
Abstract
West African livestock keepers need forage sources to feed animals without degrading the local fragile ecosystems. Establishing high-density forage protein banks for animal feeding was evaluated in three contrasting zones of Burkina Faso. The agronomic performance of the fodder species Leucaena leucocephala (leucaena) and Morus alba (mulberry), planted for intensive forage production (25,000 plants/ha) was monitored through plant height (HEI), stem diameter (DIA), branch number (BrN) and branch length (BrL). Once plots were considered established (at 7 months) all trees were coppiced. Biomass yield subsequently was monitored over three bi-monthly harvests which began approximately 1 year after planting. For both species, a highly significant interaction between experimental site and dates was observed for all variables except BrN. Average HEI (103 cm) were similar. DIA (7 ± 0.4 cm) and BrL (55 ± 2.2 cm) were higher in mulberry (6 ± 0.1 and 29 ± 1.0 cm, respectively), whereas the BrN remained higher in leucaena (7 ± 0.2 vs. 2 ± 0.1 cm). No differences were observed in HEI between sites from June to August. However, from September to October, both species displayed a stronger growth rate at site 1. The biomass yields (t DM/ha/year) in three consecutive harvests were lower than values reported in other tropical regions (1.8 ± 1.2 and 8.1 ± 3.9, for mulberry and leucaena, respectively). Differences between sites were due mainly to soil properties, rainfall pattern and termite attacks. This study demonstrated that L. leucocephala and M. alba, planted in high-density forage banks, successfully establish and have high forage yield across several edaphoclimatic conditions in Burkina Faso. Our findings suggest that this technology could significantly contribute to livestock feeding systems in the region, thus reducing negative effects of over-grazing on landscape degradation.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
In Africa, the main challenge for making progress in livestock production is a lack of productive and locally affordable feeding systems. Current dominant agro-pastoral systems are based on extensive grazing of native grasslands which are characterized by seasonality, low productivity and variable nutritive value (Khan and Habib 2012). In other tropical regions, such as Latin America, the Caribbean, and Asia, experience with alternative systems exists. One strategy is a type of agroforestry where various strata and species of multipurpose trees and shrubs at varied densities and arrangements are included on the farm. These technologies are widely recognized and accepted because of their positive economic and environmental impacts across a large spectrum of situations (Franzel et al. 2014; Murgueitio et al. 2015).
Even if shrubs and trees are present in African agricultural landscapes, their presence is in most cases deliberate and planted at relatively low densities for living fences, alley cropping, and/or soil fertility or soil erosion control purposes (Bationo et al. 2012; Bayala et al. 2014; Franzel et al. 2014). In West Africa, some fodder species are informally wild-harvested and used to feed animals during the dry season when availability of green natural grasses start to decline.
However, ecosystems in this region are fragile and trends in the Sahelian and Sudanian woody vegetation are controversial. Some authors report a steady decline and a vegetation impoverishment despite resource protection measures and recovery programs (Paré et al. 2010; Herrmann and Tappan 2013), indicating an overall reduction in woody species richness, a loss of large trees, an increasing dominance of shrubs, and a shift towards more arid-tolerant, Sahelian species since 1983 (Herrmann and Tappan 2013). Other authors, however, document a greening trend over the past 30 years in semi-arid West Africa (Kusserow 2017). The assertion of the vicious cycle of the Sahel crisis prompted by population pressure and low rainfall is now replaced by an adaptive cycle theory, with the traditional picture of a unidirectional process of land degradation and system collapse in Sahelian agro-pastoral systems considered as a simplification of more complex realities (Vang Rasmussen and Reenberg 2012). Brandt et al. (2016) argued that this variability is driven by precipitation, independent of the human dimension and argued that land use modalities provide greater insights on the spatial changes. Nevertheless, the main causes affecting the complex trends of woody vegetation in these ecosystems are droughts, savanna clearing to expand cropped lands or to meet the increased demand for charcoal and wood-fuel in urban centers (Brandt et al. 2016). In this context, among the main land use modalities we can mention the livestock production, with predominance of extensive transhumant systems reliant upon available herbaceous and woody vegetation. During the dry season, overgrazing is frequent due to a significant lack of biomass for animal feeding. Filling this gap is highly critical both to improve livestock production and to avoid negative effects of cattle on ecosystems.
Among existing technologies that might be used to increase forage production and improve farm system autonomy, high-density forage protein banks (PB) is one alternative that has produced good results in different regions including Cuba, Costa Rica and others in Asia and Latin-America (Datta 2000; González-García and Martín-Martín 2016; Mullen et al. 2003; Saddul et al. 2004). To date, the prevalence of these PB in West Africa or Burkina is not well documented.
