
Abstract
The predatory mite Amblyseius swirskii quickly became one of the most successful biocontrol agents in protected cultivation after its introduction into the market in 2005 and is now released in more than 50 countries. There are several key factors contributing to this success: (1) it can control several major pests including the western flower thrips, Frankliniella occidentalis, the whiteflies Bemisia tabaci and Trialeurodes vaporariorum and the broad mite, Polyphagotarsonemus latus, simultaneously in vegetables and ornamental crops; (2) it can develop and reproduce feeding on non-prey food sources such as pollen, which allows populations of the predator to build up on plants before the pests are present and to persist in the crop during periods when prey is scarce or absent; and (3) it can be easily reared on factitious prey, which allows economic mass production. However, despite the fact that A. swirskii provides growers with a robust control method, external demands were initially a key factor in promoting the use of this predator, particularly in Spain. In 2006, when exports of fresh vegetables from Spain were stopped due to the presence of pesticide residues, growers were forced to look for alternatives to chemical control. This resulted in the massive adoption of biological control-based integrated pest management programmes based on the use of A. swirskii in sweet pepper. Biological control increased from 5 % in 2005, 1 year before A. swirskii was commercially released, to almost 100 % of a total 6,000 ha of protected sweet pepper in Spain within 3 years. Later, it was demonstrated that A. swirskii was equally effective in other crops and countries, resulting in extensive worldwide use of A. swirskii in greenhouses.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Biological control has been one of the most environmentally safe and most economical modes of pest management for growers (Cock et al. 2010). Augmentative biological control, i.e. the release of mass-reared natural enemies in large numbers to obtain control of pests, has been an environmentally and economically sound alternative to chemical control in several agricultural systems, mainly in protected cultivation (van Lenteren and Bueno 2003). The biocontrol industry has made great advances in the last decades in the identification of natural enemies and development of commercial products, leading to more than 230 species of natural enemies available for augmentative biological control worldwide (van Lenteren 2012). The predatory mite Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) is one of the most successful of these biocontrol agents and is currently released in more than 50 countries all over the world.
In this paper we describe the development of A. swirskii into a biocontrol agent, putting emphasis on factors we consider important for its success in the market as well as on the importance of A. swirskii for the breakthrough of augmentative biological control in greenhouse vegetable production in Spain first, and other countries thereafter. The Spanish case constitutes an example for the successful switch from purely chemical pest management to biocontrol-based integrated pest management (IPM) stimulated by the availability of an effective biocontrol agent and other factors.
Use of Phytoseiidae as biocontrol agents
The Phytoseiid mite Phytoseiulus persimilis Athias-Henriot (Acari: Phytoseiidae), used for biological control of the two spotted spider mite Tetranychus urticae Koch (Acari: Tetranychidae), was one of the earliest biocontrol agents commercially available in Europe in 1968. However, the use of Phytoseiidae as biocontrol agents for insect pests started considerably later with the use of Neoseiulus barkeri Hughes and Neoseiulus cucumeris (Oudemans) (Acari: Phytoseiidae) for thrips control, which have been on the market since 1981 and 1985, respectively (van Lenteren 2012). Further research on the use of phytoseiids as biocontrol agents of insect pests was conducted in the 1990s after the invasion of the western flower thrips, Frankliniella occidentalis (Pergande) (Thysanoptera: Thripidae), into Europe (van Houten et al. 1993, 1995). The invasion of new biotypes of Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) into the New World (Brown et al. 1995) and Europe (Fransen 1994) triggered the interest in Phytoseiidae as biocontrol agents of whiteflies (Nomikou et al. 2001).
The development of Amblyseius swirskii into a biocontrol agent
Origin, distribution and early research
Amblyseius swirskii originates from the East Mediterranean coast and was described in 1962 from almond (Prunus amygdalus) in Bet Dagan, Israel (Athias-Henriot 1962). It naturally occurs in citrus crops along the Israeli coast (Porath and Swirski 1965). In addition, it has been found on other fruit trees, grapes, vegetables, cotton, wild trees and shrubs, as well as in various annual and perennial plants in this country (Swirski and Amitai 1997). In addition to Israel, it has been reported from other Middle Eastern countries, Southern Europe, West, Central and East Africa and North, Central and South America (Demite et al. 2014).
The first short study on its biology was published by Teich (1966) who discovered that eggs and larvae of B. tabaci in a laboratory culture were eaten by A. swirskii and Amblyseius rubini Swirskii and Amitai (Acari: Phytoseiidae). Shortly thereafter, Swirski et al. (1967) published a detailed study showing that A. swirskii has a wide prey range and can feed on several mite and insect species as well as pollen of many plant species. Thereafter, only a few scattered laboratory studies were conducted on the biology of A. swirskii feeding on different prey and non-prey food (Ragusa and Swirski 1975, 1977; Metwally et al. 1984; Hoda et al. 1986; El-Laithy and Fouly 1992; Momen and El-Saway 1993; for details see Table 1).
Development of Amblyseius swirskii as a biocontrol agent for whiteflies and thrips
Amblyseius swirskii can develop and reproduce on a wide range of arthropod prey (Table 1; see section Feeding on other pests and non-prey food), but it is mainly used for augmentative biological control of the whiteflies B. tabaci and Trialeurodes vaporariorum (Westwood) (Hemiptera: Aleyrodidae) and the thrips F. occidentalis (Cock et al. 2010). After the publications of Teich (1966) and Swirski et al. (1967) several other papers were published on the ability of Phytoseiidae to prey on whiteflies mainly in the 1960s and early 1970s (see Nomikou et al. 2001 for a summary); however, these findings were not taken up by the biocontrol industry. Biocontrol of whiteflies largely relied on the parasitoids Encarsia formosa Gahan, Eretmocerus eremicus Rose and Zolnerowich and Eretmocerus mundus Mercet (all Hymenoptera: Aphelinidae), and predatory bugs such as Macrolophus pygmaeus (Rambur) (then named Macrolophus caliginosus Wagner) and Nesidiocoris tenuis Reuter (both Hemiptera: Miridae) (van Lenteren and Martin 1999; Calvo and Urbaneja 2004; Stansly et al. 2004, 2005; Calvo et al. 2009, 2012).
