
Abstract
Forests in the highlands of Ethiopia have declined considerably, and the supply of forest-based ecosystem services is eroding. Managing agricultural landscapes as well as enhancing plant biodiversity in agro-ecosystems is and will continue to be one possible strategy to preserve biodiversity, ensure an ecosystem service supply and sustain agricultural productivity. This study investigated the current status and prospects of plant diversity and its determinants in an agricultural landscape dominated by smallholder farmers in Southern Ethiopia. Specifically, the study investigated effects of land use, altitudinal gradient, wealth status and household attributes on plant diversity in agricultural landscapes. A complete count, Y-frame transect sampling method and household interviews were used for the study involving 39 households and 115 sample plots. A total of 166 plant species belonging to 134 genera and 56 families were recorded in all land use types. Of these, 101 were woody plant species (51 trees and 50 shrubs), while 65 were cultivated herbs and grasses. Land use, altitude and household wealth status significantly influenced tree and shrub species richness. Among land uses, home gardens hosted the highest number of tree and shrub species. Upper altitudes and rich households also had the highest tree and shrub species richness compared to others. Plant diversity indices (Shannon, Simpson and Margalef) were affected by altitude, wealth status and land use types. Household location, wealth status, the household attributes of landholding and family size had strong and positive influences on the tree species diversity and woody stem density of households, while educational background and off-farm income were negatively related with household-owned tree stem density. Species preference ranking, seedling demand and importance value index computations indicated the dominance of exotic tree species, which may suggest their economic importance over indigenous tree species. In the long term, this might lead to dominance of exotic tree species in the landscapes, which could cause a potential future threat to the remnant indigenous plant diversity that is currently finding refuge in the agricultural landscapes.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Background
Deforestation and the consequent losses of biodiversity are the top global environmental problems today. Agro-ecosystem-based biodiversity conservation has received considerable attention as a supplement to the conventional conservation methods following the continued contraction of natural ecosystems (Boffa et al. 2008). This approach can be considered an integrated strategy aimed at ensuring the supply of multiple ecosystem goods and services including biodiversity conservation (MEA 2005; Ranganathan et al. 2008; Negash et al. 2012; Negash 2013; Tadesse et al. 2014a, b). The conservation of plant biodiversity in the agricultural landscapes dominated by smallholder farmers is receiving growing attention, popularity and priority in Africa (Garrity and Verchot 2008; CRGE 2010; Lemenih and Kassa 2014).
Ethiopia is one of the globally important centers of plant diversity, endemism and plant domestication owing to its diverse climates, altitudes, topography, soil conditions, natural vegetation and cultures (Edwards and Kelbessa 1999; Teketay 2001; Hurni 2007). However, the diverse natural vegetation and biodiversity have been and still are under intense pressure, particularly in the highlands of Ethiopia, as a result of the long history of human settlement and cultivation, persistent deforestation, over-grazing and growing agricultural intensification (Tadesse 2001; Lemenih and Teketay 2006; Kindu et al. 2013, 2015; Telila et al. 2015). Forest resources in Ethiopia have been declining at an alarming rate of 140,000 ha per annum (FAO 2010; Teketay et al. 2010), and the biodiversity and other forest-based ecosystem services are apparently eroding (Tadesse et al. 2014a; Kindu et al. 2015, 2016). Agricultural intensification through the adoption of improved seeds, inorganic fertilizers and pesticides is also exacerbating the environmental degradation, which in turn leads to a decline in agricultural productivity and agro-biodiversity (Hadgu et al. 2009). Landscape and agricultural production transformations caused by the rapid expansion of exotic tree species and economically important cash or food crops are threatening the traditional farming systems, which are more biodiversity friendly, such as the parkland and multistrata agroforestry systems common in Southern Ethiopia (Achalu 2004; Abebe 2005; Dessie 2007; Negash et al. 2012).
Given the continued high rural population growth in the country, at the rate of 2.6%, the pressure on natural ecosystems and the associated biodiversity resources will continue unabated (Abebe 2005; Negash et al. 2012; Kindu et al. 2013, 2015). Strengthening the management of biodiversity in agricultural landscapes is crucial to complement other biodiversity conservation initiatives through national parks, wildlife sanctuaries, priority forest areas or protected forests, area enclosures and gene banks in the country (Zomer et al. 2009; Boffa et al. 2005; Negash et al. 2012; Tadesse et al. 2014a). Agro-bio-diverse ecosystems have contributed to biodiversity conservation via (1) provision of supplementary habitats/refuges for native flora and fauna; (2) minimizing the pressure on natural forests and their biodiversity through provision of alternative sources; (3) connecting wild populations of fragmented forests with those of continuous forests through landscape connectivity; (4) conserving the gene pools of native woody species such as wild coffee by providing habitats; (5) enhancing the gene flow of native flora by providing habitats for pollinators and seed-dispersing animals (Boffa et al. 2005; Abebe 2005; Senbeta and Denich 2006; Negash et al. 2012; Tadesse et al. 2014a).
However, the potential of agro-ecosystems in biodiversity conservation varies greatly depending on the land management, land use, and environmental and socioeconomic conditions of an area (Abebe 2005; Tolera et al. 2008; Abebe et al. 2009; Duguma and Hager 2010; Negash et al. 2012; Tadesse et al. 2014a). For instance, home garden and agricultural-forest mosaic landscapes in Southern and Southwest Ethiopia host a remarkably higher number of woody species than the cereal-based agricultural landscape common to Central and Northern Ethiopia (Tolera et al. 2008; Duguma and Hager 2010; Tefera et al. 2014). The altitude, soil conditions, slope and aspects, and household attributes also affect the role of the agricultural landscape in biodiversity conservation (Nair 1993; Abebe 2005; Mengistu 2008; Hadgu 2008; Negash et al. 2012; Kebede et al. 2013; Yirdaw et al. 2015). For instance, several studies reported the positive relationship between household wealth status on wood species diversity and abundance (e.g., Jarvis et al. 2000; Kindt et al. 2004; Asfaw and Ågren 2007). However, in some cases such positive relationships were not observed (e.g., Boffa et al. 2008; Duguma and Hager 2010). The variation in plant species composition and diversity among different locations and land uses indicated that more studies are needed across diverse ecological and socioeconomic settings to comprehensively understand the role of agro-ecosystems in plant diversity preservation and its determinants. This study investigated the role of smallholder agricultural landscapes for plant diversity conservation in part of Ethiopia’s Central Rift Valley. The specific objectives were to: (1) assess plant species composition and diversity; (2) identify determinant factors of the presence of plant diversity and woody stems; (3) assess future prospects of woody species among smallholder farmers; (4) understand the importance of managing plant biodiversity, i.e., local uses.
