
Abstract
Recent research undertaken in Mediterranean pine forests suggests that the establishment of Pinus pinea L. and Pinus pinaster Ait. natural regeneration is determined by different spatial and temporal factors. In this context, we analyzed the establishment and success of natural regeneration in these two species and we examined the hypothesis that the spatial pattern of natural regeneration depends on adult tree presence, density or stand composition, while temporal patterns are typically driven by climatic conditions. For the purposes of the experiment we used 1936 plots of 0.02 ha established in mixed and pure forests of P. pinea and P. pinaster in the Northern Plateau of Spain. Plots were installed following regeneration fellings, and regeneration was monitored annually between 2001 and 2015. We used contingency tables and principal component analysis (PCA) to evaluate the main factors affecting the spatial distribution of the regeneration. PCA was also used to assess the temporal variation as a function of climatic variables. Finally, we checked for the existence of interspecific and intraspecific synchrony through Tau-Kendall synchrony analysis. We found that natural regeneration of P. pinea was more successful than that of P. pinaster over the whole of the studied period. Contingency tables showed that P. pinaster regeneration was associated with conspecific adult trees, while the relationship between P. pinea regeneration and adult trees varied depending on the regeneration development stage, i.e., seedling or saplings. Furthermore, we found that natural regeneration in both species could be enhanced through the presence of mixtures of adult trees. The spatial variability, analyzed through PCA, showed similar responses in both species, although temporal variability associated with climate variables differed between P. pinea and P. pinaster. Finally, we found different synchrony patterns affecting natural regeneration, depending on the species and the regeneration stages. In this context, temporal patterns seem to be species specific during the sapling stage of regeneration. These findings allow the observed regeneration patterns in Mediterranean pinewoods to be generalized at forest management scale.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Many of the projected future changes in climate, as well as the indirect effects of these changes, are likely to have a negative impact on forest systems (Keenan 2015). Mediterranean forests are considered among the most vulnerable to climate change (Bernier and Schoene 2009), and the increase in drought frequency, which is already being observed, has been shown to have a negative impact on the long-term establishment of this type of forests. Among other effects, an increase in drought events might have an adverse impact on plant regeneration (Doblas-Miranda et al. 2016), which is considered a key factor when evaluating the capacity of forest systems to adapt (Gómez-Aparicio et al. 2008). Forest regeneration is a successional process in which the success of the establishment phase is essential to ensure the long-term persistence of the forest. During this process, the ontogenic developmental stages in which natural regeneration is recognized and identified (seedling or sapling) are influenced differently by the vegetation structure, species composition and functional attributes (Walker et al. 2007). In this context, the species-specific requirements and limitations (incoming light, site and climate conditions, vegetation structure, etc.) play a key role, which may explain why the regeneration of one species is more successful than the other in forests where the two species coexist. Moreover, these specific requirements and limitations could hold the key to developing a strategy to improve the resilience of Mediterranean forests. This idea has been proposed in several studies (Kolström et al. 2011; Pretzsch et al. 2013; Forrester 2014; del Río et al. 2016), which advocate the use of mixed species stands as one of the main adaptation strategies in forest management due to the greater efficiency of these stands as regards resource uptake, resistance to pests and diseases and greater resilience in the face of climate change.
Through appropriate management we can control the overstory vegetation that intercepts the incoming light and competes for ground resources (Stancioiu and O’hara 2006a, b), thus achieving vigorous regeneration in both mixed and pure forests. It is therefore essential to determine how regeneration fellings contribute to and interact with the regeneration process, taking into account the species-specific regeneration strategies (Yoshida et al. 2005; Stancioiu and O’Hara 2006a, b; Webb and Jarrett 2013). These strategies mainly relate to the degree of shade tolerance, water use efficiency or resistance to drought of the species, although other factors such as the frequency of biotic and abiotic disturbances (e.g., pests and diseases, fire or windthrow) also influence regeneration success. The joint occurrence of these intrinsic and extrinsic factors leads to a diversity of forest dynamics (Oliver and Larson 1996). Thus, it is important not only to identify the factors that affect natural regeneration but also to determine how the effects can be modified through different management options.
Natural regeneration of Mediterranean pine forests is often unsuccessful due to the existence of certain limiting factors. The main bottlenecks for regeneration are directly linked to either the intrinsic characteristics of the species (e.g., seed size, masting), to those of the habitat (e.g., soil compaction) or to climatic conditions (mainly summer drought). However, inadequate forest management can also lead to regeneration failure (Calama et al. 2017). In the Northern Plateau of Spain, Pinus pinea L. and P. pinaster Ait. occur in both pure and mixed stands and share not only territory, but also ecological conditions and historical management traits (Calama et al. 2017). P. pinea and P. pinaster have traditionally been favored over other species due to the high economic importance of edible pine nut production in the case of P. pinea and timber and resin collection associated with P. pinaster (Gordo et al. 2012). Unfortunately, in certain areas, natural regeneration in both species is far from successful and the problem is getting worse under the current climate change scenarios. Various studies have been undertaken in this field in order to identify the main limitations affecting natural regeneration (Gordo et al. 2012). Manso et al. (2014) observed that the regeneration of P. pinea is limited by mast-year occurrence, seed dispersal limitations and seed predation, all of which are influenced by climatic conditions. Furthermore, Calama et al. (2015) found that the joint occurrence of severe water stress and negative rates of net assimilation (circumstances commonly found during the summer), result in P. pinea seedling mortality. Natural regeneration in P. pinaster is mainly limited by seedling establishment and survival during the first summer after emergence (Del Peso et al. 2012). It is important to consider the initial conditions required by each species, in accordance with the species-specific shade needs for germination and initial seedling growth and survival. In this regard, regeneration in both species is highly favored by water availability in soils (Rodríguez-García et al. 2011a, b). Mid-shade positions provide the optimal conditions required by P. pinaster regeneration (Ruano et al. 2009), while P. pinea regeneration is favored by the presence of dense shade environments during the first stages of regeneration, followed by progressive and flexible release of the seedlings (Manso et al. 2014). These conditions are especially important during the hot, dry summer, since shade and adequate levels of water availability in soils prevent water stress conditions, reducing the air temperature and evaporative demand (Valladares et al. 2005).