Interest into phytoseiid mites as biocontrol agents for whiteflies re-emerged in the late 1990s. Worldwide outbreaks of B. tabaci caused serious yield losses in many crops as a result of direct feeding damage and transmission of viruses (Brown et al. 1995; Gerling and Mayer 1996; De Barro et al. 2011). In addition, development of resistance to most of the available insecticides made control more difficult (Cahill et al. 1996a, b; Nauen and Denholm 2005; Fernández et al. 2009). Hence, B. tabaci rapidly became a serious pest in protected crops and there was renewed interest in alternative control methods. As part of this search for alternative control options, surveys were started in Israel and Jordan to search for predatory mites feeding on B. tabaci in the field (Nomikou et al. 2001).
In laboratory experiments conducted with the species collected during these surveys, A. swirskii, Typhlodromus athiasae Porath and Swirski, N. barkeri and Euseius scutalis (Athias-Henriot) (all Acari: Phytoseiidae) oviposited with B. tabaci as prey. A. swirskii and E. scutalis were selected for further evaluation because they had higher intrinsic rates of increase (rm) than the other two species (Nomikou et al. 2001). Both A. swirskii and E. scutalis were able to suppress B. tabaci populations on isolated cucumber plants in a greenhouse experiment when Typha sp. pollen was provided as supplementary food (Nomikou et al. 2002).
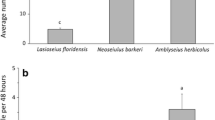
In a comparison with other predatory mite species, A. swirskii also provided good control of F. occidentalis in cucumber. It was much better than N. cucumeris, the predatory mite commonly used for augmentative biological control of thrips in cucumber and other crops at that time, although Amblydromalus limonicus Garman and McGregor (Acari: Phytoseiidae) provided even better thrips control than A. swirskii. Thrips control with E. scutalis was not successful despite the fact that this mite developed much higher populations on cucumber plants than N. cucumeris. Euseius scutalis was mainly present on the lower leaves of the cucumber plants whereas the thrips reached high densities on the upper leaves (Messelink et al. 2005, 2006). Further research in The Netherlands focused on A. swirskii because it had the potential to control both pests and because it was easier and cheaper to rear than A. limonicus and E. scutalis (Nomikou et al. 2003, 2010; Messelink et al. 2008, 2010; Knapp et al. 2013).
The results of these experiments caught the attention of Koppert Biological Systems, a commercial biocontrol company. Amblyseius swirskii was obtained from the University of Amsterdam and additional experiments were conducted. Major outcomes of these trials were that A. swirskii also developed and multiplied with the greenhouse whitefly T. vaporariorum as prey and was able to establish on flowering sweet pepper with only pollen as food source in commercial greenhouses in The Netherlands (Bolckmans et al. 2005). Control of F. occidentalis in sweet pepper with A. swirskii was better than with N. cucumeris and Amblyseius andersoni (Chant) and as good as with Iphiseius degenerans (Berlese) (both Acari: Phytoseiidae) under Dutch summer conditions (van Houten et al. 2005). At the same time a mass-rearing system based on the use of the factitious prey Carpoglyphus lactis (L.) (Acari: Carpoglyphidae), a stored-product mite that is easy to rear, was developed and patented (Bolckmans and van Houten 2006).
Despite the impressive data, the uptake of A. swirskii by Dutch growers initially started slowly. Many of the sweet pepper growers were content with the combination of N. cucumeris and Orius laevigatus Fieber (Hemiptera: Anthocoridae) they were using for thrips control. Whiteflies were not a big problem in Dutch sweet pepper greenhouses. An additional disadvantage was that, in contrast to N. cucumeris, no slow-release sachets were available for A. swirskii.
The success of Amblyseius swirskii in Spain
In the early 2000s there were increasing problems with F. occidentalis and B. tabaci control in protected sweet pepper crops in south-eastern Spain. The growers still largely relied on synthetic insecticides to control these pests and were caught in a vicious circle: increasing resistance of both pests to insecticides (e.g. Espinosa et al. 2002; Fernández et al. 2009) and pest resurgence (i.e. pesticide-induced pest outbreaks) led to increasing application frequencies and dose rates, and ultimately to the use of illegal pesticides. This, in turn, caused even more pest problems, increasing residue levels and environmental impact, and decreasing food safety. Biocontrol companies had tried to introduce IPM in greenhouse vegetable production in southern Spain for several years without success, mainly because they initially tried to copy the strategies used in The Netherlands, based on the use of E. formosa and E. eremicus for whitefly control and N. cucumeris and O. laevigatus for thrips control. However, the climate and greenhouse technology in southern Spain was different from that in The Netherlands—the Dutch solutions did not work sufficiently under these conditions, leading to poor establishment of beneficials, and they were too expensive. Later attempts to adapt these methods to local conditions resulted in the replacement of the parasitoids by the indigenous parasitoid E. mundus, which was more effective in B. tabaci control than the above-mentioned parasitoids in southern Spain (Stansly et al. 2004, 2005). However, results were still disappointing, especially in summer plantings and thus, growers continued to rely on chemical control. In 2005, initial semi-field experiments demonstrated that biological control of B. tabaci on sweet pepper with A. swirskii was possible in southern Spain and promising results were also obtained for control of F. occidentalis (Hoogerbrugge et al. 2005; Belda and Calvo 2006). However, it was still difficult to convince the growers that biocontrol could work in practice.
In 2006 this situation changed. Greenpeace Germany had sent sweet pepper from German supermarkets to a laboratory for residue analysis. The results were dramatic. Residues of isofenphos-methyl were detected in 60 % of the samples from Spain. This active ingredient was and is not registered in the European Union; in 40 % of the samples the isofenphos-methyl values were above the maximum residue level. In samples of other origins (Turkey, The Netherlands, Morocco) isofenphos-methyl was not detected (CVUA Stuttgart 2007). Greenpeace published these results widely (see e.g. Krautter 2007) and German consumers largely stopped buying Spanish sweet peppers. The major market for the Spanish growers collapsed, and suddenly their interest in biocontrol re-emerged.
