
Abstract
Fossil bioerosion traces at a new site of Les Bez (Jura Department) and those already known from Verrières-de-Joux (Doubs Department) in the central French Jura record an ancient Miocene cliffed shoreline cut into Lower Cretaceous limestones. Both localities are at about 900 m above the present sea level. Caulostrepsis, Gastrochaenolites and Circolites are the ichnogenera identified at Les Bez, whereas only Gastrochaenolites is present at Verrières-de-Joux. This ichnoassociation identifies the Entobia ichnofacies, which corresponds to the upper infralittoral marine environment, biologically associated with the biocenosis of photophilic algae. By means of digital analysis of high-resolution field images, the percentage of the surface affected by the erosive activity of polychaete worms, bivalves and echinoids has been calculated. This information permits a better evaluation of their biological activity, thus complementing the data on the number of specimens per surface area. The cliff preserved at Les Bez was more affected, showing Gastrochaenolites with diameters smaller than those at Verrières. The Circolites–Caulostrepsis association is well developed at Les Bez, where polychaete borings are located within echinoid bowl-shaped pits with particular patterns, suggesting an ecological relationship between involved organisms. Moreover, the distribution of the borings along this outcrop is not uniform and digital analysis at three sections (A, B and C) suggests a lower and an upper zone developed on the outcrop under slightly different environmental conditions within the upper infralittoral. The exceptional preservation of the Les Bez fossil cliff-site demands protection in order to ensure its future existence and accessibility.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The current marine rocky coastline occupies a very restricted area on the surface of our planet and represents a particular environment inhabited by a characteristic biota adapted to harsh living conditions. Rocky shores are subject to high-energy levels, with active wave erosion that produces a changing landscape. Such conditions prevent an extensive fossil record of these locations, limiting it to local fossils of resistant encrusting skeletons (barnacles, oysters, calcareous algae, serpulids) and ichnofossils. Bioerosion traces generated by several groups of invertebrates on rocky substrates represent an excellent indicator of both ancient marine rocky-shorelines and the position of the coast throughout the geological past, as pioneering papers by Johnson (1988a, b) and Johnson and Baarli (1999) have already proved. Furthermore, their positioning furnishes the best opportunities to develop palaeoecological, palaeoenvironmental and palaeogeographic interpretations, in addition to tectonic reconstructions.
Gibert et al. (2012) and Johnson and Baarli (2012) provided exhaustive reviews of the published literature on this topic. According to these compilations and to newer contributions (Baarli et al. 2013; Aguirre et al. 2014, 2017; Domènech et al. 2014; Pineda-Salgado et al. 2015; Rodríguez-Tovar et al. 2015; Santos et al. 2015, among others), Neogene and Quaternary coastal rocky environments are by far the most abundant and best documented. In particular, there is a wealth of studies dealing with ichnoassemblages in European Neogene rocky shores and associated hard substrates (Radwański 1970; Bałuk and Radwański 1977; Gutowski and Machalski 1984; Martinell and Domènech 1986; Silva et al. 1995; Aguirre and Jiménez 1997; Gibert et al. 1998,2012; da Silva et al. 1999; Domènech et al. 2001; Uchman et al. 2002; Titschack et al. 2005; Radwański et al. 2006,2011; Cachão et al. 2009; Santos et al. 2008, 2010, 2011a, b, 2016; Johnson et al. 2011; Demircan 2012; Brlek et al. 2016; Aguirre et al. 2017; Rosso et al. 2015; Steinthorsdottir and Håkansson 2017, among others).
In the central Jura Massif (eastern France), the ancient Miocene shoreline has been identified in cliffs cut into Mesozoic limestones and in their associated sediments from fossil bioerosion traces. Specifically, the best localities are situated in the Jura département, with one new outcrop, and in the Doubs département, where documented sites (Rangheard et al. 1985; Pharisat and Rangheard 1986; Martin et al. 1991) have been examined. Apart from these references, there is little detailed information on the ancient bored rocky shores in the French Jura. Only Charollais et al. (2006) mentioned the presence of bioperforations in the Urgonian limestones of the Combe d'Evuaz-la Pesse basin. However, various outcrops of the Jurassic basement with borings of a Miocene age have been known for a long time in the German Jura. In particular, Reiff (1989) and Hoffmann and Friedrich (2017) dealt with several sites in the Swabian Alb, the northeastern geological extension of the French Jura, together with the Franconian Alb.
The main objectives of the present research are as follows: (a) to provide new ichnological localities that refine the position of the Burdigalian coast in eastern France; (b) to calibrate the extent of bioerosion through image analysis; (c) to characterise ichnologically and environmentally the preserved ichnoassemblages; (d) to study the ecological relationships between producers by means of their traces and (e) to highlight the heritage value of the outcrops described here.
Geographical and geological settings
General overview
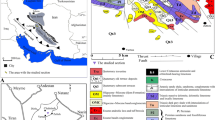
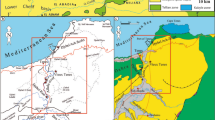
The study sites are located on the French side of the Jura Massif, a crescent-like fold and thrust belt (Fig. 1). The Jura constitutes a foreland chain located in front of the western part of the Alpine arch and is surrounded by various Cenozoic basins (the Upper Rhine Graben plain, the Bresse Graben and the Swiss Molasse Basin). The Jura Mountains are constructed of Mesozoic limestones and marls which were folded by the Alpine orogeny in the Late Miocene (Homberg et al. 2002; Laubscher 2010). The Jura are divided into two main domains characterised by their different altitudes and geological structures. The Western domain (so-called Jura des plateaux) constitutes the external Jura, with altitudes ranging from 400 to 700 m. Structurally, the Jura des plateaux consists of tabular zones (plateaux) separated by fault and fold structures named faisceaux by the locals. The Eastern zone or inner Jura (so-called Haut Jura) presents an alternation of well-developed anticlines and synclines ranging from 800 to 1700 m in height. On a large scale, deformation is structurally characterized by folds, thrusts and strike-slip faults.

Geographic and geological location of the studied sites
The Jura Massif is flanked to the SE by the flexural Swiss Molasse foreland basin of the Alps (Ziegler and Fraefel 2009). The remnant part of this Alpine basin can be found in the core of synclines of the Haut Jura and it is known as Jura Molasse (Rangheard et al. 1985; Homewood et al. 1989). Nevertheless, the infilling molasse crops out very rarely in the synclines (Weidmann 2008) due to the deposition of glacial tills during the last glacial maximum (Buoncristiani and Campy 2004).
Localities with bioerosion features
Three outcrops corresponding to ancient shoreline cliffs at two different sites (Les Bez, Commune de la Grande-Rivière Château -Jura département, and Les Verrières-de-Joux-Doubs département) are documented here (Fig. 1). Domènech et al. (2014) named and described them Les Bez site, but a more in-depth analysis was pending. In the meantime, the site has been registered as Geotope N°39–59 in the Geological Heritage of the Franche-Comté Région (Bichet et al. 2014). The neighbouring outcrop of l'Abbaye en Grandvaux (Rangheard et al. 1985) is included as the same site in the present study. The Les Verrières-de-Joux area, already documented in the literature by Pharisat and Rangheard (1986), Rangheard et al. (1990) and Martin et al. (1991) is now revisited. All of them are described in the following section.
Description of the new and revisited sites
The new outcrop is located near the Les Bez hamlet (46°31′32″ N, 5°53′47″ E) (Grande-Rivière Château, Jura), in a 140 m-long and 2–3 m high section that is visible along the D146 road, at 898.5 masl. (Fig. 2). The basement consists of limestones corresponding to the Urgonian facies of Barremian age (Lower Cretaceous) (Guillaume and Guillaume 1968). The strata show a vertical to overturned dip due to their location in the Grand-Rivière Syncline. As in other areas of the Jura Massif, the core of the SW–NE oriented syncline contains Burdigalian marine molasse (Pierçon 1932; Rangheard et al. 1990; Charollais et al. 2006). As a result of differential erosion, the contact between the marine molasse and the limestones of the pre-Cenozoic Urgonian facies is morphologically well expressed in the topography, although outcrops are rare due to vegetation cover. The molasse conglomerates, composed of limestone and flint clasts in a micaceous sandstone, are in contact with the Barremian substratum (Pierçon 1932) but it is uncertain if they infill morphological pits of the palaeorelief or if, alternatively, secondary faults are responsible for the contact. Rangheard et al. (1990) noted that in the Les Bez site the base of the outcropping molasse corresponds to a puddingstone with Crassostrea gryphoides (Schlotheim 1820).