The objectives of this study were to: (1) test the feasibility of establishing PB for livestock feeding under the constrained conditions of Burkina Faso; (2) study the agronomic performance during the first year of establishment of two recognized woody fodder species (one leguminous: leucaena, Leucaena leucocephala; and one non-leguminous: mulberry, Morus alba) planted for intensive forage production; and (3) evaluate the effects and differences between three contrasting edaphoclimatic conditions on the main morphagronomic parameters evaluated. To do this, the successful experiences of establishing this technology in Latin America and, more specifically Cuba, were extrapolated, with the hypothesis that these systems could be adapted to West Africa.
Materials and methods
Edaphoclimatic conditions of the region
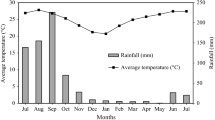
In order to fit within the scope of the STRADIV project (system approach for the TRAnsition to bio-DIVersified agroecosystems this study was developed in western Burkina Faso. The work was conducted in Houet and Tuy provinces (Fig. 1), both characterized by a Sudanese climate and a unimodal rainfall pattern. The rainy season occurs from mid-May to late October with a precipitation regime ranging between 800 and 1000 mm per year. Average temperatures are between 23 and 35 °C, and relative humidity is between 27 and 80% (Fig. 2). During the 2 years of the study, the most abundant rains were registered in August for the three sites during the first year, whereas for the second year it was in May, June and August in sites 1, 2 and 3, respectively (Fig. 2). The dry season starts after the last remaining rains of late October, and finishes with the first little rains at late April or early May of the following year (Fig. 2). Beyond the scarcity of water, dry seasons are also characterized by lower relative atmospheric humidity (~ 40%), high temperatures from January to April and lower temperatures from October to January (Fig. 2).

Localization of the protein banks established in three contrasting sites belonging to the region of Hauts-Bassins (Bobo-Dioulasso), in the west of Burkina Faso (West Africa)

Average monthly temperatures, rainfall and relative humidity during the monitoring year on each experimental site (site 1 = PB1, site 2 = PB2 and site 3 = PB3)
In this region of Sahel, the main limiting factors for developing agriculture are the sandy soils and the lack of water. The vegetation mainly consists of wooded and grassy savanna, dominated by various types of steppe (grassy, shrubby, wooded), woodland and forest galleries along watercourses. Typical Saharan and Sahelien species present in the landscape include Acacia ehrenbergiana, A. raddiana, Grewia tenax, Maerua crassifolia, Andropogon gayanus var., Tridentatus, Aristida stipoides, Hyphaene thebaica (“palmier doum”) and Cenchrus sp., etc.
The main crop systems leading the agriculture in the region of the study are cotton, millet, maize, sorghum, mucuna, peanut, and cowpea (Vigna unguiculata). Livestock are mostly informally fed and/or supplemented when available with good fresh or dried mucuna, peanut, or cowpea leaves and crop residues such as maize, sorghum, millet or bush straws, cotton hulls. The most used supplements locally are cotton cakes or other cereals straws.
The hydrographic network in this region (Hauts-Bassins) of Bobo-Dioulasso is organized around two basins: The Kou and the Tuy sub-basins. There were no irrigation systems for the three experimental sites used in the study.
Particularities of the experimental sites
Three contrasting locations were selected for the study where mixed crop-livestock production systems already were established. The site selection criteria were: (1) soil type and properties; (2) differences in rainfall pattern and other climatological parameters such as humidity or temperature. Thus, the experimental plots were located on plain reliefs at altitudes that reached 386 m above sea level (m.a.s.l.).
The experimental site 1 (PB1; 11°19″42″N, 4°13″25″W; 386 m.a.s.l.) is located 10 km north of Bobo-Dioulasso. Site 2 (PB2; 11°18″5″N, 3°44″48″W; 309 m.a.s.l.) is in Tuy province (Koumbia, 67 km from Bobo-Dioulasso), while site 3 (PB3; 11°13″56″N, 4°27″23″W; 386 m.a.s.l.) is located at Camp Peul Nasso, in the northwest of Bobo-Dioulasso. Details of soil properties and rainfall pattern for each experimental site appears in Table 1 and Fig. 1, respectively, and differences are considered when discussing the experimental results. The main differences between the experimental sites are the soil characteristics (i.e. more clay and silt particles, and less available P and pH in PB3; and sandy soils in PB1 and PB2 with higher concentrations of available P and pH).
Before starting the experiment, soil samples were collected on each experimental site for analysis of chemical and physical characteristics. Samples were representative of different points of the plot and at two horizons i.e. superficial layer (0–20 cm) and a deeper layer (20–40 cm). Soil analyses were completed and the available phosphorus was determined according to the method proposed by Bray and Kurtz (1945). Soil characteristics for the three experimental sites are summarized in Table 1.
The farmers who volunteered to participate in the trials needed to have a plot that they committed to establish the forage bank and they must have all required rights for conducting the agronomic activities in those areas. They were asked that the plot be as close as possible to the family home, the cattle stall and to a water source.