Studies concerning the natural regeneration of forests have generally adopted one of the two following approaches: (1) to study the different processes involved in successful natural regeneration (seed production, seed dispersal, seed predation, germination, emergence, survival and initial seedling growth) (Pukkala and Kolström 2008; Price et al. 2001; Ordóñez et al. 2006; Manso et al. 2013) or (2) to study regeneration by focusing on the established regeneration; linking the probability of success to site conditions and adult tree composition (Grassi et al. 2004; Rodríguez-García et al. 2007; Eerikäinen et al. 2007). In the first approach, the regeneration process is considered a set of sequential states defined by the probability of transition from one to another (Pardos et al. 2012). In the second approach a wider range of spatial heterogeneity can be addressed since the sampling tends to be easier (Pardos et al. 2012). However, scarce research has been undertaken into the way in which different environmental factors and silvicultural practices can modulate natural regeneration at greater spatial scales.
In this study we used a network of 1936 permanent plots established in pure and mixed forests to study the natural regeneration of P. pinea and P. pinaster in the Northern Plateau of Spain. The plots cover 3529.9 ha in which regeneration fellings were carried out and were monitored over a 15-year period to identify the key factors influencing natural regeneration in these Mediterranean forests. We hypothesize that: (1) P. pinea and P. pinaster seedlings and saplings have different ecological requirements and limitations, which could result in interspecific differences in the rate of establishment and success of the natural regeneration; (2) the occurrence and the abundance of adult trees of one of the species could have a significant effect on the natural regeneration not only of that species but also of the other species; hence, regeneration fellings could be regulated to modify future stand composition; (3) the effect of climatic drivers on natural regeneration is not constant across species and/or regeneration stages. Thus, the specific aims of the study were (1) to evaluate the establishment and success of natural regeneration of P. pinea and P. pinaster in pure and mixed Mediterranean forests following regeneration fellings; (2) to analyze the recruitment of seedlings and saplings of both species, depending on adult stand composition, stocking and maturity; (3) to identify whether there is intra- and interspecific synchrony in regeneration establishment and developmental stage change and to determine how temporal variation in establishment is related to climatic conditions.
Materials and methods
Study site
The study was conducted in pure and mixed forests of P. pinea and P. pinaster located at 700 m a.s.l. on the Northern Plateau of Spain. In this region, the two species cover approximately 68.000 ha, with pure P. pinea stands covering 70% of the area, pure P. pinaster 10% and mixed forest of the two species, 20% (PORF 2008). The climate is continental-Mediterranean characterized by absolute temperatures in summer and winter of 40 °C and − 10 °C, respectively, with a mean annual temperature of 11.2 °C. Mean annual precipitation is 435 mm, and there is a period of summer drought (July to September) with mean precipitation of 66 mm. Frosts may occur from September to May. The main soil types found in the area are shown in Table 1. From an ecological perspective, Calama et al. (2008) proposed a stratification for the region based on soil, lithology and climatic attributes, resulting in ten groups denominated natural units. Hence, the plots used in this study are distributed among three very homogenous natural units 6, 7 and 8, so no spatial pattern was observed in the studied area (Table 1).
Clear-cutting followed by direct sowing of seeds collected from adjacent stands in rows was the system most commonly used in the Northern Plateau for both species until the 1970s. However, this method was replaced by the uniform shelterwood system, involving an initial seeding felling in which dominated, co-dominant and poor condition dominant trees are removed. Secondary fellings are subsequently applied every 5–10 years over the following 20 years (Montero et al. 2008). Rotations in pure stands of P. pinea currently range from 100 to 120 years, while for pure stands of P. pinaster the rotation is around 80–100 years (Calama et al. 2017). In mixed stands, regeneration fellings tend to mimic the practices proposed for the dominant species, since no specific guidelines for these mixtures are currently available.
Experimental design
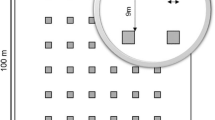
The data were gathered from a network of permanent plots established to monitor natural regeneration following regeneration fellings in the study area (Gordo et al. 2012). The plots were systematically located in 32 public forests (13,951 ha) within the study area (Fig. 1), which are divided into blocks and stands for management planning. The first plots were installed in 2001, covering forest blocks where the first shelterwood felling had already been carried out, but they were still in the regeneration period. Since then, new blocks where the regeneration period has begun have gradually been included in the monitored area. At the end of 2015 the number of available plots was 1936. The plots are circular with a radius of 8 m (0.02 ha). The stock quadrant method was used to monitor the regeneration annually (Stein 1992), dividing the plots into four quadrants according to cardinal points.

Distribution of the monospecific and mixed stands of P. pinea and P. pinaster as well as the distribution of the permanent plots that comprise the dataset from Valladolid province. The upper right corner of the figure shows a zoom of the systematic distribution of the plots
Dataset
Depending on the size and viability of the monitored plants, natural regeneration was classified into two stages of development: well-established plants with the presence of adult needles, hereafter termed “saplings”; and young plants showing uncertain future, with juvenile needles and height commonly below 20 cm, hereafter termed “seedlings.” The number of plants of each category in each quadrant of the plot was recorded annually, between October and December from 2001 to 2015, along with the level of grass cover (high, medium and low) and the number of adult trees per species in each plot. According to the composition of the adult stratum of each plot they were classified as pure adult P. pinea plots (accounting for 56.8% of the plots, 1100), pure adult P. pinaster plots (16.4%, 318 plots) and plots with mixed composition, and the proportion was 5.9% (115 plots). The remaining 20.9% (402 plots) corresponds to plots where no adult trees of any of the species remain standing in the plot after regeneration fellings were applied.
