
Abstract
Expanding the collection of Planctomycetes by characterisation of novel species is key to better understanding of their complex lifestyles, uncommon cell biology and unexplored metabolism. Here, we isolated three novel planctomycetal strains from a kelp forest on the California Coastline at Monterey Bay or from plastic surfaces submerged in the Baltic Sea and the estuary of the river Warnow in the northeast of Germany. According to our phylogenetic analysis, the isolated strains Poly21T, Pla100T and CA85T represent three novel species within the genus Rhodopirellula. All three show typical planctomycetal traits such as division by budding. All are aerobic, mesophilic chemoheterotrophs and show genomic features comparable to other described Rhodopirellula species. However, strain CA85T is exceptional as it forms cream colonies, but no aggregates, which is a notable deviation from the pink- to red-pigmented and aggregate-forming Rhodopirellula species known thus far. We propose the names Rhodopirellula heiligendammensis sp. nov., Rhodopirellula pilleata sp. nov., and Rhodopirellula solitaria sp. nov. for the novel strains Poly21T (DSM 102266T = LMG 29467T = CECT 9847T = VKM B-3435T), Pla100T (DSM 102937T = LMG 29465T) and CA85T (DSM 109595T = LMG 29699T = VKM B-3451T), respectively, which we present as the respective type strains of these novel species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Planctomycetes are ubiquitous microorganisms often found in aquatic environments. They are Gram-negative bacteria that belong to the PVC (Planctomycetes–Verrucomicrobia–Chlamydiae) superphylum (Spring et al. 2016; Wagner and Horn 2006). The phylum Planctomycetes itself is divided into the classes Phycisphaerae and Planctomycetia (and Candidatus Brocadiae). The order Planctomycetales is currently the only validly described order within the class Planctomycetia and is further subdivided into three families, namely Planctomycetaceae, Isosphaeraceae and Gemmataceae, to which the vast majority of described planctomycetal species belong.
In order to survive in oligotrophic ‘deserts’ such as seawater, species of the family Planctomycetaceae have been shown to attach to many different biotic surfaces (Bengtsson et al. 2012; Bondoso et al. 2015, 2014, 2017; Vollmers et al. 2017). For this purpose, many perform a lifestyle switch between planktonic swimming daughter and sessile mother cells (Jogler et al. 2011). In particular, species of the genus Rhodopirellula are believed to be potent degraders of polymeric sugars released from algal surfaces (Jeske et al. 2013; Wegner et al. 2013). A large set of glycolytic enzymes such as sugar hydrolases, esterases, deacylases and sulfatases encoded in their genomes supports the hypothesis that such species are capable of using naturally-occurring complex sugars or other highly decorated compounds as carbon and energy sources (Ivanova et al. 2017; Wallner et al. 2005; Wegner et al. 2013). Unique pili-forming crateriform structures and an enlarged periplasm are likely involved in the uptake and probably also cleavage of such compounds (Boedeker et al. 2017). Such a unique uptake mechanism allows the intracellular degradation of high molecular weight carbon sources. This could be a decisive advantage compared to extracellular cleavage by secreted enzymes, a strategy that would provide easily accessible carbon sources also to competing microorganisms occupying the same ecological niches (Frank et al. 2014; Wiegand et al. 2018).
Several members of the family Planctomycetaceae are predominant in biofilms on marine biotic surfaces (Bengtsson and Øvreås 2010; Kohn et al. 2019), which is rather unexpected when considering that many planctomycetal strains grow considerably slower than most of their natural competitors. Earlier studies revealed the presence of several Rhodopirellula species in European coastal sediments (Žure et al 2015). Sediments close to Sylt Island in the North Sea were investigated, a location which is quite close to the isolation source of Rhodopirellula baltica, the first Rhodopirellula species, isolated from brackish water of Kiel Fjord in the Baltic Sea (Schlesner et al. 2004). In addition to the specialised machinery for the uptake of polymeric compounds, production of metabolites with anti-microbial or anti-oxidative activity might also explain the observed planctomycetal predominance (Kallscheuer et al. 2019c). This notion is supported by their large genomes, e.g. of Rhodopirellula species, and predicted gene clusters involved in small molecule production (Graca et al. 2016; Jeske et al. 2016). The presence of unusual giant genes might correlate with small molecule production as well (Faria et al. 2018; Kohn et al. 2016; Wiegand et al. 2019).
Not only metabolically, but also structurally, Planctomycetes have been found to possess exceptional traits compared to canonical bacteria. Many of those, such as the lack of peptidoglycan (König et al. 1984), a compartmentalised cell plan (Lindsay et al. 1997), a nucleus-like structure (Fuerst and Webb 1991) and performance of endocytosis (Lonhienne et al. 2010) did however not prove to be verifiable. With the exception of Candidatus Brocadiaceae (Jogler 2014; Neumann et al. 2014), the proposed compartments turned out to be invaginations of the cytoplasmic membrane (Acehan et al. 2013; Boedeker et al. 2017). Also, the presence of peptidoglycan in Planctomycetes was confirmed (Jeske et al. 2015; van Teeseling et al. 2015). Such findings contributed to the reinterpretation of the cell plan of Planctomycetes as Gram-negative (Boedeker et al. 2017; Devos 2014). Nonetheless, Planctomycetes remain exceptional in comparison to well-characterised bacteria of other phyla, e.g. due to the lack of canonical divisome proteins including FtsZ (Jogler et al. 2012; Pilhofer et al. 2008) and an unusual mode of division, either by budding, binary fission or even alternation of both modes (Wiegand et al. 2018, 2019). The unique and highly diverse biology of Planctomycetes is a driving force for expanding the collection of planctomycetal strains, which is prerequisite for getting a better insight into their physiology.
