
Abstract
The shea tree, Vitellaria paradoxa, shields people, crops and livestock in West African parkland agroforestry systems from climate variability. Accurate estimates of accumulated biomass of such key species may support ways to secure financial incentives within global climate policies. In this quest, variation in allometric relations used for biomass (carbon stock) estimates on the basis of stem diameter matters, but parameters a and b of the standard format (AGB = a Db; AGB = aboveground biomass, D = stem diameter) are correlated and are directly related to tree shapes. Functional branch analysis (FBA) allows non-destructive derivation of allometrics. For Koutiala and Yanfolila shea populations empirical branching parameters were scale-independent, matching FBA assumptions. Allometry (AGB = 169 (D/20)2.64 and AGB = 146 (D/20)2.65, kg tree−1) implied 22%, 16% and 11% larger vegetative aboveground biomass in Koutiala than in Yanfolila at stem diameters (D) of 10, 20 and 32 cm, respectively. Below-ground biomass predictions (BGBi = 8.73 (Di/10)2.35 and BGBi = 8.16 (Di/10)2.38, kg per proximal root) differed − 6% and − 15% for root diameters Di of 10 and 32 cm, respectively. On a dry weight basis, the shoot:root ratio was 2.7 and 2.9, respectively for the two sites. Stand-level above-and below-ground carbon stocks in Koutiala (2.16 ± 0.44 and 0.8 ± 0.15 Mg C ha−1) were not significantly different from those in Yanfolila (3.21 ± 0.60 and 1.26 ± 0.21 Mg C ha−1), respectively. Further research is required to include (potential) fruit production to the plant architectural model.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Parkland agroforestry systems are the dominant woody vegetation in much of the drier parts of West Africa, including southern Mali. These systems are one of the key resilience strategies for smallholder farmers to reduce food insecurity and obtain income, alleviating poverty, through ecosystem services from selectively retained and managed trees. In these parklands Vitellaria paradoxa is the most selected and preserved tree species because of its socio-economic importance (oil, food, wood, fodder, medicine, charcoal, fuelwood, etc.) for rural people (Tom-Dery et al. 2017). In west Africa, the trade of shea nuts and butter contribute to opportunities for industry expansion and the region represents 99.8% of global exports of shea nuts (Agúndez et al. 2020). Nowadays, climate change and human population pressure are threatening tree cover (including shea) in agroforestry systems in the Sahel (Dimobe et al. 2020).
Expectations that the role of agroforestry in the global carbon cycle can translate to financial incentives for land users (in line with efforts for Reducing Emissions from Deforestation and Forest Degradation or REDD++, Minang et al. 2014) has led to increased efforts in refining methods for quantifying the standing biomass, both above and below-ground (Kuyah et al. 2014). The accuracy of biomass estimates with common methods depends on errors in tree diameter frequencies, uncertainty on wood density (Henry et al. 2011) and choice of an allometric equation. Generic equations that use species-specific wood density estimates can over- or underestimate biomass and carbon storage of trees, by a factor of 2 or more (Ekoungoulou et al. 2014).
Empirical data for testing and deriving allometric equations generally require destructive sampling, which involves costs and effort that need to be weighed against the possible gains and benefits from accuracy. In the absence of specific equations for Vitellaria paradoxa, for example, a generic allometric equation has been used to determine its above-ground biomass in previous parkland studies (Saïdou et al. 2012). As previous estimates of biomass of this species were not based on specific allometric equations (Takimoto et al. 2008) and did not attempt to evaluate the below-ground part of the biomass (Saïdou et al. 2012), we designed our study to test the influence of site variation on allometric equations and resulting biomass estimates. As it is not desirable to cut down the trees of this valuable species, the non-destructive method based on characterizing and reconstructing the branching pattern of trees as basis for allometrics called ‘functional branch analysis’ (FBA) was used (Van Noordwijk et al. 2002). Its use depends on when an independence relationship of parent diameters and two branching parameters, p (transfer of cross-sectional area) and q (relative size of largest branch) is found (MacFarlane et al. 2014). Besides being non-destructive, the FBA model separately estimates biomass into different categories (twigs, branches and wood) using standing tree measurements and derives species-specific and mixed-species biomass equations (MacFarlane et al. 2014). FBA can also be applied to estimate root length and root biomass in standing trees based on root diameters at stem base (Van Noordwijk and Mulia 2002).