Field activities and protein bank design
Prior to study establishment, we acquired seed for each fodder species (leucaena and mulberry) and for the living fences (Gliricidia sepium, Albizia lebbeck, and Samanea saman) to be planted on the perimeter of each paddock. Nurseries were prepared to develop the plant material while land for each experimental site was prepared for fodder and living fence establishment. Planting was coordinated based on rains and labor availability and a robust monitoring and evaluation system was instituted to manage the sites and measure plant growth (Fig. 2).
In the three farms, a common identical plot size (625 m2) and field design was considered for planting and establishing the protein banks (Fig. 2). The plant density was 20,000 plants per hectare (plantation framework of 1 m between rows and 0.5 m between plants).
Seeds of the mulberry (M. alba cv YU-62), and the majority of leucaena (L. leucocephala) and gliricidia (G. sepium) plantings were acquired from the Experimental Station Indio Hatuey in Cuba. Seeds of albizia (Albizia lebbeck) and samanea (Samanea saman) were bought from the National Centre of Forest Seed (CNSF) or from the Association for the Promotion of Fertilizing Trees, Agroforestry and Forestry Trees (APAF), both located in Ouagadougou (Burkina Faso).
Seeds were planted and seedlings developed under specific, controlled conditions during the nursery phase, which was carried out in a pre-determined site in Bobo-Dioulasso. Seedlings were grown in small pots (50 cl at the start and 1.5 L once the plants started to grow) filled with 50% soil and 50% compost. No physical or chemical seed treatments were used during the whole nursery phase, which lasted from late December until the planting dates (Fig. 4). In this nursery phase, the non-scarified seeds emerged at different stages after sowing, depending on the species (i.e. 4 days after for leucaena and gliricidia, 7 days for samanea, 8 days for albizia, and 2 weeks for mulberry).
Plot conditioning and arrangement
Concurrent to the nursery phase, conventional soil preparation started at each site using animal traction. Two tillage activities were carried out at each site: the first in late May or early June when the first rains started, and the second one a few days before the planting date. Manual work was required to finish adequate individual plant holes (20–30 cm depth) according to the forage bank design. Each experimental site (or village) was considered as a replicate with identical forage bank plots established. Each plot (on each site) was divided into two uniform blocks (one per fodder species i.e. one for mulberry and the other for leucaena; Fig. 3). Thus, a total of 3 similar plots with 6 sub-plots (1 per fodder specie per site) was planted and used for the study.

Field designs of the experimental plots. Bloc 1 (Morus alba) and Bloc 2 (Leucaena leucocephala). A living fence was also planted and established all around each forage bank. M = mulberry (Morus alba), L = leucaena (Leucaena leucocephala), A = albizia (Albizia lebbeck), G = gliricidia (Gliricidia sepium), S = samanea (Samanea saman)
The plantings were made during the rainy season according to the planting schedule (Fig. 4). To prepare for planting, approximately 150 kg of dry cattle manure was applied on each experimental plot to enhance the startup conditions for in-field plant growth.

Schematic representation of the experimental design, including the dates of monitoring visits for performing the plot measurements. An establishment and an exploitation (harvest) periods were included. The PB1, PB2, PB3 = protein banks on experimental sites 1, 2 and 3, respectively
Experimental design and monitoring schedule
Measured variables and sampling dates
At each experimental site, a representative sample of 10% of the plants per species (that is 50 mulberry and 50 leucaena) was selected on the planting date, and individually identified and labelled for monitoring purposes, scheduled from the date of planting until 7 months after, when the main variables of plant growth attained a plateau (and the PBs were thus considered established). Overall, data were accumulated for each experimental site from July (first field monitoring visit) until January of the following year (last plant growth monitoring visit date considered for this study). A total of eight visits (two in August) were performed for plant growth monitoring. Three forage harvests occurred solely at the PB2 and PB3 sites (Fig. 4), due to an accidental wildfire that occurred at PB1 following establishment.
Four plant growth and development variables were monitored: plant height (HEI), stem diameter (DIA), branch number (BrN) and branch length (BrL). The plant HEI and the length of the main branches (BrL; up to the middle of the crown) were measured with a ruler and expressed in centimeters. The number of primary branches was counted plant by plant and expressed in units. The DIA was measured at half-height by using a caliper gauge and expressed in millimeters.
For each location, the climatological variables monitored throughout the experiment were temperature (°C) as well as precipitation (mm) and relative humidity (%). At each field visit, plants with damage and plant mortality were recorded for the entire plantation. Pest (e.g. termite) and disease attacks were expressed and recorded as the percentage of the total plant population for each forage species.
Biomass yield after protein bank establishment
Once the PBs were considered established (i.e. 7 months after planting date), a standardization harvest was carried out in the whole plot and for both species at the height of 1.20 m with the aim of getting ready for the “forage exploitation” phase, after standardizing the plot and eliminating the eventual heterogeneity in plant sizes. Thereafter, the monitoring of biomass yield commenced and was carried out during the first three harvests scheduled during the exploitation phase for animal feeding and made at 45–50 cm of the plant height (from the ground level). Total biomass production was measured by weighing the total forage produced in the whole PB area at each harvest. The results are expressed in dry matter basis as fresh forage samples and were immediately sent to the laboratory after each harvest and dried at 60 °C for 48 h in a forced air stove.