Materials and methods
Description of the study area
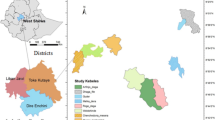
This study was conducted in the Meskan district, which is located between 38°15′0.7″–38°33′50.9″E and 8°1′58.8″–8°16′29.6″N in Central Ethiopia (Fig. 1). Butajira is the district town and is located about 130 km south of Addis Ababa. The study area is characterized by diverse topographic features consisting of plains (55%), sloping land (35%) and mountains (10%) (WARD 2013).

Map of the study area and location of the studied households
The major land uses in the district include: cultivated land (23,234 ha), natural vegetation (5425 ha), plantation forests (4668 ha), grazing land (3346 ha) and settlements and roads (11,703 ha). The characteristic agricultural practice in the study area is a subsistence mixed crop-livestock farming system. The most commonly grown crops are maize, teff, wheat, enset, pulse and sorghum (Table 1). The area is characterized by bimodal rainfall. It receives rain from March to September with major rains usually taking place in July and August (Fig. 2). The average mean annual rainfall is 1167 mm, and mean monthly maximum and minimum temperatures are 27.3 and 10 °C, respectively (Fig. 2).

Climatic condition of the study area (Source: NMAE 2015)
The district's altitude ranges from 1700 to 3500 m above sea level (asl). The dominant soil types of the district include eutric cambisols, chromic luvisols, pellic vertisols, chromic vertisols, eutric fluvisols and leptosols (FAO 1994). A few scattered tree species of highland types (e.g., Acacia abyssinica, Cordia africana, Juniperus procera, Croton macrostachyus) are observed in the upper and mid-altitude areas. In the lower altitude areas, tree species of woodland types such as Acacia seyal and Faidherbia albida are scattered in the landscapes (Table 1). Economically important exotic tree species such as Eucalyptus camaldulensis are also grown and managed widely.
Methods
Household selection
Three rural kebelesFootnote 1 were selected along an altitudinal gradient. These were Goyban from a high-altitude area (2200–2400 m asl), Mekicho from a mid-altitude area (2000–2200 m asl) and Ile from a low-altitude area (1800–2000 m asl) (Table 1). Three geo-referenced points were purposively selected from each kebele. From each geo-referenced point, a Y-framed transect line was established (Henry et al. 2009). A list of households residing in the selected kebeles was obtained from the kebele administration office and divided into three wealth classes (poor, medium and rich) with the help of the local elders and development agents. Local criteria such as land holding size, housing status and livestock holdings were used for the classification of the households (Table 2). Then, sample households from the three wealth classes were selected along the transect lines at approximately 0, 300, 600 and 900 m distances from the original point. An equal number of households from each wealth class was selected. A total of 39 (i.e., 13 poor, 13 medium and 13 rich) households were selected. Farms of the selected households (i.e., land managed by a household) were categorized into different land use types such as home garden, farmland, grazing land and woodlot. Home garden describes land use in and around a homestead, often demarcated by a live fence, commonly used for growing enset, vegetables, cereals, trees and shrubs. Farmland is a plot far from home dominantly used for growing annual crops. Grazing land is land allocated for private grazing purposes, and woodlots are land allocated for tree planting.
Data collection
The selected households were visited to assess the spatial variability of plant biodiversity from November 2013 to October 2014. As farmers own two or more pieces of plots, data were collected from all management units by each household, and the data were averaged for analysis. In each plot, parameters such as species richness, abundance (individuals) and tree diameter at breast height (cm) were collected. For tree and shrub species, individuals with diameter at breast height (DBH) ≥3 cm were counted and their respective DBH measured. However, for those tree and shrub individuals with DBH ≤3 cm and height greater than 20 cm, only the number of individuals were counted. All trees and shrubs with a height above 20 cm were considered for biodiversity analysis. In addition, the area coverage of major herbaceous crops was measured using a meter tape. The collected species were identified in the field based on vernacular names using a supplementary field guide (Bekele 2007). For species difficult to identify in the field, specimens were collected and taken to the National Herbarium at Addis Ababa University for identification. The nomenclature follows the Flora of Ethiopia and Eritrea, volume 1–7. Local uses were recorded using key informants.
To evaluate the effect of socioeconomic variables on having woody plants, household attributes such as wealth status, household location (altitude), family size, farm size (ha), livestock holding (TLU), age of the household head, housing status, educational background and off-farm income were recorded using semi-structured questionnaires.
Additionally, households were asked to list the woody species they would like to plant in the future and the number of seedlings they would potentially be planting. This was used as an indicator to predict whether there will be changes in the trend of woody species composition in the landscape in the future. Specially, eight of the woody plants with the highest frequency in the area were evaluated concerning whether they were included in the farmers' responses. In addition, 21 key informants (7 from each location) were sampled among the 39 sample households based on their knowledge and enthusiasm regarding woody plants. A preference-ranking exercise was conducted with each key informant separately. The six most frequently mentioned local uses and two most commonly cited criteria for cultivation were included as attributes for preference ranking. The local names of the species and attributes were printed along columns and rows, respectively, on separate sheets of paper, and then each key informant was asked to vote based on his/her preference. The informants assigned points on a scale from 1 to 8 (the highest ranked species received a score of 8 and the lowest a score of 1). The response was computed by summing the points given by 21 key informants for each species, and then an overall rank was assigned to the respective species.
Data analysis
The ecological importance of each species in a given land use and landscape was demonstrated using parameters such as relative frequency, relative abundance (%) and relative dominance (%) as well as the importance value index. Relative dominance is the percentage share of the basal area of a given species out of the total measured stem basal areas for all species. Relative abundance is the percentage of the abundance of each species out of the total stem numbers for all species. Relative frequency is the percentage of the frequency (the percentage of the total number of plots containing the species compared to all plots) of a species compared to the total frequencies of all the species added together. The importance value index of each species was computed by summing the relative frequency, relative abundance and relative dominance. For multiple stem plants such as coffee, dominance was calculated by measuring the root collar diameter of all stems and by summing the square root of the cross-sectional areas of all stems at that height (Snowdon et al. 2002). The same approach was applied for other species as well. Basal area (m2) is the sum of the basal areas of all stems in the assessed area of land calculated as:
where DBHi is the diameter at breast height of the ith stem.