General view of the Les Bez site (Grande-Rivière Châteaux, Jura)
Near the former Abbaye en Grandvaux, located some 1.5 km NE from the main outcrop, the molasse appears beside the road D437 (Fig. 3). It occupies a small location and Rangheard et al. (1990) also described the materials as a puddingstone with Crassostrea gryphoides.

General view of Abbaye en Grandvaux site (Grande-Rivière Châteaux, Jura)
The second outcrop is located near Les Verrières-de-Joux, specifically on the N slope of the road D67B (46°53′13″ N, 6°25′52″ E), at 900 masl. (Fig. 4). Aubert (1972), Rangheard et al. (1985), Pharisat and Rangheard (1986) and Martin et al. (1991) identified two sites with bioerosion traces (borings of lithophagid molluscs and cupules of sea-urchins) in the Hauterivian limestones that constitute the pre-Cenozoic basement within the Verrières syncline at this locality. Strata are oriented N60°E and show a 40–60°SE dip (Rangheard et al. 1985). The section of the so-called Chemin du Larmont is no longer available and, at present, only one site with evidence of bioerosion has been located in the area, equivalent to that described by Rangheard et al. (1985) some 2 km to the SW of Les Verrières. It is exposed over 40 m along the road, with some 3 m exposure height. Following Rangheard et al. (1985), the Miocene succession is composed of conglomerates, sandstones, coquinas, limestones, shales and marls, with a quite abundant although poorly preserved fossil content (calcareous nanoplankton, rhodophyceans, foraminiferans, oysters, pectinids, bryozoans). These materials uncomformably overlap the Cretaceous limestones dipping 20°SE (Rangheard et al. 1985).

General view of the Les Verrières-de-Joux site (Doubs)
Material and methods
The main object of this study was to document the assemblage of recognizable boring traces on the rocky slopes that constitute the Les Bez and Les Verrières-de-Joux outcrops. Two approaches have been undertaken: (a) characterisation, physical distribution analysis and calibration of the trace density (number of borings/m2 and percentage of the bored surface) along the outcrop by means of digital image treatment of the ichnofossil assemblages, and (b) study of the (palaeo)ecological relationships among the corresponding trace-makers.
Specially related to the first approach, we consider that erosion other than that produced by marine organisms in the Miocene (waves, storms, but also recent subaerial weathering effects of rain, roots, etc.) may have modified the trace morphology, the penetration into the rock and their interconnections, which determine their current appearance. Consequently, quantification of the affected surface has been undertaken, and in view of that, only some selected locations have been taken into account in this research to illustrate the bioeroders’ impact on the palaeocliffs.
Digital treatment of trace fossil imaging and pixel-counting methodology are commonly used for analysis of bioturbation (Dorador and Rodríguez-Tovar 2014, 2018; Rodriguez-Tovar and Dorador 2015; Razzolini et al. 2016, 2017; Rodríguez-Tovar et al. 2018; Marty et al. 2018; Razzolini and Klein 2018, among others) and bioerosion in mobile hard substrates [shells, guards, thecas; Wisshak (2010), Gibert et al. (2012), and references therein]. Although the potential utility of this approach is promising, there are few examples of its application to outcrops. Ancient bioeroded cliffs and rocky shores provide an ideal subject to apply these techniques, although effective sampling is challenging because the research subject is the substrate itself, namely holes and pits upon it.
Cachão et al. (2011), for example, applied photogrammetric methods to study a Portuguese Miocene rocky shore exposure with a high density of Gastrochaenolites (borings produced by endolithic bivalves). The obtained images allowed the authors to analyse aspects such as density and distribution of borings and trace alignments, to discriminate between ichnospecies and to draw palaeoenvironmental deductions.
We applied a simple and practical method to evaluate the bioeroded surface at the two locations. Once the traces were identified, the visible specimens of each ichnotaxon on selected surfaces were counted and the percentage of affected surface areas was calculated. Counting and measurements were obtained by combining fieldwork with the high-resolution digital images taken in the field.
The first step consisted in obtaining images along the outcrops. This was achieved using a Nikon Coolpix camera. All the outcrop was imaged at a distance of 3 m with images of 300 ppi. Several close-up pictures completed the set, aimed to provide detailed counts inline with a palaeoecological approach. Second, selected pictures were digitally manipulated with Adobe Photoshop CS6 to a grey-scale format and bored areas were manually delimited and converted to black. With this technique dark pixels due to shadows or foreign elements are avoided. Finally, the program UNESCO Bilko 3.3 allowed us to quantify the black surface percentage with regard to the total considered surface and subsequently the boring intensity.
Results
Ichnofossil assemblages
Although rich in specimens, a very low ichnodiversity was observed for both the Les Bez and the Verrières-de-Joux sites. On the whole, bioerosion is represented by the ichnogenera Entobia,Caulostrepsis, Gastrochaenolites and Circolites, produced, respectively, by clionaid sponges, polychaete annelids, endolithic bivalves and regular echinoids. Trypanites-like structures are locally present, although they seem attributable rather to taphonomically altered Caulostrepsis. Entobia is the least represented ichnogenus in the outcrops and, moreover, the few sparsely observed specimens are poorly preserved, thus impeding their ichnospecific identification. Caulostrepsis and Gastrochaenolites correspond to penetrative traces, a condition that makes assigning all the specimens to a particular ichnospecies difficult because of its particular substrate. However, a calcareous hand-sample was cut that permitted the identification of at least the presence of Caulostrepsis cretacea (Voigt 1971), Gastrochaenolites lapidicus Kelly and Bromley, 1984 and G. torpedo Kelly and Bromley, 1984. Finally, Circolites bowl-shaped pits were assigned to C. kotoucensis Mikuláš, 1992.
The most extensive exposure occurs at the Les Bez site, which is 140 m-long and had 2–3 m of variable height along the road trench (Fig. 5). Three different N to S sectors can be differentiated at the Les Bez outcrop on a bioerosion basis: sector A—Circolites-Caulostrepsis sector, with very abundant specimens of both ichnogenera (Fig. 5a); sector B—Circolites-Caulostrepsis-Gastrochaenolites sector, where all three ichnotaxa are well represented but with small concentrations (Fig. 5b), and sector C—characterized by a great abundance of Gastrochaenolites and locally Entobia (Fig. 5c). No shell remains are found inside of the borings across the entire outcrop.

Les Bez section and the three differentiated sectors: a Circolites-Caulostrepsis sector; b Circolites-Caulostrepsis-Gastrochaenolites sector; c Gastrochaenolites sector
In sector C, a zone of 0.127 m2 was delimited and traces were counted (271) and digitised (Fig. 6a, b). Gastrochaenolites ispp. observed in that area show highly dense concentrations, theoretically as high as 2,084 specimens/m2, covering up to 28% of the surface (Table 1).

Area with Gastrochaenolites (sector C) at the Les Bez site: a General view of the documented zone; b Graphical representation of the digitised traces, covering 30% of the surface
Circolites kotoucensis mostly occurs at the southernmost zone of the outcrop (sector A), where it forms smooth hemispherical, bowl-shaped pits, some of them up to 9 cm in diameter (Fig. 7). Ninety-five specimens were counted and registered among sectors A and B in this area. Among these structures, 55% contain Caulostrepsis isp. specimens, with a variable yet significant presence. The distribution of infested C. kotoucensis is not random in the outcrop and two zones are broadly delimited on this basis: a lower zone, corresponding to the first 1/3 of the slope (which encompasses 28 traces, so 30% of the total) and an upper one, that comprises the remaining 2/3 (with 67 traces, or 70% of the total). These zones are not clearly observable along the whole road exposure, as C. kotoucensis is not present in sector C. The analysis of the Circolites-Caulostrepsis association is addressed in the next section.