Calculations and statistical analyses
An excel database was built for this study with individual sheets of a common structure corresponding to each experimental site (PB1, PB2 and PB3). Further statistical analyses were carried out using the Statistical Analysis System package (SAS; v. 9.1.3. 2002–2003 by SAS Institute Inc., Cary, NC, USA). A two-step statistical analyses approach was followed. In a first step, the main fixed effects were experimental site, fodder specie, monitoring date (month) and their first-order interactions. Effects on the variables (i.e. HEI, DIA, BrN, BrL and biomass yield) were evaluated. Data were analyzed using the PROC MIXED function with repeated measures. The least-squares means separation procedure using the PDIFF option in SAS was used and the statistical model was as follows:
where Yijk is the plant measurement at date k, for the fodder specie j at the site i, µ is the overall mean, Sitei is the fixed effect of the specific experimental site i (i = 1–3), Speciej is the fixed effect of the Fodder specie j (j = 1–2), Datek is the fixed effect of date of measurement k (k = 1–8), (Site × Specie)ij is the fixed interaction effect of the experimental site i with the fodder specie j, (Site × Date)ik is the fixed interaction effect of the experimental site i with the date of measurement k, (Specie × Date)jk is the fixed interaction effect of the fodder specie j with the date of measurement k and εijk represents the random error.
In a second step, considering the highly significant effects of the Fodder specie (p < 0.0001), we further continued by analyzing the results with a comparative approach between the two species for each parameter; for this we then used the following model:
where Yijk is the plant measurement at date j at the site i, µ is the overall mean, Sitei is the fixed effect of the specific experimental site i (i = 1–3), Datej is the fixed effect of date of measurement j (j = 1–8), (Site × Date)ij is the fixed interaction effect of the experimental site i with the date of measurement j and εijk represents the random error.
After performing the multivariate analysis of variance, the outputs were interpreted taking into consideration the particularities of each experimental location regarding soil properties, climatic parameters and the occurrence of other events.
Results
Plant growth performance
The statistical analysis showed significance levels of the main fixed effects considered for this study (i.e. experimental site, month of the year and their first order interactions) on HEI, DIA, BrN and BrL progression of leucaena (L. leucocephala) and mulberry (M. alba) plants (Table 2). A highly significant interaction (p < 0.0001) between the experimental site and month was observed for all variables except for BrN (p values = 0.90 and 0.78 for mulberry and leucaena, respectively).
Averages reached HEI were similar between fodder species after 7-months, the overall establishment period considered in this study (103 ± 1.7 and 103 ± 2.1 cm for mulberry and leucaena, respectively). However, overall DIA (7 ± 0.4 cm) and BrL (55 ± 2.2 cm) were wider in mulberry than in leucaena (6 ± 0.1 cm and 29 ± 1.0 cm, respectively). In contrast, the number of branches remained higher in leucaena (7 ± 0.2) when compared to mulberry (2 ± 0.1 cm).
In both woody plant species, no differences were observed in HEI between experimental sites from the planting date (June) until the end of August (Fig. 5). However, from the beginning of September, the mulberry planted at PB1 were taller than plants at PB2 and PB3 (Fig. 5; a left panel). In the case of leucaena, a similar non-significant pattern was observed, however, differences between PB1 and PB2 only were detected in November, when plants at PB1 had highest HEI values (Fig. 5; a right panel).

Plant growth progression, as illustrated by the agronomic parameters plant height (a), main stem diameter (b) and branch length (c) in Morus alba and Leucaena leucocephala mixed high-density protein banks, established under three contrasting edaphoclimatic conditions of Burkina Faso (Western Africa). PB1, PB2, PB3 = protein banks or experimental sites 1, 2 and 3, respectively. Asterisks or superscripts indicate significant differences between sites (p < 0.05)
With regard to the DIA, the highest values were also observed in PB1, from October in mulberry and from September in leucaena (Fig. 5; b left and right panels, respectively). No differences between PB2 and PB3 were found in mulberry but, in leucaena plants, the PB2 stems were consistently wider compared to PB3 (Fig. 5; b right panel).
Average BrL was consistently higher (p < 0.0001) in PB1 mulberry plants from the second measurement date in August, whereas no differences in BrL were observed between plants grown at PB2 and PB3 (Fig. 5; c left panel). In the case of leucaena plants, BrL differed (p < 0.0001) among sites from September and until the end of the experiment (Fig. 5; c right panel).
