
Abstract
This research aims to study the variation in phytosociology and plant diversity of large cardamom-based traditional agroforestry systems along an altitudinal gradient (700–2000 m) in the Darjeeling Himalayas. We analyzed the changes in phytosociology and plant diversity by adopting stratified random nested quadrate sampling method. The agroforestry managers were interviewed for their perception of ecosystem service following Millennium Ecosystem Assessment guidelines. The present study showed altitudinal location significantly influences plant diversity. Overall, 130 plant species were documented, of which 37 were trees, 25 shrubs, 46 herbs, 8 ferns, 11 climbers and 3 orchids. The low-, mid- and high-altitude classes were documented with 76, 60 and 52 plant species, respectively. Overall, the study system was highly heterogeneous and diverse with a higher Shannon and Wiener diversity index of 4.09 which decreased progressively with increasing altitude as evidenced from significant negative relationship of altitude with diversity (r =− 0.582**), species richness (r =− 0.648**) and plant population (r =− 0.587**). Of the total listed plant species, about 68% were data deficit, 29% were least concerned; two species (Cryptomeria japonica and Cupressus cashmeriana) were near threatened, and one species (Brugmansia suaveolens) was extinct in the wild. This indicates that the study system plays a vital role in harbouring and conserving regional plant diversity. The plant species documented were also classified based on their ecosystem services with 120, 47, 34 and 33 species providing provisional, cultural, regulatory and supporting services, respectively.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Traditional agroforestry farming forms a chief component in the livelihoods and economy of the rural poor population (Reang et al. 2022; Roy et al. 2022a), evolved in response to various factors such as social, economic, cultural, and environmental. In the mountains, the traditional agroforestry systems mimic the natural forests and offer similar ecosystem services to those under the forests (Sharma et al. 2007; Vineeta et al. 2021). One such traditional system is large cardamom (Amomum subulatum Roxb.) based agroforestry in the north eastern Indian states, including the Darjeeling Himalayas (Sharma et al. 2016). Large cardamom, the oldest of spices, is native to Sikkim and Darjeeling Himalaya, including the eastern hills of Nepal (Shrestha et al. 2018) and distributed in minimal areas and mainly found in the Eastern Himalayan region of India, Nepal and Bhutan (Mehta et al. 2015). The crop is sciophyte and traditionally inter-mixed as an understorey of natural forest trees on marginal lands and slopes with high moisture in areas of high rainfall between 1500 and 3500 mm at an altitude of 600 and 2000 m above mean sea level (Vineeta et al. 2021). Of the many shade trees associated with the large cardamom, Himalayan Alder (Alnus nepalensis) is prominent because of its nitrogen-fixing capacity (Negi et al. 2018).
In the tropical regions, some traditional agroforestry systems show high floristic diversities compared with other agroecosystems (Reang et al. 2021). However, plant density, species diversity and composition vary from place to place due to ecological and socio-economic factors (Roy et al. 2022b). Locality and topographic factors alter the microclimate and edaphic settings of a site and are accountable for shaping the position of vegetation in a particular habitat. Phytosociological characters vary among aspects and location, even in a similar type of vegetation (Roy et al. 2022b).
The ecosystem services provisioned in the form of food, timber, fiber, medicine, drinking and irrigation water from the Himalayan ecosystems are the source of livelihood to the people residing there (Sharma et al. 2015). In addition, some studies have reported that the mountains' forests and agroforestry systems help stabilize headwaters, prevent flooding, landslide and maintain steady year-round water flows vital for the densely populated downstream areas (Molden et al. 2014). However, the quality and quantity of ecosystem services have decreased due to anthropogenic activities and natural phenomena in the recent years (Sharma and Chettri 2021).
Among the numerous traditional farming systems in Darjeeling and Sikkim Himalayas, the large cardamom-based traditional agroforestry system has gained research attention in recent years due to its associated ecosystem services, which has sustained livelihood and supported economic growth of the farming households in the region (Sharma et al. 2000, 2016; Avasthe et al. 2011; Singh et al. 2018; Tarafder et al. 2018). However, few studies on large cardamom-based agroforestry systems covering phytosociological analysis and biomass production were reported mainly from Sikkim Himalayas (Sharma et al. 2007; 2016). As a result, little has been studied to understand the floristic diversity, structure and composition, and ecosystem services along an altitudinal gradient. Moreover, to date, there is no data available from Darjeeling Himalaya. Therefore, the study was carried out to generate precise and systematic quantitative data on the potential of large cardamom-based agroforestry systems in the Darjeeling Himalayas along the altitudinal gradient for their role in biodiversity conservation and ecosystem services. The specific objectives of this study are (i) to document plant diversity along an altitudinal gradient under the large cardamom-based agroforestry system, (ii) to classify the plant species based on IUCN status along the altitudinal gradient, and (iii) to explore the plant species for their known ecosystem services along the altitudinal gradient based on Millennium Ecosystem Assessment guidelines.
