
Abstract
This study was carried out to determine bud and phloem tissue damage rates after cold treatments in 12 different grape cultivars. The single-bud cuttings tested at −10 °C and −15 °C during 18 h in freezer. The study was conducted in two group; one group is related with the damage rates of the winter buds and phloem tissue, the second group tested in cold applied cuttings in the growing room to be determined of the cuttings performance. The damage ratio of the winter buds and phloem tissue was evaluated by the cross-sectioning method. Freeze-tested single-bud cuttings were forced to sprouting in the growing room condition. The sprouting rate (%), cluster per shoot (%), root and shoot length (cm), root scale (0–4), root number values, and basal callus formation rate (%) were determined in the cuttings. According to results; while ‘Karaerik’ and ‘Ugni Blanc’ cultivars showed the highest primary bud viability in −15 °C (83.3% and 78.3%, respectively); ‘Çavuş’ showed the lowest primary bud viability rates with 0.02%. The highest phloem viability at −15 °C was determined in ‘Narince’ (70.0%) and ‘Alphonse Lavallee’ cultivars (65.0%). In the growing room condition; the highest root number, root length, and root scale were determined as in ‘Ugni Blanc’ cultivar subjected to −15 °C. In this study; according to bud cross-sectioning and growing room results, the least affected cultivars by cold applications were found as ‘Karaerik’, ‘Ugni Blanc’, ‘Cardinal’ and ‘Alphonse Lavallee’ cultivars.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The cultivation of grapevines, or viticulture, is an integral and historic part of agriculture worldwide, with the European grapevine species, Vitis vinifera, playing a central role (Santos et al. 2020; Petitpierre et al. 2023). Within this extensive spread, there are regions highly suitable for vine growth, but also areas where environmental stress factors limit productivity and quality, posing economic challenges for vine cultivation (Bernardo et al. 2018). Particularly in continental climates, successful viticulture faces many limiting factors, one of which is freezing temperatures. Recent global climate changes, though seemingly reducing the risk of low-temperature damage with rising temperatures, also bring significant concerns (Droulia and Charalampopoulos 2021; Mirás-Avalos and Araujo 2021; Benítez-Cabello et al. 2023). The associated droughts and the reduction of protective snow cover, especially in areas that rely on this cover, are thought to increase the risk of frost damage (Frank et al. 2015; Walsh et al. 2020; Wan et al. 2023). The issue of frost damage in viticulture is multifaceted, involving the interplay of the vine’s physiological, morphological, and genetic characteristics with varying environmental factors and cultural practices (Eriş 1982; Kara et al. 2005). Vines adopt strategies like leaf fall or dormancy, essentially halting growth in all tissues, to protect sensitive tissues from frost damage (Howell and Shaulis 1980; Lang 1987; Bates et al. 2002; Zapata et al. 2004). However, these adaptations do not render the vines entirely cold-tolerant, especially under prolonged exposure to freezing temperatures, leading to cane and trunk injuries (Wolfe 2001; Dami et al. 2012). This study also considers the phenomenon of deacclimation, where warm periods during the acclimation stage can cause vines to lose their cold tolerance prematurely (Antivilo et al. 2018). The implications of low temperatures on crop yield, quality, and survival are profound (Alberdi and Corcuera 1991; Zabadal et al. 2007). Most Vitis vinifera cultivars are sensitive to winter cold, presenting a significant challenge in cold-continental climates. These cultivars may suffer frost damage at temperatures below −15 to −25 °C, depending on the period and variety (Fennell 2004; Kaya and Köse 2018; Karimi 2020). In Turkey, for instance, winter temperatures have caused severe damage to vine buds. In the winter of 2006–2007, temperatures of −23.4 °C in Diyarbakır province’s vineyards resulted in up to 99% death in primary buds (Karataş et al. 2008). Similarly, in the Üzümlü district of Erzincan province, the ‘Karaerik’ cultivar suffered around 64% damage in primary buds due to temperatures dropping to −22.2 °C in 2008 (Köse and Güleryüz 2009). Furthermore, Kalkan et al. (2011) reported an 81.6% primary bud damage in the ‘Karaerik’ grape cultivar during the 2007–2008 winter season, with recorded temperatures as low as −30.7 °C. he cultivation of frost-tolerant grape cultivars is crucial for adapting viticulture to the increasing severity of winter conditions due to climate change, ensuring both the economic viability and environmental sustainability of this important agricultural sector. This approach not only mitigates the risk of significant crop losses but also supports the preservation of diverse grape varieties essential for the wine industry’s resilience and future growth. Therefore, this study focuses on understanding the challenges faced by grapevines in cold-continental climates, particularly the threat posed by severe winter conditions. The aim is to assess the level of frost damage across 12 different Vitis vinifera cultivars and provide insights into enhancing their resilience in suitable climatic regions.