The BrN was affected by the experimental site and, as expected, by the month of the year, for both fodder species (Fig. 6a, b). In the case of mulberry, significantly (p < 0.0001) greater BrN were observed in PB2, which was followed by PB1 and PB3, respectively (Fig. 6; a left panel). No differences were observed between PB1 and PB2 for this parameter in leucaena plants, but PB3 showed lower values (Fig. 6; a right panel). The BrN decreased in mulberry from October to November, then stabilized thereafter (Fig. 6; b left panel). However, leucaena plants increased their number of branches linearly until November, and then stabilized from that point until January (Fig. 6; b right panel).

Effects of the experimental site (a top graphics) and month of the year (b bottom graphics) on the average number of branches (BrN) in Morus alba and Leucaena leucocephala mixed high-density protein banks. Values represent the average BrN (i.e., average of all sites) over time. PB1, PB2, PB3 = protein banks or experimental sites 1, 2 and 3, respectively
Forage yield at the first year of exploitation in PB2 and PB3
The forage yields (Fig. 7) of PB2 and PB3 were calculated at three consecutive dates, every 2 months (i.e. July, September and November). As explained, an accidental fire occurred in PB1 that impeded the ability to carry out the measurements in this site, which ironically had the best growth parameters. Average forage yield per harvest was greater (p < 0.001) at PB2 than at PB3 for both fodder species (1.1 vs. 0.8, and 0.3 vs. 0.1 t DM/ha in leucaena and mulberry, respectively). It was also systematically higher in leucaena as compared to mulberry (0.91 vs. 0.20 t DM/ha, respectively).

Forage yield progression of Morus alba (top) and Leucaena leucocephala (bottom) in the three consecutive first harvests (from July to November) in protein banks (PB) 2 and 3
For both fodder species and at both experimental sites, forage yield reached a peak in September (5.7 and 3.7 in leucaena; 2.0 and 0.3 t DM/ha in mulberry, for PB2 and PB3, respectively). Forage yields were lower in November, averaging 1.2 and 0.45 t DM/ha for leucaena and mulberry, respectively.
Discussion
Feasibility of high density protein banks in Burkina Faso
In West Africa, intensive cropping of trees and shrubs for forage production is rarely documented. Previous work reported by ICRAF (Samanko, Mali) had been carried out under controlled conditions at experimental stations, using low plant densities (Bayala et al. 2014). The high-density forage banks presented in this study are, to the authors’ knowledge, the first to be attempted in this region, with results confirming the hypothesis that adapting the technology to such edaphoclimatic conditions would be feasible. This statement is supported by the observed good growth rates, development of morphological parameters, high plant survival and overall adaptation to local conditions, as well as forage yields with a low input strategy.
In order to evaluate the feasibility, a similar sequence of agronomic activities and labour schedules to those reported by González-García and Martín-Martín (2016) and others in Cuba (Noda et al. 2007; Pentón et al. 2007) or Latin-America (Benavides 1995) was followed and adopted on each experimental site. This study was unique in using seeds for plant propagation, whereas most of the other studies used vegetative propagules for their plantations (Pentón et al. 2007; Noda et al. 2007; Datta 2000). Using seeds was expected to encourage more vigorous plants (Marcos Filho 2015). Seedling trees may have benefited by the use of animal traction during land preparation, which both induced less deep soil preparation but also caused less compaction when compared to using tractors (González-García and Martín-Martín 2016). The dairy cattle manure was applied dried at a rate of 2.5 t/ha. In India, Datta (2000) reported fertilization rates in the range of 10–20 t/ha for different varieties of mulberry. Our values are lower than those reported by Datta (2000), but higher compared to the hen manure used at different doses (100, 300 and 500 kg N/ha/year) by González-García and Martín-Martín (2016) in Cuba.
Effects of different edaphoclimatic conditions on plant growth and biomass production performances
Mulberry and leucaena are known for intrinsic genetic and physiological characteristics of adaptability in tropical conditions, but their agronomic responses from one site to another depend on soil quality, rainfall, temperature, relative humidity and the cultural practices applied (Bertrand 2009; Chotchutima et al. 2016). Although trees (within species) were uniform at establishment, site differences became apparent over time.
Mulberry HEIs obtained at 7 months were 175, 127 and 128 cm in PB1, PB2 and PB3, respectively. These values are substantially higher than those obtained by Noda and Martín (2008) in Cuba. In contrast, the BrNs obtained in this experiment (1.9, 2.4 and 1.7 in PB1, PB2 and PB3, respectively) were lower than the 4.7 reported by these authors in a high-density protein bank of 25,000 plants/ha. The average HEI (154 cm) at 120 days reported by Pentón et al. (2007) with the variety Indonesia is similar to the value found for PB1 in this study (156 cm) at the same age, using the variety Yu-62. Overall, the results with mulberry are close to those published by several authors in Latin America, regardless of variety used. However, the differences in the material used for planting (seeds vs. young mulberry stakes) may create bias in the comparison.