Circolites kotoucensis in Sector A at the Les Bez site
Caulostrepsis isp. does not only appear inside Circolites specimens, but also affects the Cretaceous limestones in other areas of the outcrop (Fig. 8), although generally showing poor preservation. It becomes locally very abundant, as seen in the case of a small surface of 413.6 cm2 in sector B that contains 186 borings (that is, some theoretical 4500 borings/m2), affecting 20% of the area.

Surface with several Caulostrepsis isp. out of Circolites bowls at the Les Bez site
In the neighbouring outcrop of Abbaye en Grandvaux, the molasse contains several Crassostrea gryphoides shells, and boulders show some Gastrochaeonolites lapidicus and G. torpedo.
The Verrières-de-Joux site is located some 70 km to the NE with respect to Les Bez and it is smaller than that outcrop, with an exposure of some 40 m along the road (Fig. 4). Only the ichnogenus Gastrochaeonolites is identified there, covering a significant area of the exposed surface (Fig. 9a). In line with the Les Bez site, no shell remains are found inside the borings along the outcrop.

Section with Gastrochaenolites at the Verrières-de-Joux outcrop: a General view of the documented zone; b graphical representation of the digitised traces, covering 15% of the surface
For a selected 0.35 m2 surface area, 512 specimens are present (some theoretical 1,600 borings/m2) that affect 15% of the total area (Fig. 9b, Table 1). The contrast in bored surfaces between the Les Bez and Les Verrières-de-Joux sites should be noted. Although the Gastrochaenolites density (number of borings/m2) in the examined surface at Les Bez is 1.4 times that of Verrières-de-Joux (2084 versus 1600), the bored surface rate increases to 1.9 times (30% versus 15%). Despite the presence of Gastrochaenolites with diameters smaller than those of Verrières-de-Joux, the cliff was more precisely affected at Les Bez in terms of its surface.
Rangheard et al. (1985), Pharisart and Rangheard (1986), and Martin et al. (1991) also mentioned the presence of sea-urchin depresions (i.e., Circolites) in different outcrops of the Verrières-de-Joux zone. Unfortunately, these localities are no longer accessible or have disappeared and the only present outcrop in the area does not show Circolites at all.
Circolites–Caulostrepsis association
Circolites kotoucensis is a dwelling trace produced by regular boring echinoids (Martinell 1981; Mikuláš 1992; Santos et al. 2011b, 2012a, b; Belaústegui et al. 2017, among others) and consists of bowl-shaped, hemispherical or deeper pits bored in hard or firm substrates in shallow waters. Trace-makers usually inhabit them permanently and only move for short distance around to search for food. These bioerosive structures are mechanically produced by using jaws and spines in both sedimentary and non-sedimentary hard rocks: beach and carbonate rocks (e.g. Martinell and Domènech 1986; Domènech et al. 2014), granite (e.g. Martinell 1981), metamorphic (Watkins 1990; Santos et al. 2011a), and volcanic rocks (Ramalho et al. 2010, 2013; Santos et al. 2011b, 2012a, b, 2015; Baarli et al. 2013; Carey et al. 2020). In the Mediterranean Sea these structures are mainly produced by Paracentrotus lividus (Lamarck 1816), an herbivorous species that fed on calcareous and non-calcareous algae and seagrass, no deeper than 20 m (Kemp 1962; Pérès and Picard 1964; Martinell 1981). In the Neogene fossil record, examples of sinuous grooves (Ericichnus) excavated by sea-urchins and associated with Circolites also exist (Santos et al. 2015; Aguirre et al. 2017). Ericichnus is interpreted as an agrichnion-pascichnion structure (Santos et al. 2015), and it has not been identified in the studied area. The erosive activity of regular echinoid populations produces significant amounts of sediment in a short-medium term, as demonstrated by Russell et al. (2018) through experimental studies.
In the geological record the ichnogenus Circolites ranges from the Jurassic to the Holocene (Mikuláš 1992; Kemps, 1962; Pérès and Picard 1964; Martinell and Domènech 1995; Gibert et al. 1998, 2007; Bromley 2004; Dermican 2012; Doyle et al. 1998; Santos et al. 2008; Santos and Mayoral 2009; Johnson et al. 2011; Baarli et al. 2013; Belaústegui et al. 2017).
Caulostrepsis corresponds to narrow penetrative U-shaped galleries attributed to the boring activity of polychaete annelids (Bromley and D’Alessandro 1983; Domènech et al. 2008). The apertures of the traces appear contiguous and are oval or eight-shaped, whereas their inner galleries vary in morphology from rectilinear to contorted tubes, which in some cases show vanes connecting the inner sides of their limbs (Fig. 10a, c, d). Epoxy casts produced in the laboratory (Fig. 10b, d, f) demonstrate that the boring penetration in the dolomite at Les Bez is between 6 and 8.5 mm and the trace morphology coincides with those described by Boekshoten (1966a, b), Voigt (1971), Bromley and D’Alessandro (1983) and Hanken et al. (2012).

Caulostrepsis isp., Les Bez: a, c, e Details of three limestone samples with the ‘eight-shaped’ openings of the trace. b, d, f Their corresponding epoxy casts showing the typical U-shaped morphology of the polychaete borings
As mentioned above, 55% of Circolites kotoucensis specimens in sectors A and B at the Les Bez site host Caulostrepsis isp., affecting up to 11% of the surface in the pit bottom. The presence of the ichnogenus Caulostrepsis in the concave surfaces of Circolites was mentioned for the first time in the fossil record by Johnson et al. (2011). These authors found rare specimens inside C. kotoucensis in the Upper Miocene rocky shores along the flanks of the headland at the north coast of Menorca (Balearic Islands). By contrast, Caulostrepsis isp. is locally very abundant at the Les Bez site. Unlike the Menorca example, here it is especially present inside the echinoid bowls: the inner surface of 70% of C. kotoucensis specimens located at the upper part of the outcrop contains Caulostrepsis isp., with very variable densities. No comprehensive analysis has been carried out due to the difficulty of distinguishing specimens due to the high surface alteration, but illustrative examples can be presented from three random samples (Table 1). Amongst them, the most affected Circolites (nº 2 in Table 1), which covers a surface of 135.6 cm2, contains up to 420 specimens of Caulostrepsis isp. that represent 12% of the bored surface (Table 1). Within those samples, the least affected one (nº 1 in Table 1) contains 40 Caulostrepsis isp. on a surface of 32.1 cm2, affecting 5% of it.
The percentages of C. kotoucensis with Caulostrepsis isp. differ within the outcrop. Sectors A and B, in the upper part of the cliff, concentrate 70% of the Circolites, whereas the remaining 30% are located in the lower part of the outcrop (sector C). At the top, 72% of them show Caulostrepsis isp. inside, whereas this percentage drops to 15% at the bottom. With regard to the top percentage, it should be noted that 30% of the pits concentrate the borings in the outermost band of the cuvette and the centre is clean (Fig. 11a, b), whereas the remaining borings are spread all over the background in a random way (Fig. 11c), just as happens to all the C. kotoucensis on the visible bottom.