Materials and Methods
Plant Materials
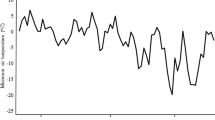
This research was conducted in Ondokuz Mayıs University in 2020–2021 winter season. In this study, it was aimed to determine the degree of damage caused by freeze on the bud and phloem tissue during the deacclimation period in some grape cultivars. The weather temperatures oc the vineyard recorded with HOBO data logger (Onset Computer Corp., Pocasset, MA). The average temperatures for the year in which the experiment was conducted are shown in Fig. 1.

Air temperature values of the vineyard
Maturity times of the cultivars are presented in Table 1.
Methods
The winter canes were taken from the grapevines belonging to the Vitis vinifera L. grape cultivars in the deacclimation stage (second week of the March) at Bafra Research Vineyard in Samsun, Turkiye. One-year-old canes containing 10 buds were taken from each vines and divided into two groups. First group used for artificial frost test. The second group were planted in the moist perlite filled floam boxes were mantained in the growing room after freeze applications. The planted cuttings were kept at ± 26 °C, 80% humidity and 16 h photoperiod for a month.
In the study, −10 °C and −15 °C temperatures tested in the cuttings. After the freezing test was performed on the single-bud cuttings in the first group, cross sections were taken from the bud and phloem tisse with a razor blade and the damage rates in the primary, secondary and tertiary buds and phloem viability rates were determined. The single-bud canes in the second group were planted in the foam boxes filled with perlite medium after the freeze test application. The sprouting, and rooting parameters of the cuttings were examined in the growing room.
Freeze Applications
Freeze applications were performed according to the modified method applied by Sivritepe et al. (2001). There were 4 replications and 40 single-bud canes used in each replication. Single-bud cuttings were prapared from annual canes and they wrapped in aluminum foil to prevent water loss (Antivilo et al. 2020). The freeze application was made with the help of a cooling cabinet (Elektro-mag M7040R, Turkiye) with a sensitivity of 0.1 °C. The single bud cuttings were kept at +4 °C for 24 h before the cold test.
The freeze test, the single-bud cuttings were placed in a freezer cabinet with an initial temperature of +4 °C. After the samples were kept at this temperature for 1 h, the freezer temperature was reduced to +3 degrees. From this temperature, the temperatures were gradually reduced by 3 degrees every two hours. Temperatures were applied as +3, 0, −3, −3, −6, −9, −12, −15 °C degrees. When the cabin temperature reached −10 and −15 degrees, the samples were kept at these temperatures for 18 h and they were removed from the freezing cabinet and placed back into the cooler at +4 degrees. 12 h after, the single-bud cuttings were removed from the freezer cabinet, removed from the aluminum foil and placed in paper cups filled with distilled water in order to detect living and dead buds and phloem tissue easily for 24 h at laboratory temperature of +25 degrees (Wolf and Pool 1987; Stergios and Howell 1977; Kaya 2020; Kalkan et al. 2017).
Enzymatic Browning Test in Buds and Phloem (Cross-Sectional Method)
Each bud was cut horizontally with the help of a razor blade. The rate of alive or dead primary, secondary, and tertiary buds and phloem tissue was examined under a stereomicroscope. The buds seen as green under the stereomicroscope were evaluated as alive; the buds seen as black or brown-black were evaluated as dead (Fig. 2). The percentage of death in the buds was determined by dividing the number of dead buds by the total number of buds (Köse and Güleryüz 2009; Karimi 2020).

Freeze damage in primary, secondary, and tertiary buds (a Tertiary bud alive; b Secondary bud dead, other two buds alive; c Primary bud dead; d all three buds alive Photos taken by Bülent Köse)
Determination of The Water Content of The Buds (%)
The fresh weight (FW) of the vine buds was determined on the canes taken from vineyard befor freeze test. After take the fresh weight of the samples, the dry weights (DW) of the samples calculated after dried at 65 °C for 48 h. It was calculated with the equation [% water content= (DW) × 100/FW] (Ershadi et al. 2016; Kaya 2020).
Determinig of The Climate Room Performance
To determine the damage caused by cold application to the winter buds and phloem tissue of the cultivars, the sprouting and rooting performances of single-bud cuttings in the growing room were examined. The sprouting process was applied to the winter buds in the climate room. For this purpose, single-bud cuttings at −10 °C and −15 °C were subjected to force sprouting in the growing room. The cuttings were planted in 400 × 40 × 25 cm foam boxes in moist perlite with 4 replications and 40 single-bud cuttings in each replication. The planted cuttings were kept at ± 26 °C, 80% humidity and 16 h photoperiod for a month (Fig. 3).