The positive results obtained in semi-field trials in Spain mentioned above were confirmed in further experiments. Releases of A. swirskii in combination with E. mundus significantly improved control of B. tabaci compared to using the parasitoid only in protected sweet pepper (Calvo and Belda 2007; Calvo et al. 2009), which was later confirmed in commercial greenhouses (Calvo et al. 2012). Releases of A. swirskii significantly reduced F. occidentalis densities in sweet pepper, but the thrips was not sufficiently controlled in sweet pepper flowers to mitigate the risk of Tomato-spotted wilt virus infections (Belda and Calvo 2006). As in The Netherlands, the mite therefore needed to be combined with O. laevigatus (Weintraub et al. 2011; Calvo et al. 2012). Overall, biological control of both pests with the system based on A. swirskii was not only more efficient, but also cheaper than the previously used strategy consisting of parasitoids for whitefly control and N. cucumeris and O. laevigatus for thrips control (Calvo et al. 2012). A. swirskii also provided excellent control of B. tabaci and F. occidentalis in cucumber, eggplant, melon and courgette in southern Spain (Calvo et al. 2008, 2011).
Within 3 years, nearly all sweet pepper growers switched from chemical control to biological control of both pests (van der Blom 2005; Merino-Pacheco 2007; van der Blom et al. 2008). The area of sweet pepper under biological control increased from a mere 200 ha in the season of 2005–2006 to about 6,000 ha in 2008–2009; Fig. 1). At the same time, the percentage of sweet pepper samples with pesticide residues decreased from 33.3 % in January 2007 to <1 % throughout the season 2007–2008 (Glass and Gonzalez 2012). In 2012–2013, the total greenhouse area in Spain under IPM in which A. swirskii was released was around 18,000 ha, including more than 2,100 ha cucumber, more than 950 ha eggplant, around 1,100 ha zucchini and about 6,000 ha of other crops (Fig. 1).

Greenhouse area (ha) by crop in Spain under biological control-based integrated pest management programmes based on the use of Amblyseius swirskii (Frutas y Hortalizas 2014). Amblyseius swirskii was first commercially released in 2006–2007 in sweet pepper
Further characteristics of Amblyseius swirskii making it a successful biocontrol agent
Wide range of application: host plants and temperature
In addition to various wild plants (see section Origin, distribution and early research), A. swirskii is able to establish in many vegetable crops, including peppers, cucumber or eggplants as well as in ornamentals and fruit trees (Calvo and Belda 2007; Calvo et al. 2012; Gerson and Weintraub 2012; Juan-Blasco et al. 2012). In life-table studies with Typha latifolia pollen as food source in the laboratory, no development occurred at 13 °C and the rm was negative at 15 °C. The highest rm (0.160) was recorded at 32 °C; the estimated lower threshold for population growth was 15.5 °C and the estimated upper threshold 37.0 °C. This implies that populations should grow quickly between 20 and 32 °C, a temperature range common in many agro-systems, but population growth could be slow below 20 °C (Lee and Gillespie 2011).
Feeding on other pests and non-prey food
Amblyseius swirskii is a generalist predatory mite feeding on many different small insects and mites (Table 1) as well as on pollen from many plants (Ragusa and Swirski 1975; Goleva and Zebitz 2013) and other non-prey food, including eggs of the Mediterranean flour moth Ephestia kuehniella Zeller (Lepidoptera: Pyralidae), decapsulated dry cysts of the brine shrimp Artemia franciscana Kellogg, mango powdery mildew Oidium mangiferae Berthet and various artificial diets (Abou-Awad et al. 2011; Nguyen et al. 2013, 2014a, b). Its ability to feed on pollen greatly helps to establish in high numbers in flowering plants, for instance in sweet pepper, even before pests are present (Bolckmans et al. 2005). This ability to build up populations on pepper pollen makes it also possible to use ornamental peppers as banker plants for A. swirskii in ornamental greenhouses to increase predatory mite densities (Xiao et al. 2012a; Avery et al. 2014; Buitenhuis et al. 2015). As a generalist, A. swirskii can be used for the control of different pests simultaneously. Although the presence of whitefly can lead to a short-term escape of thrips from predation (van Maanen et al. 2012), thrips control is not negatively affected by the presence of B. tabaci or T. vaporariorum, or vice versa, in greenhouses (Messelink et al. 2008; Calvo et al. 2011). To the contrary, Messelink et al. (2008), (2010) observed that the presence of thrips and spider mites enhanced T. vaporariorum control by A. swirskii by increasing predator densities, because the generalist A. swirskii performs better on a mixed pest diet.
Amblyseius swirskii can also feed and reproduce on T. urticae (El-Laithy and Fouly 1992; Xiao et al. 2012b); however, it has a clear preference for F. occidentalis (Xu and Enkegaard 2010). It is not able to control spider mites in the absence of other pests on greenhouse cucumbers, mainly because the predatory mites cannot enter dense spider mite webbing and therefore can only feed on spider mites outside or near the edges of the webbing (van Houten et al. 2007a; Messelink et al. 2010). However, spider mite damage was much lower in the presence than in the absence of F. occidentalis and/or T. vaporariorum, probably due to the strong numerical response of the predator when these pests were present (Messelink et al. 2010).
Good results have been achieved with A. swirskii in the control of broad mites, Polyphagotarsonemus latus (Banks) (Acari: Tarsonemidae), in sweet pepper, hot pepper, eggplant and gboma eggplant (Solanum macrocarpon) (Tal et al. 2007; Stansly and Castillo 2009; van Maanen et al. 2010; Onzo et al. 2012; Abou-Awad et al. 2014a, b). Preliminary experiments have indicated that A. swirskii could play an important role in the control of the invasive red palm mite, Raoiella indica Hirst (Acari: Tenuipalpidae) (Peña et al. 2009). In phalaenopsis (Orchidaceae), Brevipalpus sp. (Acari: Tenuipalpidae) has been controlled with releases of A. swirskii (H. Nennmann, Pflanzenschutzdienst Nordrhein Westfalen, pers. comm.). The chilli thrips, Scirtothrips dorsalis Hood (Thysanoptera: Thripidae), can also be controlled by A. swirskii in pepper (Arthurs et al. 2009; Dogramaci et al. 2011).