Here, we describe three novel planctomycetal strains isolated from a kelp forest on the California Coastline at Monterey Bay or from plastic surfaces submerged in the Baltic Sea and the estuary of the river Warnow in the northeast of Germany.
Materials and methods
Isolation of the novel strains
The strains were isolated as described before (Wiegand et al. 2019). Briefly, for isolation of Poly21T and Pla100T, incubators containing polyethylene particles were stored for two weeks at different locations in Northern Germany: Poly21T was isolated on 8 October 2015 from an incubator placed at a depth of 2 m below Heiligendamm pier (Seebrücke Heiligendamm) (exact location 54.146 N 11.843 E). Pla100T was isolated on 4 September 2014 from an incubator placed in the Unterwarnow, the estuary of the Warnow river located in the city of Rostock (sampling location 54.106 N 12.096 E), close to a wastewater treatment plant discharge. The exact setup of sampling was described in detail earlier (Oberbeckmann et al. 2018). CA85T was isolated on 28 November 2014 from leaves of a giant bladder kelp (Macrocystis pyrifera) on the Californian coastline in Monterey Bay, CA, USA (36.619 N 121.901 W). Kelp pieces were initially washed with 100 mg/L cycloheximide dissolved in sterile-filtered natural seawater to prevent fungal growth, then swabbed over M1H N-acetyl glucosamine (NAG) artificial seawater (ASW) media plates containing 8 g/L gellan gum, 1000 mg/L streptomycin, 200 mg/L ampicillin and 20 mg/L cycloheximide and incubated at 20 °C for two to three weeks.
Cultivation conditions
After isolation, the strains were cultivated in M1 medium with 4-(2-hydroxyethyl)-1-piperazineethanesulfonic acid (HEPES) as buffering agent and additionally supplemented with ASW and NAG. Designation of the final medium is M1H NAG ASW and it was prepared as previously described (Kallscheuer et al. 2019a).
Light and electron microscopy
Phase contrast (Phaco) analyses and field emission scanning electron microscopy were performed according to a previously published protocol (Kallscheuer et al. 2019a).
Genome information
Genome information of the three isolated strains is available from GenBank under accession numbers SJPU00000000 (Poly21T), SJPM00000000 (Pla100T) and SJPK00000000 (CA85T). 16S rRNA gene sequences are available from GenBank under accession numbers MK554550 (Poly21T), MK554546 (Pla100T) and MK554540 (CA85T). Methods used for DNA isolation and genome sequencing were described before (Wiegand et al. 2019).
Phylogenetic analysis
16S rRNA gene phylogeny was computed for strains Pla100T, CA85T and Poly21T, the type strains of all described planctomycetal species (as of May 2019) and all isolates recently published (Wiegand et al. 2019) and validly named (Boersma et al. 2019; Kallscheuer et al. 2019a, b, d; Kohn et al. 2019). The 16S rRNA gene sequences were aligned with SINA (Pruesse et al. 2012) and the phylogenetic inference was calculated with RAxML (Stamatakis 2014) with a maximum likelihood (ML) approach with 1000 bootstraps, nucleotide substitution model GTR, gamma distributed rate variation and estimation of proportion of invariable sites (GTRGAMMAI option). Three 16S rRNA genes of bacterial strains from the PVC superphylum were used as outgroup. The average nucleotide identity (ANI) was calculated using OrthoANI (Lee et al. 2016). Average amino acid identity (AAI) was calculated using the aai.rb script of the enveomics collection (Rodriguez-R and Konstantinidis 2016) and the percentage of conserved proteins (POCP) was calculated as described (Qin et al. 2014). The rpoB nucleotide sequences were taken from the above-mentioned genomes as well as other publicly available genome annotations and the sequence identities were determined as described (Bondoso et al. 2013). Upon extracting only those parts of the sequence that would have been sequenced with the described primer set the alignment and matrix calculation was performed with Clustal Omega (Sievers et al. 2011). For the multi-locus sequence analysis (MLSA) the unique single-copy core genome of all analysed genomes was determined with proteinortho5 (Lechner et al. 2011) with the ‘selfblast’ option enabled. The protein sequences of the resulting orthologous groups were aligned using MUSCLE v.3.8.31 (Edgar 2004). After clipping, partially aligned C- and N-terminal regions and poorly aligned internal regions were filtered using Gblocks (Castresana 2000). The final alignment was concatenated and clustered using the maximum likelihood method implemented by RaxML (Stamatakis 2014) with the "rapid bootstrap" method and 500 bootstrap replicates.