Growing room investigations after freeze tested cuttings
Statistical Analysing
The experiment was conducted according to the randomized blocks design. One-year-old canes taken from each vine with 10 buds and divided into two groups. First group used for artificial frost test, and the second group were planted in the moist perlite filled floam boxes after freeze applications and mantained in the growing room. For each group, 4 replications and 40 single bud canes were used. The rate of alive or dead primary, secondary, and tertiary buds and phloem tissue was examined after cold application. The winter buds and phloem tissue of the cultivars, were evaluated in terms of sprouting and rooting performances of single-bud cuttings in the growing room. Statistical analysis program SPSS 16.0 was used to evaluate the data. Differences between the cultivars and the cold applications tested as compared by Duncan’s multiple range test. Differences between the cultivars and the applications were determined by the LSD (P ≤ 0.05) test.
Results
The viability rates of primary, secondary, and tertiary buds were determined after freeze applications. It has been observed that the freeze test has statistically significant effects on primary, secondary, and tertiary buds of the data obtained as a result of the applications (p < 0.05). When primary bud viability was evaluated, the highest viability rate was obtained in ‘Karaerik’ cultivar as 96.7% at −10 °C and 83.3% at −15 °C (Table 2). Although the highest primary bud viability was obtained at −10 °C with 90.0% in the ‘Çavuş’ cultivar, when the temperature dropped to −15 °C all primary buds died.
The cultivars with more than 80% viability observed in the primary buds at −10 °C temperature were determined as ‘Karaerik’ (96.7%), ‘Narince’ (95.0%), ‘Alphonse Lavallée’ (91.7%), ‘Ugni Blanc’ (90.0%), ‘Çavuş’ (90.0%), respectively. In the study; the highest viability rate in primary buds at −15 °C temperature was found in the ‘Karaerik’ (83.3%) and ‘Ugni Blanc’ (78.3%) cultivars, while the lowest value was determined in the ‘Çavuş’ (0.02%). ‘Ugni Blanc’, ‘Karaerik’, ‘Alphonse Lavallée’, ‘Narince’, and ‘Victoria’ showed more than 50% primary bud viability at −15 °C temperature. The cultivars showing less than 10% primary bud viability rate at −15 °C temperature in the primary buds determined in ‘Michele Palieri’, ‘Royal’, ‘Early Sweet’, and ‘Çavuş’ (0.2%) cultivars, respectively (Table 1).
The differences between the cultivars was found to be statistically significant in terms of secondary bud viability (p ≤ 0.05). According to the results of secondary buds; the highest secondary bud viability at −10 °C was at ‘Red Globe’ (66.7%) cultivar and the lowest was obtained from ‘Early Sweet’ (0.15%) (Table 3). Cultivars with more than 50% viability in secondary buds were respectively; ‘Çavuş’ (95.0%), and ‘Ugni Blanc’ (90.0%), ‘Alphonse Lavallee’ (85.5%), and ‘Karaerik’ (80.0%) cultivars. When the temperature is reduced to −15 degrees, the viability rates in the secondary buds decreased significantly. However, in the ‘Ugni Blanc’ cultivar, secondary buds were able to maintain 70% of their viability at −15 degrees. Apart from ‘Ugni Blanc’, the cultivars determined to have high viability in secondary buds were ‘Alphonse Lavallee’ (53.3%), ‘Karaerik’ (48.3%), ‘Narince’ (35.0%), ‘Cardinal’ (31.7%), ‘Red Globe’ (31.7%) and ‘Victoria’ (26.7%). On the other hand, the viability of the secondary buds was below 10% in the ‘Çavuş’, ‘Tekirdağ Seedless’, ‘Early Sweet’, ‘Michele Palieri’ and ‘Royal’ cultivars (Table 3).
As a result of freeze applications, tertiary bud viability was show differences according to cultivars and cold degrees (P ≤ 0.05). While the highest tertiary buds vitality value was found in ‘Çavuş’ (96.7%), and ‘Ugni Blanc’ (91.7%), the lowest was found in ‘Tekirdağ Seedless’ (8.3%) at −10 °C temperature (Table 4). Among the cultivars, rates showing tertiary bud viability over 50% at −15 degrees were determined in ‘Ugni Blanc’ (65.0%), ‘Karaerik’ (58.3%) and ‘Narince’ (56.7%), respectively. The grape cultivars showing less than 10% tertiary bud viability at −15 °C were identified as ‘Royal’, ‘Tekirdağ Seedless’, ‘Early Sweet’ and ‘Michele Palieri’, respectively.