Material and methods
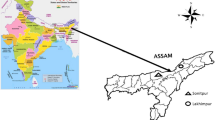
The study was carried out in the Darjeeling Himalayan region of West Bengal, India (Fig. 1) from January 2019 to April 2021 in an altitudinal range of 132–3660 m. The region is humid and sub-tropical to sub-alpine, with an average annual rainfall of 2700–3100 mm (Moktan and Das 2013). A reconnaissance survey was conducted to explore the large cardamom based traditional agroforestry systems. A total of 25 traditional holdings of 1–3 ha size were found during the survey. In this traditional farming, large cardamom is cultivated under the canopy of reserved or protected forest leased out to the growers by the State Forest Department with no rights to cut the trees (Sharma et al. 1994, 2009). Following the altitudinal chronosequence (Moktan and Das 2013; Cajee 2018), three altitudinal classes were considered: low (700–1200 m asl), mid (1200–1700 m asl) and high (> 1700 m asl) with distribution of eleven, nine and five large cardamom agroforestry holdings, respectively. In these large cardamom holdings, 1–4 quadrates were laid for vegetation sampling depending on its size.

Map of the study area showing different sampling points
Phytosociological analysis
In each large cardamom holding, 10 m × 10 m sized quadrats were laid for trees within which two 5 m × 5 m sub-quadrats were laid at diagonal corners for shrubs and five 1 m × 1 m sub-quadrats at four corners and centre of the main quadrat for herbs. Individual plant species were counted species-wise in each quadrant and sub-quadrant while, the plant species richness in each quadrant was also recorded. In addition, the name of the species, family and genera were documented following the Plant List of World Flora Online at www.worldfloraonline.org and www.tropicos.org of Missouri Botanical Garden.
Diversity indices
The number of plant species documented is the species richness (S). Species diversity or Menhinick’s index (Dʹ = S/ √N) is based on species richness (S) in proportion to the total number of individuals of all species (N) (Menhinick 1964). Shannon–Wiener diversity index (Hʹ = ∑ni/N ln ni/N); where ‘ni' is the number of individuals of a species, 'N' is the number of individuals of all species and 'ln' is the natural logarithm (Shannon and Weaver 1949; Shannon and Weiner 1963). Concentration of dominance or Simpson's index (C = ∑ (ni/N) 2 where ‘ni' is the number of individuals of a species and 'N' is the number of individuals of all species documented from the sampled area, and its value is 0–1 (Simpson, 1949). Species evenness index (EI = Hʹ/ln N) indicates the degree of species distribution (Pielou 1975) where 'Hʹ' is the Shannon–Wiener diversity index, and 'ln N' is the natural logarithm of a number of individuals of all species.
Sorenson’s similarity index
This index (SI) estimates the degree of similarity in species among the habitats or plant assemblages, and the equation used following Sorenson (1948) was:
where, A is the number of species unique in area A, B is the number of species unique in area B, C is the number of species unique in area C, and D is the number of species common in all the areas A, B and C.
Conservation status
The conservation status of the documented species was based on the IUCN Red List category. Among the mentioned nine IUCN categories, the documented plant species represented only four categories which are data deficit (DD), least concern (LC), near threatened (NT) and extinct in the wild (EW) (Anonymous 2021).
Ecosystem services
Ecosystem services provided by different species in the traditional large cardamom-based agroforestry system were collected through personal interviews with the agroforestry managers (Lepcha et al. 2018, 2019), and species were then classified following Millennium Ecosystem Assessment guidelines (MEA 2005; Supplementary Table 1). The information was gathered through personal interviews with the holding owners by an enumerator (a local resident) in the presence of any of the authors of this study. The interview focused on the utilization of the documented plant species by the respondents. Based on the utilization of plant species by the respondents, we broadly classified the species as provisionary, regulatory, supporting and cultural ecosystem services.
Data analysis
We analyzed the research data using the range, mean, Karl Pearson’s coefficient of correlation and multiple regressions with analysis of variance (Goldstein and Dillon 1985) using IBM SPSS version 2020 using R 3.6.3 (R Core Team 2020) statistical software.