More recent research has shown that A. swirskii can also feed on citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae), and significantly reduce citrus psyllid populations on isolated plants in a greenhouse (Juan-Blasco et al. 2012). It still remains to be investigated whether A. swirskii can contribute to the control of citrus psyllid in the field. It is important to note that the ability to feed and reproduce on a specific pest in laboratory trials does not guarantee successful control of this pest. Amblyseius swirskii develops and reproduces very well on the tomato russet mite, Aculops lycopersici (Massee) (Acari: Eriophyidae), on leaf discs in Petri dishes (Park et al. 2010, 2011); however, it cannot control this pest on tomato due to the glandular trichomes on stems and leaves of tomato plants, which impair the movement of the predatory mite (van Houten et al. 2013).
Combination with other control agents
Amblyseius swirskii has been combined with other biocontrol agents in various crops, with no detrimental effects on biological control of whitefly, thrips, spider mites and other pests (van Houten et al. 2007b; Chow et al. 2010; Messelink et al. 2010; Calvo et al. 2012), with one known exception. When it is combined with the aphidophagous gall midge Aphidoletes aphidimyza (Rondani) (Diptera: Cecidomyiidae), predation of the predatory mite on the eggs of the gall midge can disrupt biological control of aphids (Messelink et al. 2011). In addition, there are many pesticides that are compatible with A. swirskii (Calvo et al. 2007; Gradish et al. 2011), which allows the simultaneous control of pests and diseases that cannot be controlled biologically.
Release systems
Good coverage and dispersal of predatory mites is important for biological control, especially in ornamentals, where the pest levels that can be tolerated are lower than in vegetables. Therefore, patches of prey can be sparsely distributed and there is often not enough prey to maintain a predator population (Skirvin and Fenlon 2003; Van Driesche and Heinz 2004; Buitenhuis et al. 2010, 2015). To achieve this, various release methods have been developed for A. swirskii since its introduction. Initially, the predatory mites were only available in bottles with wheat bran as carrier material, and needed to be distributed by hand. Later, release systems based on mechanical blowers were developed, which distribute the mites more uniformly and reduce labour cost (Opit et al. 2005; Pezzi et al. 2015; see also http://www.koppert.com/products/distribution-appliances/). Another release method is the ‘slow release’, using breeding sachets. These sachets contain a carrier material such as wheat bran, and a food source for the factitious prey mites. The predatory mites feed and multiply in the sachet and can leave through a small hole that is punched into the sachets and disperse into the crop for several weeks (Midthassel et al. 2014). Sachets are therefore a valuable option in systems where no alternative food in form of pollen is available and preventive releases help to keep pest levels low (Midthassel et al. 2014). For A. swirskii, commercial slow-release sachets can contain the factitious prey C. lactis, Suidasia medanensis (Oudemans) (Acari: Suidasiidae) or Thyreophagus entomophagus (Laboulbène and Robin) (Acari: Acaridae) (Bolckmans and van Houten 2006; Fidgett and Stinson 2008; Baxter et al. 2011; Midthassel et al. 2013; Nguyen et al. 2013). Recently, a slow-release sachet containing two prey mites, C. lactis and Lepidoglyphus destructor (Schrank) (Acari: Glycyphagidae), was developed—this combination increases the lifetime of the breeding sachet significantly from 3–4 to 6–8 weeks (Bolckmans et al. 2013).
Conclusions
The pathway for uptake of innovations in pest management is often envisaged as a linear process of (1) basic research, (2) laboratory trials, (3) greenhouse research trials, (4) commercial trials, and (5) adoption (Murphy 2014). The development of A. swirskii into a successful biocontrol agent largely followed this pathway. Initial laboratory experiments were carried out at the University of Amsterdam, greenhouse trials at the Glasshouse Horticulture Division of Applied Plant Research in The Netherlands (now WUR Glasshouse Horticulture). Based on the results of these trials, A. swirskii caught the attention of Koppert Biological Systems, and more semi-field experiments and commercial trials were conducted by this company, both in The Netherlands and Spain. As shown above, adoption was initially largely driven by external factors, i.e. pesticide resistance and residue problems. After the initial successes in Spain, biocontrol strategies based on A. swirskii were quickly adopted by many growers of vegetable and ornamental crops in various parts of the world. This, in turn, stimulated further research and the development of better rearing and release systems, which again opened further possibilities for using this predatory mite.
Amblyseius swirskii was put on the market in 2005. In 2009 it was already used in more than 20 countries (Cock et al. 2010) and more than 18,000 ha of protected crops in Spain (Fig. 1) and in 2014 it is sold in more than 50 countries. The large-scale success of A. swirskii would have never been possible without the enthusiasm and expertise of advisors and consultants both from private companies and government agencies, such as universities and research stations, and last but not least, motivated and knowledgeable growers. Growers are the last link in the chain and how they use natural enemies will ultimately determine their success.