According to phloem tissue viability rates, significantly differences were obtained in terms of cultivars and cold degrees (P ≤ 0.05). Pholem tissue viability rates changed between 93.3% and 41.7% at −10 °C cold applied cuttings. Phloem viability rates was as 93.3% in ‘Alphonse Lavallee’, and 90.0% in ‘Çavuş’ cultivar. On the other hand, ‘Tekirdağ Seedless’ and ‘Early Sweet’ was take lowest phloem viability rate at −10 °C. The highest phloem viability rate obtained in ‘Narince’ (70.0%), ‘Alphonse Lavallee’ (65.0%), ‘Karaerik’ (61.7%) cultivars at −15 °C. The study found that the ‘Çavuş’ and ‘Early Sweet’ grape cultivars exhibited high sensitivity in terms of phloem tissue hardiness (Table 5).
Water content (%) of the buds was measured before the cold test. The highest bud water content was found in the ‘Early Sweet’ cultivar (53.7%). ’Ugni Blanc’ had the lowest water content among all grape cultivars. (25.2%) (Fig. 4). There is a negative correlation between tissue water content and cold tolerance (Wolpert and Howel 1986; Kaya 2020; Zhao et al. 2020). A decrease in tissue water content is associated with an increase in frost tolerance and is thought to increase the ability to supercooling (Ambroise et al. 2020; Karimi 2020; Babajamali et al. 2022). In our conducted this study, although they were in the same group, the water content of these cultivars such as ’Karaerik’, ’Ugni Blanc’, ’Cardinal’, ’Narince’, ’Alphonse Lavalle’ were slightly lower than the others. Similarly, the cold hardiness of these cultivars with lower water content was also higher than the others.

Water contents of the grape buds. Different letters in the same column indicate statistically significant differences (P ≤ 0.05)
Regarding bud and phloem viability results after cold treatments at the cuttings, as the cold hardy grape cultivars as evaluated in Table 6. Given viability rates, the highest bud viability rates for primary, secondary and tertiary buds has been determined in ‘Ugni Blanc’, ‘Karaerik’, ‘Alphonse Lavallee’, and ‘Narince’ cultivars. Considering the bud and phloem viability rates, it was determined that the ‘Ugni Banc’ stands out in terms of cold hardiness.
In the research; the effects of cold temperatures on the sprouting, shoot growth and rooting performances of cold applied cuttings in the growing room were examined (Table 7). Statistically significant differences were obtained in the tested samples (p ≤ 0.05). Within the −10 °C applied group, the highest sprouting rate was determined in ‘Ugni Blanc’ (100%) and ‘Çavuş’ (100%) cultivars, while the lowest rate was determined in ‘Early Sweet’ (30%) cultivar. The highest sprouting rates at 10 °C was determined in the ‘Ugni Blanc’ (100%), ‘Çavuş’ (100%), ‘Karaerik’ (96.7%), ‘Cardinal’ (96.7%), ‘Red Globe’ (96.7%), ‘Narince’ (93.3%), ‘Alphonse Lavallee’ (90%) cultivars. When the tested effects of −15 °C on cuttingsi the sprouting rates was decrease dramatically among all cultivars. In the group with application temperature of −15 °C, the highest sprouting rate was found in ‘Ugni Blanc’ (96.7%) and ‘Karaerik’ (93.3%). On the other hand, the lowest was in ‘Early Sweet’ (0.02%). The sprouting rates of cultivars were under 50% were determined as ‘Tekirdağ Seedless’ (16.7%), ‘Çavuş’ (6.7%), ‘Michelle Palieri’ (6.7%), ‘Royal’ (3.3%), ‘Early Sweet’ (0.02%), respectively.
The differences were found in terms of shoot length of the cuttings both cultivars and cold levels (P ≤ 0.05). As a result of freeze applications, at −10 °C, the longest shoot was determined in ‘Çavuş’ (11.4 cm), ‘Cardinal’ (10.7 cm), ‘Karaerik’ (11.1 cm) and cultivars. The shortest shoot was determined in ‘Early Sweet’ (0.01 cm) cultivar at a temperature −10 °C. The shoot lengths of the cultivars were decrease dramatically when the cuttings tested in −15 °C. The longest shoot was obtained in ‘Karaerik’ (9.6 cm) and ‘Cardinal’ (7.9 cm) cultivars. As a result of the study, there is almost no shoot length was measured in the cultivars ‘Çavuş’, ‘Royal’, ‘Tekirdağ Seedless’, ‘Early Sweet’ and ‘Michele Palieri’. The cultivars showing a shoot length than 5 cm were determined as ‘Karaerik’ (9.7 cm), ‘Cardinal’ (7.9 cm), ‘Ugni Blanc’ (5.3 cm), respectively, in −15 °C.