Examples of Caulostrepsis distribution inside Circolites kotoucensis: a, b Circolites showing the polychaetes’ proclivity to concentrate in the outermost band of some echinoid bowls. c C. kotoucensis with a random distribution of the polychaete borings
All these distributions suggest that the association between C. kotoucensis and Caulostrepsis isp. in the outcrop is reflecting an ecological relationship between the organisms producing these traces. As mentioned above, Caulostrepsis has been largely attributed to the activity of polychaete annelids (Boekschoten 1966a, b; Bromley and D’Alessandro 1983, 1990; Martin and Britayev 1998; Domènech et al. 2008; Bromley 2004; Hanken et al. 2012). Families Spionidae, Sabellidae, Cirratulidae and Eunicidae are the most reported present boring groups. In the fossil record, Caulostrepsis has often been attributed to the spionid Polydora or the cirratulid Dodecaceria. At present, there is no documented relationship between Polydora and the echinoid cavities, while different authors have highlighted this association with Dodecaceria. Since this section focuses on the Circolites–Calustrepsis relationship, only literature regarding Dodecaceria and echinoids is summarized below.
Kempf (1962) and Schoppe and Werding (1996) described Circolites-like borings generated by the modern echinoid Paracentrotus lividus in Marseille (France), and Echinometra lucunter (Linnaeus, 1758) and E. vanbrunti A. Agassiz, 1863 and on the Caribbean coast of Colombia, respectively; they pointed out the presence of several organisms (including algae, crabs, gastropods, polyplacophorans, clingfish, and/or brittle stars) cohabiting within these boreholes. Kempf (1962) noted the presence of the boring cirratulid polychaete Dodecaceria concharum inside the cavities (cupules), concluding that the boring activity of P. lividus was probably favoured by the action of these worms.
For the first time in the fossil record, Voigt (1970) identified boring activity attributable to Dodecaceria consisting of small borings in Santonian (Late Cretaceous) pebbles from Germany. Voigt (1971) also described “casts of tunnels burrowed by polychaetes into the colonies of scleractinians” in the Maastrichtian (Late Cretaceous) chalk-tuff of the Netherlands and identified the flat “burrows” (in fact, borings) as Dodecaceria cretacea n. isp. Dodecaceria is a genus of cirratulid polychaete that impedes its use in ichnology. In view of this, Voigt’s Dodecaceria cretacea is included in Caulostrepsis cretacea (see Bromley and D’Alessandro 1983).
Gibson (2017) studied the boring activity of the Holocene polychaete Dodecaceria and compared their traces with fossil Caulostrepsis from several Cretaceous and Neogene localities. He concluded that they are different from those produced by Holocene Dodecaceria species. Nevertheless, the author provided several images of their Holocene traces in shales and encrusting calcareous algae that perfectly coincide with the morphology of Caulostrepsis isp. in the Les Bez Miocene outcrop. Finally, Gravina et al. (2019) concluded that Dodecaceria concharum is responsible for Caulostrepsis and Maeandropolydora traces that affect a submerged mosaic Roman floor in the Gulf of Naples (Italy), which is covered by calcareous algae.
Dodecaceria infests encrusting calcareous algae, whereas Paracentrotus, Echinometra and other echinoids feed on them. Therefore, in this case the association polychaete-regular echinoids could be explained by their mutual relationship to such algae. In several cases at Les Bez outcrop, the position of Caulostrepsis is limited to the distal zone of Circolites. That can be interpreted as a result of the cohabitation between the producers, the echinoid offering passive protection to the worm against predators and/or the high-energy environment (Davidson and Grupe 2014).
Palaeoenvironmental interpretation of the ichnoassemblage
Bioerosion provides evidence of a strong relationship between the behaviour of the producers and the resulting traces based on their almost permanent emplacement (Taylor and Wilson 2003; Tapanila 2008). From an ethological point of view, the ichnoassemblage at the Les Bez site consists of domichnion bioerosion traces produced by sessile endolithozoans (annelids and bivalves), as well as semi-endolithozoans (sponges) and vagile epilithozoans (regular echinoids). Although there is a token presence of Entobia, the association Caulostrepsis-Gastrochaenolites-Circolites allows one to attribute the ichnoassemblage to the archetypical Entobia ichnofacies in hard substrates (Bromley and Asgaard 1993a, b; Gibert et al. 1998, 2012). Gastrochaenolites lapiducus and G. torpedo are also present in the small neighbouring outcrop of Abbaye en Grandvaux.
At Les Verrières-de-Joux only Gastrochaenolites, produced by the activity of sessile endolithic bivalves, was recorded on this occasion, but Rangheard et al. (1985), Pharisart and Rangheard (1986) and Martin et al. (1991) also noted the former presence of sea-urchins bowls (i.e., Circolites). Ichnoassociation assimilation to the Entobia ichnofacies is, therefore, also feasible.
The calculated theoretical abundance of Gastrochaenolites per m2 is 2084 at Les Bez and 1600 at Verrières. These amounts are comparable with concentrations of 1250–1500 Gastrochaenolites per m2 calculated for similar Miocene bioeroded surfaces on the marine palaeocliffs of Foz da Fonte (Portugal) (da Silva et al. 1999) and Tarragona (NE Spain) (Gibert et al. 1998). Nevertheless, differences are remarkable in relation to other sites, such as the Miocene outcrop of Oura (Portugal) (Cachão et al. 2009), where the abundance is reduced to 300 specimens per m2, or to the bored surfaces studied in numerous Pliocene basins of Western Mediterranean (Spain: Baix Llobregat and Alt Emporda; France: Vence and Gard), with theoretical densities of 100–400 specimens per m2 (Martinell and Domènech 1995; Clauzon et al. 1995; Gibert et al. 1996, 1998). The higher concentrations in Les Bez and Verrières could indicate a similar environment to that observed in the Miocene Iberian Mediterranean cliffs, which is clearly shallower than that of the Pliocene in the same area, or in the Atlantic Miocene.
The Entobia ichnofacies, which may contain Trypanites (not identified in this study), has been classically interpreted as an indicator of shallow to very shallow-water settings with a low or null sedimentation (Bromley and Asgaard 1993a, b; Martinell and Domènech 1995; Aguirre and Jiménez 1997; Gibert et al. 1998, 2012; Silva et al. 1999; Domènech et al. 2001; Santos et al. 2008, 2011a, b, 2012a , b; Cachão et al. 2009; Johnson et al. 2011; Aguirre et al. 2014, among many others). Although similar ichnofossil assemblages have been described in deep-water macroids collected in 60–100 m water depth (Bassi et al. 2011, 2012), which could confer upon it a greater value as an indicator of frequent turbulence and low sedimentation rates rather than as a water-depth indicator, both numerous quantitative studies and qualitative observations show a decrease in the bioerosion rates as a function of depth, both in the tropics and in high-latitude settings (see summary in Wisshak 2010). Generally, the presence of macroborers increases in shallow waters as well as with exposure time (e.g., Tribollet and Golubic 2005; Wisshak 2010). As an example, polychaetes are important macroborers from 1 to 7 m water depths and so Caulostrepsis isp. occurs in shallow-water environments and significantly decreases below its highest abundance at 15–30 m in cold temperate areas (Wisshak et al. 2005; Wisshak 2010).
Kemp (1962) already described the boring activity of Dodecaceria in Lithophyllum incrustans Phillippi, 1837 crusts in Mediterranean shallow environments. Gibson (2017) described it for Lithothamnion crusts at low spring tides on the coast of Great Britain, and Gravina et al. (2019), for a Roman mosaic submerged at 2.5 m on the Italian coast. Moreover, Dodecaceria disappears below salinities of 34‰ (Gibson 1996).
In addition, bioeroding activity by echinoids is constrained to high-energy very shallow waters worldwide, close to the low-tide line (see a review in Asgaard and Bromley, 2008). In the western Mediterranean Sea, Martinell (1981) described dense concentrations of Paracentrotus lividus within their characteristic cup-shaped traces (Circolites) in rockgrounds of different lithologies (limestone, granite, metamorphic rock), in very shallow waters (0.5–5 m). Therefore, the significant presence of Circolites kotoucensis (with Caulostrepsis inside, in addition) in the Le Bez outcrop permits us to constrain the palaeodepth of the identified ichnoassemblage to very shallow waters (in any case, not deeper than 5 m). Moreover, based on observations by Bassi et al. (2011, 2012), this ichnoassemblage indicates low or null sedimentation rates as well as high water turbulence, inherent to these shallow-water settings.