Species richness and diversity for each land use, altitude and wealth class were calculated using a popular index of alpha diversity (Magurran 1988): Shannon’s index (H’), Simpson’s evenness index (E1/D) and Margalef’s index (Dmg) were also calculated (Shannon 1949; Magurran 1988). The former index was selected because of its sensitivity to sample size since it gives more weight to rare species, whereas the latter two indices were selected because they had less sensitivity to plot size.
Three-way analysis was conducted using the generalized linear model (GLM) to investigate the effect of land use, altitudinal gradient and wealth status on plant diversity and woody stem possession among households. Altitude was considered the main factor, wealth a sub-factor and land use a sub-sub-factor. Altitude comprised three levels, i.e., upper, middle and lower, and wealth comprised three levels, i.e., poor, medium and rich, while land use comprised four levels, i.e., home garden, farmland, grazing land and woodlot. Separate analyses were performed for each plant attribute, namely species richness, the Shannon index, Margalef index and Simpson evenness index, and the density of tree and shrub species, using SPSS software. When the analysis of variance (ANOVA) showed a significant difference (P < 0.01 or P < 0.05), mean separation was made using Tukey’s pairwise comparisons (SPSS Inc 2006).
The potential influence of explanatory variables such as farm size, wealth status, livestock holding, family size, off-farm income, educational background, housing status and age of the household on having woody plants was assessed by applying regression models. However, as wealth status, livestock and landholding were strongly correlated with each other, the former two variables were not used in the analysis. A separate analysis was performed for households' tree and shrub possession using the GLM of the quasi-Poisson family with the log link function in biodiversity. R software developed by the R Development Core Team (2010, 2012) was used for the analysis. In spite of their ordinary nature, wealth status and household locations were treated as categorical variables during the GLM procedure. The GLM was chosen for the analysis because of its ability to accommodate different types of predictor variables within the same function. This helped us to include both the continuous and categorical predictor variables mentioned earlier into the same model. Quasi-Poisson distribution was chosen to take into account the over-dispersion of the data, and this helped to estimate the dispersion parameter from the data rather than forcing it to be unity as in normal Poisson distribution.
Results and discussion
Species composition and its importance
A total of 166 plant species belonging to 134 genera and 56 families were identified. Of these, 101 (60.4%) were woody plants and 65 (39.5%) were cultivated crops (Online Appendix). In terms of life form, 51 were trees, 50 were shrubs, 52 were herbs, and 13 were grass species. The family Fabaceae was the most dominant, represented by 26 (46.4%) species, followed by Poaceae with 10 (19.6%) and Solanaceae with 9 (16%).
Home gardens hosted the largest woody species composition (96 species), followed by grazing land (30), eucalyptus woodlots (29) and farmland (25) (Online Appendix). Overall, this study identified a greater number of woody species than previous similar studies in Ethiopia. For instance, Tolera et al. (2008) recorded 70 woody species; Duguma and Hager (2010) recorded 54 species, while Tefera et al. (2014) recorded only 48 species from agricultural landscapes in Central and Northern Ethiopia, respectively. The number of woody species identified in this study was also higher than that reported from multistrata agroforest systems in Southern Ethiopia (Negash et al. 2012). However, Abebe (2005) and Tadesse et al. (2014a) found more woody species, 120 and 155 species, in enset-based home garden and agroforestry-forest mosaic landscapes in Southern and Southwestern Ethiopia, respectively. Our results concerning the proportion of native tree species (68%) hosted in agricultural landscapes was comparatively lower than those recorded in the typical multistrata agro-forestry system of the Sidama and Gedeo Zone of Southern Ethiopia, respectively (Abebe 2005; Negash et al. 2012).
Home gardens hosted the largest number of both domesticated and non-domesticated plant species, 82 and 74, followed by farmlands (30, 22), while grazing land and woodlots supported the fewest domesticated plants (Table 3).
The variation in woody species richness among different locations may be the result of socio-cultural differences, land use and management intensities, farmers' management strategies, environmental factors and the presence of adjacent fragmented forests (Abebe 2005; Tolera et al. 2008; Duguma and Hager 2010; Negash et al. 2012; Tadesse et al. 2014a, b).
Exotic tree species ranked high in terms of frequency, abundance and dominance and had an importance value index in the home gardens and woodlots, whereas native trees were dominant in farm and grazing lands (Table 4). The three commonly planted exotic tree species, namely E. camaldulensis, C. lusitanica and G. robusta, account for about 76.8% of the relative abundance of all recorded tree species across the land use systems investigated. This trend signals a tendency toward favoring exotic plant species over indigenous ones; hence, this calls for precautionary measures to manage and conserve native tree species. Variation in species, an importance value index, was observed among the investigated land uses, altitudes and wealth classes. E. camaldulensis contributes the most to the importance value index in home gardens (79.33%) and woodlots (218.08%) (Table 4). The importance value index of E. camaldulensis showed a declining trend with increased altitudinal gradient, i.e., 96.21% in upper, 129.54% in lower and 139.96% in middle altitudes.
On the other hand, its contribution increased with decreasing household wealth status, i.e., 84.08% in rich, 104.02% in medium and 106.22% in poor households (Table 4). The dominance of exotic species may be due to their economic and domestic importance for use as construction wood, firewood and income generation (Online Appendix). Moreover, their fast-growing nature and adaptation to marginal environments (degraded, i.e., poor soil nutrients, water logging and free grazing situations) are also contributing factors (Jagger and Pender 2003; Achalu 2004; Lemenih 2010). Several other studies (e.g., Abebe 2005; Dessie 2007; Tolera et al. 2008; Duguma and Hager 2009, 2010; Tefera et al. 2014) showed the dominance of exotic tree species in the agricultural landscape in the highlands of Ethiopia. Farmers in the study area also retain some native trees species, mainly as parkland, agroforestry on farmlands and grazing lands (Table 4). C. africana (97.37%), C. macrostachyus (73.68%), A. abyssinica (65.79%) and A. albida (44.74%) were the most frequently occurring native tree species among others (Online Appendix). The highest importance value index of A. abyssinica (77.19%) and A. albida (91.23%) was in farmland. A. abyssinica also had the highest importance value index of 151.67% in grazing land (Table 4).
The importance value index of A. abyssinica was highest (52.55%) in the upper altitude and declined with decreasing altitude, while that of A. albida was highest in lower altitude (39.69) and declined with increasing altitudinal gradient (Table 4). This may indicate the environmentally related distributional pattern of the two species.