In the growing room, the number of clusters on the shoots of single-bud cuttings was determined. In both tested cold temperatures, the highest cluster number per shoot was obtained in ‘Karaerik’, ‘Ugni Blanc’ and ‘Cardinal’ cultivars. ‘Early Sweet’, ‘Çavuş’, ‘Narince’, ‘Tekirdağ Seedless’, and ‘Michele Palieri’ cultivars were not show any cluster on the shoot in −15 °C. In the study, While ‘Çavuş’, ‘Narince’, ‘Royal’, ‘Tekirdağ Seedless’, ‘Early Sweet’, and ‘Michele Palieri’ evaluated as very sensitive, ‘Karaerik’, ‘Ugni Blanc’ and ‘Cardinal’ cultivars were evaluated as cold hardy grapes in terms of Sprouting, shoot length and cluster numbers of the shoot (Table 7).
One month later in the growth room; the cuttings were removed from the perlite and the number of root number, root lengths, rooting and basal callus rates were counted (Table 8). Significantly differences were found both cultivar and cold degrees (p ≤ 0.05). The rooting ratios were changes between 96.7% (Ugni Blanc) and 20.0% (Early Sweet) at −10 °C. When the cold degrees ingrease, the rooting rates decreased except for ‘Ugni Blanc’ (96.7%). In −15 °C; ‘Tekirdağ Seedless’ and ‘Early Sweet’ cultivars were not any rooting performance. The cultivars showing rooting performance of more than 50% are respectively; ‘Ugni Blanc’ (96.7%), ‘Karaerik’ (53.3%) and ‘Red Globe’ (53.3%). On the other hand, the highest rate of basal callus formation of cuttings at −15 °C was ‘Alphonse Lavalle’ (66.7%). There were no basal callus formation occurred in ‘Çavuş’ and ‘Early Sweet’ cultivars.
Root lengths of single-bud cuttings subjected to cold applied showed significantly differences according to the cultivars and the cold degrees (P ≤ 0.05). When the effect of application temperatures was evaluated, −10 °C temperature caused a decrease in root length compared to the control group. When the effects of −10 °C temperature application on root length were evaluated, the longest root was found in ‘Red Globe’ (3.8 cm) cultivar and the shortest root was found in ‘Tekirdağ Çekirdeksiz’ (0.01 cm) cultivar. The longest root length in cuttings applied at −10 °C was measured in ‘Red Globe’ (3.8 cm) and ‘Alphonse Lavallee’ (3.1 cm) cultivars. While the longest root length at −15 °C was in the ‘Ugni Blanc’ (2.1 cm), measurable rooting could not be achieved in the ‘Çavuş’, ‘Tekirdağ Seedless’, ‘Early Sweet’ and ‘Michele Palieri’ cuttings (Table 8).
The root numbers and root development level of the cuttings removed from the perlite environment after freeze applications was determined using the 0–4 scale (Table 9). When the results were evaluated, the cultivar with the highest root scale value at −10 °C temperature was ‘Ugni Blanc’ (3.0), while the lowest root scale value was found in ‘Tekirdağ Seedless’, ‘Early Sweet’, and ‘Çavuş’ (0.3) cuttings. When the effect of −15 °C temperature application on the root scale was evaluated, the highest ratio was found in ‘Ugni Blanc’ (2.5), ‘Cardinal’ (1.0) and ‘Karaerik’ (0.8). Since rooting did not occur in the ‘Çavuş’, ‘Royal’, ‘Tekirdağ Seedless’, ‘Early Sweet’ and ‘Michele Palieri’, the root scale could not be recorded.
Among the grape cultivars examined in the study, the highest root numbers at both cold temperatures was determined in ‘Ugni Blanc’ cultivar. In terms of the average root number, ‘Karaerik’, ‘Red Globe’, ‘Cardinal’ and ‘Alphonse Lavalle’ cuttings had one or more roots, while almost no roots were formed in ‘Çavuş’, ‘Royal’, ‘Tekirdağ Seedless’, ‘Early Sweet’ and ‘Michele Palieri’ cuttings (Table 9).
Means followed by similar letters are not statistically different (P ≤ 0.05) as compared by Duncan’s multiple range test.
When evaluated according to the bud sprouting and rooting performance of single-bud cuttings kept in the growth chamber for a month; ‘Ugni Blanc’ and ‘Karaerik’ cultivars were evaluated to be more cold hardy than other tested grapes (Table 10).