The objectives of this study were to (1) test the validity of the assumptions of the FBA model as a non-destructive method for deriving allometrics, and (2) to use FBA to develop allometric equations of above and below-ground V. paradoxa carbon stocks at two locations of contrasting climate in the parkland zone.
Materials and methods
Site description and species
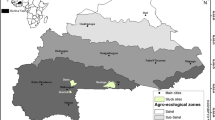
The study was carried out in two districts (Koutiala and Yanfolila) in southern Mali (Fig. 1). Koutiala is located at 12° 38′ N and 5° 66′ W in the Sudano-Sahelian zone and Yanfolila is located at 11° 10′ N and 8° 09′ W in the Sudano-Guinea zone. The rainfall pattern at both sites is unimodal, with no important difference in the length of the rainy season and temperature (3–4 months and 28.0 ± 0.42 °C in Koutiala and 4–5 months and 27.8 ± 0.48 °C in Yanfolila) but the mean annual rainfalls differ with 889 ± 173 mm in Koutiala and 1126 ± 174 mm for Yanfolila.

Location of the study sites in southern Mali, West Africa
Soils at both study sites are characterized by an average bulk density of 1.5 g cm−3 at 0–20 cm soil horizon and by a weak water retention capacity (Traore and Birhanu 2019).
Vitellaria paradoxa C.F Gaertn. (synonym of Butyrospermum paradoxum or parkii) (Naughton et al. 2015), is commonly known as shea butter tree or karité (French), belonging to the Sapotaceae family. The ecological zone of shea trees (V. paradoxa) covers a 500–750 km wide area stretching 6000 km from Senegal/Guinea to South Sudan and Uganda (Naughton et al. 2015).
Field sampling
A total of 70 trees of V. paradoxa were selected in three land-use types (parklands, fallows and protected area) in southern Mali with 30 trees in Koutiala and 40 trees from Yanfolila. As no protected area was found in Koutiala, only parklands and fallows were considered whereas in Yanfolila samples were collected from three land-use types (parklands, fallows and a protected forest). The 70 sample trees were grouped into six classes based on their diameter at breast height (DBH) as following: 3 to 7; > 7 to 10; > 10 to 14; > 14 to 24; > 24 to 32 and > 32 cm. For each individual tree, we aimed at a minimum of 50 measurements but this was not possible for trees with diameter less than 3 cm. Thus, we considered only trees with diameter ≥ 3 cm. The different classes of DBH were built at each 5 cm increment in diameter but some classes presented few individuals and we extended such classes to 10 cm increment. These six classes of DBH have been identified based on data collected on individuals in 200 plots of 2500 m2 (50 m × 50 m) in parklands and fallows, and 5 plots of 1,000 m2 (50 m × 20 m) in protected area in the study sites. Within each plot we recorded the DBH using diameter tape and height using hypsometer (Nikon Forestry Pro). Then, the sample size for each class of diameter was determined by prorating the total sample size of each site to the total number of each diameter class (Table 1).