The average DIAs were 9.3, 8.1 and 8.3 mm at 7 months, whereas the length of branches was 127, 63 and 56 cm for PB1, PB2 and PB3, respectively. The results for the monitored morphagronomic parameters are close to those found by Pentón et al. (2007) who highlighted the performance displayed by some mulberry varieties, Although this confirms the adaptability of mulberry to the conditions of Burkina Faso, Martín-Martín et al. (2016) reported some unsatisfactory results when using Tigreada and Yu-62 mulberry seeds in similar field conditions in Cuba.
For leucaena plots, the average HEIs obtained at the end of the experiment were 165, 160 and 109 cm in PB1, PB2 and PB3, respectively. Leucaena BrN (7.5, 7.4 and 6.0 mm), average DIA (9.3, 7.9 and 6.0 mm) and BrL (54.6, 47.5 and 36.5 cm) were also affected by the study site. In a clay-sandy soil with a pH of 5.2, Chotchutima et al. (2016) reported HEI of 79 cm and 231 cm at 4 and 12 months of establishment of leucaena, respectively. These HEI are similar to values obtained at PB3 at 4 months and lower than those obtained in PB1 (104 cm) and PB2 (92 cm).
The physical–chemical properties of the soil were variable from one site to another in the present study (Table 1). The PB1 contained the highest content of available P (7.3 mg/kg−1 soil) followed by PB2 (6.7 mg/kg) and PB3 (3.8 mg/kg). Such differences in soil between the experimental sites could explain in part the differences in performance observed between the three protein banks. Ouédraogo et al. (2015) argued that, in the same way as N and K, P plays a key role in plant growth and development. However, P deficiency is a characteristic element of West African soils (Lompo et al. 2009). This mineral, essential for plant nutrition, is absorbed by the roots in the soil solution, and pH plays an important role in its solubility (Bernoux et al 2011). According to Datta (2000), the optimal pH required for mulberry is between 6.5 and 6.8, but between 7 and 8.5 for leucaena (Bertrand 2009). This, in part, could explain the observed gap in plant growth parameters at PB3 (pH 5.4) compared with the other two sites (Table 1). Comparison of soil textures between the three sites revealed a predominance of sandy particles with more than 80% in PB1 and PB2 (Table 1). The site PB3 had a much more balanced texture. According to Datta (2000) mulberry is adapted to well drained and clayey soils that have good porosity and water retention capacity. The rains, which restarted between April and September at PB1 (918 mm), and between May and October with 718 and 992.5 mm for PB2 and the PB3, respectively (Fig. 1), fall in the theoretically accepted interval of pluviometry reported for mulberry (Datta 2000) and for leucaena (Bertrand 2009).
The attack of termites was a significant negative factor affecting the establishment of protein banks under these conditions in Burkina Faso. This was a particular problem at PB3, with attacks occurring just after planting at the beginning of the establishment period. Termite attacks had less impact during the peak of the rainy season, and most important with the arrival of the cold dry season. Gueye (1987) in forest plantations of Cape Verde made a similar observation. Absence of mulch in PB3 may have increased the magnitude of the termite attacks. While mulch may provide a food resource that supports the development of termite colonies, it may reduce their dependence on new seedlings (Han and Ndiaye 1996).
Trees at PB1 benefited from relatively better management or attention (due to differences in farmers’ attitudes, motivation and initiatives). In particular, PB1 farmer made wider and deeper plantation beds; this was facilitated by greater ease of working the soil (Table 1). Cultural practices such as mulching in early November, followed by irrigation (approximately 1000 L of water, by the farmer initiative) in December, could partially explain the superiority of the performance in the PB1. However, labor availability was also different among farmers or sites. The site PB1 relied on permanently hired field workers, whereas PB2 and PB3 depended mainly on family labor, which became problematic during cropping seasons.
A combination of soil-plant-environment factors, farmer motivation and the related specific management (“man effect”) was likely to have direct effect on biomass production and forage yields, which was proportional to the plants’ growth performance in each protein bank. Unfortunately, after being considered as established, the PB1 suffered an accidental wildfire. This situation impeded to monitor the forage volume production and thus prevent us from making further comparisons of PB1 with PB2 and PB3 during the first 3 harvests.
Biomass of leucaena was higher than that of mulberry in PB2 and PB3. Overall, total accumulated forage yields reached after the first 3 harvests in the first year of exploitation (1.8 t DM/ha and 8.2 t DM/ha, respectively for mulberry and leucaena), are lower than those obtained by González-García and Martín-Martín (2016) in Cuba (10 t DM/ha/year for mulberry) and those of González-García et al. (2009) in Cuba (11 to 15 t DM/ha/year for leucaena). Brewbaker et al. (1985) reported that in subtropical conditions where the growth rate is limited, the yield of edible forage of leucaena could range from 1.5 to 10 t DM/ha/year. Our results are in the top side of that range.