Result and discussion
Floristic composition and diversity
Species richness
Overall, the plant species enlisted for the large cardamom based traditional agroforestry systems in the Darjeeling Himalayas were 130 species represented by 63 families and 107 genera. Altogether, there were 37 tree species represented by 23 families and 30 genera; 25 shrubs species represented by 15 families and 21 genera; 46 herbs species represented by 23 families and 38 genera; 8 ferns species represented by 8 families and eight genera; 11 climbers species represented by 8 families and 9 genera and 3 orchids species represented by 2 families and 3 genera (Supplementary Table 2). The low-altitude class was documented with 76 plant species represented by 63 genera and 43 families. In comparison, the mid-altitude class was represented by 60 species belonging to 57 genera and 40 families and in the high-altitude, class 52 species represented by 45 genera and 35 families were listed. The plant species richness ranges from 8 to 50 species at low altitudes, 8–30 at mid-altitudes and 9–21 at high altitudes (Table 1). The species richness, genus and family of the life forms in the altitudinal classes are given in Table 1. The major plant form found in all the altitude classes was herb, followed by tree, shrub, climber, fern and orchid.
Overall, Asteraceae was the prominent family with 11 species while, the prominent genera were Pilea and Ficus, each with 4 species (Supplementary Table 2). Asteraceae, represented by 8 species was the principal family at low altitudes while, the most prominent genera were Piper, Ficus and Oxalis, each represented by 3 species. In the mid-altitude, Asteraceae, Acanthaceae and Rosaceae, each represented by 4 species, were the prominent families and Ageratum and Rubus, each represented by 2 species, were the major genera. The prominent families recorded in the high-altitude were Asteraceae and Urticaceae, each represented by 6 species and Magnolia with 3 species was the major genus documented in the high-altitude class.
The plant species richness and plant population decreased significantly with increasing altitude. Altitude was found significantly and negatively correlated with both species' richness (r =− 0.648**, Table 2) and plant population (r =− 0.587**, Table 2). However, a significant positive relationship between species richness and plant population was observed (r = 0.890**, Table 2). The traditional large cardamom agroforestry systems are forest-based and very close to natural ecosystems, which are not intervened by human interference except the natural factors, including climate and soil (Sharma et al. 1994, 2007). In such a condition, species richness and population are prominently governed by the temperature and modified by the altitude of the geographical area (Odum 1971). Species richness and plant population of woody species in the large cardamom based traditional agroforestry system thus varied depending on geographical location (topography) and climatic conditions of the region (Kumar and Nair 2004). The inverse relationship of plant species richness with altitude has been reported by other researchers from different studies (Bachman et al. 2004; Trigas et al. 2013; Pandey et al. 2018). Distribution of vegetation exhibit decreasing trend with elevation due to decrease in temperature with the increase in altitude (Wangda and Ohsawa, 2006a, 2006b).
Diversity indices
The diversity indices like Menhinick's species diversity index, the concentration of dominance, the Shannon–Wiener index and evenness index estimated for the Darjeeling Himalayan traditional large cardamom-based agroforestry systems at different altitude classes are given in Table 3. The Menhinick's species diversity index of the studied systems at Darjeeling Himalayas was 1.36, while the index estimated for the different altitude classes was lesser than the overall index, i.e., 1.16, 1.16 and 1.03 for low-, mid-and high-altitude class, respectively (Table 3). The diversity index decreased with increasing altitude class (r =− 0.648**, Table 2) which can be attributed to significant inverse relationship of species richness (r =− 0.648**, Table 2) and plant population (r =− 0.587**, Table 2) with altitude. A higher index indicates that plant species in the low-altitude class were more diverse but less frequently present than those in the higher altitude class. This is because the index is a function of both the total number of species and a total number of individuals of all the species of the sampled sites and lower altitude classes were more species rich than the higher altitude classes due to decreasing temperature with increasing altitude. The diversity index estimated at all the altitude classes was much lesser than the other agroforestry systems of Darjeeling Himalayas, particularly the home gardens (Sarkar 2020), because the species present in the large cardamom based traditional agroforestry systems were more frequent than the species at home gardens of Darjeeling Himalayas.