The presence of Circolites, Caulostrepsis and Gastrochaenolites, with local Entobia, would place sectors B and C of Les Bez in the uppermost infralittoral zone, which would be covered with photophilic algae, such as Lithophyllum and Lithothamion. Comparatively, sector A could be identified with the deepest part of the mesolittoral zone, with a high abundance of Circolites and Caulostrepsis, but a near absence of bivalve borings. On the whole, the palaeoenvironment could be attributed to the classical biocoenosis of photophilic algae described by Pérès and Picard (1964). The sole presence of scarce Gastrochaenolites at the l’Abbaye en Grandvaux site does not permit a precise environmental attribution, although an infra-meso littoral position is indeed compatible with the fossil content. At Verrières-de-Joux, the presence of Gastrochaenolites and Circolites (known from the literature) permits placing the outcrop in the infralittoral zone.
Rocky shores in the Jura mountains (s.l.) and geoheritage considerations
As mentioned in the introduction, there is still little information about ancient bored rocky shores in the French Jura. Charollais et al. (2006) noted the presence of “bioperforations” (Ø 5–8 mm) in the Urgonian limestone that crops out in the southern area (Ain Department) of the Combe d'Evuaz-la Pesse Basin, which is unconformably covered by Miocene sediments. Tentatively, these borings are identified as possible small Gastrochaenolites. Although the authors do not explictly mention the traces’ position above present sea-level, it is inferred from their maps as under 900 m.
Les Bez and Verrières-de-Joux outcrops are also located at about 900 masl. No other localities of bioeroded palaeocliffs are known at similar altitudes above current sea level within the European Miocene of the Tethys domain. The presence of features indicative of littoral environments at this altitude provides useful information contributing towards interpreting the evolution of the coastline itself during the Burdigalian, reconstructing the regional palaeogeography of that time and subsequently, our understanding of the tectonic evolution (uplift, tilting) of the Jura Massif. These aspects are beyond the research presented here, but could be explored in future studies.
In the German Jura, various outcrops of Jurassic basement with borings of Miocene (Burdigalian) age have been known for a long time. Although they belong to the Paratethyan domain, these outcrops are comparable with the studied ones because of the same age for the rocky shores (Burdigalian) and their geological location in the NE branch of the Jura mountains. Reiff (1989) described the Heldenfingen site in the northern part of the Swabian Alb (Baden-Württemberg state). More recently, Hoffmann and Friedrich (2017) studied this site and a neighbouring and similar outcrop at Burgmagerbein. Both localities are part of the so-called Swabian Alb cliff-line, which runs discontinuously for about 100 km from E to W, and are located at 645 and 455 m above present sea level, respectively. Hoffmann and Friedrich (2017) assigned the borings to sponges and lithophagous bivalves and record the spatial pattern and the dimensions of bivalve boreholes by defining classes of borehole diameters ranging from < 0.4 cm to > 2 cm, while counting those holes in defined squares. Moreover, they recorded the positions of holes of different sizes relative to each other. Ichnotaxonomic aspects are not considered in this approach. Hoffmann and Friedrich (2017) distinguished two different bioerosion levels in these outcrops, determining that the marine features of the Heldenfingen site suggest a greater water depth compared to Burgmagerbein site. The Heldenfinger site (known as Heldenfinger Kliff) makes up part of the UNESCO Global Geopark Swabian Alb and was designated as a Geotope in 2019.
Beyond its ichnofossil record, the rocky cliff identified at the Les Bez site provides convincing evidence of the Miocene palaeo-shoreline in the Jura chain, thanks to the bioerosion features documented here. The Les Bez outcrop deserves a high level of protection to avoid any destruction of the rock face and its exceptional fossil traces, resulting from their location at the side of a main road. The section corresponds to the actual surface of the cliff, and any damage would completely eliminate the fragile, yet exceptional, fossil record. At present, this locality is recognized as Geotope nº 39–59 “Littoral burdigalien à Grande-Rivière”. The patrimonial consideration of this geotope in the Franche-Comté Regional Geological Heritage Inventory is 2 stars (on a scale of 0–3), with a heritage value of 29 (on a scale of 0 to 48) and with a necessary protection level of 7 (from 0 to 12) [see Bichet et al. (2014) for an explanation of the scales used].
Conclusions
Following an ichnological study of the bored Miocene rocky surfaces outcropping in the central area of the Jura Massif (eastern France), we draw the following conclusions:
-
(1)
In Les Bez (Jura département) and Verrières-de-Joux (Doubs département) two Miocene marine rocky shores have been identified on the Cretaceous limestones on the basis of the bioerosion traces. The Les Bez one has been characterised for the first time. The second—already known in the geological literature—has been revisited and revised.
-
(2)
The Entobia ichnofacies is found at both sites, with extensive Gastrochaenolites and Circolites (known only from the literature at Verrières), and Caulostrepsis at Les Bez. This ichnofacies corresponds to sdecidedly littoral marine environment (upper infralittoral), which in this area seems to point to a shallower level than in other already described, and indeed southernmost basins, in any case, not deeper than 5 m.
-
(3)
Traces are not evenly distributed along the Les Bez site. Their occurrences at sections A, B and C have been characterised and differentiated on the basis on their ichnological composition. Digital analysis provides data to compare different zones with respect to the percentage of affected surface, allowing to distinguish a lower and an upper zone in the outcrop, which were affected by slightly different palaeoenvironmental conditions.
-
(4)
A Circolites–Caulostrepsis association is well developed at the Les Bez outcrop. Annelid borings are distributed in different patterns inside echinoid bowls: in some cases covering all the surface, and in others located in the most external part. This situation suggests an ecological relationship between the involved organisms.
-
(5)
The calibration of the percentage of surface affected by endolithozoans (boring bivalves) provides a new way to evaluate their biological activity, which complements the data on the number of specimens/surface. In these examples, whereas the Gastrochaenolites density (number of borings/m2) at Les Bez is 1.4 times higher than in Verrières-de-Joux, the same relation to the bored surface increases by 1.9 times. In terms of the surface, the cliff was more affected at Les Bez, even showing Gastrochaenolites with diameters smaller than those at Verrières.
-
(6)
The palaeoenvironment deduced from the identified Entobia ichnofacies and the ecological requirements of their producers suggest their affiliation with the classical biocoenosis of photophilic algae.
-
(7)
The outcrop of Abbaye du Grandvaux, in the vicinity of the Les Bez site, is ichnologically characterised by the presence of rare Gastrochaenolites in cobbles of the bedrock.
-
(8)
The Les Bez and Verrières-de-Joux outcrops are located at about 900 m a.s.l. The presence at this altitude of features indicative of littoral environments provides useful information in order to interpret the evolution of the Burdigalian coastline itself, the regional palaeogeography and the tectonic evolution of the Jura Massif. Finally, further studies should look to combine data from the French and German Miocene rocky shores.
-
(9)
The rarity and the exceptional preservation of the Les Bez site demand a reevaluation of its degree of protection, in order to ensure its future existence.
Data availability
Data available from the authors.
Code availability
Not applicable.