Discussion
Cold damage to buds may vary from year to year due to genetic factors, the effect of cultural practices, and the severity and duration of freezing (Fennell 2004; Rende et al. 2018). Browning of the bud structure after a frost is a good indicator that the buds and tissues have died (Kaya 2020). Although it varies according to different climatic characteristics, the winter buds on the vine generally sprout in the spring and the primary bud located in the central part of the buds, which has fully realized the morphological separation period, occurs. If the primary bud is affected by winter or spring frosts secondary buds come into play and sprouting takes place (Poni et al. 2022). On the other hand, there is a slight decrease in productivity in the secondary bud that replaces the primary bud (Matti et al. 1989). If both buds die, sprouting takes place from the tertiary bud (Pratt 1979).
In this study; ‘Ugni Blanc’, ‘Karaerik’, ‘Alphonse Lavallée’, ‘Narince’, and ‘Victoria’ showed more than 50% primary bud viability at −15 °C. Similarly, Küpe and Köse (2019) calculated that the mortality rate of ‘Karaerik’ cultivar in primary buds were as 68.24% by cross sectioning method when the temperature dropped under −13.6 °C in Erzincan province, Turkiye between 2013 and 2014 years. Işık (2002) reported that while the ‘Yalova İncisi’, ‘Trakya İlkeren’, ‘Hamburg Muscat’, ‘Royal’, ‘Libol’, ‘Kozak Siyahi’, ‘İtalia’ and ‘Grosvert’ cultivars were resistant to low temperatures in Tekirdağ province, Turkiye which were −10.5 °C in 2000 and −12.2 °C in 2001. It was determined that ‘Hafızali’, ‘Razaki’, ‘Kozak Beyazı’, ‘Çınarlı Karasi’, ‘Manda Gözü’, ‘Danam’, ‘Michele Palieri’ and 29/2 (Yalova) cultivars were found as susceptible. Kaya (2020), In the research year when the air temperature was measured as −19.8 degrees, the viability rate in primary buds of the ‘Cardinal’ grape was determined with 78% as cold hardy and should be consider in future breeding programs in regions under high risk of winter cold. Çelik et al. (2008), in their study to determine the freeze damages caused by the 2005–2006 winter season in Kalecik conditions, Turkiye that due to low temperatures between −12 and −22 °C, the bud damage rates changed as; in ‘Narince’ (15%), ‘Gamay’ (16%), ‘Muscat Hamburg’ (19%), ‘Kalecik Karası’, ‘Emir’ (25%) and ‘Trakya İlkeren’ (39%) cultivars.
Topçu et al. (2013) in their study where they examined the degree of damage in the winter buds of the ‘Narince’ grape cultivar in 2012, when the minimum temperature was measured as −17.5 °C; the bud sporuting rate in the buds was found to be between 63% and 93%. As a matter of fact, in our research, the sprouting rate at −15 °C was determined to be 66.7% in the ‘Narince’ cultivar. This result is consistent with the findings of Topcu et al. (2013).
Sivritepe et al. (2001) ‘Atasarı’, ‘Uslu’ and ‘Yalova İncisi’ grape cultivars have determined the degrees of freeze resistance of the cultivars with the application of artificial freeze. As a result, it was reported that although primary, secondary, and tertiary bud viability showed a significant decrease in −15 °C application compared to the control, it showed viability rates above 50%, on the other hand, complete death occurred in all cultivars at −20 °C. The results obtained from the our research are similar to the results of that researchers at −15 °C.
Secondary buds will be carry cluster after emerge, but tertiary buds are usually unproductive, and even if sprouting occurs, clusters do not occur (Pratt 1979). In a study where the effect of different temperature applications on bud damage was investigated, it was found that the viability of tertiary buds decreases with a decrease in temperature (Sivritepe et al. 2001; Doğan et al. 2018). Different studies have reported that secondary buds may vary in terms of yield based on cultivars and years (Aydin 2001). Küpe and Köse (2019) was found that in ‘Karaerik’ cultivar, while secondary bud viability rates was as 50.7%, thertiary bud viability rates was as 65.7 according to cros sectioning method. In a study, it was reported that the application of −18 °C temperature for 24 h caused 100% death in the secondary buds of the ‘Antep Karasi’ grape cultivar (Doğan et al. 2018).