References
Abou-Awad BA, El-Sawaf BM, Abdel-Khalek AA (1999) Impact of two eriophyoid fig mites, Aceria ficus and Rhyncaphytoptus fucifoliae, as prey on postembryonic development and oviposition rate of the predacious mite Amblyseius swirskii. Acarologia 40:367–371
Abou-Awad BA, Metwally AM, Al-Azzazy MM (2010) Typhlodromips swirskii (Acari: Phytoseiidae) a predator of eriophyid and tetranychid mango mites in Egypt. Acta Phytopathol Entomol Hung 45:135–148
Abou-Awad BA, Afia S, Al-Azzazy MM (2011) Mango powdery mildew Oidium mangiferae an alternative food for the predatory mites Typhlodromus mangiferus and Typhlodromips swirskii (Phytoseiidae) in absence or presence increasing prey density of Oligonychus mangiferus (Tetranychidae) in Egypt. Arch Phytopathol Plant Prot 44:1703–1710
Abou-Awad BA, Hafez SM, Farhat BM (2014a) Biological studies of the predacious mite Amblyseius swirskii, a predator of the broad mite Polyphagotarsonemus latus on pepper plants (Acari: Phytoseiidae: Tarsonemidae). Arch Phytopathol Plant Prot 47:349–354
Abou-Awad BA, Hafez SM, Farhat BM (2014b) Bionomics and control of the broad mite Polyphagotarsonemus latus (Banks) (Acari: Tarsonemidae). Arch Phytopathol Plant Prot 47:631–641
Abou-Ellela GM, Saber SA, El-Sawi SA (2013) Biological aspects and life tables of the predacious mites, Typhlodromips swirskii (Athias-Henriot) and Euseius scutalis (Athias-Henriot) feeding on two scale insect species and plant pollen. Arch Phytopathol Plant Prot 46:1717–1725
Ali FS, Zaher MA (2007) Effect of food and temperature on the biology of Typhlodrompis swirskii (Athias-Henriot) (Acari: Phytoseiidae). Acarines 1:17–21
Arthurs S, McKenzie CL, Chen J, Dogramaci M, Brennan M, Houben K, Osborne L (2009) Evaluation of Neoseiulus cucumeris and Amblyseius swirskii (Acari: Phytoseiidae) as biological control agents of chilli thrips, Scirtothrips dorsalis (Thysanoptera: Thripidae) on pepper. Biol Control 49:91–96
Athias-Henriot C (1962) Amblyseius swirskii, un nouveau phytoseiide voisin d’ A. andersoni (Acariens Anactinotriches). Ann Ec Natl Agric Alger 3:1–7
Avery PB, Kumar V, Xiao Y, Powell CA, McKenzie CL, Osborne LS (2014) Selecting an ornamental pepper banker plant for Amblyseius swirskii in floriculture crops. Arthropod Plant Interact 8:49–56
Baxter I, Midthassel A, Stepman W, Fryer R, Puerto-Garcia F, Lewis J, Walker P, Hulshof J (2011) Field results of a sachet release system using the predator Amblyseius swirskii (Athias-Henriot) (Acari: Phytoseiidae) and the factitious prey, Suidasia medanensis Oudemans (Acari: Astigmata). IOBC WPRS Bull 68:1–4
Belda JE, Calvo J (2006) Eficacia de Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) en el control biológico de Bemisia tabaci (Genn.) (Hom.: Aleyrodidae) y Frankliniella occidentalis (Pergande) (Thys.: Thripidae) en pimiento en condiciones de semicampo. Bol Sanid Veg Plagas 32:283–296
Bolckmans KJF, van Houten YM (2006) Mite composition, use thereof, method for rearing the phytoseiid predatory mite Amblyseius swirskii, rearing system for rearing said phytoseiid mite and methods for biological pest control on a crop. WO Patent WO/2006/057552
Bolckmans K, van Houten Y, Hoogerbrugge H (2005) Biological control of whiteflies and western flower thrips in greenhouse sweet peppers with the phytoseiid predatory mite Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae). In: Proceedings 2nd International Symposium on Biological Control of Arthropods, pp 555–565
Bolckmans KJF, van Houten YM, van Baal AE, Stam AT (2013) Phytoseiid predatory mite releasing system and method for production. WO Patent WO/2013/043050
Brown JK, Frohlich DR, Rosell RC (1995) The sweetpotato or silverleaf whiteflies: Biotypes of Bemisia tabaci or a species complex? Ann Rev Entomol 40:511–534
Buitenhuis R, Shipp L, Scott-Dupree C (2010) Dispersal of Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae) on potted greenhouse chrysanthemum. Biol Control 52:110–114
Buitenhuis R, Murphy G, Shipp L, Scott-Dupree C (2015) Amblyseius swirskii in greenhouse production systems: a floriculture perspective. Exp Appl Acarol. doi:10.1007/s10493-014-9869-9
Cahill M, Gorman K, Day S, Denholm I, Elbert A, Nauen R (1996a) Baseline determination and detection of resistance to imidacloprid in Bemisia tabaci (Homoptera: Aleyrodidae). Bull Entomol Res 86:343–349
Cahill M, Jarvis W, Gorman K, Denholm I (1996b) Resolution of baseline response and documentation of resistance to buprofezin in Bemisia tabaci (Hom.: Aleyrodidae). Bull Entomol Res 86:117–122
Calvo FJ, Belda JE (2007) Amblyseius swirskii, un depredador para el control de mosca blanca y trips en cultivos hortícolas. Phytoma España 190:58–62
Calvo J, Urbaneja A (2004) Nesidiocoris tenuis, un aliado para el control biológico de la mosca blanca. Hortic Int 44:20–25
Calvo J, Peña A, Belda JE (2007) Efectos secundarios de productos fitosanitarios sobre el ácaro depredador Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae). In: Proceedings V Congreso Nacional de Entomología Aplicada, Cartagena, Spain
Calvo FJ, Bolkmans K, Belda JE (2008) Controlling the tobacco whitefly Bemisia tabaci (Genn.) (Hom.: Aleyrodidae) in horticultural crops with the predatory mite Amblyseius swirskii (Athias-Henriot). J Insect Sci 8:4
Calvo FJ, Bolckmans K, Belda JE (2009) Development of a biological control-based IPM method for Bemisia tabaci for protected sweet pepper crops. Entomol Exp Appl 133:9–18
Calvo FJ, Bolckmans K, Belda JE (2011) Control of Bemisia tabaci and Frankliniella occidentalis in cucumber by Amblyseius swirskii. Biocontrol 56:185–192