In addition, other general information as regards stand structure was obtained from the forest management inventories at block and stand level—i.e., forest management unit. The variables used in this study at this scale were age, dominant height, quadratic mean diameter, basal area and number of adult trees per ha by species, along with the Reineke stand density index. Finally, we used the number of years since the regeneration felling as a potential explanatory covariable (Table 2).
Climate data were obtained from the nearby meteorological stations (Valladolid, 04°46′27″W; 41°38′402N; 735 m and Olmedo 04°41′15″W; 41°17′19″N; 769 m [data available at www.inforiego.org]). Seasonal and yearly precipitation and mean temperatures were calculated for the study period (2001–2015) (Table 3).
Regeneration occurrence and success
The occurrence of regeneration during the studied period was analyzed as a proxy to evaluate the capacity of the territory to engender regeneration. For this purpose, plots were classified into: Pp, plots where only P. pinea regeneration has been identified during the studied period; Pt, plots in which only P. pinaster regeneration has occurred; Mixed, plots where regeneration of both species has taken place; and No_Reg, plots in which no regeneration has been recorded during the studied period. In this analysis, the identification of a single plant during the whole period was sufficient to consider that regeneration had occurred.
In addition, to evaluate the success of the regeneration at the end of period, we studied the presence of saplings of both species in the last year of the study (2015), analyzing the distribution of observed frequencies and presenting the percentage of plots in which saplings of P. pinea, P. pinaster, both species or no saplings at all were present.
Statistical analysis
The spatial pattern of natural regeneration was studied using the records from the last year of the study (2015) in order to observe the most recent spatial relationships at two different spatial scales: plot and forest stand. Spearman’s correlation analysis was carried out to determine whether there was a significant correlation among the two species and development stages at plot level. To this end, we applied the analysis in plots where recruitment of at least one species is present, leaving out the 0, 0 combinations (no presence of regeneration at all) which could obscure the results. We also analyzed the association at plot level between the presence/absence of natural regeneration of P. pinea and P. pinaster and the composition of the stratum of adult trees by means of χ2 contingency tables. The composition of the adult tree stratum was classified into four categories: (1) pure P. pinea, (2) pure P. pinaster, (3) mixed and (4) plots without adult trees. This analysis was carried out separately for seedlings and saplings.
Principal component analysis (PCA) was used to determine the dependence between adult stand characteristics and the natural regeneration of both species at stand level. Variables included in the PCA were the stand variables presented in Table 2, the number of years since regeneration fellings started, the level of grass and the average number of seedlings and saplings of each species recorded in the last year of the study (2015).
We also used PCA to identify the main temporal-varying factors influencing the natural regeneration. In this case we used the annual ingrowth, defined as the observed difference between the number of plants recorded in two consecutive years for each developmental stage and species. In the case of seedlings, a positive value can be interpreted as an indicator of seedling recruitment, whereas for saplings a positive value in this covariate is a proxy of the change from seedling to sapling, thus a change in ontogeny phase. As potential temporal influencing factors, we evaluated the annual and seasonal averaged values of the different climatic variables corresponding to the current and the previous year (Table 3).
The Kendall coefficient of concordance, W (Sokal and Rohlf 1995), was used to evaluate the degree of concordance in interannual variation of regeneration (synchrony). Kendall’s W is a nonparametric measure of agreement among several complete sets of rankings of a number of objects, in this case, series of number of individuals by species or development stages—seedlings or saplings. Kendall’s W ranges from 0 (no association) to 1 (perfect association). In order to determine the existence of common temporal pattern, the analysis was performed in three different ways. First, we calculated the Kendall coefficient of concordance, W, to determine whether each regeneration stage, seedlings or saplings, of each of the two species follows the same temporal pattern between the three natural units (Case A). Here, the P value indicates whether synchrony exists (or not) between the three natural units in each of the analyzed cases. Subsequently, within each natural unit, we used Kendall’s W to study: (1) whether the two developmental stages of regeneration of the same species shared the same temporal pattern (intraspecific synchrony, Case B); and (2) the occurrence of an interspecific synchrony between species at the same developmental stage (interspecific synchrony, Case C).
The statistical analyses were carried out using the R software environment (R Core Team 2017), specifically the “descr” package for contingency table analysis (Aquino 2016) and the “vegan” package for multivariate PCAs and synchrony analysis (Oksanen et al. 2018).
Results
Regeneration success
During the whole studied period (15 years), natural regeneration of either of the two studied species at any developmental stage was recorded in 83% of the sampling plots. Hence, 332 out of the 1939 plots (17%) showed no regeneration at all of either P. pinea or P. pinaster during the whole period (Fig. 2a).

Diagrams showing frequencies of occurrence of regeneration for a seedlings and saplings during the complete study period and b saplings during the last studied year, 2015. (Pp) refers to plots where only regeneration of P. pinea has been observed; (Pt) refers to plots where only regeneration of P. pinaster has been observed; (Mix) plots where regeneration of both species has been observed; (No Reg) plots without regeneration
When each species was analyzed separately, the number of plots with P. pinea regeneration was 1598 plots, while P. pinaster regeneration was recorded in 445 plots (23% of the plots) during this period (Fig. 2a). Only 40 (2%) of these plots presented pure P. pinaster regeneration, while in the rest of plots, 405 (21%), the natural regeneration observed was of mixed composition (Fig. 2a).
As regards successfully established saplings, only 35% of the plots had established saplings of any species at the end of the monitoring period (Fig. 2b). P. pinaster saplings were only observed in 5% of the plots, while the figure was 33% in the case of P. pinea. In this regard, it was possible to identify a pattern of zero abundance and asymmetry in the distribution of frequencies for the number of established saplings per plot, which would indicate that most of the successful natural regeneration occurred in only a few parts of our study area (Fig. 3).