The overall concentration of dominance was estimated at 0.03, and it increased with the increase in altitude (Table 3). The highest index value at the high-altitude class indicates that the chances of a species encountered during sampling in this altitude class were highest than in the lower altitude classes. However, the chances of species encountered during sampling in the present study area decreased significantly with increasing altitude (r =− 0.580**, Table 2) as both species’ richness and plant population significantly decreased with altitude (r =− 0.648**, − 0.587**, respectively; Table 2). Shannon Wiener's index of the traditional large cardamom-based agroforestry farming of Darjeeling Himalayas was estimated at 4.09 (Table 3), which was less than the traditional home gardens from a similar geographical region (4.75) (Sarkar 2020). The plant diversity decreased with altitude in a progressive manner from 3.79 in low-altitude class to 3.64 in mid-altitude class and 3.24 in high-altitude class, respectively. Altitude inversely influenced diversity, as evidenced by their significant negative relationship (− 0.582**, Table 2).
The evenness index of the traditional large cardamom-based agroforestry systems (0.45) was slightly less than traditional home gardens (0.51) (Sarkar 2020). The diversity indices of trees, shrubs, herbs, climbers, orchids and ferns estimated at different altitude class are given in Table 4. Herbs were more diverse than any other life form and estimated with the highest Shannon–Wiener diversity index compared to other plant forms. The traditional large cardamom agroforestry systems are forest-based with minimal anthropogenic interference, which supports the growth and development of under storey vegetation (Sharma et al. 1994, 2009). Therefore, forest sites with less disturbances have higher herb and under-storey life form diversity (Han et al. 2011). Following herbs, trees were estimated with an overall Shannon–Wiener diversity index of 3.24, and at low-, mid-and high-altitude class, the values were 2.91, 2.69 and 2.35, respectively. The diversity of herbs, trees, climbers and orchids decreased gradually with increasing altitude, but the diversity of shrubs and ferns increased.
Orchids were the least diverse among the plant forms with an overall Shannon–Wiener diversity index of 1.06, and at low- and high-altitude classes, the values were 0.68 and 0.63, respectively. We did not find orchid in the mid-altitude. However, trees were less frequently present with higher estimated Menhinick's species diversity index (overall value of 1.36 and at low-, medium- and high-altitude class with values of 1.10, 1.17 and 1.12, respectively) than the shrubs, climbers, herbs, orchids and ferns. Trees were most evenly distributed with the highest evenness index (with an overall value of 0.49 and at low-, mid-and high-altitude class with values of 0.49, 0.49 and 0.46, respectively) followed by shrubs, herbs, climbers, orchids and ferns. The diversity and dominance of species in the large cardamom based traditional agroforestry systems were lesser than the home gardens at all altitude classes (Sarkar 2020). Similar diversity indices of plant species, particularly the trees, were also reported from large cardamom based traditional agroforestry system of Sikkim Himalayas (Sharma et al. 2008) and other traditional agroforestry systems in India (Salve et al. 2018; Taran and Deb 2019) and elsewhere (Udawatta et al. 2019; Wari et al. 2019).
Sorenson's similarity index suggests a lesser similarity (0.18) for species encountered in large cardamom based traditional agroforestry systems (Fig. 2). In addition, only 17 species were common throughout the altitudinal gradient (Supplementary Fig. 1). Seven species of herbs (Amomum subulatam, Ageratina adenophora, Ageratum conyzoides, Ageratum houstonianum, Brachiaria reptans, Dichroa febrifuga and Drymaria cordata); 4 species of trees (Alnus nepalensis, Cryptomeria japonica, Cupressus cashmeriana and Terminalia myriocarpa), 3 species of shrubs (Boehmeria platyphylla, Girardinia palmata and Phlogacanthus thyrsiformis); two species of ferns (Diplazium esculentum and Selaginella denticulate) and 1 species of climber (Piper boehmeriaefolium) were found in all the three altitudes.

Similarity index of plant species in the large cardamom based traditional agroforestry systems
The similarity of the species decreased gradually with increasing altitude as 36 species were common between low- and mid-altitude class with a similarity index of 0.53, while only 16 species were common between mid- and high-altitude class with a similarity index of 0.29, and 15 species were common between low- and high-altitude class with a similarity index of 0.23 (Fig. 2 and Supplementary Fig. 1). In total, 91 unique or specialist species were listed, of which 39, 21 and 31 species were unique to low, mid and high altitudes (Supplementary Fig. 1). Most of the species documented (70%) in Darjeeling Himalayas' traditional large cardamom-based agroforestry systems were specialists or unique to the altitude class (i.e., low, mid and high). This indicates vast differences or variations among the floristic elements of these traditional systems. This is because topographic factors affect the microclimate and edaphic condition across the altitudinal gradient, thus influencing the vegetation composition (Jha 2001; De 2007).