With respect to shrub species diversity, indigenous shrub species dominate over exotics in frequency, abundance, dominance and importance value index in home gardens (Online Appendix). For instance, five of the indigenous shrub species, Catha edulis, Rhamnus prinoides, Dovyalis abyssinica, Justicia schimperiana and Vernonia amygdalina, together account for about 79% of the relative abundance of all shrub species in home gardens (Table 5). This result fits well with the findings of Duguma and Hager (2010) who found a higher abundance of the former two economically important shrub species. According to the same authors, the two shrub species account for about 87% of the abundance of all shrub species in the home gardens of Central Ethiopia. C. edulis makes the highest importance value index contribution of 63.28% of all shrub species in home gardens, 128.01% in middle altitude and 122.29% in medium households (Table 5). In addition to these shrub species, Coffee arabica was also the most frequently encountered economically important indigenous shrub species with a relative frequency of 97.37% in agricultural landscapes (Online Appendix).
Farmers also dominantly grow annual food crops such as maize, teff and wheat and perennial food crops such as enset (Ensete ventricosum) (Online Appendix). Of the total cropped land of the studied households, maize had the highest area coverage of 25.19%, followed by teff (22.54%), wheat (15.35%) and enset (14.17%) (source: own field survey, 2014). Maize had the highest relative frequency of 81.58%, followed by enset (78.95%), teff (76.32%) and wheat (63.16) (Online Appendix ). We recorded about 22 landrace varieties of enset ranging from 3 to 13 per farm. This demonstrates the presence of higher numbers of landrace varieties of enset crops per farm.
Preserving diverse trees, shrubs and crops serves several functions including wood for different purposes (e.g., firewood, timber, local construction, farm implements, household utensils), fodder, food and medicine. They also play beneficial ecological roles such as erosion control and soil fertility improvement. For instance, 46 (27.8%) plants are used for medicine (both human and veterinary medicines) and 40 (25%) for food, of which 23 (13.9%) are herbaceous and grasses and 17 (10.2%) are fruit trees and shrubs; 32 (19.28%) are used for fuel wood, 29 (17.47%) for soil fertility maintenance, 17 (10.24%) for live fencing, 13 (7.8%) for fodder, 14 (8.4%) for shade and 12 (7.2%) for construction; 7 (4.2%) woody plants are grown intentionally for income generation and the rest for various local uses (Online Appendix). However, according to local informants, lack of seedlings, land shortage, wild animal pests and diseases, such as coffee berry disease and enset bacterial wilt, lack of water availability, lack of tree crop compatibility and labor shortages were the most frequently cited constraints in decreasing order of frequency to promote agro-plant diversity. Several authors (e.g., Abebe 2005; Duguma and Hager 2010; Hailu and Asfaw 2011; Abebe 2013; Lemmesa et al. 2013; Ango et al. 2014) have noted similar constraints that influence biodiversity conservation in Central and Southern Ethiopia.
Influence of altitude, wealth status and land use on plant diversity
The existence of statistical differences along altitudinal levels, wealth classes and land use types for plant species richness, diversity indices (Shannon, Margalef and Simpson evenness indices), density of tree and shrub stems per ha and land use was tested using three-way (ANOVA) and Tukey’s pairwise comparisons (Online Appendix and Table 6). The results show that the number of tree and shrub species varied significantly among land use types, wealth and altitudinal classes (Online Appendix). All three first-order interactions, altitude × wealth, altitude × land use and wealth × land use, were significant for tree species richness (P < 0.05), while shrub species richness was significant for first-order interactions of altitude × land use and wealth × land use ((P < 0.05) (Online Appendix).
Home gardens had the highest number of 11.57 ± 0.72 tree and 11.33 ± 0.86 shrub species richness followed by grazing land, farmland and woodlots (Table 6). This is consistent with the findings of several other similar studies (e.g., Abebe 2005; Tolera et al. 2008; Henry et al. 2009; Duguma and Hager 2010; Baral et al. 2013) that found significantly higher woody species richness and diversity indices in home gardens than in other land use types. Rich households had the highest tree and shrub species richness of 5.78 ± 0.87 and 4.5 ± 0.99 per field, respectively, compared to other wealth categories (Table 6).
Plant diversity indices were also affected by altitude, wealth and land use types and by first-order interaction (Online Appendix). The Shannon index of tree species was significantly affected by altitude, wealth and land use (P < 0.05) and by all the first-order interactions of altitude × wealth, altitude × wealth and altitude × land use (Online Appendix), while the Shannon index of shrub species was only significantly affected (P < 0.05) by wealth and land use types (Online Appendix). Home gardens had the highest Shannon index value of 1.43 ± 0.06 and 1.42 ± 0.07 for tree and shrub species, respectively (Table 6). This is comparable to the Shannon index value of tree species recorded in tree-crop-based and enset-coffee-based home gardens of Western Kenya and Southern Ethiopia, respectively (Henry et al. 2009; Abebe 2005). However, the recorded Shannon index value was higher than that recorded by Negash et al. (2012) in the multistrata agro-forestry system of the Southeastern Rift Valley escarpment of Ethiopia.
The Margalef index values of tree and shrub species were significantly affected by altitude, land use and wealth (P < 0.05) and by the first-order interaction of altitude × wealth and altitude × land use (P < 0.05) (Online Appendix). This is in concordance with the results of Negash et al. (2012), who found a significant influence of land use on the Margalef index of woody species in the Southeastern Rift Valley escarpment of Ethiopia.
Home gardens had the highest Margalef index of 2.39 ± 0.22 and 1.88 ± 0.11 tree and shrub species, respectively (Table 6). Likewise, rich households had the highest Margalef index of 1.1 ± 0.16 and 1.07 ± 0.15 for tree and shrub species, respectively, compared to other wealth categories (Table 6). However, the Margalef index value recorded in this case was lower than that recorded by Negash et al. (2012) in multistrata agroforestry in the Southeastern Rift Valley of Ethiopia. Similarly, Simpson’s evenness index of tree species was significantly (P < 0.05) affected by altitude, wealth and land use and by the first-order interaction of altitude × wealth, while Simpson’s evenness index of shrub species was significantly affected (P < 0.05) by wealth and the first-order interaction of altitude × wealth (Table 6, Online Appendix). Farmland had a significantly higher Simpson’s evenness index value of 0.61 ± 0.06, followed by home gardens, grazing land and woodlots (Table 6). The Margalef index increased with increased woody species richness, while Simpson’s evenness index increased with decreased species richness (Table 6). In contrast, Negash et al. (2012) showed an increasing trend of both the Simpson’s evenness and Margalef indices with decreasing species richness in multistrata agroforestry in the Southeastern Rift Valley of Ethiopia.
