RG, Apanaskevich DA et al (2014) The hard ticks of the world. Springer, New York, p 738
Guglielmone AA, Petney TN, Robbins RG (2020) Ixodidae (Acari: Ixodidea): descriptions and redescriptions of all known species from 1758 to December 31, 2019. Zootaxa 4871(1):1–322
Hsu HF (1930) Preliminary note on a new variety of Cyclemys flavomarginata from China. Contributions from the Biological Laboratory of the Science Society of China. Zool Ser 6(1):1–7
Jongejan F, Uilenberg G (2004) The global importance of ticks. Parasitology 129:S3–S14
Kaenkan W, Nooma W, Chelong IA et al (2020) Reptile-associated Borrelia spp. in Amblyomma ticks, Thailand. Ticks Tick-Borne Dis 11:101315
Keirans JE, Clifford CM (1978) The genus Ixodes in the United States: a scanning electron microscope study and key to the adults. J Med Entomol Suppl 2:1–149
Kumar S, Stecher G, Li M et al (2018) MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol Bio Evol 35:1547–1549
Lin YF, Wu SH, Lin TE et al (2010) Population status and distribution of the endangered yellow-margined box turtle Cuora flavomarginata in Taiwan. Int J Conserv 44(4):581–587
Qiu Y, Kidera N, Hayashi M et al (2021) Rickettsia spp. and Ehrlichia spp. in Amblyomma ticks parasitizing wild amphibious sea kraits and yellow-margined box turtles in Okinawa, Japan. Ticks Tick-Borne Dis 12(2):101636
Rich SM, Rosenthal BM, Telford SR III et al (1997) Heterogeneity of the internal transcribed spacer (ITS-2) region within individual deer ticks. Insect Mol Biol 6:123–129
Robbins RG, Phong BD, McCormack T et al (2006) Four new host records for Amblyomma geoemydae (Cantor) (Acari: Ixodida: Ixodidae) from captive tortoises and freshwater turtles (Reptilia: Testudines) in the turtle conservation center, Cuc Phuong National park. Vietnam Proc Entomol Soc Wash 108:726–729
Robbins RG, Platt SG (2011) Amblyomma geoemydae (Cantor) (Acari: Ixodida: Ixodidae): first report from the Arakan forest turtle, Heasemys depressa (Anderson) (Reptilia: Testudines: Emydidae) and first documented occurrence of this tick in the Union of Myanmar. Int J Acaril 37:103–105
Sonenshine DE (1991) Biology of ticks. Oxford University Press, New York
Sun Y, Zhang SG, Ye XD et al (2016) Systemic classification and pictorial key of Amblyomma ticks (Ixodida: Ixodidae) in China. Acta Parasitol Med Entomol Sin 23:111–121 (In Chinese)
Takahashi M, Misumi H, Tamura H (2017) First record in Japan of tortoise tick Amblyomma geoemydae (Cantor 1847) (Acari: Ixodidae) parsitizing Prier’s keelback snake Hebius pryeri (Boulenger, 1887) (Repile: Colubridae). Int J Acarol 43:314–319
Takano A, Sugimori C, Fujita H et al (2012) A novel relapsing fever Borrelia sp. infects the salivary glands of the molted hard tick Amblyomma Geoemydae. Ticks Tick Dis 3:259–261
Teng KF, Jiang ZJ (1991) Economic insect fauna of China Fasc 39 Acari: Ixodidae. Science Press, Beijing (In Chinese)
Tsai YL, Shyu CL, Yao CT et al (2012) The ixodid ticks collected from dogs and other animals in Taiwan and Kinmen Island. Int J Acarol 389(2):110–115
Thompson JD, Higgins DG, Gibson TJ (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nuc Acids Res 22:4673–4680
Yamaguti N, Tipton VJ, Keegan HL, Toshioka S (1971) Ticks of Japan, Korea, and the Ryukyu Islands. Brigham Young University Sci. Bul. Biol. Ser., 15
Zahler M, Filippova NA, More PC et al (1997) Relationships between species of the Rhipicephalus sanguineus group: a molecular approach. J Parasit 83:302–306
Zhao E (1998) China red data book of endangered animals, amphibian and reptilia. Science Press, Beijing
Acknowledgements
We thank the staff of Taipei Fei-tsui Reservoir Administration for their technical assistance and permission for collecting ticks from the Reservoir area based on a research project (FL110026) sponsored by the Taipei Fei-tui Reservoir Administration. This work was supported in part by a Grant (NPUST-KMU-111-P007) from the cooperative project between National Pingtung University of Science and Technology, and Kaohsiung Medical University, Taiwan, Republic of China.
Author information
Authors and Affiliations
Contributions
LLC and CMS wrote the main manuscript text and prepared the figures and tables, THC worked on the field collection of ticks, and LLC performed the molecular analysis. All authors have read and agree its submission.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Chao, LL., Chen, TH. & Shih, CM. First zootiological survey of Amblyomma geoemydae ticks (Acari: Ixodidae) infesting a wild turtle (Cuora flavomarginata) in northern Taiwan. Exp Appl Acarol 88, 349–359 (2022). https://doi.org/10.1007/s10493-022-00758-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-022-00758-z