Calvo FJ, Bolckmans K, Belda JE (2012) Biological control-based IPM in sweet pepper greenhouses using Amblyseius swirskii (Acari: Phytoseiidae). Biocontrol Sci Technol 22:1398–1416
Chow A, Chau A, Heinz KM (2010) Compatibility of Amblyseius (Typhlodromips) swirskii (Athias-Henriot) (Acari: Phytoseiidae) and Orius insidiosus (Hemiptera: Anthocoridae) for biological control of Frankliniella occidentalis (Thysanoptera: Thripidae) on roses. Biol Control 53:188–196
Cock MJW, van Lenteren JC, Brodeur J, Barratt BIP, Bigler F, Bolckmans K, Consoli FI, Haas F, Mason PG, Parra JRP (2010) Do new access and benefit sharing procedures under the convention on biological diversity threaten the future of biological control? Biocontrol 55:199–218
CVUA Stuttgart (2007) Residues of the unauthorized insecticide isofenphos-methyl detected in sweet peppers from Spain. http://www.cvuas.de/pub/beitrag.asp?ID=646&subid=1&Thema_ID=5 (accessed 20 Oct 2014)
De Barro PJ, Liu SS, Boykin LM, Dinsdale AB (2011) Bemisia tabaci: a statement of species status. Ann Rev Entomol 56:1–19
Demite PR, de Moraes GJ, McMurtry JA, Denmark HA, Castilho R de C (2014) Phytoseiidae database. www.lea.esalq.usp.br/phytoseiidae (accessed 20 Oct 2014)
Dogramaci M, Arthurs SP, Chen J, McKenzie C, Irrizary F, Osborne L (2011) Management of chilli thrips Scirtothrips dorsalis (Thysanoptera: Thripidae) on peppers by Amblyseius swirskii (Acari: Phytoseiidae) and Orius insidiosus (Hemiptera: Anthocoridae). Biol Control 59:340–347
El-Laithy AYM, Fouly AH (1992) Life table parameters of two phytoseiid predators Amblyseius scutalis (Athias-Henriot) and A. swirskii Athias-Henriot. J Appl Entomol 113:8–12
Espinosa PJ, Bielza P, Contreras J, Lacasa A (2002) Insecticide resistance in field populations of Frankliniella occidentalis (Pergande) in Murcia (south-east Spain). Pest Manag Sci 58:967–971
Fernández E, Gravalos C, Haro PJ, Cifuentes D, Bielza P (2009) Insecticide resistance status of Bemisia tabaci Q-biotype in south-eastern Spain. Pest Manag Sci 65:885–891
Fidgett MJ, Stinson CSA (2008) Method for rearing predatory mites. WO Patent WO/2008/015393
Fouly AH, Al-Deghairi MA, Abdel Baky NF (2011) Biological aspects and life tables of Typhlodromips swirskii (Acari: Phytoseiidae) fed Bemisia tabaci (Hemiptera: Aleyroididae). J Entomol 8:52–62
Fransen JJ (1994) Bemisia tabaci in the Netherlands; here to stay? Pestic Sci 42:129–134
Frutas y Hortalizas (2014) Anuario Hortícola 2014. Frutas y Hortalizas, Almeria
Gerling D, Mayer RT (eds) (1996) Bemisia: 1995. Taxonomy, biology, damage, control and management. Intercept, Andover
Gerson U, Weintraub PG (2012) Mites (Acari) as a factor in greenhouse management. Ann Rev Entomol 57:229–247
Glass R, Gonzalez FJE (2012) Biological control in the greenhouses of Almeria and challenges for a sustainable intensive production. Outlooks Pest Manag 2012:276–278
Goleva I, Zebitz CPW (2013) Suitability of different pollen as alternative food for the predatory mite Amblyseius swirskii (Acari, Phytoseiidae). Exp Appl Acarol 61:259–283
Gradish AE, Scott-Dupree CD, Shipp L, Harris CR, Ferguson G (2011) Effect of reduced risk pesticides on greenhouse vegetable arthropod biological control agents. Pest Manag Sci 67:82–86
Hoda FM, El-Naggar ME, Taha AH, Ibrahim GA (1986) Effect of different types of food on fecundity of predaceous mite Amblyseius swirskii Athias-Henriot (Acari: Phytoseiidae). Bull Soc Entomol Egypt 66:113–116
Hoogerbrugge H, Calvo J, van Houten Y, Bolckmans K (2005) Biological control of the tobacco whitefly Bemisia tabaci with the predatory mite Amblyseius swirskii in sweet pepper crops. IOBC WPRS Bull 28(1):119–122
Hoogerbrugge H, Oude Lenferink K, van Houten Y, Bolckmans K (2014) Screening of three phytoseiid mite species as biocontrol agents of Echinothrips americanus. IOBC WPRS Bull 102:97–101
Ji J, Lin T, Zhang Y, Lin J, Sun L, Chen X (2013) A comparison between Amblyseius (Typhlodromips) swirskii and Amblyseius eharai with Panonychus citri (Acari: Tetranychidae) as prey: developmental duration, life table and predation. Syst Appl Acarol 18:123–129
Juan-Blasco M, Qureshi JA, Urbaneja A, Stansly P (2012) Predatory mite, Amblyseius swirskii (Acari: Phytoseiidae), for biological control of Asian Citrus Psyllid, Diaphorina citri (Hemiptera: Psyllidae). Fla Entomol 95:543–551
Knapp M, van Houten Y, Hoogerbrugge H, Bolckmans K (2013) Amblydromalus limonicus (Acari: Phytoseiidae) as a biocontrol agent: literature review and new findings. Acarologia 53:191–202
Krautter M (2007) Essen ohne Pestizide. Greenpeace, Hamburg
Kumar V, Wekesa VW, Avery PB, Powell CA, McKenzie CL, Osborne LS (2014) Effect of pollens of various ornamental pepper cultivars on the development and reproduction of Amblyseius swirskii (Acari: Phytoseiidae). Fla Entomol 97:367–373
Lee H, Gillespie DR (2011) Life tables and development of Amblyseius swirskii (Acari: Phytoseiidae) at different temperatures. Exp Appl Acarol 53:17–27
Merino-Pacheco M (2007) Almeria finally forced to turn green. Fruit Veg Technol 7(1):23–25
Messelink G, van Steenpaal S, van Wensveen W (2005) Typhlodromips swirskii (Athias-Henriot) (Acari: Phytoseiidae): a new predator for thrips control in greenhouse cucumber. IOBC WPRS Bull 28(1):183–186
Messelink GJ, van Steenpaal SEF, Ramakers PMJ (2006) Evaluation of phytoseiid predators for control of western flower thrips on greenhouse cucumber. Biocontrol 51:753–768
Messelink GJ, van Maanen R, van Steenpaal SEF, Janssen A (2008) Biological control of thrips and whiteflies by a shared predator: two pests are better than one. Biol Control 44:372–379