Similarly, numerous studies also had documented plant species diversity of traditional agroforestry systems across the globe (Nath et al. 2016; Salve et al. 2018; Taran and Deb 2019; Udawatta et al. 2019). The traditional agroforestry system of Darjeeling and Sikkim Himalayas was biodiversity-rich due to the association of shade trees (Sharma et al.1994). Large cardamom based indigenous agroforestry system supports higher tree diversity than other agroforestry systems in the Sikkim Himalayas (Sharma and Sharma 1997; Das et al. 2012). Moreover, traditional agroforestry systems, particularly large cardamom-based agroforestry, are culturally associated with the hill farming community.
Conservation status
The ICUN conservation status of plant species (130 species) from the large cardamom-based agroforestry systems, 89 species were data deficit (68.46%), 38 species were least concerned (29.23%); 2 species (Cryptomeria japonica and Cupressus cashmeriana) were near threatened, and 1 species (Brugmansia suaveolens) was extinct in the wild (Supplementary Table 2, Supplementary Fig. 2). These near threatened and extinct from the wild species are exotics, introduced to India and their IUCN profile indicates their global status, particularly in their native habitat. For instance, Brugmansia suaveolens is extinct from the wild in Brazil (Hay 2014) but occur in Darjeeling and neighbouring foothill regions of West Bengal, India (Mallick 2020).
The highest number of data deficit (47 species) and least concerned (26 species) was recorded from the low-altitude, followed by mid-altitude (41 and 17 species, respectively) and high-altitude (35 and 15 species, respectively; Fig. 3). The two documented near-threatened species were found throughout the altitudinal classes, while Brugmansia suaveolens was found only at the low-altitude class. The inventory of diverse native plant species along with ICUN red-listed ones indicate that these forest-based traditional large cardamom-based agroforestry systems play an important role in harbouring and conserving both native and exotic plant genetic resources (Campos-Salas et al. 2016; Rendón-Sandoval et al. 2020).

ICUN status of the documented plant species at altitude classes
Ecosystem services
The documented plant species were also classified based on their ecosystem services, i.e., provisional, regulatory, supportive, and cultural service (Figs. 4, 5). Overall, in the large cardamom based traditional agroforestry systems 120, 47, 34 and 33 species were classified to provide provisional, cultural, regulatory and supporting ecosystem services, respectively. The agroforestry managers opined that the shade trees ameliorate the microclimate of the area, while agroforestry patches along the mountain slopes prevent soil erosion. The agroforestry managers generally collect dead wood and fallen branches for firewood and non-timber forest products (NTFPs), particularly for food, folk medicine, and other daily provisionary and cultural use. Such agroforestry systems have also been reported to support a variety of regional avifauna and other small wildlife species and ensure green fodder during the lean periods as well (Pandey and Singh, 1984; Sharma and Sharma, 1997). The large cardamom traditional agroforestry systems being very close to natural ecosystems, has the potential to offer a variety of ecosystem services from provisional to cultural services like NTFPs, biodiversity conservation, water regulation and purification, biomass production, carbon sequestration, nutrient cycling and socio-cultural service for the wellbeing of the society (Singh et al. 2018; Vineeta et al., 2021). The present study documented only the preliminary information about the ecosystem services provided by the agroforestry managers and which may differ from farmer to farmer based on their actual utilization, experiences and perceptions. The study tried associating the system at a micro-landscape level with perceived ecosystem services (Kamiyama et al. 2016) rather than its multidimensional roles in enhancing human well-being (Mosina and Maroyi 2016). Inconsistencies lie in perceived information that must be validated comprehensively through interdisciplinary studies at inter- and intra-specific levels within diversified environmental and cultural contexts (Agbogidi and Adolor, 2013). This is because the intangibility of provisioning services may undermine the regulatory services because the perception of provisions (food or medicines) is considered more beneficial than other services (Caballero-Serrano et al., 2016).