Histogram of frequencies for the number of P. pinea and P. pinaster saplings observed per plot in the last year of the study (2015)
Spatial analysis
Using Spearman’s rank correlation coefficient to check for a significant spatial correlation between the regeneration stages and species at plot level, we identified a significant negative correlation between the number of P. pinea seedlings and P. pinaster seedlings and saplings. Conversely, we found a significant positive correlation between the number of P. pinea and P. pinaster saplings. Moreover, a significant positive, strong correlation was observed between seedlings and saplings belonging to the same species (Table 4).
The association between natural regeneration and the composition of the stratum of adult trees at plot level is presented in Table 5. The significant values indicate the existence of dependence between different natural regeneration stages and the presence of adult trees. In the case of P. pinaster a strong association was observed between the natural regeneration and the composition of the adult stratum, specifically with conspecific adult trees (203 plots with regeneration vs. 63.4 expected plots under a random distribution of regeneration). Moreover, there was scarce presence of P. pinaster regeneration in plots with only P. pinea adult trees (66 observed plots vs. 219.4 expected under a random distribution). As regards P. pinea, the contingency table showed no significant differences among observed and expected values for saplings, while an influence of conspecific adult trees over the presence of P. pinea seedlings was observed. A mixed composition of adult trees had a beneficial effect on natural regeneration, especially in the case of P. pinaster.
The PCA showed that in the four analyzed cases—P. pinea seedlings and saplings/P. pinaster seedlings and saplings—the PC1 explained approximately 25% of the variability and the PC2 around 20%. In the analysis we observed that seedlings of both species showed a close, positive relationship with the basal area of conspecific adult trees (Fig. 4a, b). For P. pinaster seedlings, we also found a positive influence of the number of P. pinaster adult trees (Fig. 4b). In the case of saplings, these variables—basal area and number of conspecific adult trees—had less impact (Fig. 4c, d). Moreover, negative relationships were observed between the number of seedlings of each species and the stand density index (SDI). Similarly, saplings of the two species displayed negative relationships with this variable, although in the case of P. pinaster this relationship was closer. Other variables that seemed to have an influence on natural regeneration were the number of years since fellings had been carried out as well as the age of the stand and the mean squared diameter of adult trees. In this case, the magnitude of the variables used in the analysis is related to the degree of relevance of these same variables in PC1 and PC2. Thus, it is likely that SDI, which is at least one order of magnitude larger than all the other variables, will also have a large variance and, therefore, a greater probability of becoming highly relevant in the first component. This could also occur with the basal area and the number of trees in conspecific situations in the second component.

Results for the first two axes of the PCA combining stand variables and natural regeneration; aP. pinea seedlings; bP. pinaster seedlings; cP. pinea saplings; and dP. pinaster saplings. Abbreviations as shown in Table 2
Temporal association
The percentage of variability explained by PCA using climatic variables was around 30% for the first axis and 20% for the second, for both P. pinea and P. pinaster. It was observed that the association between climate and natural regeneration changed depending on the time lag used for the climatic variable as well as on the species (Fig. 5). PCA revealed that there was a perfect, opposite relationship between the annual ingrowth (observed difference in the number of seedlings or saplings between two consecutive years) in both species and the mean annual and spring temperature of the current year (Fig. 5a, c). In this regard, sapling ingrowth also showed a negative relationship with the mean autumn temperature and a positive, close relationship with the mean autumn precipitation of the current year.

Results for the first two axes of the PCA combining climatic variables and annual ingrowth natural regeneration; aP. pinea and P. pinaster seedlings and current year climatic variables, bP. pinea and P. pinaster seedlings and previous year climatic variables, cP. pinea and P. pinaster saplings and current climatic variables, dP. pinea and P. pinaster saplings and previous year climatic variables. Abbreviations as shown in Table 3
Interestingly, mean summer precipitation of the previous year seems to have a perfect, positive relationship with the annual ingrowth of seedlings, especially for P. Pinaster seedlings. In contrast, an opposite relationship was found between the current value of this variable and the annual ingrowth of saplings. Similar relationships (positive association with the value for the previous year and opposite relationship with the value for the current year) were observed for the rest of analyzed climatic variables (Fig. 5b, c).
The Tau-Kendall analysis revealed an intraspecific regeneration synchrony between the three natural units for the variation in number of seedlings of the two species (P value < 0.05, Case A of Table 6). In the case of P. pinaster, the value of W (0.851) reveals an almost perfect synchrony over the studied period. In contrast, in the case of saplings we only observed this intraspecific synchrony among natural units in P. pinea. No synchronized temporal patterns were observed within each natural unit between seedlings and saplings belonging to the same species (Case B of Table 6). Lastly, the analysis revealed an interspecific synchrony in the variation of number of seedlings in two of the natural units (6 and 8), with values of W (0.838 and 0.881) close to 1. No interspecific synchrony was observed in the case of saplings (Case C of Table 6).
Discussion
Natural regeneration in pinewood forests on the Northern Plateau of Spain exhibited large heterogeneity linked to density and composition of adult stands and to interannual variability in climatic conditions. Although the density of established saplings is at a critical level for the two studied species, our results related to the spatio-temporal analyses show that regeneration success was greater for P. pinea than for P. pinaster regardless of the structure of the adult stand (Table 5), confirming our first hypothesis and pointing to better adaptation of P. pinea to current climatic conditions. The large scale used in our study for the analysis of the spatial and temporal regeneration variability complements the findings obtained in previous studies, mainly designed at smaller scales, allowing us to generalize the observed regeneration patterns at forest management scale.
Drivers of spatial variation in regeneration success
The significant negative spatial correlation between P. pinea and P. pinaster seedlings indicates differences related to the initial conditions required by each species, in accordance with the species-specific shade needs for germination and initial seedling growth and survival (Ruano et al. 2009; Manso et al. 2014). However, the observed positive correlation between saplings of both species suggests similar requirements for sapling development. These requirements could be related to a similar demand for light at this stage and the density of adult trees in the plot. This idea is supported by Barbeito et al. 2008 and Rodríguez-García et al. (2011a), who conducted studies concerning P. pinea and P. pinaster and found similar requirements at more advanced stages of regeneration. Furthermore, we must consider the existence of other uncontrolled factors, such as soil water retention, which could have a significant influence on regeneration success under these conditions. Surprisingly, we detected a significant negative correlation between the presence of P. pinea seedlings and P. pinaster saplings, but a nonsignificant relationship between P. pinaster seedlings and P. pinea saplings. This finding could be related to the fact that the denser and more shaded conditions required for P. pinea seedling establishment prevent not only the establishment of P. pinaster seedlings, but also the transition to the sapling developmental stage of the established P. pinaster seedlings. All these results confirm our first hypothesis, by defining how the different light and shade requirements of the species affect the establishment and success of the natural regeneration.