The highest production of leucaena and mulberry in PB2 could also be explained by the less termites attack, resulting in less mortality unlike the occurrence in PB3 plants especially on mulberry trees (Fig. 8).

Schematic representation showing the progression of termite attacks per experimental site and monitoring dates. At the top of the figure, damages provoked on mulberry (B1) and leucaena (B2) are illustrated with regard to healthy plants (A1 and A2, respectively)
Conclusions
Our results demonstrated for the first time the feasibility of achieving well-established high-density mixed protein banks for livestock feeding under the harsh conditions of the sub-humid savannas of West Africa. Both ligneous fodder species evaluated here (leucaena and mulberry) displayed good adaptation to the local environment, which was characterized by poor soils, and fluctuating rainfall pattern and climatic conditions. Plant growth performances obtained in this study are comparable to those reported for the same technology in previous Latin-American experiences. However, the forage biomass yields were lower compared to those reported. Therefore, to achieve good protein-bank establishment and forage production levels, the farmer must pay special attention to key factors, such as the right choice of the site and planting dates, adequate soil preparation, the application of appropriate fertility inputs, and management for, weed, pest and disease controls, a key factor during plantation establishment. Under the conditions of this experiment, plant growth and production performances of both species were affected by the study site, reflecting differences in soil properties (pH, available P and texture) rainfall distribution and management inputs. Control of termite attacks was crucial, and adopting strategic mulching and biological control would help to minimize the negative effects provoked during the establishment period.
Perspectives
In a long-term perspective, pursuing this monitoring work would allow obtaining new relevant insights in terms of forage quality, animal acceptability and productive response, as well as capacity to adapt to dynamic, regular and well-defined harvest frequency regimes during further consecutive productive cycles. It would also permit checks on the protein bank responses when facing climatic alias such as long and unexpected dry periods. The holistic interpretation will contribute to defining alternative strategies for the optimal use of the high quality volumes of forage possible to be obtained with this technology to support local livestock feeding systems with an agroecological and sustainable approach.
References
Bationo BA, Kalinganire A, Bayala J (2012) Potentialités des ligneux dans la pratique de l’agriculture de conservation dans les zones arides et semi-arides de l’Afrique de l’Ouest: Aperçu de quelques systèmes candidats.ICRAF. Technical Manual, No 17. World Agroforestry Centre, Nairobi
Bayala J, Ky-Dembele C, Kalinganire A, Olivier A, Nantoumé H (2014) A review of pasture and fodder production and productivity for small ruminants in the Sahel. ICRAF occasional paper no. 21. World Agroforestry Centre, Nairobi
Benavides I (1995) Manejo y utilización de la morera (Morus alba) como forraje. Agrofor Am 2(7):27–30
Bernoux M, Chenu C, Blanchart E, Eglin A, Bispo T, Bardy M, King D (2011) Le programme GESSOL 2: impact des pratiques agricoles sur les matières organiques et les fonctions des sols. Etude et Gestion des Sols 18(3):137–145
Bertrand G (2009) Caractérisation des réponses adaptatives a la contrainte hydrique dans le Sud-Est de l’Amazonie chez trois: espèces fourragères cultivées en monoculture et en association: Brachiaria brizantha, Leucaena leucocephala et Arachis pinto. Thèse de doctorat, Université Paris-Est. https://tel.archives-ouvertes.fr/tel-00504177
Brandt M, Hiernaux P, Kjeld R, Cheikh M, Laurent K, Torbern T, Yahaya ZI, Abdoulaye W, Compton JT, Rasmus F (2016) Assessing woody vegetation trends in Sahelian drylands using MODIS based seasonal metrics. Remote Sens Environ 183:215–225
Bray RH, Kurtz LT (1945) Determination of total, organic, and available forms of phosphorus in soils. Soil Sci 59:39–45
Brewbaker JL, Hegde N, Hutton EM, Jones RJ, Lowry JB, Moog F, van den Beldt R (1985) Leucaena forage production and use. Hawai, NFTA, p 39
Chotchutima S, Tudsri S, Kangvansaichol K, Sripichitt P (2016) Effects of sulfur and phosphorus application on the growth, biomass yield and fuel properties of leucaena (Leucaena leucocephala (Lam.) de Wit.) as bioenergy crop on sandy infertile soil. Agric Nat Res 50:54–59
Datta RK (2000) Mulberry cultivation and utilization in India. FAO Electronic conference on mulberry for animal production (Morus L.). http://www.fao.org/DOCREP/005/X9895E/x9895e04.htm#TopOfPage. Accessed 20 Sept 2017