Results and discussion
Phylogenetic analysis
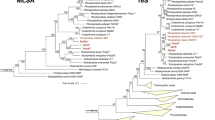
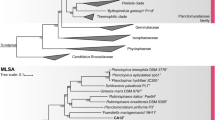
The phylogenetic inference of the novel isolates shows that the three strains Poly21T, Pla100T and CA85T cluster monophyletically within the recently emended genus Rhodopirellula (Kallscheuer et al. 2019c) (Fig. 1). The 16S rRNA gene identity of the three isolated strains and the type species R. baltica SH1T lies between 95.7% and 96.3%, which is above the genus threshold of 94.5% (Yarza et al. 2014) (Table 1, Fig. 2). The indicated proposal of the novel isolates belonging to the Rhodopirellula genus is also supported by POCP values > 50% against R. baltica SH1T (Qin et al. 2014) (Table 1). The AAI analysis gave values of 58.7–59.2% (Table 1). These values are somewhat below the proposed AAI genus threshold of 60–80%, but nevertheless well within the ranges found for other bacterial genera (Luo et al. 2014), thereby also allowing the novel isolates an affiliation to the genus Rhodopirellula.

Maximum likelihood phylogenetic analysis. Phylogenetic trees showing the position of the three novel strains Pla100T, CA85T and Poly21T. Recently described species are included (Kallscheuer et al. 2019b, d). 16S rRNA gene- and MLSA-based phylogeny was computed as described in the Material and Methods section. The outgroup consists of three 16S rRNA genes from the PVC superphylum. In the MLSA-based tree Bythopirellula goksoyri Pr1d served as outgroup. The Gimesia clade includes species of the genera Gimesia, Planctopirus, Fuerstiella, Schlesneria, Rubinisphaera and Planctomicrobium, while the thermophilic clade includes species of the genus Thermostilla, Thermogutta and Thermopirellula

Similarity values of the novel isolates Poly21T, Pla100T and CA85T in relation to type species Rhodopirellula baltica SH1T. The black font gives the 16S rRNA gene identity, the orange font gives the rpoB sequence identity and the turquoise font the whole genome-based average nucleotide identity (ANI)
The last criterion for the affiliation to the genus Rhodopirellula is the rpoB sequence identity of Poly21T, Pla100T and CA85T compared to R. baltica SH1T of 79.8%, 82.0% and 79.0%, respectively. Values in this range are above the genus threshold (Kallscheuer et al. 2019d) (Table 1). The rpoB sequence identities < 95.5% (Fig. 1 and Table 1) also confirm that the novel isolates form separate species within the genus Rhodopirellula. The same can be shown by the comparison of the ANI, as all values are clearly below the species threshold of 95–96% (Kim et al. 2014) and by 16S rRNA gene identities below the species threshold of 98.7% (Stackebrandt and Ebers 2006) (Fig. 2).
Genomic characteristics
The genomes of strains Poly21T (7.1 Mb) and CA85T (6.8 Mb) are similar in size, whereas Pla100T has a larger genome (8.5 Mb). The genome sizes are in the same range as for closely related species, e.g. R. baltica (7.1 Mb), Rhodopirellula bahusiensis (7.8 Mb) and Rhodopirellula sallentina (8.2 Mb). Detailed genomic characteristics data for the novel strains in comparison to the type species R. baltica SH1T are summarised in Table 2. The most striking difference when comparing the three isolated strains is the number of tRNAs. Pla100T harbours 114 tRNA genes, while the number is between 40 and 55 in the other two strains and 91 in R. baltica. CA85T harbours two copies of the 16S rRNA-encoding gene, while only single copies are present in the other three strains. The G + C content of the genomes is between 55 and 58%, which is in the same range as observed for previously characterised Rhodopirellula species.
Morphology and physiology
We analysed the cell morphology of strains Poly21T, Pla100T and CA85T employing light microscopy (LM, Fig. 3) and scanning electron microscopy (SEM, Fig. 4). For this purpose, cells were cultivated in M1H NAG ASW medium and harvested during the exponential growth phase.

Light microscopy images and cell size plots of strains Pla100T, CA85T and Poly21T. The mode of cell division (a, c, e) and a general overview of cell morphology (b, d, f) is shown in the pictures. The scale bar is 1 µm. For determination of the cell size (g, h, i) at least 100 representative cells were counted manually or by using a semi-automated object count tool

Scanning electron microscopic pictures of the three novel strains Pla100T, CA85T and Poly21T. The scale bar is 1 µm
Cells of strain Poly21T are pear-shaped to round with an average size of 2.4 ± 0.3 µm in length and 1.9 ± 0.2 µm in width (Fig. 3a, c). Colonies have a coral pink colour. Poly21T cells form loose aggregates. Cell division is performed by polar budding with the daughter cell having the same shape as the mother cell (Fig. 3a). The strain is aerobic and grows at pH values ranging from 5.5 to 9.0 (Fig. 5). An optimal growth at pH 8.5 suggests that strain Poly21T is slightly alkaliphilic. Growth was observed at temperatures ranging from 10 to 33 °C with optimal growth at 30 °C (Fig. 5). In M1H NAG ASW medium a maximal growth rate of 0.064 h−1 was observed, which corresponds to a doubling time of approximately 11 h.