As stated in our second hypothesis, the presence and composition of adult tress had a significant influence over the two regeneration stages. P. pinea seedlings were mainly favored by the presence of conspecific adult trees (Table 5), in accordance with previous findings in monospecific stands at smaller scales. Calama et al. (2015) found that crown cover favors the survival of P. pinea seedlings, mitigating the negative effect of high irradiance and drought. In this species, the primary dispersal agent is gravity (Manso et al. 2012), which could also explain this positive association. However, our results for P. pinea saplings revealed that presence was only lower than expected under conspecific conditions, suggesting that P. pinea saplings must be progressively released from overstory cover, probably due to an increase in light requirements. Manso et al. (2014) found that as seedlings grow, their performance is optimized in more exposed environments. The observed negative relationship of P. pinea saplings with the SDI variable in the PCA reinforces this theory. In addition, P. pinea recruits manage to survive beneath P. pinaster crowns, probably as a consequence of the higher light availability under P. pinaster as opposed to P. pinea crowns (Barbeito et al. 2008), suggesting that the mixed composition of adult trees is a plausible alternative.
In the case of P. pinaster, we found that both seedling and sapling establishment occurred in close proximity to conspecific adult individuals (Table 5; Fig. 3), and even though regeneration in P. pinaster is generally less successful, recruits of this species were still found in about half of the 318 pure conspecific plots. Although P. pinaster generally exhibits a shade-intolerant behavior, our results suggest that under the light and climate conditions present in the study site, canopy protection by conspecific trees against the extreme summer drought favors regeneration in this species. Rodríguez-García et al. (2011a) and Ruano et al. (2009) reported that a certain level of canopy cover is required for P. pinaster seedling survival and establishment, particularly during the water stress season. However, despite the fact that P. pinaster seeds may be able to reach P. pinea stands as they are mainly wind-dispersed, we found limited P. pinaster regeneration under nonconspecific trees. This fact could be related to the dense shade conditions caused by P. pinea crowns, which are much denser than those of P. pinaster and therefore probably hamper successful natural regeneration in the latter species. Rodriguez et al. (2008) confirm the shade-intolerant nature of this species, providing examples of greater seedling densities at sites with open rather than closed canopies. These somewhat contradictory results highlight the importance of considering foliar distribution and overstory canopy structure to characterize species as shade-tolerant or not, according to abiotic conditions (Martínez and Tapias 2005). Given the positive influence of P. pinaster adult trees over conspecific regeneration, retaining a sufficient number of P. pinaster adult trees to ensure successful regeneration should be considered when performing regeneration fellings.
The positive effect of mixed composition in adult stands on natural regeneration of the two species is noteworthy, even though this mixed composition was only observed in a few plots. Canopy cover comprising species with different crown features can create different light and environmental conditions, allowing the development of species with distinct shade tolerances (Stancioiu and O’hara 2006a). Note also that this situation could be advantageous over pure composition due to differences in existing seed sources. These facts would explain the results obtained in mixed plots. In our species composition, light availability in mixtures can differ considerably from that of monospecific stands due to the differences in crown widths and foliar densities of the studied species, providing different niches for the development of each (Moreno-Fernández et al. 2017). However, quantifying the availability of light and its effect on natural regeneration in complex stands requires detailed measurements (Lochhead and Comeau 2012), often carried out at local scales (Moreno-Fernández et al. 2017). Such measurements are not feasible at large scales like that of our study. Conversely, by exploring relationships between observed regeneration and adult stand characteristics at landscape scale it would be possible to provide meaningful information regarding the role of mixed species composition on natural regeneration.
Intra- and interspecific synchrony
The results of Kendall coefficient of concordance, W (Table 6), highlight the important role of climate variation on the success of natural regeneration in Mediterranean pinewoods (Calama et al. 2017). The synchrony found in the seedling and sapling time series for each species among the three natural units, with the exception of P. pinaster saplings, suggests that temporal variation in the number of seedlings and saplings per species was mainly driven by interannual climatic conditions, which are similar across the three natural units. Conversely, the lack of synchrony among natural units in P. pinaster saplings (Case A, Table 6) may indicate a stronger dependence at this stage of regeneration on other factors apart from annual climatic conditions. However, this result could be affected by the large number of plots that presented no natural regeneration at all (Fig. 2a, b), making it difficult to detect synchrony. Accordingly, the lack of intraspecific synchrony between young seedlings and saplings in these two species, analyzed inside each natural unit (Case B, Table 6), underlines the different climatic requirements shown by the two regeneration stages, as has been reported for other Mediterranean species (Davis et al. 2011). As regards interspecific synchrony, our results again highlight the difference between regeneration stages (Table 6). In the two natural units with lower water retention (6 and 8), there was synchrony between the seedling time series of the two species (case C), suggesting a similar dependence on climatic conditions, probably linked to successful germination and seedling emergence in the previous year, as well as summer drought survival in the current year (Calama et al. 2017). These findings show the complexity of the natural regeneration process, since our observations suggest that regeneration in P. pinea and P. pinaster presents similar dependence on climatic conditions while at the same time displays differences with regard to the initial spatial conditions. In this sense, the probability of natural regeneration occurring in one or other species could be more related to the presence/absence of adult trees, i.e., with a spatial requirement. After this initial life stage, both species seem to display similar variations with climate, revealing the dependence of this developmental stage on climatic conditions. In contrast, the lack of interspecific synchrony in the sapling stage could suggest that this stage is more closely related to stand characteristics than with climatic factors.