Ecosystem services of overall documented plant species

Ecosystem services of the documented plant species at altitude classes. P = Provisional (70, 53 & 48 species respectively at low-, mid- and high altitude class); C = Cultural (22, 20 & 17 species respectively); R = Regulatory (19, 19 & 14 species respectively); S = Supportive (16, 12 & 15, species respectively); P + C = 13, 9 & 5, species respectively; C + R = 2, 3 & 2, species respectively; R + S = 1, 0 & 1 species respectively; S + P = 5, 4 & 6, species respectively; P + R = 6, 7 & 3, species respectively; S + C = 1, 0 & 0, species respectively; P + C + R = 2, 3 & 3, species respectively; C + R + S = 0, 0 & 1, species respectively; R + S + P = 6, 4 & 3, species respectively; S + P + C = 2, 2 & 4, species respectively; P + C + R + S = 1, 2 & 1, species respectively
Conclusions
The present study showed that the altitudinal gradient significantly influenced the species richness, population, and diversity indices. The diversity of herbs, trees, climbers and orchids decreased gradually with increasing altitude, but the diversity of shrubs and ferns increased with increasing altitude. However, trees were less frequently present with higher estimated Menhinick’s species diversity index than the other plant life forms. Trees were most evenly distributed with the highest evenness index, followed by shrubs, herbs, climbers, orchids and ferns. The system's heterogeneity can be attributed to the richness of 130 plant species represented by 63 families and 107 genera. Documentation of ICUN red-listed species indicates that the large cardamom-based agroforestry system plays an essential role in harbouring and conserving regional plant diversity. Additionally, the system was found to offer a variety of ecosystem services.
References
Agbogidi OM, Adolor EB (2013) Homegardens in the maintenance of biological diversity. Appl Sci Rep 1:19–25
Anonymous (2021) Red list of threatened species. Version 2018–2. IUCN. 17 January 2021.
Avasthe RK, Singh KK, Tomar JMS (2011) Large cardamom Amomum subulatum (Roxb.) based agroforestry systems for production, resource conservation and livelihood security in the Sikkim Himalayas. Indian J Soil Conser 39:155–160
Bachman S, Baker WJ, Brummitt N, Dransfield J, Moat J (2004) Elevational gradients, area and tropical island diversity: an example from the palms of New Guinea. Ecography 27:299–310
Caballero-Serrano V, Onaindia M, Alday JG, Caballero D, Carrasco JC, McLaren B (2016) Plant diversity and ecosystem services in Amazonian homegardens of Ecuador. Agr Ecosyst Environ 225:116–125
Cajee L (2018) Physical aspects of the Darjeeling Himalaya: understanding from a geographical perspective. IOSR J Humanities Soc Sci 23:66–79
Campos-Salas N, Casas A, Moreno-Calles AI, Vallejo M (2016) Plant Management in Agroforestry Systems of Rosetophyllous Forests in the Tehuacán Valley, Mexico. Econ Bot 70:254–269
Das A, Ram Krushna GI, Choudhury BU, Munda GC, Patel DP, Ngachan SV, Ghosh PK, Tripathi AK, Das S, Kumar M (2012) Natural resource conservation through indigenous farming systems: wisdom alive in North East India. Indian J Tradit Knowl 11:505–513
De A (2007) Patterns of plant species diversity in the forest corridor of Rajaji-Corbett National Park, Uttaranchal, India. Curr Sci 92:90–93
Goldstein M, Dillon WP (1985) Multivariate analysis: methods and applications. Wiley, New York
Han H, Jang K, Song J, Seol A, Chung W, Chung J (2011) The effect of site factors on herb species diversity in Kwangneung Forest stand. For Sci Technol 7:1–7
Hay A (2014) Brugmansia suaveolens. The IUCN red list of threatened species: 2014: e. T51247699A58913403. https://doi.org/10.2305/IUCN.UK.2014-1.RLTS.T5 1247699A58913403.en
Jha SK (2001) On upper Bias in forest cover data of hilly terrain obtained through satellite imagery with special reference to Mizoram. Indian Forester 127:871–878
Kamiyama C, Hashimoto S, Kohsaka R, Saito O (2016) Noo-market food provisioning service via homegarden and communal sharing in satoyama socio-ecological production landscapes on Japan’s Noto peninsula. Ecosyst Serv 17:185–196
Kumar BM, Nair PKR (2004) The enigma of tropical homegardens. Agrofor Syst 61:135–152