Temperature and pH optima of the isolated strains Pla100T, CA85T and Poly21T. The graphs show the average growth rates obtained from cultivation of the three isolated strains in M1H NAG ASW medium in biological triplicates. Cultivations at different pH values were conducted at 28 °C and cultivations at different temperatures were performed at pH 7.5
Colonies of strain Pla100T have a similar colour as Poly21T but the cell shape is different (Table 2). In contrast to Poly21T, Pla100T cells have an elongated shape, which becomes obvious when comparing the cells sizes of both strains. Although similar in length (Poly21T: 2.4 µm, Pla100T: 2.3 µm), cell width of Pla100T (1.0 µm) is only half of the width of Poly21T (1.9 µm) (Figs. 3c, h, 4c). Strain Pla100T divides by budding with the bud being slimmer than the mother cell. Stronger aggregate formation of Pla100T compared to Poly21T was observed, which is also reflected in the formation of a polar extracellular matrix by strain Pla100T. Pla100T grows at a pH range of 5.5 to 8.5 (optimum 7.0) and a temperature range of 10–33 °C (optimum 30 °C) (Fig. 5). The maximal observed growth rate of Pla100T was 0.033 h−1, which is approximately half of the growth rate of Poly21T, thus the doubling time of Pla100T is 21 h.
The third strain, CA85T, has a cell size of 2.0 µm × 1.3 µm and is slightly smaller than Poly21T, but similar in shape. Although producing an extracellular matrix covering the entire cell surface (Fig. 4f), CA85T did not form aggregates (Fig. 4e). Instead, individual cells were observed during SEM and light microscopy. Cells divide by budding (Fig. 3g). Strain CA85T reached a maximal growth rate of 0.066 h−1 (generation time: 11 h), which is very similar to that obtained for Poly21T. The strain grew in the same pH and temperature range as Pla100T with optimal growth at pH 7.5 and 27 °C (Fig. 5). Some characteristics of CA85T are exceptional compared to other Rhodopirellula species. CA85T forms cream colonies and is the first known Rhodopirellula strain not showing a pink or red pigmentation. Additionally, the lack of aggregate formation is also a major difference to most of the Rhodopirellula species characterised thus far. Basic aspects of morphology and mechanism of cell division for all strains are summarised in Table 2.
Based on the genome sequences we analysed key metabolic capabilities in the primary and secondary metabolism of the three strains in comparison to R. baltica SH1T. In our analysis, we focused on the central carbon metabolism comprising glycolytic pathways, pentose phosphate pathway, citric acid cycle (TCA cycle), gluconeogenesis and anaplerotic reactions, such as the glyoxylate shunt (Table 3). Obtained results suggest that all four strains use the Embden–Meyerhof–Parnas pathway for the degradation of sugars as we were able to assign genes to all required enzymatic reactions of this most common glycolytic pathway. The absence of additional glycolytic pathways, e.g. the Entner–Doudoroff pathway, was reinforced by the absence of 2-dehydro-3-deoxy-phosphogluconate aldolase in all four strains. The pentose phosphate pathway also appears to be functional in all four strains. This was to some extent expected as important precursors for the biosynthesis of L-histidine, the three aromatic amino acids and nucleotides are provided by this pathway. All enzymes of the TCA cycle are encoded, however, growth of Rhodopirellula species with acetate or fatty acids as sole carbon and energy source appears unlikely as all four strains lack the glyoxylate shunt enabling anaplerosis during growth on the above-mentioned carbon sources. Remarkably, key gluconeogenic enzymes also seem to be absent in the four strains. This further supports the prediction that Rhodopirellula species are unable to use acetate or e.g. TCA cycle intermediates as sole carbon and energy source since an incomplete gluconeogenesis would not allow for production of sugars, which are essential for biosynthesis of cell wall components. Taken together, our analysis leads to the hypothesis that the metabolism of the here analysed Rhodopirellula strains is rather adapted to sugars as substrates for biomass formation and does not seem to support good growth with short- or long-chain carboxylic acids.
To gain a first insight into the potential for secondary metabolite biosynthesis an AntiSMASH analysis was performed (Blin et al. 2019). Here, we focused on terpenoid-forming proteins, polyketide synthases (PKS) and non-ribosomal peptide synthetases (NRPS) as these enzyme classes are known to be responsible for production of bioactive small molecules (Table 4). It is likely that Planctomycetes follow other, yet uncharacterised, pathways for secondary metabolite production, however these will most likely escape the AntiSMASH prediction. In the genomes of the three strains isolated here, 6–9 clusters potentially involved in small molecule production were found. This number is comparable to R. baltica SH1T (six clusters). The distribution of the clusters into different types of PKSs, NRPSs, mixed PKS-NRPSs and terpenoid-forming clusters is similar, but not identical. All four species have in common the lack of type II PKSs, and presence of one type III PKS and two terpenoid clusters. R. baltica lacks an NRPS-encoding cluster, while one putative NRPS was identified in the three novel isolates. The number of clusters correlated with the genome size of the strains. The genome of Pla100T is around 20% larger than the genome of the other three strains and this strain also contains the highest number of clusters.