References
Aguirre J, Jiménez AP (1997) Census assemblages in hard-bottom coastal communities: a case study from the Plio-Pleistocene Mediterranean. Palaios 12:598–608
Aguirre J, Belaústegui Z, Domènech R, de Gibert JM, Martinell J (2014) Snapshot of a lower Pliocene Dendropoma reef from Sant Onofre (Baix Ebre Basin, Tarragona, NE Spain). Palaoegeogr Palaeoclimatol Palaeoecol 395:9–20
Aguirre J, Domènech R, Martinell J, Mayoral E, Santos A, Pérez-Asensio JN (2017) Witnesses of the early Pliocene sea-level rise in the Manilva Basin (Málaga, S Spain). Span J Palaeontol 32:35–52
Asgaard U, Bromley RG (2008) Echinometrid sea urchins, their trophic styles and corresponding bioerosion. In: Wisshak M, Tapanila L (eds) Current developments in bioerosion. Springer-Verlag, Berlin, pp 279–303
Aubert D (1972) Le lapié fossile des Verrières (Doubs). Ann Sci Univ Besançon 3 Geol 17:85–88
Baarli BG, Santos AG, Mayoral EJ, Ledesma-Vázquez J, Johnson ME, Da Silva CM, Cachão M (2013) What Darwin did not see: Pleistocene fossil assemblages on a high-energy coast at Ponta das Bicudas, Santiago, Cape Verde Islands. Geol Mag 150:183–189
Bałuk W, Radwański A (1977) Organic communities and facies development of the Korytnica basin (Middle Miocene; Holy Cross Mountains, Central Poland). Acta Geol Pol 27:85–123
Bassi D, Humblet M, Iryu Y (2011) Recent ichnocoenosis in deep water macroids, Ryukyu Islands, Japan. Palaios 26:232–238
Bassi D, Iryu Y, Humblet M, Matsuda H, Machiyama H, Sasaki K, Matsuda S, Arai K, Inoue T (2012) Recent macroids on the Kikai-jima shelf, Central Ryukyu Islands, Japan. Sedimentol 59:2024–2041
Belaústegui Z, Muñiz F, Nebelsick JH, Domènech R, Martinell J (2017) Echinoderm ichnology: bioturbation, bioerosion and related processes. J Paleontol 91:643–661
Bichet V, Campy M, Cordier S, Riot G (2014) Inventaire régional, Patrimoine Géologique de Franche-Comté. https://zaaj.univ-fcomte.fr/IMG/pdf/1_synthese_irpg_fc_sept2014.pdf. Accessed 15 June 2020.
Boekschoten GJ (1966a) Shell borings of sessile epibiontic organisms as palaeoecological guides (with examples from the Dutch coast). Palaoegeogr Palaeoclimatol Palaeoecol 2:333–379
Boekschoten GJ (1966b) Shell boring of sessile epibiontic organisms as Palaeoecological guides (with examples from the Dutch coast). Palaeogeogr Palaeoclimatol Palaeoecol 2:333–379
Brlek M, Špišić M, Brčić V, Mišur I, Kurečić T, Miknić M, Avanić R, Vrsaljko D, Slovenec D (2016) Mid-Miocene (Badenian) transgression on Mesozoic basement rocks in the Mt. Medvednica área of northern Croatia. Facies 62:18. https://doi.org/10.1007/s10347-016-0470-z
Bromley RG (2004) A stratigraphy of marine bioerosion. Geol Soc London Spec Publ 228:455–479
Bromley RG, Asgaard U (1993a) Two bioerosion ichnofacies produced by early and late burial associated with sea-level change. Geol Rundsch 82:276–280
Bromley RG, Asgaard U (1993b) Endolithic community replacement on a Pliocene rocky coast. Ichnos 2:93–116
Bromley RG, D’Alessandro A (1983) Bioerosion in the Pleistocene of southern Italy: ichnogenera Caulostrepsis and Maeandropolydora. Riv Ital Paleontol Stratigr 89:283–309
Bromley RG, D’Alessandro A (1990) Comparative analysis of bioerosion in deep and shallow water, Pliocene to recent, Mediterranean Sea. Ichnos 1:43–49
Buoncristiani JF, Campy M (2004) Expansion and retreat of the Jura ice sheet (France) during the last glacial maximum. Sediment Geol 165:253–264
Cachão M, Silva CM, Santos A, Domènech R, Martinell J, Mayoral E (2009) The bioeroded megasurface of Oura (Algarve, south Portugal): Implications for the Neogene stratigraphy and tectonic evolution of southwest Iberia. Facies 55:213–225
Cachão M, Redweik P, Barreira E, Dinis J, Catita C, da Silva CM, Santos A, Mayoral E, Linder W (2011) Photogrammetric and spatial analysis of a bioeroded Early Miocene rocky shore, western Portugal. Facies 57:417–429
Carey S, Sparks RSJ, Tucker ME, Li T, Robinson L, Watt SFL, Gee M, Hastie A, Barfod DN, Stinton A, Leng M, Raineault N, Ballard RD (2020) The polygenetic Kahouanne Seamounts in the northern Lesser Antilles island arc: evidence for large-scale volcanic island subsidence. Mar Geol 419:106046. https://doi.org/10.1016/j.margeo.2019.106046
Charollais J, Wernli R, Chene RJD, Von Salis K, Steiner F (2006) La molasse marine supérieure de la Combe d’Evuaz et de la Pesse (Jura méridional, France). Arch Sci 59:21–46
Clauzon G, Rubino, JL, Savoye B (1995) Marine Pliocene Gilbert-type fan deltas along the French Mediterranean coast. A typical infill feature of preexisting subaerial Messinian canyons. In: 16th IAS Reg Meet Sedimentol-5ème Congr Fr Sédimentol, ASF Guideb, Publ Assoc Sedimentol Fr, Paris, 23:143–222
da Silva CM, Cãchao M, Martinell J, Domènech R (1995) Estruturas Bioerosivas como indicadores de Paleolitorais rochosos. O exemplo do Miocenico da Foz da Fonte (Sesimbra, Portugal). Dados preliminares. Mem Mus Lab Miner Geol FCUP 4:133–137
da Silva CM, Cachão M, Martinell J, Domènech R (1999) Bioerosional evidence of rocky palaeoshores in the Neogene of Portugal: Environmental and stratigraphical significance. Bull Geol Soc Den 45:156–160
Davidson TM, Grupe BM (2014) Habitat modification in tidepools by bioeroding sea urchins and implications for fine-scale community structure. Mar Ecol. https://doi.org/10.1111/maec.12134
de Gibert JM, Domènech R, Martinell J (2007) Bioerosion in shell beds from the Pliocene Roussillon Basin (SE France): significance for their paleoenvironmental interpretation and implications for the (macro)bioerosion Ichnofacies model. Acta Palaeontol Pol 52:783–798
de Gibert JM, Domènech R, Martinell J (2012) Rocky shorelines. In: Bromley RG, Knaust D (eds) Trace fossils as indicators of sedimentary environments, Developments in Sedimentology. Elseiver, Amsterdam, pp 441–462
Demircan H (2012) Determination of a Late Miocene rocky palaeoshore by bioerosion trace fossils from the Bozcaada Island, Çanakkale, Turkey. C R- Palevol 11:331–344
Domènech R, de Gibert JM, Martinell J (2001) Ichnological features of a marine transgression: middle Miocene rocky shores of Tarragona, Spain. Geobios 34:97–107
Domènech R, Martinell J, de Porta J (2008) Bioerosión por poliquetos espiónidos (Polychaeta, Spionidae) en moluscos marinos del Cuaternario caribeño de Colombia. Rev Acad Colomb Cienc 32(124):411–419
Domènech R, Juilleret J, Martinell J (2014) Delimitando la costa burdigaliense en el Jura francés mediante la bioerosión sobre acantilados. Fundamental 24:83–86
Dorador FJ, Rodríguez-Tovar FJ (2014) Digital image treatment applied to ichnological analysis of marine core sediments. Facies 60:39–44
Dorador FJ, Rodríguez-Tovar FJ (2018) High-resolution image treatment in ichnological core analysis: Initial steps, advances and prospects. Earth-Sci Rev 177:226–237
Doyle P, Bennett M, Cocks F (1998) Borings in a boulder substrate from the Miocene of southern Spain. Ichnos 5:277–286