Field data collection
The data on branching points required for the FBA model calibration were collected in two steps for each sample tree. For the above-ground part, the main stem was considered as the first link and its diameter was recorded at three levels, which are lower diameter (Dproximal), the diameter in the middle of the link (Dmiddle) and the diameter at the distal end of the link (Ddistal). The length of each subsequent link (branch) was recorded with a diameter tape in addition to its link number as well as the parent number of each link. The number zero was attributed to the parent of the main stem with the link number 1, whereas its offspring was 2 and 3 and so on. From the first branch point, the largest branch was selected and followed to a terminal branching point. The measures were repeated on each successive link in the path by climbing on the sample tree (Fig. 2). When 100 measurements were not obtained, this process was repeated on a second or third branch until the target number is reached (Van Noordwijk et al. 2002). The DBH, height of each sample tree and land-use types were recorded. For the below-ground part the root system was excavated and exposed for the sample trees. Thereafter the diameters, link lengths and link number of the roots were measured as for the above-ground part. Data for at least 100 branching points were recorded for each sample tree in above as well as in below-ground parts as recommended by van Noordwijk and Mulia (Van Noordwijk et al. 2002). However, it was not possible to record 100 measures on some sample trees with a small diameter, as a result at least 50 branching points were recorded. On average 2 trees could be measured per day (of 8 h) by a field team of 10 persons.

Measurements for above and belowground biomass of Vitellaria paradoxa in southern Mali, West Africa. a Above-ground biomass measurements of V. paradoxa, b, c exposed roots of V. paradoxa, d below-ground biomass measurements of V. paradoxa, e, f covering of exposed roots
Functional branching analysis model calibration
Our first objective is to test the before applicability of the FBA model by testing for statistical independence of parent diameters and two branching parameters, p (transfer of cross-sectional area) and q (relative size of largest branch). The two parameters can be derived from diameters before and after branching (Van Noordwijk et al. 2002) as:
and
A mean p value should be centered around 1 and explains a wide range of values of cross-sectional area. A q value of 0.5 indicates a perfect fork and higher values that one branch was larger than the others. The value of q is at least 1/n where there are more than 2 links after a branching point. The Eqs. 1 and 2 mentioned above were applied to above-ground as well as for below-ground data. The analysis of variance did not show statistically significant difference between the values of p and q for land-use types within sites, we combined them into a single group in each site to increase sample size. The woody part of the tree was stratified into three diameter categories: twig (link with diameter of less than 2 cm), branch (link between 2 and 10 cm) and wood (link above 10 cm diameter) (MacFarlane et al. 2014). Thus, the FBA model was parametrized for above and below-ground parts of a tree with the following inputs:
-
Average number of branches at each branching point;
-
Link length–diameter relationships;
-
The scaling factor (p);
-
The allocation parameter (q);
-
Tapering coefficient (D2distal/D2prox)/L; where L is the length of link diameter
-
Twigs length (cm);
-
Branch length (cm);
-
Wood length (cm);
-
Wood density (g cm−3).
The wood density used to parametrize this model was derived from twenty trees of V. paradoxa in a previous study at the same sites (Sanogo et al. 2016).
Generation of biomass equation coefficients and carbon stocks estimation
The allometric model derived from FBA based on diameter and allometric scaling used to assess biomass and carbon stocks of the trees is:
where B is biomass (kg tree−1) of the stem or root, D is diameter (cm) at breast height for above and proximal root diameter for below-ground and Dref is a reference diameter used to center the data prior to regression analysis (Tjeuw et al. 2015). Normalization by the mean prior to regression analysis is advised in statistical textbooks to reduce correlation between the parameter estimates and to obtain more meaningful parameter values for a, reflecting the Y value for a tree with D equal Dref. Please note that the default choice of Dref = 1 refers to a tree with diameter of 1 cm, which is generally outside the calibration range.
The parameter b is the allometric exponent. Both parameters a and b were generated with input data for p, q and link lengths through FBA (Van Noordwijk et al. 2002) rules using some stochastic variation within measured parameter ranges. A mean Carbon (C) concentration of dry woody biomass of 0.50 was used for above and below-ground biomass (Basuki et al. 2014). The mean biomass per diameter class was multiplied by the density (ha−1) of trees in this class to obtain its biomass. Biomass per diameter classes was summed up to obtain total biomass per hectare.
Statistical analysis
A pair-wise test was used to compare the carbon stocks between sites using STATA (version 14.1.0). The site-specific allometric equations were evaluated based on the coefficient of determination (R2), standard error of the estimate (SEE), F-values and the root mean square error (RMSE) generated from linear regression analysis.