Filho JM (2015) Fisiologia de Sementes de Plantas Cultivadas. Londrina, PR: Associação Brasileira de Tecnologia de Sementes - ABRATES, ISBN: 978-85-64895-03-4
Franzel S, Carsan S, Lukuyu B, Sinja J, Wambugu C (2014) Fodder trees for improving livestock productivity and smallholder livelihoods in Africa. Curr Opin Environ Sustain 6:98–103
González-García E, Martín-Martín G (2016) Biomass yield and nutrient content of a tropical mulberry forage bank: effects of season, harvest frequency and fertilization rate. Grass Forage Sci 72:248–260
González-García E, Cáceres O, Archimede H, Santana H (2009) Nutritive value of edible forage from two Leucaena leucocephala cultivars with different growth habit and morphology. Agrofor Syst 77:131–141. https://doi.org/10.1007/s10457-008-9188-4
Gueye N (1987) Rôle des termites dans des plantations forestières du Cap-vert (mbao, Sénégal). Thèse de doctorat Université Paris 6. http://horizon.documentation.ird.fr/exl-doc/pleins_textes/divers09-12/010045991.pdf. Accessed 12 Sept 2017
Han SH, Ndiaye AB (1996) Dégâts causés par les termites (Isoptera) sur les arbres fruitiers dans la région de Dakar (Sénégal). Actes Coll Insectes Soc 10:111–117
Herrmann SM, Tappan GG (2013) Vegetation impoverishment despite greening: a case study from central Senegal. J Arid Environ 90:55–66
Khan NA, Habib G (2012) Assessment of Grewia oppositifolia leaves as crude protein supplement to low-quality forage diets of sheep. Trop Anim Health Prod 44:1375–1381
Kusserow H (2017) Desertification, resilience, and re-greening in the African Sahel—a matter of the observation period? Earth Syst Dyn 8:1141–1170
Lompo F, Segda Z, Gnankambary Z, Ouandaogo N (2009) Influence des phosphates naturels sur la qualité et la biodégradation d’un compost de paille de maïs. Tropicultura 27(2):105–109
Martín-Martín GJ, Noda LY, Olivera CY, Pentón FG (2016) Uso de estimuladores en la supervivencia y desarrollo foliar de propágulos de Morus alba. Use of stimulators in the survival and leaf development of Morus alba propagules. Pastos y Forrajes 39(1):14–18
Mullen BF, Shelton HM, Gutteridge RC, Basford KE (2003) Agronomic evaluation of Leucaena. Part 1. Adaptation to environmental challenges in multi environment trials. Agrofor Syst 58:77–92
Murgueitio E, Barahona R, Chará JD, Flores MX, Mauricio RM, Molina JJ (2015) The intensive silvopastoral systems in Latin America sustainable alternative to face climatic change in animal husbandry. Cub J Agric Sci 49(4):541–554
Noda Y, Martín G (2008) Efecto de la densidad de siembra en el establecimiento de morera para su inclusión en sistemas ganaderos. Zootec Trop 26(3):339–341
Noda Y, Martín G, Mesa A (2007) Nota técnica: influencia de la densidad de siembra en el establecimiento de la morera. Pastos y Forrajes 30(4):449–454
Ouédraogo J, Nacro HB, Ouédraogo E, Youl S, Sedogo MP (2015) Amélioration de la disponibilité du phosphore par la gestion de la macrofaune du sol: cas d’un lixisol en zone semi-aride du Burkina Faso. Int J Biol Chem Sci 8(4):1838–1846
Paré S, Savadogo P, Tigabu M, Ouadba JM, Odén PC (2010) Consumptive values and local perception of dry forest decline in Burkina Faso, West Africa. Environ Dev Sustain 12:277–295
Pentón G, Martín G, Pérez A, Noda Y (2007) Comportamiento morfoagronómico de variedades de morera (Morus alba L.) durante el establecimiento. Pastos y Forrajes 30(3):315–325
Saddul D, Jelan ZA, Liang JB, Halim RA (2004) The potential of mulberry (Morus alba) as a fodder crop: the effect of plant maturity on yield, persistence and nutrient composition of plant fractions. Asian Aust J Anim Sci 17(12):1657–1662
SAS (2002–2003) Statistical Analysis Systems institute (SAS) language guide for personal computers. SAS Institute Inc., Cary, NC, USA, Version 9.3
Vang Rasmussen L, Reenberg A (2012) Collapse and recovery in Sahelian agro-pastoral systems: rethinking trajectories of change. Ecol Soc 17(1):14
Acknowledgements
Authors appreciate the financial grant STRADIV (System approach for the TRAnsition to bio-DIVersified agroecosystems) from Agropolis Foundation (France) (Grant No. 1405-018). They also acknowledge their colleagues from the DP ASAP, the Research Station Indio Hatuey (Matanzas, Cuba) and CIRDES of Bobo-Dioulasso, Burkina Faso for their contribution.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Sib, O., Vall, E., Bougouma-Yaméogo, V.M.C. et al. Establishing high-density protein banks for livestock in Burkina Faso (West Africa): agronomic performance under contrasting edaphoclimatic conditions. Agroforest Syst 94, 319–333 (2020). https://doi.org/10.1007/s10457-019-00394-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10457-019-00394-4