Conclusion
Based on our physiological and phylogenetic analyses, we conclude that the three characterised strains represent three novel species within the genus Rhodopirellula. Thus, we propose the names Rhodopirellula heiligendammensis sp. nov., Rhodopirellula pilleata sp. nov., and Rhodopirellula solitaria sp. nov. with strains Poly21T, Pla100T and CA85T as the respective type strains of these novel species.
Description of Rhodopirellula heiligendammensis sp. nov.
Rhodopirellula heiligendammensis (hei.li.gen.dam.men’sis. N.L. fem. adj. heiligendammensis of Heiligendamm; referring to the origin of the bacterium from Heiligendamm, Germany).
Cells are pear-shaped to round (length: 2.4 ± 0.3 µm, width: 1.9 ± 0.2 µm), form aggregates and divide by polar budding. Cells grow at 10–33 °C (optimum 30 °C) and at pH 5.5–9.0 (optimum 8.5). Colonies are coral pink. The genome sequence (acc. no. SJPU00000000) and 16S rRNA gene sequence (acc. no. MK554550) of the type strain are available from GenBank. The type strain is Poly21T (DSM 102266T = LMG 29467T = CECT 9847T = VKM B-3435T), isolated from polyethylene particles incubated in 2 m depth in the Baltic Sea close to Heiligendamm, Germany in October 2015.
Description of Rhodopirellula pilleata sp. nov.
Rhodopirellula pilleata (pil.le.a’ta. L. fem. adj. pilleata wearing a headgear; referring to the hat-like shape of the fibers).
Cells are elongated pear-shaped (length: 2.3 ± 0.4 µm, width: 1.0 ± 0.2 µm), form aggregates and divide by budding. Colonies are brink pink. The temperature optimum is 30 °C (growth observed from 10–33 °C). Growth is observed at pH 5.5–8.5 (optimum 7.0). The genome sequence (acc. no. SJPM00000000) and 16S rRNA gene sequence (acc. no. MK554546) of the type strain are available from GenBank. The type strain is Pla100T (DSM 102937T = LMG 29465T), isolated from polyethylene particles submerged in the Unterwarnow close to a wastewater treatment plant discharge in Rostock, Germany in September 2014.
Description of Rhodopirellula solitaria sp. nov.
Rhodopirellula solitaria (sol.i.ta’ri.a. L. fem. adj. solitaria lonely; referring to the solitary occurrence of the cells and the absence of aggregates).
Cells are pear-shaped (2.0 ± 0.3 × 1.3 ± 0.2 µm) and divide by budding. Colonies have a cream colour. Preferred temperature and pH are 27 °C and 7.5, respectively, while growth is observed in the range of 10–33 °C and at pH 6.0–8.5. The genome sequence (acc. no. SJPK00000000) and 16S rRNA gene sequence (acc. no. MK554540) of the type strain are available from GenBank. The type strain is CA85T (DSM 109595T = LMG 29699T = VKM B-3451T), isolated from a giant bladder kelp (M. pyrifera) in a kelp forest in Monterey Bay, CA, USA in November 2014.
Emended description of the genus Rhodopirellula Schlesner et al. 2004
The description of the genus Rhodopirellula is as given previously (Schlesner et al. 2004), with the following modification: Colony colours range from cream to pink. Crateriform structures are not always present.