In our conducted study, in the ‘Ugni Blanc’ cultivar, secondary bud visbility rate was 70% of at −15 degrees. Apart from ‘Ugni Blanc’; ‘Alphonse Lavallee’ and (53.3%), ‘Karaerik’ (48.3%) cultivars were the highest secondary buds viability rate. Köse et al. (2014); during late spring frosts in Samsun, the shoots emerged from secondary buds was found to be lowest in the ‘Alphonse Lavalle’ (12.4%) and highest in the ‘Çavuş’ (61.0%). These results obtained by the researchers showed similar with our results. In our study, a serious decrease in the bud viability of the ‘Çavuş’ and ‘Early Sweet’ cultivars were observed in −15 °C application. The reason why the ‘Çavuş’ and ‘Early Sweet’ encounters such a high decline may be that the riding rate from the primary bud is 90.0% and the riding rate from the primary bud is 0.0%. As a result of damage to the primary buds under natural conditions, it can be explained that this cultivars started deacclimation stage before than the other tested cultivars. In our findings in this study shows paralel results in terms of ‘Michele Palieri’ (Işık 2002), ‘Narince’ (Çelik et al. 2008; Topçu et al. 2013), and ‘Cardinal’ (Kaya 2020), ‘Karaerik’ (Küpe and Köse 2019). In this way; Eriş et al. (1997) reported that samples taken in March, can survive at rates as low as 5–10% at −20 °C. Reserarcher determined that when the ‘Çavuş’ and ‘Kalecik Karası’ cultivars were exposed to cold temperatures −20 °C, those taken in January and February had higher frost resistance than those taken in November and March.
Sivritepe et al. (2005), in their study to determine the cold resistance of ‘Ergin Çekirdeksizi’, ‘Yalova Beyazı’ and ‘Yalova Misketi’ grape cultivars, found that only ‘Yalova Beyazı’, among these cultivars, could tolerate a 24-hour −15 °C freeze application during the dormant period.
In a study where the effects of cold temperatures on vine development were examined in ‘Ata Sarısı’, ‘Yalova İncisi’, and ‘Uslu’ cultivars, a decrease in the number of roots occurred with a decrease in temperatures and they determined the maximum number of roots in −15 °C at ‘Uslu’ (8.98) cultivar (Sivritepe et al. 2001). In our conducted study, phloem damage was measured minimum in ‘Alphonse Lavallee’ and ‘Karaerik’ cultivars compare the others. Similarly, the highest callus rate occurring in the basal part of the cuttings, which is an indicator of the phloem tissue viability, was determined in the ‘Alphonse Lavalle’. On the other hand, when root scale and phloem viability rate were correlated, the highest root scale was determined in ‘Ugni Blanc’ cultivar. It has been determined that the application of −20 °C temperature on the number of roots has a lethal effect (Sivritepe et al. 2001). The results of our study obtained showed parallel results with the other studies. While the control group received the highest root count values, low application temperatures showed a negative effect on the number of roots. In line with the results obtained; the ‘basal callus formation’ of the cuttings showed an inverse ratio with parameters such as rooting rate, number of roots, and root length. As the callus rate increases in the basal, there is a decrease in the rooting rate. On the other hand, seen callus development at the basal side, shows the phloem tissues are alive. In this subject, Fennell (2004) stated that the cold hardiness degrees of phloem tissues can differ between −15 °C and −32 °C among cultivars. In this study, ‘Alphonse Lavallee’, ‘Karaerik’ and ‘Ugni Blanch’ cultivars showed the highest phloem viability rate when the growing room results evaluated.
Plants that respond to low-temperature stress increase their cold tolerance ability by initiating a series of signal transduction reactions, reducing tissue water content, accumulating osmoregulatory substances, and changing antioxidant enzyme activity levels (Chinnusamy et al. 2007). The amount of water contained in plant buds is connected with frost resistance. Water in plant tissues is found as free or bound water. It is thought that the bound water content is higher in plants with good cold resistance compared to sensitive plants (Burke et al. 1976). It has been shown in many studies that frost resistance decreases with an increase in the amount of water in the buds (Rapacz 2002; Webster and Ebdon 2005). Although Kaya (2020) stated a positive correlation between cold hardiness and water content of tissues in grapevine buds; In contrast, Hamman et al. (1990) reported that tissue water content was not strongly correlated with changes in dormant bud cold hardiness in grape cultivars. Our obtained data shows parallels results with previous studies. While the ‘Early Sweet’ cultivar which was found to very sensitive as a result of tests, had the highest water content, the ‘Ugni Blanc’ which showed hardy are seen to have the lowest water content.
According to growing room performance of the cuttings which were cold applied, ‘Ugni Blanc’ and ‘Karaerik’ cultivars were evaluated to be more cold hardy grapes. A similar study was conducted and reported by Wang et al. (2022) that the LT50 values of the some different grape cultivars were between −12.5 and −21.7 °C, according to the bud sprouting method in the growing room condition. Researchers determined the LT50 value in the ‘Ugni Blanc’ as −17.56 °C according to bud germitation results in the growing room.