Messelink GJ, van Maanen R, van Holstein-Saj R, Sabelis MW, Janssen A (2010) Pest species diversity enhances control of spider mites and whiteflies by a generalist phytoseiid predator. Biocontrol 55:387–398
Messelink GJ, Bloemhard CMJ, Cortes JA, Sabelis MW, Janssen A (2011) Hyperpredation by generalist predatory mites disrupts biological control of aphids by the aphidophagous gall midge Aphidoletes aphidimyza. Biol Control 57:246–252
Metwally AM, Abou-El-Naga MM, Taha HA, Hoda FM (1984) Studies on feeding, reproduction and development of Amblyseius swirskii (A.H.) (Acarina: Phytoseiidae). Agric Res Rev 62:323–326
Midthassel A, Leather SR, Baxter IH (2013) Life table parameters and capture success ratio studies of Typhlodromips swirskii (Acari: Phytoseiidae) to the factitious prey Suidasia medanensis (Acari: Suidasiidae). Exp Appl Acarol 61:69–78
Midthassel A, Leather SR, Wright DJ, Baxter IH (2014) The functional and numerical response of Typhlodromips swirskii (Acari: Phytoseiidae) to the factitious prey Suidasia medanensis (Acari: Suidasiidae) in the context of a breeding sachet. Biocontrol Sci Technol 24:361–374
Momen FM, Abdel-Khalek A (2008) Effect of the tomato russet mite Aculops lycoeprsici (Acari: Eriophyidae) on the development and reproduction of three predatory phytoseiid mites. Int J Trop Insect Sci 28:53–57
Momen FM, El-Saway SA (1993) Biology and feeding behavior of the predatory mite, Amblyseius swirskii (Acari: Phytoseiidae). Acarologia 34:199–204
Murphy G (2014) Grower adoption of biological control in greenhouse ornamentals and the role of technology transfer. IOBC WPRS Bull 102:163–167
Nauen R, Denholm I (2005) Resistance of insect pests to neonicotinoid insecticides: current status and future prospects. Arch Insect Biochem Physiol 58:200–215
Nguyen DT, Vangansbeke D, Lü X, De Clercq P (2013) Development and reproduction of the predatory mite Amblyseius swirskii on artificial diets. Biocontrol 58:369–377
Nguyen DT, Vangansbeke D, De Clercq P (2014a) Artificial and factitious foods support the development and reproduction of the predatory mite Amblyseius swirskii. Exp Appl Acarol 62:181–194
Nguyen DT, Vangansbeke D, De Clercq P (2014b) Solid artificial diets for the phytoseiid predator Amblyseius swirskii. Biocontrol. doi:10.1007/s10526-014-9607-6
Nomikou M, Janssen A, Schraag R, Sabelis MW (2001) Phytoseiid predators as potential biological control agents for Bemisia tabaci. Exp Appl Acarol 25:271–291
Nomikou M, Janssen A, Schraag R, Sabelis MW (2002) Phytoseiid predators suppress populations of Bemisia tabaci on cucumber plants with alternative food. Exp Appl Acarol 27:57–68
Nomikou M, Janssen A, Sabelis MW (2003) Phytoseiid predators of whiteflies feed and reproduce on non-prey food sources. Exp Appl Acarol 31:15–26
Nomikou M, Sabelis MW, Janssen A (2010) Pollen subsidies promote whitefly control through the numerical response of predatory mites. Biocontrol 55:253–260
Onzo A, Houedokoho AF, Hanna R (2012) Potential of the predatory mite, Amblyseius swirskii to suppress the broad mite, Polyphagotarsonemus latus on the gboma eggplant, Solanum macrocarpon. J Insect Sci 12(7):1–11
Opit GP, Nechols JR, Margolies DC, Williams KA (2005) Survival, horizontal distribution, and economics of releasing predatory mites (Acari: Phytoseiidae) using mechanical blowers. Biol Control 33:344–351
Park H, Shipp L, Buitenhuis R (2010) Predation, development, and oviposition by the predatory mite Amblyseius swirskii (Acari: Phytoseiidae) on tomato russet mite (Acari: Eriophyidae). J Econ Entomol 103:563–569
Park H, Shipp L, Buitenhuis R, Ahn JJ (2011) Life history parameters of a commercially available Amblyseius swirskii (Acari: Phytoseiidae) fed on cattail (Typha latifolia) pollen and tomato russet mite (Aculops lycopersici). J Asia-Pac Entomol 14:497–501
Peña JE, Rodrigues JCV, Roda A, Carrillo D, Osborne LS (2009) Predator-prey dynamics and strategies for control of the red palm mite (Raoiella indica) (Acari: Tenuipalpidae) in areas of invasion in the Neotropics. IOBC WPRS Bull 50:69–79
Pezzi F, Martelli R, Lanzoni A, Maini S (2015) Effects of mechanical distribution on survival and reproduction of Phytoseiulus persimilis and Amblyseius swirskii. Biosyst Eng 129:11–19
Porath A, Swirski E (1965) A survey of phytoseiid mites (Acarina: Phytoseiidae) on citrus with a description of one new species. Isr J Agric Res 15:87–100
Ragusa S, Swirski E (1975) Feeding habits, development and oviposition of the predacious mite Amblyseius swirskii Athias-Henriot (Acarina: Phytoseiidae) on pollen of various weeds. Isr J Entomol 10:93–103
Ragusa S, Swirski E (1977) Feeding habits, post-embryonic and adult survival, mating, virility and fecundity of the predacious mite Amblyseius swirskii (Acarina: Phytoseiidae) on some coccids and mealybugs. Entomophaga 22:383–392
Skirvin D, Fenlon J (2003) Of mites and movement: the effects of plant connectedness and temperature on movement of Phytoseiulus persimilis. Biol Control 27:242–250
Stansly PA, Castillo J (2009) Control of broad mite, Polyphagotarsonemus latus and the whitefly Bemisia tabaci in open field pepper and eggplant with predaceous mites. IOBC WPRS Bull 49:145–152
Stansly PA, Sánchez PA, Rodríguez JM, Cañizares F, Nieto A, López MJ, Fajardo M, Suárez V, Urbaneja A (2004) Prospects for biological control of Bemisia tabaci (Homoptera, Aleyrodidae) in greenhouse tomatoes of Southern Spain. Crop Prot 23:701–712