References
Acehan D, Santarella-Mellwig R, Devos DP (2013) A bacterial tubulovesicular network. J Cell Sci 127:277–280
Bengtsson MM, Øvreås L (2010) Planctomycetes dominate biofilms on surfaces of the kelp Laminaria hyperborea. BMC Microbiol 10:261
Bengtsson MM, Sjøtun K, Lanzén A, Øvreås L (2012) Bacterial diversity in relation to secondary production and succession on surfaces of the kelp Laminaria hyperborea. ISME J 6:2188–2198
Blin K, Shaw S, Steinke K, Villebro R, Ziemert N, Lee SY, Medema MH, Weber T (2019) AntiSMASH 5.0: updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res 47:W81–W87
Boedeker C, Schuler M, Reintjes G, Jeske O, van Teeseling MC, Jogler M, Rast P, Borchert D, Devos DP, Kucklick M, Schaffer M, Kolter R, van Niftrik L, Engelmann S, Amann R, Rohde M, Engelhardt H, Jogler C (2017) Determining the bacterial cell biology of Planctomycetes. Nat Commun 8:14853
Boersma A, Kallscheuer N, Wiegand S, Rast R, Peeters S, Mesman R, Heuer A, Boedeker C, Jetten M, Rohde M, Jogler M (2019) Alienimonas californiensis gen. nov. sp. nov., a novel Planctomycete isolated from the kelp forest in Monterey Bay. Antonie van Leeuwenhoek. https://doi.org/10.1007/s10482-019-01367-4
Bondoso J, Harder J, Lage OM (2013) rpoB gene as a novel molecular marker to infer phylogeny in Planctomycetales. Antonie Van Leeuwenhoek 104:477–488
Bondoso J, Balague V, Gasol JM, Lage OM (2014) Community composition of the Planctomycetes associated with different macroalgae. FEMS Microbiol Ecol 88:445–456
Bondoso J, Albuquerque L, Nobre MF, Lobo-da-Cunha A, da Costa MS, Lage OM (2015) Roseimaritima ulvae gen. nov., sp. nov. and Rubripirellula obstinata gen. nov., sp. nov. two novel planctomycetes isolated from the epiphytic community of macroalgae. Syst Appl Microbiol 38:8–15
Bondoso J, Godoy-Vitorino F, Balague V, Gasol JM, Harder J, Lage OM (2017) Epiphytic Planctomycetes communities associated with three main groups of macroalgae. FEMS Microbiol Ecol 93:fiw255
Castresana J (2000) Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol Biol Evol 17:540–552
Devos DP (2014) Re-interpretation of the evidence for the PVC cell plan supports a gram-negative origin. Antonie Van Leeuwenhoek 105:271–274
Edgar RC (2004) MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res 32:1792–1797
Faria M, Bordin N, Kizina J, Harder J, Devos D, Lage OM (2018) Planctomycetes attached to algal surfaces: insight into their genomes. Genomics 110:231–238
Frank O, Michael V, Pauker O, Boedeker C, Jogler C, Rohde M, Petersen J (2014) Plasmid curing and the loss of grip—the 65-kb replicon of Phaeobacter inhibens DSM 17395 is required for biofilm formation, motility and the colonization of marine algae. Syst Appl Microbiol 38:120–127
Fuerst JA, Webb RI (1991) Membrane-bounded nucleoid in the eubacterium Gemmata obscuriglobus. Proc Natl Acad Sci USA 88:8184–8188
Graca AP, Calisto R, Lage OM (2016) Planctomycetes as novel source of bioactive molecules. Front Microbiol 7:1241
Ivanova AA, Naumoff DG, Miroshnikov KK, Liesack W, Dedysh SN (2017) Comparative genomics of four Isosphaeraceae planctomycetes: a common pool of plasmids and glycoside hydrolase genes shared by Paludisphaera borealis PX4T, Isosphaera pallida IS1BT, Singulisphaera acidiphila DSM 18658T, and strain SH-PL62. Front Microbiol 8:412
Jeske O, Jogler M, Petersen J, Sikorski J, Jogler C (2013) From genome mining to phenotypic microarrays: Planctomycetes as source for novel bioactive molecules. Antonie Van Leeuwenhoek 104:551–567
Jeske O, Schüler M, Schumann P, Schneider A, Boedeker C, Jogler M, Bollschweiler D, Rohde M, Mayer C, Engelhardt H, Spring S, Jogler C (2015) Planctomycetes do possess a peptidoglycan cell wall. Nat Commun 6:7116
Jeske O, Surup F, Ketteniß M, Rast P, Förster B, Jogler M, Wink J, Jogler C (2016) Developing techniques for the utilization of Planctomycetes as producers of bioactive molecules. Front Microbiol 7:1242
Jogler C (2014) The bacterial 'mitochondrium'. Mol Microbiol 94:751–755
Jogler C, Glöckner FO, Kolter R (2011) Characterization of Planctomyces limnophilus and development of genetic tools for its manipulation establish it as a model species for the phylum Planctomycetes. Appl Environ Microbiol 77:5826–5829
Jogler C, Waldmann J, Huang X, Jogler M, Glöckner FO, Mascher T, Kolter R (2012) Identification of proteins likely to be involved in morphogenesis, cell division, and signal transduction in Planctomycetes by comparative genomics. J Bacteriol 194:6419–6430
Kallscheuer N, Jogler M, Wiegand S, Peeters S, Heuer A, Boedeker C, Jetten M, Rohde M, Jogler C (2019a) Rubinisphaera italica sp. nov. isolated from a hydrothermal area in the Tyrrhenian Sea close to the volcanic island Panarea. Antonie van Leeuwenhoek. https://doi.org/10.1007/s10482-019-01329-w