Climatic drivers
The results of the PCA support our third hypothesis that the effect of climatic drivers is not constant across species and/or regeneration stages. Favorable conditions as regards annual and spring mean temperatures could promote seedling establishment and the ontogenic step to the next regeneration stage. Calama et al. (2015) highlighted the importance of optimum root system development in P. pinea seedlings during the period prior to summer to improve water uptake in the summer months. Our findings may reflect this theory in the sense that an increase in spring temperatures and scarce spring precipitation could limit the development of the root system, affecting water uptake during the following months and therefore hindering the transition to the next ontogenic stage. In addition, a notable positive relationship was observed between warmer winters and ingrowth for both seedlings and saplings, which given the high frost tolerance observed for pine species in the region (Pardos et al. 2014) may be related to a lengthening of the period with optimal conditions for seedling and sapling growth (Calama et al. 2015). Furthermore, the relationship with the annual and spring mean temperatures is stronger in the case of P. pinaster than for P. pinea, possibly due to the better adaptation of P. pinea to sandy soils (Moreno-Fernández et al. 2017), which restrict the natural establishment and development of P. pinaster in years with extreme climatic conditions such as high temperatures and severe drought (Rodríguez-García et al. 2011b; Calama et al. 2017). In general, this climatic effect may be more direct at the seedling stage, since they are more sensitive to extreme climatic conditions than saplings (Cavender-Bares and Bazzaz 2000; Castro et al. 2004).
Overall, P. pinaster is a species with a narrow range of optimal precipitation conditions (Gandullo and Sánchez-Palomares 1994). The significance of this climatic variable on natural regeneration of P. pinaster has been addressed in previous studies (Rodríguez et al. 2008; Rodríguez-García et al. 2011a, b). Ruano et al. (2009) observed that summer rainfall had a significant influence on germination, early development, and on total and viable seedling density in P. pinaster; the availability of water being of greater influence than light on the growth of P. pinaster regeneration. These authors also found differing effects of water supply on seedling development according to the time lag used. We detected a significant positive relationship between current year seedling ingrowth and the precipitation levels of the summer and fall seasons of the previous year, probably due to the positive effect that this water availability has on the emergence and survival of recently emerged seedlings which, in the following year, will enter the seedling stage. In contrast, precipitation of the previous year has no effect on sapling ingrowth, indicating that changes in ontogeny from seedling to sapling are mainly linked to current year conditions. According to future climate scenario predictions for the Mediterranean basin (Brunet et al. 2009), it is likely that this narrow range of optimal precipitation conditions will be reduced, which could seriously limit the long-term establishment of P. pinaster in the study area.
However, although these adverse climatic factors predicted for the studied area are not controllable, stand management can be used to facilitate natural regeneration. Our results could help managers to decide when and how to open the forest, releasing regeneration in order to favor different species in the future stand. Thus, to favor P. pinaster regeneration it would be necessary to maintain an adequate number of P. pinaster trees when regeneration fellings are performed. To favor P. pinea regeneration, however, it would be necessary to maintain denser conditions, provided mainly by conspecific crown cover, along with progressive release of the overstory cover once establishment of P. pinea seedlings is observed.
These findings could constitute a starting point for future studies on the role of ontogeny in the regeneration process. In this sense, the identification of the spatial and temporal factors controlling the growth and survival on each ontogenic phase and triggering the change from seedling to sapling is considered a key issue in the studies of natural regeneration on forest tree species.
Conclusions
Based on our findings, we conclude that natural generation in P. pinea is more successful than that of P. pinaster in the studied area, at both seedling and sapling stages. Species-specific differences during the seedlings stage, shown by the negative correlations between both species, are probably related to the initial conditions required by each species. In the two developmental stages of both species, the composition of the stratum of adult trees seems to play a significant role. The interspecific synchrony shown by seedlings reveals the higher sensitivity of this developmental stage to extreme climatic conditions. In contrast, the lack of synchrony shown by saplings, as well as the existence of a spatial correlation in the occurrence of saplings of both species, suggests that at landscape scale there is a greater dependence at this stage on site and spatially explicit attributes. Optimal regeneration establishment seems to be related to favorable conditions as regards annual and spring mean temperatures. Precipitation levels of the summer and fall seasons of the previous and current year were found to have different effects on seedling and sapling ingrowth, reflecting the complex interaction existing between environmental factors and the optimum conditions for natural regeneration.