Results
Tree frequency and empirical diameter–height relationship
The mean DBH and standard deviation of sample trees ranged from 4.6 ± 1.1 to 36.5 ± 2.0 cm in Koutiala whereas the range for Yanfolila was from 5.5 ± 1.4 to 38.0 ± 4.1 cm (Table 1). As a convenient reference diameter Dref for the allometrics a value of 20 cm was selected. Tree height for a given stem diameter was lower in the drier Koutiala site but converged for larger stem diameters (Fig. 3). The allometric relationship for tree height (m) was 5.10 (D/20)0.72 (R2 = 0.96) and 5.58 (D/20)0.61 (R2 = 0.90), for Koutiala and Yanfolila, respectively.

Shea tree diameter-height relationships in Koutiala (a) and Yanfolila (b) in southern Mali, West Africa
Applicability of FBA for Vitellaria paradoxa
In total, 3075 and 2857 branching points were observed on 30 trees in above- and below-ground in Koutiala, respectively whereas 3986 and 3761 branching points on 40 trees in above- and below-ground in Yanfolila, respectively. The p and q derived from each branching point were plotted against parent diameters (Figs. 4, 5). The results revealed a very low R2 values (ranging from 0.01 to 0.06 for p and from 0.001 to 0.02 for q) of the relationship between p and q in above and below-ground at both sites and showed that the null-hypothesis of independence could not be rejected. This implies that the FBA model is applicable for our studied species (V. paradoxa) as it suggests self-similarity across multiple scales.

Relationship between parent diameter and p and q values in Koutiala (a, c) and Yanfolila (b, d) for Vitellaria paradoxa in above-ground in southern Mali, West Africa

Relationship between root diameter and p and q values in Koutiala (a, c) and Yanfolila (b, d) for Vitellaria paradoxa in below-ground in southern Mali, West Africa
Branching parameters inputs for FBA model
Table 2 shows the tree parameters for above as well as below-ground biomass in the study sites revealing some similarities in the values between parameters of these two compartments. In both Koutiala and Yanfolila, the average number of branches per branching point in above-ground (2.2 vs 2.5) and below-ground parts of trees (2.3 vs 2.65) are of the same order of magnitude. The p value is lower for above than below-ground, which indicated the link cross-sectional area before branching was smaller than the sum of the cross-sectional areas of links after branching point. The tapering coefficient of V. paradoxa was higher in above-ground at each site compared to that of the below-ground.
The average link lengths derived from each tree components (twig, branch and wood) for above and below-ground with length values of the same magnitude were used in FBA. These link lengths increase from the twigs to the wood in above-ground for V. paradoxa. The length–diameter relationship for tree components (twig, branch and wood) was not linear (Table 3). Therefore, the parametrization did not use an average value for link length for all components but used an average link length for each component for above and below-ground. The wood density was included for all components and this value was 0.72 (± 0.03) in Yanfolila and 0.74 (± 0.01) in Koutiala (Sanogo et al. 2016).
Ratio of cross-sectional area of the stem and sum of proximal roots
The ratios of cross-sectional area of the stem and sum of proximal roots varied little according to site. In the Sudano-Sahelian zone (Koutiala) with lower rainfall, the value of the ratio of cross-sectional area (CSA) of the stem and sum of proximal roots was 0.68 whereas for the Sudano-Guinean zone (Yanfolila) the value was 0.70. Figure 6 shows that the sum of proximal roots explained 92% (R2 = 0.92) of the variations in the CSA of the stem in Koutiala and 91% (R2 = 0.91) in Yanfolila.

Relationship between cross-sectional area of the stem and sum of proximal roots of V. paradoxa in Koutiala (a) and Yanfolila (b) in southern Mali, West Africa
Fractional branch analysis outputs and allometric equations
The power coefficient b of allometrics for above-ground biomass (2.64 and 2.65) was greater than that for root biomass (2.35 and 2.38) in Koutiala and Yanfolila, respectively (Table 4). The full equations for above-ground biomass (kg tree−1) of V. paradoxa were AGB = 169 (D/20)2.64 and AGB = 146 (D/20)2.65 and below-ground biomass (kg root−1) were BGBi = 8.73 (Di/10) 2.35 and BGBi = 8.16 (Di/10)2.38 in Koutiala and Yanfolila, respectively.