Gibert JM, de Martinell J, Domènech R (1996) El Mioceno marino entre las playas de L’Arrabassada y El Miracle (Tarragona): aspectos paleontológicos e implicaciones sedimentológicas. Acta Geol Hisp 29:133–148
Gibert JM, Martinell J, Domènech R (1998) Entobia ichnofacies in fossil rocky shores, lower Pliocene, northwestern Mediterranean. Palaios 13:476–487
Gibson PH (1996) Distribution of the cirratulid polychaeta Dodecaceria fimbriata, D. concharum and D. diceria in European waters between latitudes 48°N and 70°N. J Mar Biol Assoc UK 76:625–635
Gibson PH (2017) A search for trace fossils of the burrowing cirratulid polychaetes Dodecaceria fimbriata and D. concharum. Ichnos 24:83–90
Gravina MF, Antonelli F, Sacco Perasso C, Cesaretti A, Casoli E (2019) The role of polychaetes in bioerosion of submerged mosaic floors in the Underwater Archaeological Park of Baiae (Naples, Italy). Facies 65:19. https://doi.org/10.1007/s10347-019-0563-6
Guillaume S, Guillaume A (1968) Carte géologique détaillée de la France à échelle 1:50.000. Feuille 605, Morez-Bois d’Amont. BRGM edit, Paris
Gutowski J, Machalski M (1984) A new littoral locality within the Middle Miocene (Badenian) Koytnica Basin (Holy Cross Mountains, Central Poland). Acta Geol Pol 34:195–201
Hanken NM, Uchman A, Jakobsen SL (2012) Late Pleistocene-early Holocene polychaete borings in north-east Spitsbergen and their palaeoecological and climatic implications: an example from the Basissletta area. Boreas 41:42–55. https://doi.org/10.1111/j.1502-3885.2011.00223.x
Hoffmann M, Friedrich AM (2017) Erosional and tectonic overprint of the mid-Miocene marine cliff line and its applicability as a paleo-geodetic marker of regional-scale tilting, Swabian Alb, southwestern Germany. In: Hoffmann M Young tectonic evolution of the Northern Alpine Foreland Basin, southern Germany, based on linking geomorphology and structural geology. Dissertation, Universität München, pp 67–116
Homberg C, Bergerat F, Philippe Y, Lacombe O, Angelier J (2002) Structural inheritance and Cenozoic stress fields in the Jura fold-and-thrust belt (France). Tectonophys 357:137–158
Homewood P, Rigassi D, Weidmann M (1989) Le bassin molassique Suisse. In: Assoc Sedimentol Fr (ed) Dynamique et méthodes d‘étude des bassins sédimentaires. Technip, Paris, pp 299–314
Johnson ME (1988a) Why are ancient rocky shores so uncommon? J Geol 96:469–480
Johnson ME (1988b) Hunting for ancient rocky shores. J Geol Educ 36:147–154
Johnson ME, Baarli BG (1999) Diversification of rocky shore biotas through geologic time. Geobios 32:257–273
Johnson ME, Baarli BG (2012) Development of intertidal biotas through Phanerozoic time. In: Talent JA (ed) Earth and Life. Global biodiversity, extinction intervals and biogeographic perturbations through time. Springer, Berlin, pp 63–128
Johnson ME, Baarli BG, Santos A, Mayoral E (2011) Ichnofacies and microbial build-ups on Late Miocene rocky shores from Menorca (Balearic Islands), Spain. Facies 57:255–265
Kempf M (1962) Recherches d’écologie comparée sur Paracentrotus lividus (Lmk) et Arbacia lixula (L.) (I.). Rec Trav Stat Mar Endoume 25:47–116
Laubscher H (2010) Jura, Alps and the boundary of the Adria subplate. Tectonophys 483:223–239
Martin D, Britayev TA (1998) Symbiotic polychaetes: Review of known species. Oceanogr and Mar Biol 36:217–340
Martin J, Pharisat A, Rangheard Y (1991) Le sinclinal des Verrières (Haute-Châine Jurassienne): Nouvelle interpretation structurale. Ann Sci Univ Franche-Comté Bésançon Géologie 4:99–112
Martinell J (1981) Actividad erosiva de Paracentrotus lividus (Lmk) (Echinodermata, Echinoidea) en el litoral gerundense. Oecol Acuat 5:219–225
Martinell J, Domènech R (1986) Actividad bioerosiva en el Plioceno del Empordà (Catalunya). Paleont Evol 20:247–251
Martinell J, Domènech R (1995) Bioerosive structures on the Pliocene rocky shores of Catalonia (Spain). Rev Esp Paleontol 10:37–44
Marty D, Belvedere M, Razzolini NL, Lockley MG, Meyer CA, Paratte G, Lovis C, Cattin M (2018) The tracks of giant theropods (Jurabrontes cuertedulensis and ichnosp. nov.) from the Late Jurassic of NW Switzerland: palaeocological and palaeogeographical impliations. Hist Biol 30:928–956
Mikuláš R (1992) Early Cretaceous borings from Štramberk (Czechoslovakia). Čas Miner Geol 37:297–312
Peres JM, Picard J (1964) Nouveau manuel de bionomie benthique de la la mer Méditerranée. Trav Stat Mar Endoume 31:1–137
Pharisat A, Rangheard Y (1986) Analyses granulométriques de sediments du Miocène inférieur de la Haute chaine Jurassienne. Ann Sci Univ Besançon 4 Geol 7:3–33
Pierçon M (1932) Le Miocène marin de la Haute-Chaîne du Jura. Bull Soc Hist Nat Doubs 41:54–85
Pineda-Salgado G, Quiroz-Barroso SA, Sour-Tovar F (2015) Analysis of bioerosion in clasts from a Miocene rocky-shore, Concepción Formation, Veracruz, México. Palaeogeogr Palaeoclim Palaeoecol 439:50–62
Radwański A (1970) Dependence of rock-borers and burrowers on the environmental conditions within the Tortonian littoral zone of Southern Poland. In: Crimes TP, Harper JC (eds) Trace fossils. Geol Jour Spec Issue. Wiley, New Jersrey, pp 371–390
Radwański A, Górka M, Wysocka A (2006) Middle Miocene coralgal facies at Maksymivka near Ternopil (Ukraine): a preliminary account. Acta Geol Pol 56:89–103
Radwański A, Wysocka A, Górka M (2011) ‘Entobia balls’ in the Medobory biohermal complex (Middle Mio-cene, Badenian; western Ukraine). Acta Geol Pol 61:265–276
Ramalho R, Helffrich G, Schmidt DN, Vance D (2010) Tracers of uplift and subsidence in the Cape Verde Archipelago. J Geol Soc Lond 167:519–538
Ramalho R, Quartau R, Trenhaile AS, Mitchell NC, Woodroffe CD, Ávila SP (2013) Coastal evolution on volcanic oceanic islands: a complex interplay between volcanism, erosion, sedimentation, sea-level change and biogenic production. Earth Sci Rev 127:140–170
Rangheard Y, Demarcq G, Mongereau N, Pharisat A, Pouyet S, Truc G (1985) Le Miocène du Val des Verrières et du Bief des Lavaux (Jura central, Haute-Chaîne): événements paléobiogéographiques et géodynamiques. Geobios 18:769–785
Rangheard Y, Demarq G, Müller C, Poignant A, Pharisat A (1990) Données Nouvelles sur le Burdigalien du Jura interne: paléobiologie, biostratigraphie et evolution structural. Bull Soc Géol Fr VI:479–486
Razzolini NL, Klein H (2018) Crossing slopes: Unusual trackways of recent birds and implications for tetrapod footprint preservation. Ichnos 25:252–259
Razzolini N, Vila B, Díaz-Martínez I, Manning PL, Galobart À (2016) Pes shape variation in an ornithopod dinosaur trackway (Lower Cretaceous, NW Spain): New evidence of an antalgic gait in the fossil track record. Cret Res 58:125–134. https://doi.org/10.1016/j.cretres.2015.10.012
Razzolini NL, Belvedere M, Marty D, Meyer C, Paratte G, Lovis C, Cattin M (2017) Megalosauripus transjuranicus ichnosp. nov. A new Late Jurassic theropod ichnotaxon from NW Switzerland and implications for tridactyl dinosaur ichnology and ichnotaxomy. PLoS ONE. https://doi.org/10.1371/journal.pone.0180289