Kallscheuer N, Jogler M, Wiegand S, Peeters S, Heuer A, Boedeker C, Jetten M, Rohde M, Jogler C (2019b) Three novel Rubripirellula species isolated from artificial plastic surfaces submerged in the German part of the Baltic Sea and the estuary of the river Warnow. Antonie van Leeuwenhoek. https://doi.org/10.1007/s10482-019-01368-3
Kallscheuer N, Moreira C, Airs R, Llewellyn CA, Wiegand S, Jogler C, Lage OM (2019c) Pink-and orange-pigmented Planctomycetes produce saproxanthin-type carotenoids including a rare C45 carotenoid. Environ Microbiol Rep 11:741–748
Kallscheuer N, Wiegand S, Peeters SH, Jogler M, Boedeker C, Heuer A, Rast P, Jetten MSM, Rohde M, Jogler C (2019d) Description of three bacterial strains belonging to the new genus Novipirellula gen. nov., reclassificiation of Rhodopirellula rosea and Rhodopirellula caenicola and readjustment of the genus threshold of the phylogenetic marker rpoB for Planctomycetaceae. Antonie van Leeuwenhoek (accepted manuscript ANTO-D-19-00304)
Kim M, Oh HS, Park SC, Chun J (2014) Towards a taxonomic coherence between average nucleotide identity and 16S rRNA gene sequence similarity for species demarcation of prokaryotes. Int J Syst Evol Microbiol 64:346–351
Kohn T, Heuer A, Jogler M, Vollmers J, Boedeker C, Bunk B, Rast P, Borchert D, Glöckner I, Freese HM, Klenk HP, Overmann J, Kaster AK, Wiegand S, Rohde M, Jogler C (2016) Fuerstia marisgermanicae gen. nov., sp. nov., an unusual member of the phylum Planctomycetes from the German Wadden Sea. Front Microbiol 7:2079
Kohn T, Wiegand S, Boedeker C, Rast P, Heuer A, Jetten MSM, Schüler M, Becker S, Rohde C, Müller R-W, Brümmer F, Rohde M, Engelhardt H, Jogler M, Jogler C (2019) Planctopirus ephydatiae, a novel Planctomycete isolated from a freshwater sponge. Syst Appl Microbiol. https://doi.org/10.1016/j.syapm.2019.126022
König E, Schlesner H, Hirsch P (1984) Cell wall studies on budding bacteria of the Planctomyces/Pasteuria group and on a Prosthecomicrobium sp. Arch Microbiol 138:200–205
Lechner M, Findeiss S, Steiner L, Marz M, Stadler PF, Prohaska SJ (2011) Proteinortho: detection of (co-)orthologs in large-scale analysis. BMC Bioinform 12:124
Lee I, Ouk Kim Y, Park SC, Chun J (2016) OrthoANI: an improved algorithm and software for calculating average nucleotide identity. Int J Syst Evol Microbiol 66:1100–1103
Lindsay MR, Webb RI, Fuerst JA (1997) Pirellulosomes: a new type of membrane-bounded cell compartment in planctomycete bacteria of the genus Pirellula. Microbiol UK 143:739–748
Lonhienne TG, Sagulenko E, Webb RI, Lee KC, Franke J, Devos DP, Nouwens A, Carroll BJ, Fuerst JA (2010) Endocytosis-like protein uptake in the bacterium Gemmata obscuriglobus. Proc Natl Acad Sci USA 107:12883–12888
Luo C, Rodriguez RL, Konstantinidis KT (2014) MyTaxa: an advanced taxonomic classifier for genomic and metagenomic sequences. Nucleic Acids Res 42:e73
Neumann S, Wessels HJ, Rijpstra WI, Sinninghe Damste JS, Kartal B, Jetten MS, van Niftrik L (2014) Isolation and characterization of a prokaryotic cell organelle from the anammox bacterium Kuenenia stuttgartiensis. Mol Microbiol 94:794–802
Oberbeckmann S, Kreikemeyer B, Labrenz M (2018) Environmental factors support the formation of specific bacterial assemblages on microplastics. Front Microbiol 8:2709
Pilhofer M, Rappl K, Eckl C, Bauer AP, Ludwig W, Schleifer KH, Petroni G (2008) Characterization and evolution of cell division and cell wall synthesis genes in the bacterial phyla Verrucomicrobia, Lentisphaerae, Chlamydiae, and Planctomycetes and phylogenetic comparison with rRNA genes. J Bacteriol 190:3192–3202
Pruesse E, Peplies J, Glöckner FO (2012) SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28:1823–1829
Qin Q-L, Xie B-B, Zhang X-Y, Chen X-L, Zhou B-C, Zhou J, Oren A, Zhang Y-Z (2014) A proposed genus boundary for the prokaryotes based on genomic insights. J Bacteriol 196:2210–2215
Rodriguez-R LM, Konstantinidis KT (2016) The enveomics collection: a toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Preprints 4:e1900v1
Schlesner H, Rensmann C, Tindall BJ, Gade D, Rabus R, Pfeiffer S, Hirsch P (2004) Taxonomic heterogeneity within the Planctomycetales as derived by DNA–DNA hybridization, description of Rhodopirellula baltica gen. nov., sp. nov., transfer of Pirellula marina to the genus Blastopirellula gen. nov. as Blastopirellula marina comb. nov. and emended description of the genus Pirellula. Int J Syst Evol Microbiol 54:1567–1580
Sievers F, Wilm A, Dineen D, Gibson TJ, Karplus K, Li W, Lopez R, McWilliam H, Remmert M, Söding J (2011) Fast, scalable generation of high-quality protein multiple sequence alignments using clustal omega. Mol Syst Biol 7:539