References
Aquino J (2016) Includes R source code and/or documentation written by Dirk Enzmann, Marc Schwartz, Nitin Jain and Stefan Kraft (2018). descr: Descriptive Statistics. R package version 1.1.4. https://CRAN.R-project.org/package=descr
Barbeito I, Pardos M, Calama R, Canellas I (2008) Effect of stand structure on Stone pine (Pinus pinea L.) regeneration dynamics. Forestry 81:617–629. https://doi.org/10.1093/forestry/cpn037
Bernier P-Y, Schoene D (2009) Adapting forests and their management to climate change: an overview
Brunet M, Casado MJ, Castro M, et al (2009) Generación de escenarios regionalizados de cambio climático para España. Minist Medio Ambient Medio Rural y Mar, p 158
Calama R, Barbeito I, Pardos M et al (2008) Adapting a model for even-aged Pinus pinea L. stands to complex multi-aged structures. For Ecol Manag 256:1390–1399. https://doi.org/10.1016/j.foreco.2008.06.050
Calama R, Puértolas J, Madrigal G, Pardos M (2013) Modeling the environmental response of leaf net photosynthesis in Pinus pinea L. natural regeneration. Ecol Modell 251:9–21. https://doi.org/10.1016/j.ecolmodel.2012.11.029
Calama R, Puértolas J, Manso R, Pardos M (2015) Defining the optimal regeneration niche for Pinus pinea L. through physiology-based models for seedling survival and carbon assimilation. Trees Struct Funct 29:1761–1771. https://doi.org/10.1007/s00468-015-1257-5
Calama R, Manso R, Lucas-Borja ME et al (2017) Natural regeneration in Iberian pines: a review of dynamic processes and proposals for management. For Syst 26:eR02S. https://doi.org/10.5424/fs/2017262-11255
Castro J, Zamora R, Hodar JA, Gomez JM (2004) Seedling establishment of a boreal tree species (Pinus sylvestris) at its southernmost distribution limit: consequences of being in a marginal Mediterranean habitat. J Ecol 92:266–277. https://doi.org/10.1111/j.0022-0477.2004.00870.x
Cavender-Bares J, Bazzaz FA (2000) Changes in drought response strategies with ontogeny in Quercus rubra: implications for scaling from seedlings to mature trees. Oecologia 124:8–18. https://doi.org/10.1007/PL00008865
Davis FW, Tyler CM, Mahall BE (2011) Consumer control of oak demography in a Mediterranean-climate savanna. Ecosphere 2:art108. https://doi.org/10.1890/es11-00187.1
Del Peso C, Bravo F, Ruano I, Pando V (2012) Patrones de diseminación y nascencia de Pinus pinaster Ait. En la Meseta castellana. In: Gordo J, Calama R, Pardos M, Bravo F, Montero G (eds) La regeneración natural de los pinares en los arenales de la meseta castellana. Instituto Universitario de Investigación en Gestión ForestalSostenible, Valladolid, pp 161–175
del Río M, Pretzsch H, Alberdi I et al (2016) Characterization of the structure, dynamics, and productivity of mixed-species stands: review and perspectives. Eur J For Res 135:23–49. https://doi.org/10.1007/s10342-015-0927-6
Doblas-Miranda E, Alonso R, Arnan X et al (2016) A review of the combination among global change factors in forests, shrublands and pastures of the Mediterranean Region: beyond drought effects. Glob Planet Change 148:42–54. https://doi.org/10.1016/j.gloplacha.2016.11.012
Eerikäinen K, Miina J, Valkonen S (2007) Models for the regeneration establishment and the development of established seedlings in uneven-aged, Norway spruce dominated forest stands of southern Finland. For Ecol Manag 242(2–3):444–461
Forrester DI (2014) The spatial and temporal dynamics of species interactions in mixed-species forests: from pattern to process. For Ecol Manag 312:282–292. https://doi.org/10.1016/j.foreco.2013.10.003
Gandullo J., Sánchez-Palomares O (1994) Estaciones ecológicas de los pinares españoles. Minist Agric Pesca y Aliment
Gómez-Aparicio L, Pérez-Ramos IM, Mendoza I et al (2008) Oak seedling survival and growth along resource gradients in Mediterranean forests: implications for regeneration in current and future environmental scenarios. Oikos 117:1683–1699. https://doi.org/10.1111/j.1600-0706.2008.16814.x
Gordo F, Rojo LI, Calama R, Al E (2012) Selvicultura de regeneración natural de Pinus pinea L. en montes públicos de la provincia de Valladolid. In: Gordo J, Calama R, Pardos M, Bravo F, Montero G (eds) La regeneración natural de los pinares en los arenales de la meseta castellana. Instituto Universitario de Investigación en Gestión ForestalSostenible, Valladolid, pp 145–159
Grassi G, Minotta G, Tonon G, Bagnaresi U (2004) Dynamics of Norway spruce and silver fir natural regeneration in a mixed stand under uneven-aged management. Can J For Res 34(1):141–149
Keenan RJ (2015) Climate change impacts and adaptation in forest management: a review. Ann For Sci 72:145–167. https://doi.org/10.1007/s13595-014-0446-5
Kolström M, Lindner M, Vilén T et al (2011) Reviewing the science and implementation of climate change adaptation measures in European forestry. Forests 2:961–982. https://doi.org/10.3390/f2040961
Lochhead KD, Comeau PG (2012) Relationships between forest structure, understorey light and regeneration in complex Douglas-fir dominated stands in south-eastern British Columbia. For Ecol Manag 284:12–22. https://doi.org/10.1016/j.foreco.2012.07.029
Manso R, Pardos M, Keyes CR, Calama R (2012) Modelling the spatio-temporal pattern of primary dispersal in stone pine (Pinus pinea L.) stands in the Northern Plateau (Spain). Ecol Modell 226:11–21. https://doi.org/10.1016/j.ecolmodel.2011.11.028
Manso R, Calama R, Madrigal G, Pardos M (2013) A silviculture-oriented spatio-temporal model for germination in Pinus pinea L. in the Spanish Northern Plateau based on a direct seeding experiment. Eur J For Res 132(5–6):969–982
Manso R, Pukkala T, Pardos M et al (2014) Modelling Pinus pinea forest management to attain natural regeneration under present and future climatic scenarios. Can J For Res 44:250–262. https://doi.org/10.1139/cjfr-2013-0179
Martínez M, Tapias R (2005) Photosynthetic response to light and temperature of Spanish pines young seedlings. In: International conference of conservation, regeneration and restoration of mediterranean pines and their ecosystems MEDPINE