Reiff W (1989) Das Kliff in Heldenfingen und die Klifflinie auf der Heidenheimer Alb. Jahrb Mitt Oberrh Geol Ver 71:467–482
Rodríguez-Tovar FJ, Dorador J (2015) Ichnofabric characterization in cores: a method of digital image treatment. Ann Soc Geol Pol 85:465–471
Rodríguez-Tovar FJ, Uchman A, Puga-Bernabéu A (2015) Borings in gneiss boulders in the Miocene (Upper Tortonian) of the Sorbas Basin, SE Spain. Geol Mag 152:287–297
Rodríguez-Tovar FJ, Dorador J, Mena A, Hernández-Molina FJ (2018) Lateral variability of ichnofabrics in marine cores: Improving sedimentary basin analysis using Computed Tomography images and high-resolution digital treatment. Mar Geol 397:72–78
Rosso A, Sanfilippo R, Ruggieri R, Maniscalco R, Vertino A (2015) Exceptional record of submarine cave communities from the Pleistocene of Sicily (Italy). Lethaia 48:133–144
Russell MP, Gibbs VK, Duwan E (2018) Bioerosion by pit-forming, temperate-reef sea urchins: History, rates and broader implications. PLoS ONE. https://doi.org/10.1371/journal.pone.0191278
Santos A, Mayoral E (2009) Paleoacantilados y bioerosión: dos ejemplos en el Neógeno Superior de la Cordillera Bética. Rev Soc Geol Esp 22:13–22
Santos A, Mayoral E, Marques da Silva C, Cachão M, Domènech R, Martinell J (2008) Trace fossil assemblages on Miocene rocky shores of southern Iberia. In: Wisshak M, Tapanila L (eds) Current developments in bioerosion. Springer-Verlag, Berlin, pp 431–450
Santos A, Mayoral E, da Silva CM, Cachão M, Kullberg JC (2010) Trypanites ichnofacies: Palaeoenvironmental and tectonic implications. A case study from the Miocene disconformity at Foz da Fonte (Lower Tagus Basin, Portugal). Palaoegeogr Palaeoclimatol Palaeoecol 292:35–43
Santos A, Mayoral E, Bromley RG (2011a) Bioerosive structures from Miocene marine mobile-substrate communities in Southern Spain, and description of a new sponge boring. Palaeontol 54:535–545
Santos A, Mayoral EJ, da Silva CM, Cachaõ M, Johnson ME, Baarli BG (2011b) Miocene intertidal zonation on a volcanically active shoreline: Porto Santo in the Madeira Archipelago, Portugal. Lethaia 44:26–32
Santos A, Mayoral E, Johnson ME, Baarli BG, Cachão M, da Silva CM, Ledesma-Vázquez J (2012a) Extreme habitat adaptation by boring bivalves on volcanically active paleoshores from North Atlantic Macaronesia. Facies 58:325–338
Santos A, Mayoral E, Johnson ME, Baarli BG, da Silva CM, Cachão M, Ledesma-Vázquez J (2012b) Basalt mounds and adjacent pits attract contrasting biofacies on a volcanically active Middle Miocene coastline (Porto Santo, Madeira Archipelago, Portugal). Facies 58:573–585. https://doi.org/10.1007/s10347-012-0301-9
Santos A, Mayoral E, Dumont CP, da Silva CM, Ávila SP, Baarli BG, Cachão M, Johnson ME, Ramalho RS (2015) Role of environmental change in rock-boring echinoid trace fossils. Palaeogeogr Palaeoclimatol Palaeoecol 432:1–14
Santos A, Mayoral E, da Silva CM, Cachão M (2016) Dois exemplos notáveis de paleolitorais rocosos bioerosionados do Neogénico de Portugal: Novos dados e síntese. Comun Geol 103:121–130
Schoppe S, Werding B (1996) The boreholes of the sea urchin genus Echinometra (Echinodermata: Echinoidea: Echinometridae) as a microhabitat in tropical South America. Mar Ecol 17:181–186
Steinthorsdottir M, Håkansson E (2017) Endo- and epilithic faunal succession in a Pliocene-Pleistocene cave on Rhodes, Greece: record of a transgression. Palaeontol 60:663–681
Tapanila L (2008) The endolithic guild: an ecological framework for residential cavities in hard substrates. In: Wisshak M, Tapanila L (eds) Current developments in bioerosion. Erlangen Earth Conference Series. Springer, Berlin, pp 3–20
Taylor PD, Wilson MA (2003) Palaeoecology and evolution of marine hard substrate communities. Earth-Sci Rev 62:1–103
Titschack J, Bromley RG, Freiwald A (2005) Plio-Pleistocene cliff-bound, wedge-shaped, warm-temperate carbonate deposits from Rhodes (Greece): sedimentology and facies. Sed Geol 180:29–56
Tribollet A, Golubic S (2005) Cross-shelf differences in the pattern and pace of bioerosion of experimental carbonate substrates exposed for 3 years on the northern Great Barrier Reef, Australia. Coral Reefs 24:422–434
Uchman A, Demircan H, Toker V, Derman AS, Sevim S, Szulc J (2002) Relative sea-level changes recorded in borings from a Miocene rocky shore of the Mut basin, southern Turkey. Ann Soc Geol Pol 72:263–270
Voigt E (1970) Endolithische Wurm-Tunnelbauten (Lapispecus cuniculus n.g. n.sp. und Dodecaceria [?] sp.) in Brandungsgeröllen der oberen Kreide im nördlichen Harzvorlande. Geol Rundsch 60:355–380
Voigt E (1971) Fremdskulpturen an Steinkernen von Polychaeten-Bohrgängen aus der Maastrichter Tuffkreide. Paläontol Z 45:144–153
Wattkins R (1990) Paleoecology of a pliocene rocky shoreline, salton trough region, California. Palaios 5:167–175
Weidmann M (2008) La Molasse de la Vallée de Joux (Jura, Suisse et France). Bull Soc Vaud Scien Nat 91:69–101
Wisshak M (2010) High-latitude bioerosion. The Kosterfjord experiment. Lectures notes in earth sciences. Springer-Verlag, Berlin
Wisshak M, Gektidis M, Freiwald A, Lundälv T (2005) Bioerosion along bathymetric gradient in a cold temperate setting (Kosterfijord, SW Sweden): an experimental study. Facies 51:93–117
Ziegler PA, Fraefel M (2009) Response of drainage systems to Neogene evolution of Jura fold-thrust belt and Upper Rhine Graben. Swiss J Geosci 102:57–76. https://doi.org/10.1007/978-94-007-6704-1_26
Acknowledgements
Co-authors RD and JM were supported by the research project CGL 2015-66835-P/BTE of the Spanish Ministry of Economy and Competitiveness. Authors are grateful to A Gallardo (Technical Services of the Earth and Ocean Dynamics Dept., Universitat de Barcelona) for his efficient assistance in the treatment of hand samples, and to J Calvet (Earth and Ocean Dynamics Dept, Universitat de Barcelona) for his essential implication in the digital treatment of the images. Ll Martinell revised the language of a first version of the text. Final English editing by GG Buffett, physical scientist and native English speaker (www.terranova.barcelona). Special thanks to the reviewers JP Suc and A Uchman and the editor M Tucker who greately helped to improve the manuscript. In memory of Ms Juilleret Christiane, who welcomed the co-authors during the fieldwork. I love you mom (JJ).
Funding
R.D. and J.M. were supported by the research project CGL 2015-66835-P/BTE of the Spanish Ministry of Economy and Competitiveness.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Not applicable.
Rights and permissions
About this article

Cite this article
Martinell, J., Juilleret, J. & Domènech, R. Early Miocene (Burdigalian) marine rocky shores in the French Jura Massif: a palaeoecological and palaeoenvironmental analysis. Facies 67, 2 (2021). https://doi.org/10.1007/s10347-020-00610-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10347-020-00610-z