Based on the allometric equations developed above, the carbon stocks of below-ground biomass estimates were depicted against the stem diameter (Fig. 7). The relationship between below-ground carbon stocks and stem diameter was strong and accounted for 96% of the variance (Y = 34.7 (D/20)2.1212; R2 = 0.96; n = 30, p < 0.0001) and 94% (Y = 33.8 (D/20)2.1426; R2 = 0.94; n = 40, p < 0.0001) of the variation in the below-ground carbon stocks in Koutiala and Yanfolila, respectively (Fig. 7). Therefore, the stem diameter could be a good predictor for below-ground carbon stocks for V. paradoxa in the study sites. Table 5 displays the carbon stocks derived from the allometric equations developed for above and below-ground. A pair-wise test revealed no significant difference (p > 0.05) between the two sites for carbon stock in above-ground of V. paradoxa as well as for the below-ground (Table 5).

Relationship between below-ground carbon stocks and stem diameter of V. paradoxa in Koutiala (a) and Yanfolila (b) in southern Mali, West Africa
Ratio of below- to above-ground carbon stocks of Vitellaria paradoxa
The root to shoot ratio (R:S) for below-ground biomass and carbon stocks were 0.36 and 0.39 in Koutiala and Yanfolila, respectively. The above-ground carbon stocks accounted for 90% (R2 = 0.90) of the variation in the below-ground carbon stocks in Koutiala and 89% (R2 = 0.89) in Yanfolila (Fig. 8).

Relationship between above-ground and below-ground carbon stocks of Vitellaria paradoxa in Koutiala (a) and Yanfolila (b) in southern Mali, West Africa
Fractional branching analysis model validation
The regression coefficient was used to examine the strength and significance of the models. The linear model of the data for the above and below-ground carbon stocks against the diameter at breast height/proximal root diameter has yield R2 values (Table 6) ranging from 0.87 to 0.93 between the carbon stocks and the diameters at both sites indicating that the carbon prediction model provides a good fit to the data. Moreover, the standard error of the estimate (SEE) was 0.051 (5.1%) in Koutiala and 0.014 (1.4%) in Yanfolila for above-ground carbon stocks. The results showed that the F-value for the regression model is significantly different from zero F (1, 28) = 217.17, p<0.0001 in Koutiala and F (1, 38) = 355.57, p<0.0001 in Yanfolila for above-ground carbon stocks which indicated a positive linear relationship between the carbon and diameter. Similar results (high value of R2, low standard error of the estimate and F different from zero) were observed for the below-ground carbon stocks and the Root Mean Square Error (RMSE) values are low which validate the models at both sites (Table 6). Moreover, the high R2 of all the regressions (Table 6) suggested that the stem diameter could be used as a predictor variable for carbon stocks.
In addition to the mentioned above, Fig. 9 compares above-ground biomass estimation by equation AGB = 168.7 (D/20)2.64 in the site of Koutiala (a) and AGB = 145.8 (D/20)2.65 in the site of Yanfolila (Table 6) to existing equation (Chave et al. 2005) for dry forest. The equation (Chave et al. 2005) of AGB = 0.112 × (ρ DBH2 H)0.916 in Fig. 9 consistently underestimated the biomass of V. paradoxa.