Lepcha LD, Shukla G, Pala NA, Vineeta PPK, Chakravarty S (2018) Contribution of NTFPs on livelihood of forest-fringe communities in Jaldapara National Park, India. J Sustain for. https://doi.org/10.1080/10549811.2018.1528158
Lepcha LD, Shukla G, Vineeta SBC, Tamang M, Chettri R, Manohar AK, Pala NA, Chakravarty S (2019) Ethno-Botanical plant diversity in homegardens based agroforestry systems in Khangchendzonga Biosphere Reserve, Sikkim, India. J Tree Sci 38:59–69
Mallick JK (2020) An annotated checklist of dicotyledonous angiosperms in Darjeeling Himalayas and foothills, West Bengal India. J New Biol Rep 9:94–208
MEA (2005) Ecosystems and human well-being: Synthesis. Millennium Ecosystem Assessment. Island Press, Washington DC
Mehta MP, Rabgyal J, Acharya S (2015) Commodity chain analysis of large cardamom in Bhutan. FAO, Rome
Menhinick EF (1964) A comparison of some species diversity indices applied to samples of field insects. Ecology 45:858–861
Moktan S, Das AP (2013) Diversity and distribution of invasive alien plants along the altitudinal gradient in Darjeeling Himalaya, India. Pleione 7:305–313
Molden D, Verma R, Sharma E (2014) Gender equality as a key strategy for achieving equitable and sustainable development in mountains. Mt Res Dev 34:297–300
Mosina GKE, Moroyi A (2016) Edible plants of urban domestic garden in the Capricorn district Limpopo province, South Africa. Trop Ecol 57:181–191
Nath AJ, Reang D, Das AK, Brahma B, Das M (2016) Traditional practice Paan Jhum cultivation among Khasia community in Barak Valley, Assam. J Tradit Folk Pract 4:96–99
Negi BK, Joshi RK, Pandey A (2018) Status of large cardamom (Amomum subulatum Roxb.) farming systems in the changing scenario of modern economics of Sikkim, Himalaya. Glob J Bio-Sci Biotechnol 7:189–199
Odum EP (1971) Fundamentals of Ecology, 3rd edn. W. B. Saunders Co., Philadelphia
Pandey A, Rai S, Kumar D (2018) Changes in vegetation attributes along an elevation gradient towards timberline in Khangchendzonga National Park, Sikkim. Trop Ecol 59:259–271
Pandey U, Singh JS (1984) Energy-flow relationship between aground forest ecosystem in Central Himalaya. Environ Conserv 11:45–53
Pielou EC (1975) Ecological diversity. Wiley, New York
R Core Team (2020) R: A language and environment for statistical computing. A Foundation for statistical computing. Vienna, Austria. http:/www.R-project.org/
Reang D, Hazarika A, Sileshi GW, Pandey R, Das AK, Nath AJ (2021) Assessing tree diversity and carbon storage during land use transitioning from shifting cultivation to indigenous agroforestry systems: Implications for REDD+ initiatives. J Environ Manage 298:113470
Reang D, Sahoo UK, Giri K, Hazarika A, Nath AJ (2022) Ethnic homestead forests of North East India revealed as diverse land-use systems. Agrofor Syst. https://doi.org/10.1007/s10457-021-00700-z
Rendón-Sandoval FJ, Casas A, Moreno-Calles AI, Torres-García I, García-Frapolli E (2020) Traditional agroforestry systems and conservation of native plant diversity of seasonally dry tropical forests. Sustainability 12:3–27
Roy M, Sarkar BC, Shukla G, Vineeta, Debnath MK, Nath AJ, Bhat JA, Chakravarty S (2022a) Traditional homegardens and ethnomedicinal plants: insights from the Indian Sub-Himalayan region. Trees. Forest People 8:100236. https://doi.org/10.1016/j.tfp.2022.100236
Roy M, Sarkar BC, Abha MK, Shukla G, Vineeta, Nath AJ, Bhat JA, Chakravarty S (2022b) Fuelwood species diversity and consumption pattern in the homegardens from foothills of Indian Eastern Himalayas. Agrofor Syst. https://doi.org/10.1007/s10457-021-00728-1
Salve A, Bhardwaj DR, Tahkur CL (2018) Floristic composition and distribution pattern of plant communities under different agroforestry systems in Kinnaur, North-Western Himalayas. Res J Chem Environ Sci 6:17–34
Sarkar BC (2020) Ecosystem services and floristic diversity of homegardens along altitudinal gradient of Darjeeling Himalayas. Ph. D. Dissertation. Uttar Banga Krishi Viswavidyalaya, Pundibari. Unpubl
Shannon CE, Weaver (1949) The mathematical theory of communication. University of Illinois Press, Urbana, p 117p
Shannon CE, Weiner W (1963) The mathematical theory of communication. University of Illinois Press, Urbana
Sharma E, Chettri N (2021) Biodiversity conservation and ecosystem services in the Hindu Kush Himalaya. J Graph Era Univ. https://doi.org/10.13052/jgeu0975-1416.911