Spring S, Bunk B, Spröer C, Schumann P, Rohde M, Tindall BJ, Klenk H-P (2016) Characterization of the first cultured representative of Verrucomicrobia subdivision 5 indicates the proposal of a novel phylum. ISME J 10:2801
Stackebrandt E, Ebers J (2006) Taxonomic parameter revisited: tarnished gold standards. Microbiol Today 33:152–155
Stamatakis A (2014) RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30:1312–1313
van Teeseling MC, Mesman RJ, Kuru E, Espaillat A, Cava F, Brun YV, Van Nieuwenhze MS, Kartal B, van Niftrik L (2015) Anammox Planctomycetes have a peptidoglycan cell wall. Nat Commun 6:6878
Vollmers J, Frentrup M, Rast P, Jogler C, Kaster AK (2017) Untangling genomes of novel planctomycetal and verrucomicrobial species from monterey bay kelp forest metagenomes by refined binning. Front Microbiol 8:472
Wagner M, Horn M (2006) The Planctomycetes, Verrucomicrobia, Chlamydiae and sister phyla comprise a superphylum with biotechnological and medical relevance. Curr Opin Biotechnol 17:241–249
Wallner SR, Bauer M, Würdemann C, Wecker P, Glöckner FO, Faber K (2005) Highly enantioselective sec-alkyl sulfatase activity of the marine planctomycete Rhodopirellula baltica shows retention of configuration. Angew Chem Int Ed Engl 44:6381–6384
Wegner C-E, Richter-Heitmann T, Klindworth A, Klockow C, Richter M, Achstetter T, Glöckner FO, Harder J (2013) Expression of sulfatases in Rhodopirellula baltica and the diversity of sulfatases in the genus Rhodopirellula. Mar Genomics 9:51–61
Wiegand S, Jogler M, Jogler C (2018) On the maverick Planctomycetes. FEMS Microbiol Rev 42:739–760
Wiegand S, Jogler M, Boedeker C, Pinto D, Vollmers J, Rivas-Marín E, Kohn T, Peeters SH, Heuer A, Rast P, Oberbeckmann S, Bunk B, Jeske O, Meyerdierks A, Storesund JE, Kallscheuer N, Lücker S, Lage OM, Pohl T, Merkel BJ, Hornburger P, Müller R-W, Brümmer F, Labrenz M, Spormann AM, Op den Camp HJM, Overmann J, Amann R, Jetten MSM, Mascher T, Medema MH, Devos DP, Kaster A-K, Øvreås L, Rohde M, Galperin MY, Jogler C (2019) Cultivation and functional characterization of 79 planctomycetes uncovers their unique biology. Nat Microbiol. https://doi.org/10.1038/s41564-019-0588-1
Yarza P, Yilmaz P, Pruesse E, Glöckner FO, Ludwig W, Schleifer KH, Whitman WB, Euzeby J, Amann R, Rossello-Mora R (2014) Uniting the classification of cultured and uncultured bacteria and archaea using 16S rRNA gene sequences. Nat Rev Microbiol 12:635–645
Žure M, Munn CB, Harder J (2015) Diversity of Rhodopirellula and related planctomycetes in a North Sea coastal sediment employing carB as molecular marker. FEMS Microbiol Lett 362:fnv127
Acknowledgements
Part of this research was funded by the Deutsche Forschungsgemeinschaft Grants KA 4967/1-1 and JO 893/4-1, Grant ALWOP.308 of the Nederlandse Organisatie voor Wetenschappelijk Onderzoek (NWO), SIAM (Soehngen Institute for Anaerobic Microbiology) Grant No. 024002002 and the Radboud Excellence fellowship. We thank Ina Schleicher for skillful technical assistance. Brian Tindall and Regine Fähnrich from the DSMZ as well as the BCCM/LMG Bacteria collection we thank for support during strain deposition. We thank Anne-Kristin Kaster (KIT Karlsruhe, Germany) and Alfred M. Spormann (Stanford, USA) as well as the Aquarius Dive Shop Monterey and the Hopkins Marine Station for sampling support. We also thank our collaborators Sonja Oberbeckmann and Matthias Labrenz (IOW Warnemünde, Germany) for support during sampling.
Author information
Authors and Affiliations
Contributions
NK wrote the manuscript and analysed the cultivation data. SW performed the genomic and phylogenetic analysis. AH, PR and MJ isolated the strains and performed the initial cultivation and strain deposition. SHP and CB performed the light microscopic analysis and prepared the LM pictures. MSMJ contributed to text preparation and revised the manuscript. MR performed the electron microscopic analysis and prepared the SEM pictures. CJ took the samples and supervised the study. All authors read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical statement
This article does not contain any studies with animals performed by any of the authors.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Kallscheuer, N., Wiegand, S., Jogler, M. et al. Rhodopirellula heiligendammensis sp. nov., Rhodopirellula pilleata sp. nov., and Rhodopirellula solitaria sp. nov. isolated from natural or artificial marine surfaces in Northern Germany and California, USA, and emended description of the genus Rhodopirellula. Antonie van Leeuwenhoek 113, 1737–1750 (2020). https://doi.org/10.1007/s10482-019-01366-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10482-019-01366-5