Scatter plots of above-ground biomass (AGB) predicted by developed equations in the site of Koutiala (a) and Yanfolila (b) and the equation (Chave et al. 2005) for dry forests
Discussion
The lack of relationship between p and q with parent diameter/root diameter confirmed scale independence of branching and hence the applicability of FBA model to V. paradoxa. Similar results were found for Acacia mearnsii, Eucalyptus saligna and Mangifera indica in Kenya (Macfarlane et al. 2014). The average of p in above-ground was 1.01 ± 0.83 and 1.05 ± 1.03 against 1.17 ± 1.13 and 1.27 ± 1.10 in below-ground in Koutiala and Yanfolila, respectively. Values of similar magnitude of p (1.7 ± 0.4; 1.8 ± 0.4 and 1.8 ± 0.4) were found in Acacia angustissima, Gliricidia sepium and Leucaena collinsii, respectively (Koanga 2012). The value of q in below-ground in Koutiala is less than that of Yanfolila, which could be related to soil characteristics, rainfall and land-use management practices. This result is in accordance with the findings of previous studies (MacFarlane et al. 2014). Furthermore, the mean link length in above-ground increases with link diameter class for different components (twig, branch and wood) of V. paradoxa agreeing with MacFarlane et al. (2014) and Tjeuw et al. (2015).
The values of the scaling coefficient b for above-ground biomass (2.65 and 2.64) were close to the supposedly universal value 8/3 (= 2.67) (Enquist 2002), and match the expectation of Ketterings et al. (2000) that they equal 2+ the power of the diameter–height relationship (0.72 and 0.61 for the two sites). The results also revealed variation of b factor among the tree components (twig, branch and wood). Allometric equations for branch biomass had a higher value of b than those for total biomass and twig biomass. This result agrees with the FBA findings of Tjeuw et al. (2015). Our results revealed that the ratio of above-ground and below-ground carbon stocks increase with increasing stem diameter, corroborating results of Jibrin and Abdulkadir (2015).
Given the uncertainty in tree diameter frequency data, the stand-level above-ground carbon stock (3.21 ± 0.60 Mg C ha−1) in Yanfolila could not be statistically differentiated (p > 0.05) from that of the value (2.16 ± 0.44 Mg C ha−1) recorded in Koutiala. Our parkland values, considering only V. paradoxa and not including other species in the same vegetation, are lower than those of Kuyah et al. (2012) who reported 17 ± 0.02 Mg C ha−1 in above-ground of plantations (Dorstenia panamensis) in agricultural landscapes in Kenya. Other estimates in the literature for southern Mali are 22.2 to 70.8 Mg C ha−1 (Luedeling and Neufeldt 2012), 22.4 to 54.0 Mg C ha−1 (Takimoto et al. 2008), and 24 Mg C ha−1 (Traoré et al. 2004). Furthermore, carbon stocks in these studies were estimated using generic allometric equations, which are prone to overestimation. Our figures are close to those reported by Peltier et al. (2007) in Cameroon (5.05 Mg C ha−1 above-ground biomass for V. paradoxa) whereas they are smaller than those of Saïdou et al. (2012) in Benin (20.17 Mg C ha−1 for V. paradoxa). As for below-ground carbon stocks of V. paradoxa there was also no significant difference (p > 0.05) between Yanfolila (1.26 ± 0.21 Mg C ha−1) and Koutiala (0.80 ± 0.15 Mg C ha−1). These values are lower than the value recorded (4.25 ± 0.74 Mg C ha−1) by Saïdou et al. (2012). These differences of values with the current study may be explained by the difference of allometric equations used, tree diameter, tree density per hectare and climatic conditions.
The ratio of below- to above-ground biomass of 0.36 in Koutiala and 0.39 in Yanfolila were higher than those (0.26–0.28) reported by Henry et al. (2011) and Kuyah et al. (2014). They are within the range of root to shoot (R:S) recommended by IPCC (2006) for subtropical dry forests and comparable to the range from 0.35 to 0.5 for woodland of Mozambique (Ryan et al. 2011) and Jatropha curcas in Indonesia (Tjeuw et al. 2015).
Conclusions
The present study provides new insights into the biomass estimation in above and below-ground by developing site-specific allometric equations for V. paradoxa. The established allometric models can significantly improve the accuracy of carbon stocks estimation of V. paradoxa in West Africa Savanna for REDD++ activities. In addition, our models can be applied to V. paradoxa on sites with similar environmental conditions. The carbon stocks in both above-ground and below-ground did not vary with sites in southern Mali in the range of climatic conditions sampled by our study.