Sharma E, Sharma R, Sharma G, Rai SC, Sharma P, Chettri N (2008) Values and services of nitrogen-fixing alder-based cardamom agroforestry systems in the Eastern Himalayas. In: Snelder DJ, Lasco RD (eds) Smallholder tree growing for rural development and environmental services: lessons from Asia. Springer, Berlin, pp 393–408
Sharma E, Sharma R, Singh KK, Sharma G (2000) A boon to mountain populations, large cardamom farming in the Sikkim Himalaya. Mt Res Dev 20:108–111
Sharma G, Sharma E (1997) Mountain agricultural transformation process and sustainability in the Sikkim Himalayas, India. Mountain Farming Systems: ICIMOD, Kathmandu, Nepal
Sharma G, Partap E Sharma GR, Awasthi RK (2016) Agrobiodiversity in the Sikkim Himalaya: Socio-cultural Significance, Status, Practices, and Challenges. ICIMOD Working Paper 2016/2. ICIMOD, Kathmandu
Sharma G, Sharma P, Sharma E (2009) Traditional knowledge systems in large cardamom farming: Biophysical and management of diversity in Indian mountainous regions. Indian J Tradit Knowl 8:17–22
Sharma R, Jianzhu XU, Sharma G (2007) Traditional agroforestry in the Eastern Himalayas region; land management system supporting ecosystem. Trop Ecol 48:1–12
Sharma R, Sharma E, Purohit AN (1994) Dry matter production and nutrient cycling in agroforestry systems of cardamom grown under the Alnus and natural forest. Agrofor Syst 27:293–306
Sharma S, Singh B, Sikka R (2015) Changes in some physicochemical characteristics of soils under poplar-based agroforestry. Agric Res J 52:19–22
Shrestha J, Prasai HK, Timsina KP, Shrestha KP, Pokhrel D, Poudel K, Yadav M (2018) Large cardamom in Nepal: production practice and economics, processing and marketing. Nepal Agriculture Research Council, National Commercial Agriculture Program, Pakhribas, Dhankuta, Nepal
Simpson EM (1949) Measurement of diversity. Nature 163:688
Singh M, Das SK, Avasthe RK (2018) Effect of multipurpose trees on production of large cardamom and soil fertility in agroforestry systems in Sikkim, Himalaya. Indian J Agrofor 20:25–29
Sorenson T (1948) A method of establishing groups of equal amplitude on similarity of species content. Biologiske Skrifter k Danske Videnskbernes Selskab 5:1–34
Tarafder HK, Roy K, Rai B (2018) Large cardamom production scenario in mountainous region of West Bengal: Constraints, issues and strategies. Asian J Soil Sci 13:153–156
Taran M, Deb S (2019) Plant diversity and ecosystem services of wetland-based agroforestry system in Tripura, Northeast India. Indian J Agrofor 21:13–17
Trigas P, Panitsa M, Tsiftsis S (2013) Elevational gradient of vascular plant species richness and endemism in Crete- the effect of Post-isolation Mountain uplift on a continental island system. PLoS ONE 8:59425
Udawatta RP, Rankoth ML, Jose S (2019) Agroforestry and biodiversity. Sustainability 11:1–22
Vineeta TB, Siril S, Singh M, Das S, Shukla G, Chakravarty S (2021) Ecosystem services of traditional large cardamom-based agroforestry systems of Darjeeling and Sikkim Himalayas. J Tree Sci 40:78–91. https://doi.org/10.5958/2455-7129.2021.00006.6
Wangda P, Ohsawa M (2006a) Structure and regeneration dynamics of dominant tree species along altitudinal gradient in dry valley slopes of the Bhutan Himalaya. For Ecol Manage 230:136–150
Wangda P, Ohsawa M (2006b) Forest pattern analysis along the topographical and climatic gradient of the dry west and humid east slopes of Dochule, western Bhutan. Bhutan J Renew Nat Resour 2:1–17
Wari BN, Feyssa DH, Kebebew Z (2019) Assessment of woody species in agroforestry systems around Jimma Town, Southwestern Ethiopia. Int J Biodivers Conser 11:18–30
Acknowledgements
Authors are sincerely acknowledging help and support received from the large cardamom growers for completing this research work.
Funding
No.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Vineeta, Sarkar, B.C., Tamang, M. et al. Floristic diversity, and conservation status of large cardamom based traditional agroforestry system along an altitudinal gradient in the Darjeeling Himalaya, India. Agroforest Syst 96, 1199–1210 (2022). https://doi.org/10.1007/s10457-022-00778-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-022-00778-z