Stansly PA, Calvo FJ, Urbaneja A (2005) Augmentative biological control of Bemisia tabaci biotype “Q” in Spanish greenhouse pepper production using Eretmocerus spp. Crop Prot 24:829–835
Swirski E, Amitai S (1997) Annotated list of phytoseiid mites (Mesostigmata: Phytoseiidae) in Israel. Isr J Entomol 31:21–46
Swirski E, Amitai S, Dorzia N (1967) Laboratory studies on the feeding, development and oviposition of the predaceous mites Amblyseius rubini Swirski and Amitai and Amblyseius swirskii Athias-Henriot (Acarina: Phytoseiidae) on various kinds of food substances. Isr J Agric Res 17:101–119
Tal C, Coll M, Weintraub PG (2007) Biological control of Polyphagotarsonemus latus (Acari: Tarsonemidae) by the predaceous mite Amblyseius swirskii (Acari: Phytoseiidae). IOBC WPRS Bull 30(5):111–115
Teich Y (1966) Mites of the family of Phytoseiidae as predators of the tobacco whitefly, Bemisia tabaci Gennadius. Isr J Agric Res 16:141–142
van der Blom J (2005) Control biológico en cultivos hortícolas bajo abrigo. Horticultura 189:10–17
van der Blom J, Robledo A, Torres S, Sánchez JA (2008) Control biológico de plagas en Almería: revolución verde después de dos décadas. Phytoma España 198:42–48
Van Driesche RG, Heinz KM (2004) An overview of biological control in protected culture. In: Heinz KM, Van Driesche RG, Parrella MP (eds) Biocontrol in protected culture. Ball Publishing Batavia, Illinois, pp 1–24
van Houten YM, van Rijn PCJ, Tanigoshi LK, van Stratum P (1993) Potential of phytoseiid predators to control western flower thrips in greenhouse crops, in particular during the winter period. IOBC WPRS Bull 16(8):98–101
van Houten YM, van Rijn PCJ, Tanigoshi LK, van Stratum P, Bruin J (1995) Preselection of predatory mites to improve year-round biological control of western flower thrips in greenhouse crops. Entomol Exp Appl 74:225–234
van Houten Y, Østlie ML, Hoogerbrugge H, Bolckmans K (2005) Biological control of western flower thrips on sweet pepper using the predatory mites Amblyseius cucumeris, Iphiseius degenerans, A. andersoni and A. swirskii. IOBC WPRS Bull 28(1):283–286
van Houten YM, Hoogerbrugge H, Bolckmans KJF (2007a) Spider mite control by four phytoseiid species with different degrees of polyphagy. IOBC WPRS Bull 30(5):123–127
van Houten YM, Hoogerbrugge H, Bolckmans KJF (2007b) The influence of Amblyseius swirskii on biological control of two-spotted spider mites with the specialist predator Phytoseiulus persimilis (Acari: Phytoseiidae). IOBC WPRS Bull 30(5):129–132
van Houten YM, Glas JJ, Hoogerbrugge H, Rothe J, Bolckmans KJF, Simoni S, van Arkel J, Alba JM, Kant MR, Sabelis MW (2013) Herbivory-associated degradation of tomato trichomes and its impact on biological control of Aculops lycopersici. Exp Appl Acarol 60:127–138
van Lenteren JC (2012) The state of commercial augmentative biological control: plenty of natural enemies, but a frustrating lack of uptake. Biocontrol 57:1–20
van Lenteren JC, Bueno VHP (2003) Augmentative biological control of arthropods in Latin America. Biocontrol 48:123–139
van Lenteren JC, Martin NA (1999) Biological control of whitefly. In: Albajes R, Gullino ML, van Lenteren JC, Elad Y (eds) Integrated pest and disease management in greenhouse crops. Kluwer, Dordrecht, pp 202–216
van Maanen R, Vila E, Sabelis MW (2010) Biological control of broad mites (Polyphagotarsonemus latus) with the generalist predator Amblyseius swirskii. Exp Appl Acarol 52:29–34
van Maanen R, Messelink GJ, van Holstein-Saj R, Sabelis MW, Janssen A (2012) Prey temporarily escape from predation in the presence of a second prey species. Ecol Entomol 37:529–535
Weintraub P, Pivonia S, Steinberg S (2011) How many Orius laevigatus are needed for effective western flower thrips, Frankliniella occidentalis, management in sweet pepper? Crop Prot 30:1443–1448
Wimmer D, Hoffmann D, Schausberger P (2008) Prey suitability of western flower thrips, Frankliniella occidentalis, and onion thrips, Thrips tabaci, for the predatory mite Amblyseius swirskii. Biocontrol Sci Technol 18:533–542
Xiao Y, Avery P, Chen J, McKenzie C, Osborne L (2012a) Ornamental pepper as banker plants for establishment of Amblyseius swirskii (Acari: Phytoseiidae) for biological control of multiple pests in greenhouse vegetable production. Biol Control 63:279–286
Xiao YF, Osborne LS, Chen JJ, McKenzie CL (2012b) Functional responses and prey-stage preferences of a predatory gall midge and two predacious mites with twospotted spider mites, Tetranychus urticae, as host. J Insect Sci 13(8):1–12
Xu X, Enkegaard A (2010) Prey preference of the predatory mite, Amblyseius swirskii between first instar western flower thrips Frankliniella occidentalis and nymphs of the twospotted spider mite Tetranychus urticae. J Insect Sci 10(149):1–11
Yousef AEA, El-Keifl AH, Metwally AM (1982) Zur Wirkung von Temperatur und Photoperiode auf die Entwicklung, Ernährung und Eiablage der Raubmilbe Amblyseius swirskii Ath.-Henr. (Acari, Gamasida, Phytoseiidae). Anz Schädlingsk Pflanzensch Umweltsch 55:107–109
Zannou ID, Hanna R (2011) Clarifying the identity of Amblyseius swirskii and Amblyseius rykei (Acari: Phytoseiidae): are they two distinct species or two populations of one species? Exp Appl Acarol 53:339–347
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Calvo, F.J., Knapp, M., van Houten, Y.M. et al. Amblyseius swirskii: What made this predatory mite such a successful biocontrol agent?. Exp Appl Acarol 65, 419–433 (2015). https://doi.org/10.1007/s10493-014-9873-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10493-014-9873-0