The observed higher plant species composition and diversity indices in home gardens could be explained by (1) the presence of diverse plants with different uses, i.e., farmers' management strategies, (2) its closeness to home, which might favor easy management, e.g., fencing, watering, weeding/hoeing and soil fertility maintenance, and (3) the presence of better security in home gardens in terms of land tenure, theft or illegal cutting, and fewer free grazing problems.
The total plant species were significantly affected by altitude, land use and wealth (P < 0.05) and by the first-order interaction of altitude × land use and wealth × land use (Online Appendix, Table 6). Home gardens hosted a significantly higher number of plant species, 32.82 ± 2.18, than other land uses (Table 6). Upper altitudes and rich households had significantly higher total plant species of 14.1 ± 3.28 and 15.61 ± 2.64 compared to middle and lower altitudes and medium and poor households, respectively (Table 6).
Furthermore, the results from this study show that the number of tree and shrub species/household were significantly affected by wealth (P < 0.001) (Fig. 3a, b). Rich households had the highest number of tree species of 15.25 ± 0.89 followed by medium (13.72 ± 1.12) and poor (9.73 ± 0.97) per household (Fig. 3a). Likewise, rich households had the highest number of shrub species of 13.29 ± 1.36 followed by 11.5 ± 0.8 in medium and 9.08 ± 1.35 in poor households. It is logical that rich households having large farms and capital allocate more resources (e.g., space, labor and capital) for maintaining a large number of trees and shrub stems. Hence, the more land a household owns, the higher the chance is to grow more trees and shrubs. The observed significant effect of wealth on agro-plant diversity also fits well with the results of several other authors (Jarvis et al. 2000; Kindt et al. 2004) who acknowledged the influence of wealth status on farm tree diversity and farmers' tree management strategies. However, such influences were not observed in some cases (e.g., Boffa et al. 2008; Duguma and Hager 2010).

Number of tree species/hh (a) and shrub species/hh (b) across wealth class
Upper altitude had the highest number of 5.36 ± 0.34 and 4.58 ± 0.37 tree and shrub species per field followed by middle and lower altitudinal classes (Table 6). The number of tree and shrub species per household increased with increased altitudinal gradient (Fig. 4a, b), indicating that upper and middle altitudes have favorable environmental conditions for plant growth compared to lower altitudes. The observed higher tree species richness in the upper altitude also fits well with the results of several other similar studies (e.g., Abebe 2005; Mengistu 2008; Hadgu et al. 2009), who found higher tree species richness in the upper altitude. Altitude is more related to shrub species richness (r = 0.53) than tree species richness (r = 0.42) (Fig. 4a, b). It is logical that trees can often adapt better to dry environmental conditions than shrub species. In contrast, other studies (e.g., Negash et al. 2012; Yirdaw et al. 2015) have found a declining trend of woody species richness with increased altitudinal gradients in multistrata agroforestry systems of the Southeastern Rift Valley and in the Afromontane forest of the Bale Mountains of Ethiopia, respectively. Such different trends might be caused by several factors such as the variation in altitudinal ranges, differences in topography, slope and soil conditions, and land use and management, among others (Aerts et al. 2006; Duguma and Hager 2010; Negash et al. 2012). For instance, in the study areas the farming systems in the upper and middle altitudes are enset- and tree cereal-based agroforestry systems, while the lower altitude farming system is cereal dominated. Hence, the upper and middle altitudes are characterized by intensive tree management practices compared to the lower altitude. Coupled to these, the presence of more marginal lands and off-farm resources such as fragmented forests in upper altitudes might also be contributing factors (Duguma and Hager 2010; Telila et al. 2015).

Relation between altitude and tree species richness (a) and shrub species richness (b)
The number of tree stems per ha was significantly affected by land use, wealth (P < 0.001) and the first-order interaction of land use × wealth (P < 0.000) (Table 6). However, only land use significantly (P < 0.001) influenced the number of shrub stems per ha. Woodlots had the highest density of tree stems/ha (2465.39) followed by home gardens (237.27), while grazing land and farmlands had the smallest number of tree stems per ha, 41.42 and 14.83, respectively (Table 6). Home gardens also had the highest density of shrub stems/ha (625.51) compared to other land use types (Table 6). The number of tree stems per ha measured in woodlots was lower than that recorded by Abebe (2005). However, the measured density of tree stems per ha was higher than that recorded by Abebe (2005) in the enset-based agroforestry system of Southern Ethiopia. The density of tree stems per ha was also higher than that recorded by Duguma and Hager (2010), but the measured density of shrub stems per ha was lower than that recorded by the same authors in an agricultural landscape of the Suba area, indicating that the density of tree and shrub stems per ha and land use vary depending on land use and management intensity, tree planting patterns and environmental variables.
The presence of higher exotic tree stems per ha in woodlots may imply that farmers' selectiveness in decisions about planting exotic trees in separate plots to minimize tree competition for resources such as light, water and nutrients (Abebe 2005; Duguma and Hager 2010; Agidie et al. 2013). On the other hand, the relatively higher tree and shrub stems in home gardens indicated the farmers' woody planting strategies in different niches such as front yards, hedges and live fences (Duguma and Hager 2010). Farmers also plant exotic trees on farmland boundaries. However, such exotic tree-planting strategies are still debatable because of tree competition for resources against adjacent crops. Several authors (e.g., Jarvis et al. 2000; Negash 2007; Duguma and Hager 2010; Agidie et al. 2013) have acknowledged the influence of tree attributes on farmers’ decisions about tree-planting and tree-management practices.
Herb species richness and the Shannon index were significantly (P ≤ 0.01) influenced by altitude, land use and wealth and by the first-order interaction of wealth × land use and altitude × land use (Table 6). Unlike that of woody species, a higher Shannon index value of herbaceous species was found in farmlands (Table 6). This is explained by the farmers' strategy decisions to diversify food production and minimize risk by evenly allocating available cropland for different crop species.
Influence of household attributes on agro-biodiversity
The potential influence of explanatory factors on woody stems per landholding was analyzed using regression models (GLM with the quasi-Poisson family and log link function). Upper and middle altitudes had positive and strong impacts on trees and shrub stems of farm households. Farm size, which is highly correlated with farmers' economic wellbeing (r = 0.75), positively and strongly influenced tree species richness and tree stem possession (r = 0.496 and r = 0.59), respectively (Table 7; Fig. 5a, b). It is logical that households with large farm size could allocate proportionally more land for growing trees. Land holding was more related to tree stem number per household (r = 0.59) than the number of tree species per household (r = 0.50) (Fig. 5a, b).