Strong linear relationships were observed between carbon stock (above and below-ground) and stem diameter or proximal root diameter with a low SEE as well as the low RMSE and F-values significantly different from zero, which indicates that the models are valid. Fruit biomass was not included in the model so further research is required on fruit to fine-tune the model. For the many other species with no specific allometric equations, we recommend the elaboration of site-specific equations using FBA, with increased understanding of where and how species differ in allometry.
References
Agúndez D, Nouhoheflin T, Coulibaly O, Soliño M, Alía R (2020) Local preferences for shea nut and butter production in Northern Benin: preliminary results. Forests. https://doi.org/10.3390/f11010013
Basuki MT, van Laake EP, Skidmore KA, Hussin AY (2014) Allometric equations for estimating the aboveground biomass in tropical lowland Dipterocarp forests. For Ecol Manag 257: 1684–1694
Chave J, Andalo C, Brown S, Cairns MA, Chambers JQ, Eamus D, Folster H, Fromard F, Higuchi N, Kira T, Lescure JP, Puig H, Riera B, Yamakura T (2005) Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 145:87–99. https://doi.org/10.1007/s00442-005-0100-x
Dimobe K, Ouédraogo A, Ouédraogo K et al (2020) (2020) Climate change reduces the distribution area of the shea tree (Vitellaria paradoxa C.F. Gaertn.) in Burkina Faso. J Arid Environ 181:104237. https://doi.org/10.1016/j.jaridenv.2020.104237
Ekoungoulou R, Liu X, Loumeto JJ, Ifo SA (2014) Tree above and belowground biomass allometrics for carbon stocks estimation in secondary forest of Congo. JSTFT 8:9–20
Enquist BJ (2002) Universal scaling in tree and vascular plant allometry: towards a general quantitative theory linking plant form and function from cells to ecosystems. Tree Physiol 22: 1045–1064
Henry M, Picard N, Trotta C, Manlay RJ, Valentini R, Bernoux M, Saint-André L (2011) Estimating tree biomass of sub-Saharan African forests: a review of available allometric equations. SF 45: 477–569
IPCC (2006). In: Eggleston S, Beundia L, Miaw K, Ngara T, Tanabe K (eds) IPCC guidelines for national greenhouse inventories. Volume 4. Agriculture, forestry and other land use. Intergovernmental Panel on Climate change (IPCC), IPCC/IGES, Hayama, Japan, pp 48–49
Jibrin A, Abdulkadir A (2015) Allometric models for biomass estimation in Savanna Woodland Area, Niger State, Nigeria. Int J Environ Ecol Geol Geophys Eng 9:209–217
Kaonga LM (2012) Fractal analysis of canopy architectures of Acacia angustissima, Gliricidia sepium, and Leucaena collinsii for estimation of aboveground biomass in a short rotation forest in eastern Zambia. J For Res 23:1–12
Ketterings QM, Coe R, Van Noordwijk M, Ambagau Y, Palm CA (2000) Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. For Ecol Manag 146:199–209
Kuyah S, Dietz J, Murthuri C, Jammadass R, Mwangi P, Coe R, Neufeldt H (2012) Allometrics equations for estimating biomass in agricultural landscapes: belowground biomass. Agric Ecosyst Environ 158:225–234
Kuyah S, Dietz J, Muthuri C, van Noordwijk M, Neufeldt H (2014) Allometry and partitioning of above-and below-ground biomass in farm eucalyptus species dominate in Western Kenyan agricultural landscapes. Biomass Bioenergy 55:276–284
Luedeling E, Neufeldt H (2012) Carbon sequestration potential of parkland agroforestry in the Sahel. Clim Change 115:443–461
MacFarlane DW, Kuyah S, Mulia R, Dietz J, Muthuri C, Van Noordwijk M (2014) Evaluating a non-destructive method for calibrating tree biomass equations derived from tree branching architecture. Trees 28(3):807–817