Montero G, Calama R, Ruiz Peinado R (2008) Selvicultura de Pinus pinea. In: Montero G, Serrada R, Reque J (eds) Compendio de Selvicultura de Especies. Ministerio de Educación y Ciencia, Madrid, pp 431–470
Moreno-Fernández D, Montes F, Sánchez-González M et al (2017) Regeneration dynamics of mixed stands of Pinus pinaster Ait. and Pinus pinea L. in Central Spain. Eur J For Res. https://doi.org/10.1007/s10342-017-1086-8
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara LB, Simpson GL, Solymos P, Stevens MHM, Szoecs E, Wagner H (2018) Vegan: community ecology package. R package version 2.5-3. https://CRAN.R-project.org/package=vegan
Oliver CD, Larson BC (1996) Forest stand dynamics. Wiley, New York
Ordóñez JL, Molowny-Horas R, Retana J (2006) A model of the recruitment of Pinus nigra from unburned edges after large wildfires. Ecol Modell 197(3–4):405–417
Pardos M, Bravo F, Gordo J, Montero G, Calama R (2012) La investigación en regeneración natural de las masas forestales. In: Gordo J, Calama R, Pardos M, Bravo F, Montero G (eds) La regeneración natural de los pinares en los arenales de la meseta castellana. Instituto Universitario de Investigación en Gestión ForestalSostenible, Valladolid, pp 17–36
Pardos M, Climent J, Almeida H, Calama R (2014) The role of developmental stage in frost tolerance of Pinus pinea L. seedlings and saplings. Ann For Sci 71:551–562. https://doi.org/10.1007/s13595-014-0361-9
PORF (2008) Plan de ordenación de los recursos forestales de la provincia de Valladolid. Servicio Territorial de Medio Ambiente. Junta de Castilla y León. http://www.jcyl.es/web/jcyl/MedioAmbiente/es/Plantilla100DetalleFeed/1246988359553/Noticia/1200229949988/Comunicacion
Pretzsch H, Schütze G, Uhl E (2013) Resistance of European tree species to drought stress in mixed versus pure forests: evidence of stress release by inter-specific facilitation. Plant Biol 15:483–495. https://doi.org/10.1111/j.1438-8677.2012.00670.x
Price DT, Zimmermann NE, van der Meer PJ, Lexer MJ, Leadley P, Jorritsma IT, Schaber J, Clark DF, Lasch P, McNulty S, Wu J (2001) Regeneration in gap models: priority issues for studying forest responses to climate change. Clim Chang 51:475–508
Pukkala T, Kolström T (2008) A stochastic spatial regeneration model for Pinus sylvestris. Scand J For Res 7(1–4):377–385
R Core Team (2017) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Rodríguez-García E, Juez L, Guerra B, Bravo F (2007) Natural regeneration analysis of Pinus pinaster Ait. inthe sandy areas of Almazan-Bayubas. (Soria, Spain). Investigación Agraria: Sistemas y Recursos Forestales 16(1):25
Rodríguez R, Serrada R, Lucas, JA, Alejano R, Del Río M, Torres E, Cantero A (2008) Selvicultura de Pinus pinaster Ait. subp. mesogeensis Fieschi & Gaussen. In: Serrada R, Montero G, Reque J (eds) Compendio de selvicultura aplicada en España.
Rodríguez-García E, Gratzer G, Bravo F (2011a) Climatic variability and other site factor influences on natural regeneration of Pinus pinaster Ait. in Mediterranean forests. Ann For Sci 68:811–823. https://doi.org/10.1007/s13595-011-0078-y
Rodríguez-García E, Bravo F, Spies TA (2011b) Effects of overstorey canopy, plant-plant interactions and soil properties on Mediterranean maritime pine seedling dynamics. For Ecol Manag 262:244–251. https://doi.org/10.1016/j.foreco.2011.03.029
Ruano I, Pando V, Bravo F (2009) How do light and water influence Pinus pinaster Ait. germination and early seedling development? For Ecol Manag 258:2647–2653. https://doi.org/10.1016/j.foreco.2009.09.027
Sokal R, Rohlf F (1995) Biometry: the principles and practice of statistics in biological research, 3rd edn. W.H. Freemanm, New York
Stancioiu PT, O’hara KL (2006a) Regeneration growth in different light environments of mixed species, multiaged, mountainous forests of Romania. Eur J For Res 125:151–162. https://doi.org/10.1007/s10342-005-0069-3
Stancioiu PT, O’Hara KL (2006b) Morphological plasticity of regeneration subject to different levels of canopy cover in mixed-species, multiaged forests of the Romanian Carpathians. Trees Struct Funct 20:196–209. https://doi.org/10.1007/s00468-005-0026-2
Stein WI (1992) Regeneration surveys and evaluation. In: Hobbs S, Tesch S, Owston P (eds) Reforestation practices in southwestern Oregon and northern California. Forest Research Laboratory, Oregon State University, Corvallis, pp 346–382
Valladares F, Dobarro I, Sánchez-Gómez D, Pearcy RW (2005) Photoinhibition and drought in Mediterranean woody saplings: scaling effects and interactions in sun and shade phenotypes. J Exp Bot 56:483–494. https://doi.org/10.1093/jxb/eri037
Walker RF, Fecko RM, Frederick WB et al (2007) Forest health impacts of bark beetles, dwarf mistletoe, and blister rust in a Lake Tahoe Basin mixed conifer stand. West N Am Nat 67:562–571. https://doi.org/10.3398/1527-0904(2007)67%5b562:FHIOBB%5d2.0.CO;2
Webb AA, Jarrett BW (2013) Hydrological response to wildfire, integrated logging and dry mixed species eucalypt forest regeneration: the Yambulla experiment. For Ecol Manag 306:107–117. https://doi.org/10.1016/j.foreco.2013.06.020
Yoshida T, Iga Y, Ozawa M et al (2005) Factors influencing early vegetation establishment following soil scarification in a mixed forest in northern Japan. Can J For Res 35:175–188. https://doi.org/10.1139/x04-156
Acknowledgements
We are grateful to the Forest Service of the Junta de Castilla y León for conducting the field experiment. The author thanks Inforiego (www.inforiego.org) of the Instituto Tecnológico Agrario de Castilla y León for the climatic data. This contribution has been supported within the framework of projects AGL2014-51964-C2-2-R of Spanish Ministry of Economy and Competitiveness, CC16-095 PROPINEA, between INIA, ITACYL and Diputación de Valladolid and RTA2013-00011-C2.1.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Christian Ammer.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Vergarechea, M., del Río, M., Gordo, J. et al. Spatio-temporal variation of natural regeneration in Pinus pinea and Pinus pinaster Mediterranean forests in Spain. Eur J Forest Res 138, 313–326 (2019). https://doi.org/10.1007/s10342-019-01172-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10342-019-01172-8