The relation between farm size with tree species richness (a) and tree stem/hh (b)
A positive influence of large farm size on tree and shrub species possession and diversity has also been reported by several authors (e.g., Jarvis et al. 2000; Kindt et al. 2004; Abebe 2005; Duguma and Hager 2010).
Family size had strong and positive effects on tree and shrub stem possession (Table 7). The positive effects of family size could be explained by an increasing demand for wood products with increasing family size, which in turn could enhance tree planting. Our findings are also in line with several other studies on the highlands of Ethiopia (e.g., Holden et al. 2003; Duguma and Hager 2010; Mengesha 2010; Abebe 2013) that found a positive influence of family size on woody stem possession and plant diversity.
The educational level of the household head, i.e., number of years spent in school, negatively affected having tree stems, while positively affecting having shrub stems (Table 7). Another study by Duguma and Hager (2010) also revealed the positive effects of education on shrub planting in Central Ethiopia. This indicates that members of educated households might be younger people, who often have small farms. In the study area, a land shortage is the main constraint for tree planting, as explained earlier. The district is the most populated area in the Gurage Zone with a population density of 250 people per square kilometer (WARD 2015).
Off-farm income had a negative impact on tree and shrub stem possession. Our results contradict the findings of Duguma and Hager (2010) and Abebe (2005), who found a positive effect of off-farm income on vegetation density in Central and Southern Ethiopia, respectively. Our results rather reaffirm the findings of several other studies (e.g., Arnold 1987; Shaxson and Tauer 1992) that described the negative effects of off-farm income and labor shortages on managing woody plant species diversity. For instance, in the study area, because of the acute land shortage, some farmers have moved to nearby towns and engage in different off-farm activities and other work as daily laborers for neighboring farmers (e.g., plowing, enset processing); hence, these farmers might not properly manage their own farms because of labor shortages for planting and tending operations.
The age of the household head positively affected shrub stem possession (Table 7). The effects of age could be explained by (1) increasing managerial skill in tending operations with increasing age of the household; (2) older farmers may consider shrub growing an asset that assures livelihood during retirement; (3) with increasing household age, the age of home gardens may also increase, which in turn increases shrub stems (Tolera et al. 2008). The number of grass-roofed houses correlated positively with shrub stem possession and negatively with tree stem household possession. This can be explained in that households with grass-roofed houses were often poor; hence, they need fewer trees, while those with corrugated houses are often rich households with large family size and need more trees for various purposes.
Future prospects of woody plant diversity and possession
E. camaldulensis was the most preferred tree species as it has high domestic value, e.g., as fuel and construction wood and for income generation (Table 8). The fast-growing nature of the tree and seedling availability were also contributing factors. Similarly, Eucalyptus globulus and A. abyssinica species were highly preferred for planting by smallholder farmers in Northern and Central Ethiopia (Achalu 2004; Duguma and Hager 2010; Tefera et al. 2014).
A. abyssinica was the second and C. africana was the third most highly preferred native tree species for different domestic and local uses such as fuel wood and charcoal, timber, soil amendment and income generation through direct selling (Table 8).
D. abyssinica and J. schimperiana were the preferred shrub species by households for three reasons: first, the presence of thorns, providing a defense against intrusion for the formers species; second, the ability to sprout easily after cutting and easy propagation from cuttings for the latter species; third, multiple local uses such as animal feed and live fencing(Table 8). Some authors (e.g., Duguma and Hager 2010 and Ango et al. 2014) noted the influence of plant attributes such as the presence of thorns, vegetative propagation ability and local use value, e.g., as fodder or farmers' preference for woody planting and seedling demand.
The seedling demand assessment showed significant differences among woody species. Higher seedling rates were demonstrated for E. camaldulensis (2820) followed by A. abyssinica (690), D. abyssinica (608), G. robusta (304), R. prinoides (291), C. lusitanica (193), C. africana (114), C. arabica (75) and Persea americana (20). These are the average numbers of seedlings farmers were planning to plant within the time frame of 1–2 years. These results match well with the findings of Achalu (2004), who reported high seedling demand for exotic trees but less for fruit trees in Southern Ethiopia, indicating farmers' preference for planting less space-demanding tree species.
The preference ranking, seedling demand computation and importance value index showed exotic tree dominance, which may indicate continued planting of economically important tree species by smallholder farmers. However, the advantage of planting eucalyptus tree species in agricultural landscapes is debatable because of its alleged negative effects on soil and crops; the species has been rapidly expanding in the agricultural landscape in the highlands of Ethiopia including in the study area. Similar trends have also been reported by several authors in Ethiopia (e.g., Achalu 2004; Duguma and Hager 2010; Tefera et al. 2014), suggesting this trend may replace indigenous tree species in the long term. Hence, development planners may need to design appropriate tree-planting and management strategies to address this trend.
The seedling demands of the farm households was strongly and positively correlated with farm size (r = 0.499**) and livestock holding (r = 0.415*). The average seedling demands of the poor, medium and rich households were 5862, 1610 and 1683 per year, respectively.
Conclusions
Smallholder agricultural landscapes in our study sites act as complementary habitats for maintaining large numbers of native and exotic species. The plant species richness and diversity index were affected by altitude, wealth and land use types. Home gardens, upper and middle altitudes and rich households host increased plant biodiversity. Results of the study also indicated that the number of tree and shrub species increased with increased household location. Likewise, tree species richness and tree stems per household increased with increasing farm size. The importance value index of exotic trees increased with decreasing altitude, while it increased with decreased household wealth status. Other household attributes such as family size, wealth and household location and educational background also positively affected tree and shrub abundance, while off-farm income negatively affected tree and shrub abundance. Species preference ranking, seedling demand and importance value index computations indicated the dominance of exotic tree species, which may indicate continued planting of economically important trees by smallholder farmers. However, the dominance of exotic tree species in farming landscapes may replace the native tree species in the long term. Hence, development planners may need to design appropriate tree planting and management strategies to address this trend.
Notes
A kebele represents the smallest administrative unit in Ethiopia.