Minang PA, Duguma LA, Bernard F, Mertz O, van Noordwijk M (2014) Prospects for agroforestry in REDD+ landscapes in Africa. Curr Opin Environ Sustain 6:78–82
Naughton CC, Lovett PN, Mihelcic JR (2015) Land suitability modeling of shea (Vitellaria paradoxa) distribution across sub-Saharan Africa. Appl Geogr 58:217–227
Peltier R, Njiti FC, Ntoupka M, Manlay R, Henry M, Morillon V (2007) Evaluation du stock de carbone et de la productivité en bois d’un parc à karités du Nord-Cameroun. Revue Bois et Forêt des Tropiques 294:39–50
Ryan CM, Williams M, Grace J (2011) Above- and belowground carbon stocks in a Miombo woodland landscape of Mozambique. Biotropica 43:423–432
Saïdou A, Dossa EFA, Gnangle CP, Balogoun I, Aho N (2012) Evaluation du stock de carbone dans les systèmes agroforestiers à karité (Vitellaria paradoxa C.F. Gaertn.) et à néré (Parkia biglobosa Jacq. G. Don) en zone Soudanienne du Bénin. Agric Forêt 11:1025–2325
Sanogo K, Gebrekirstos A, Bayala J, Villamor GB, Kalinganire A, Dodiomon S (2016) Potential of dendrochronology in assessing carbon sequestration rates of Vitellaria paradoxa in southern Mali, West Africa. Dendrochronologia 40:26–35
Takimoto A, Nair PKR, Nair VD (2008) Carbon stock and sequestration potential of traditional and improved agroforestry systems in the West African Sahel. Agric Ecosyst Environ 125:159–166
Tjeuw J, Mulia R, Slingerland M, Van Noordwijk M (2015) Tree or shrub: a functional branch analysis of Jatropha curcas L. Agrofor Syst 89:841–856
Tom-Dery D, Eller F, Reisdorff C, Jensen K (2017) Shea (Vitellaria paradoxa CF Gaertn.) at the crossroads: current knowledge and research gaps. Agrofor Syst. https://doi.org/10.1007/s10457-017-0080-y
Traoré K, Ganry F, Olivier R, Gigou J (2004) Litter production and soil fertility in a Vitellaria parkland in a catena in southern Mali. Arid Land Res Manag 18:359–368
Traore K, Birhanu Z (2019) Soil erosion control and moisture conservation using contour ridge tillage in Bougouni and Koutiala, Southern Mali. J Environ Prot 10:1333–1360
Van Noordwijk M, Mulia R (2002) Functional branch analysis as tool for scaling above and belowground trees for their additive and non-additive properties. Ecol Model 149:41–51
Van Noordwijk M, Rahayu S, Hairiah K, Wulan YC, Farida A, Verbist B (2002) Carbon stock assessment for a forest-to-coffee conversion landscape in Sumber-Jaya (Lampung, Indonesia): from allometric equation to land use change analysis. Sci China Ser C Life Sci 45:75–86
Acknowledgements
This research was funded by the West African Science Service Center on Climate Change and Adapted Land Use (WASCAL) program; the International Foundation for Science (IFS) and the Committee on Scientific, Technological Cooperation of the Organization of Islamic Conference (COMSTECH) (Grant Number NO. D/5618-1), and Forest, Trees and Agroforestry (FTA) CRP, which are gratefully acknowledged. We extend our thanks to the staff of the World Agroforestry (ICRAF)- Mali office for logistical support during the fieldwork and supervisory work. We thank the farmers for allowing us to work in their fields.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Sanogo, K., Bayala, J., Villamor, G.B. et al. A non-destructive method for estimating woody biomass and carbon stocks of Vitellaria paradoxa in southern Mali, West Africa. Agroforest Syst 95, 135–150 (2021). https://doi.org/10.1007/s10457-020-00578-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-020-00578-3