References
Abebe T (2005) Diversity in homegarden agroforestry systems of Southern Ethiopia. Dissertation, Wageningen University
Abebe T (2013) Determinants of crop diversity and composition in Enset-coffee agroforestry homegardens of Southern Ethiopia. Agric Rural Dev Trop Subtrop 114:29–38
Abebe T, Wiersum KF, Bongers F (2009) Spatial and temporal variation in crop diversity in agroforestry homegardens of southern Ethiopia. Agrofor Syst 78:309–322
Achalu N (2004) Farm forestry decision making strategies of the Guraghe households, southern-central highlands of Ethiopia. Dissertation, University of Dresden
Aerts R, Van Overtveld K, Haile M, Hermy M, Deckers J, Muys B (2006) Species composition and diversity of small Afromontane forest fragments in north Ethiopia. Plant Ecol 187:127–142. doi:10.1007/s11258-006-9137-0
Agidie A, Ayele B, Wassie A, Hadgu KM, Aynekulu E, Mowo J (2013) Agro forestry practices and farmers’ perception in Koga watershed, upper Blue Nile basin, Ethiopia. Agric For 59:75–89
Ango TG, Börjeson L, Senbeta F, Hylander K (2014) Balancing ecosystem services and disservices: smallholder farmers’ use and management of forest and trees in an agricultural landscape in southwestern Ethiopia. Ecol Soc 9(1):30. doi:10.5751/ES-06279-190130
Arnold JEM (1987) Economic considerations in agroforestry. In: Steppler HA, Nair PKR (eds), ICRAF, Nairobi, pp 173–189
Asfaw Z, Ågren IG (2007) Farmers’ local knowledge and topsoil properties of agroforestry practices in Sidama, Southern Ethiopia. Agrofor Syst 71:35–48. doi:10.1007/s10457-007-9087-0
Baral SK, Malla R, Khanal S, Shakya R (2013) Trees on farms: diversity, carbon pool and contribution to rural livelihoods in Kanchanpur District of Nepal. Banko Janakari 23:1–11. doi:10.3126/banko.v23i1.9462
Bekele AT (2007) Useful trees and shrubs of Ethiopia. ICRAF, Nirobi, pp 1–550
Boffa JM, Turyomurugyendo L, Barnekow-Lillesø JP, Kindt R (2005) Enhancing farm tree diversity as a means of conserving landscape-based biodiversity. Mt Res Dev 25:212–217
Boffa JM, Kindt R, Katumba B, Jourget JG, Turyomurugyendo L (2008) Management of tree diversity in agricultural landscapes around Mabira Forest Reserve, Uganda. Afr J Ecol 46:24–32
CRGE (2010) Ethiopia’s climate-resilient green economy green economy strategy, Addis Ababa
Dessie G (2007) Forest decline in south central Ethiopia extent, history and process. Dissertation, Stockholm University
Duguma LA, Hager H (2009) Forest products scarcity perception and response by tree planting in the rural landscapes: farmers’ views in central highlands of Ethiopia. Ekológia 28(2):158–169. doi:10.4149/ekol_2009_02_158
Duguma LA, Hager H (2010) Woody plants diversity and possession, and their future prospects in small-scale tree and shrub growing in agricultural landscapes in central highlands of Ethiopia. Small Scale For 9:153–174. doi:10.1007/s11842-009-9108-0
Edwards S, Kelbessa E (1999) Forest genetic resources of Ethiopia: status and proposed actions. In: Edwards S, De-missie A, Bekele T, Haase G (eds) The national forest resources conservations strategy development workshop. Proceedings of national workshop from 21–22 June 1999 held in Addis Ababa. Institute of Biodiversity Conservation and Research (IBCR), GTZ, Addis Ababa, pp 101–133
ENMSA (2015) Ethiopian National Meteorological Services Agency Unpublished Rainfall and Temperature data
Food and Agricultural Organisation of the United Nations (FAO) (1994) Provisional soil map of Ethiopia. Land Use Planning Project, Addis Ababa
Food and Agricultural Organization of the United Nations (FAO) (2010) Global forest resource assessment. Country Report Ethiopia. Fra 2010/065. FAO, Rome
Garrity D, Verchot L (2008) Meeting challenges of climate change and poverty through agroforestry. World Agroforestry Centre, Nairobi
Hadgu MK (2008) Temporal and spatial changes in land use patterns and biodiversity in relation to farm productivity at multiple scales in Tigray, Ethiopia. Doctoral Dissertation, Wageningen University
Hadgu KM, Rossing WA, Kooistra L, Van Bruggen AHC (2009) Spatial variation in biodiversity, soil degradation and productivity in agricultural landscapes in the highlands of Tigray, northern Ethiopia. Food Secur 1:83–89
Hailu H, Asfaw Z (2011) Homegardens and agrobiodiversity conservation in Sabata town, Oromia Regional State, Ethiopia. SINET Ethiop J Sci 34(1):1–16
Henry M, Tittonell P, Manlay RJ, Bernoux M, Albrecht A, Vanlauwe B (2009) Biodiversity, carbon stocks and sequestration potential in aboveground biomass in smallholder farming systems of western Kenya. Agric Ecosyst Environ 129:238–252
Holden S, Benin S, Shiferaw B, Pender J (2003) Tree planting for poverty reduction in less-favoured areas of the Ethiopian highlands. Small Scale For Econ Manag Policy 2:63–80
Hurni H (2007) Challenges for sustainable rural development in Ethiopia. Faculty of Technology, Addis Abeba University, Addis Abeba
SPSS Inc (2006) Statistical package for social sciences for windows. Release 20.0.0. SPSS Inc., Chicago
Jagger P, Pender J (2003) The role of trees for sustainable management of less-favored lands: the case of eucalyptus in Ethiopia. For Policy Econ 5:83–95
Jarvis DI, Myer L, Klemick H, Guarino L, Smale M, Brown AHD, Sadiki M, Sthapit B, Hodgkin T (2000) A training guide for in situ conservation on-farm. Version 1. International Plant Genetic Resources Institute, Rome
Kebede M, Kanninen M, Yirdaw E, Lemenih M (2013) Vegetation structural characteristics and topographic factors in the remnant moist Afromontane forest of Wondo Genet, south central Ethiopia. For Res 24(3):419–430
Kindt R, Simons AJ, Van Damme P (2004) Do farm characteristics explain differences in tree species diversity among western Kenyan farms? Agrofor Syst 63:63–74
Kindu M, Schneider T, Teketay D, Knoke T (2013) Land use/land cover change analysis using object-based classification approach in Munessa-Shashemene landscape of the Ethiopian highlands. Remote Sens 5:2411–2435. doi:10.3390/rs5052411
Kindu M, Schneider T, Teketay D, Knoke T (2015) Drivers of land use/land cover changes in Munessa-Shashemene landscape of the south-central highlands of Ethiopia. Environ Monit Assess 187:452
Kindu M, Schneider T, Teketay D, Knoke T (2016) Changes of ecosystem service values in response to land use/land cover dynamics in Munessa-Shashemene landscape of the Ethiopian highlands. Sci Total Environ 547:137–147
Lemenih M (2010) Growing eucalypts by smallholder farmers in Ethiopia. In: Gil L, Tadesse W, Tolosana E, Lopez R (eds) Proceedings of the Conference on Eucalyptus Species Management, History, Status and Trends in Ethiopia, Addis Ababa, Ethiopia, 15–17 September 2010. Ethiopian Institute of Agricultural Research (EIAR), Addis Ababa, Ethiopia, pp 91–103
Lemenih M, Kassa H (2014) Re-greening Ethiopia: history, challenges and lessons. Forests 5:1896–1909
Lemenih M, Teketay D (2006) Changes in soil seed bank composition and density following deforestation and subsequent cultivation of a tropical dry afromontane forest, Ethiopia. Trop Ecol 47(1):1–12
Lemmesa D, Kristoffer H, Peter H (2013) Composition of crops and land-use types in relation to crop raiding pattern at different distances from forests. Agric Ecosyst Environ 167:71–78
Magurran AE (1988) Ecological diversity and its measurement. Croom Helm, London
Mengesha B (2010) Alternative technologies for sustainable agricultural production and agroecosystem conservation in Arsi highlands, southeastern Ethiopia. Dissertation, Addis Ababa University
Mengistu F (2008) Fruit tree species in the wild and in homegarden agroforestry: species composition, diversity and utilization in western Amhara region, Ethiopia. Dissertation, Boku University
Millennium Ecosystem Assessment (2005) Ecosystems and human well-being: synthesis. Island Press, Washington, DC
Nair PKR (1993) An introduction to agroforestry. Kluwer Academic Publisher, Dordrecht
National Meteorological Services Agency of Ethiopia (2015) National Meteorological Services Agency of Ethiopia, Addis Ababa
Negash M (2007) Trees management and livelihoods in Gedeo’s agroforests, Ethiopia. For Trees Livelihoods 17:157–168
Negash N (2013) The indigenous agroforestry systems of the south-eastern Rift Valley escarpment, Ethiopia: their biodiversity, carbon stocks, and litterfall. Dissertation, University of Helsinki
Negash M, Yirdaw E, Luukkanen O (2012) Potential of indigenous multistrata agroforests for maintaining native floristic diversity in the south-eastern Rift Valley escarpment, Ethiopia. Agrofor Syst 85:9–28. doi:10.1007/s10457-011-9408-1
Ranganathan J et al (2008) Ecosystem services: a guide for decision makers. World Resources Institute, Washington, D.C.
Senbeta F, Denich M (2006) Effects of wild coffee management on species diversity in the Afromontane rainforests of Ethiopia. For Ecol Manag 232:68–74
Shannon CE (1949) The mathematical theory of communication. University of Illinois Press, Urbana
Shaxson L, Tauer LW (1992) Intercropping and diversity: an economic analysis of cropping patterns on smallholder farms in Malawi. Exp Agric 28:211–228
Snowdon P, Raison J, Keith H, Ritson P, Grierson P, Adams M, Montagu K, Bi HQ, Burrows W, Eamus D (2002) Protocol for sampling tree and stand biomass. National carbon accounting system, technical report no. 31. Australian Greenhouse Office, Canberra
Tadesse G (2001) Land degradation: a challenge to Ethiopia. Environ Manag 27(6):815–824
Tadesse G, Zavaleta E, Shennan C (2014a) Coffee landscapes as refugia for native woody biodiversity as forest loss continues in southwest Ethiopia. Biol Conserv 169:384–391
Tadesse G, Zavaleta E, Shennan C, FitzSimmons M (2014b) Prospects for forest-based ecosystem services in forest-coffee mosaics as forest loss continues in southwestern Ethiopia. Appl Geogr 50:144–151
R Development Core Team (2010) A language and environment for statistical computing. R Foundation for Statistical Computing Vienna, Austria. http://www.R-project.org/
R Development Core Team (2012) R: a language and environment for statistical com-puting. R Foundation for Statistical Computing. Vienna. ISBN 3-900051-07-0. http://www.R-project.org/. Accessed 30 Sep 2013
Tefera B, Ruelleb ML, Asfaw Z, Abraha BT (2014) Woody plant diversity in an Afromontane agricultural landscape Debark District, northern Ethiopia. For Trees Livelihoods. doi:10.1080/14728028.2014.942709
Teketay D (2001) Deforestation, wood famine, and environmental degradation in Ethiopia's highland ecosystems: urgent need for action. Northeast Afr Stud 8(1):53–76. doi:10.1353/nas.2005.0020
Teketay D, Lemenih M, Bekele T, Yemshaw Y, Feleke S, Tadesse W, Moges Y, Hunde T, Nigussie D (2010) Forest resources and challenges of sustainable forest management and conservation in Ethiopia. In: Bongers F, Tennigkeit T (eds) Degraded forests in Eastern Africa: management and restoration. Earthscan, London, pp 19–64
Telila H, Hylander K, Nemomiss S (2015) The potential of small Eucalyptus plantations in farmscapes to foster native woody plant diversity: local and landscape constraints. Restor Ecol 23(6):918–926. doi:10.1111/rec.12257
Tolera M, Asfaw Z, Lemenih M, Karltun E (2008) Woody species diversity in a changing landscape in the south-central highland of Ethiopia. Agric Ecosyst Environ 128:52–58. doi:10.1016/j.agee.2008.05.001
Yirdaw E, Starr M, Negash M, Yimer F (2015) Influence of topographic aspect on floristic diversity, structure and treeline of Afromontane cloud forests in the Bale Mountains Ethiopia. For Res 26(4):919–931. doi:10.1007/s11676-015-0155-4
WARD (2013) Woreda Rural Redevelopment unpublished annual Report
WARD (2015) Woreda Rural Redevelopment Report unpublished annual Report
Zomer RJ, Trabucco A, Coe R, Place F (2009) Trees on farm: analysis of global extent and geographical patterns of agroforestry. ICRAF Working Paper no. 89. World Agroforestry Centre, Nairobi
Acknowledgements
We owe a particular debt of gratitude to the farmers around the Butajira area who kindly allowed us to use their fields for samples. Special thanks go to our friends Habte Telila and Kiflu Gudeta, who were helpful in utilizing the GIS and R software programs, respectively. We also thank Addis Ababa University for financial support.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Haile, G., Lemenih, M., Senbeta, F. et al. Plant diversity and determinant factors across smallholder agricultural management units in Central Ethiopia. Agroforest Syst 91, 677–695 (2017). https://doi.org/10.1007/s10457-016-0038-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-016-0038-5