Conclusions
Due to the impact of global climate change, extreme weather events will be among the problems that we will face frequently in the near future in the world and in our country. In addition, the sudden falling temperatures in winter will negatively affect the viticultural regions and it is necessary of finding new alternative regions. It is necessary to identify cultivars adapted to low winter temperatures that have adapted to these regions. In this conducted research, according to both cross-sectioning and growing room performing results of the tested cultivars; ‘Ugni Blanc’, ‘Karaerik’, ‘Cardinal’ and ‘Alphonse Lavalle’ grape cultivars were determined as the showing highest cold hardiness performance at tested freeze temperetures. As a result of the study, these cultivars can be recommended to producers in winter cold-risky areas. It has also been determined that these cultivars can be used in breeding studies to be carried out in order to develop new cold-hardy grapevines in the future.
References
Alberdi M, Corcuera LJ (1991) Cold acclimation in plants. Phytochemistry 30: 3177-3184.
Ambroise V, Legay S, Guerriero G, Hausman JF, Cuypers A, Sergeant K (2020) The roots of plant frost hardiness and tolerance. Plant Cell Physiol 61(1):3–20. https://doi.org/10.1093/pcp/pcz196
Antivilo FG, Paz RC, Echeverria M, Keller M, Tognetti J, Borgo R, Juñent FR (2018) Thermal history parameters drive changes in physiology and cold hardiness of young grapevine plants during winter. Agric For Meteorol 262:227–236. https://doi.org/10.1016/j.agrformet.2018.07.017
Antivilo FG, Paz RC, Tognetti J, Keller M, Cavagnaro M, Barrio EE, Roig Juñent F (2020) Winter injury to grapevine secondary phloem and cambium impairs budbreak, cambium activity, and yield formation. J Plant Growth Regul 39:1095–1106. https://doi.org/10.1007/s00344-019-10051-w
Aydın İ (2001) Determination of productivity levels of secondary shoots formed as a result of damage to primary shoots in some wine grape varieties grown in Ankara conditions (Ankara university, Institute of Science and Technology, Master’s Thesis, Ankara (in Turkish))
Babajamali A, Gholami M, Baninasab B (2022) Drought preconditioning improves freezing tolerance in drought-tolerant and-intolerant grape cultivars. Theor Exp Plant Physiol 34(3):395–407. https://doi.org/10.1007/s40626-022-00252-2
Bates TR, Dunst RM, Joy P (2002) Seasonal dry matter, starch, and nutrient distribution in ‘Concord’ grapevine roots. HortScience 37(2):313–316. https://doi.org/10.21273/HORTSCI.37.2.313
Benítez-Cabello A, Delgado AM, Quintas C (2023) Main challenges expected from the impact of climate change on microbial biodiversity of table olives: current status and trends. Foods 12(19):3712. https://doi.org/10.3390/foods12193712
Bernardo S, Dinis LT, Machado N, Moutinho-Pereira J (2018) Grapevine abiotic stress assessment and search for sustainable adaptation strategies in Mediterranean-like climates. A review. Agron Sustain Dev 38:1–20. https://doi.org/10.1007/s13593-018-0544-0
Burke MJ, Gusta LV, Quamme HA, Weiser CJ, Li PH (1976) Freezing and injury in plants. Ann Rev Plant Physiol 27(1):507–528. https://doi.org/10.1146/annurev.pp.27.060176.002451
Çelik H, Erdemir D, Değirmenci D (2008) Damages caused by 2005–2006 winter cold on grape cultivars grown in Kalecik (Ankara) conditions. In: Türkiye 5th National Horticulture Congress, 04–07 September, Erzurum. Volume II, pp 451–454 (in Turkish)
Chinnusamy V, Zhu J, Zhu JK (2007) Cold stress regulation of gene expression in plants. Trends Plant Sci 12(10):444–451. https://doi.org/10.5505/TurkHijyen.2012.40316
Dami IE, Ennahli S, Zhang Y (2012) Assessment of winter injury in grape cultivars and pruning strategies following a freezing stress event. Am J Enol Vitic 63(1):106–111. https://doi.org/10.5344/ajev.2011.11040
Doğan O, Kara Z, Yazar K, Sabir A (2018) Freezing tolerance of some grape cultivars and rootstocks. In: XXX International Horticultural Congress IHC2018: International Symposium on Viticulture: Primary Production and Processing 1276, pp 223–230 https://doi.org/10.17660/ActaHortic.2020.1276.32
Droulia F, Charalampopoulos I (2021) Future climate change impacts on European viticulture: A review on recent scientific advances. Atmosphere 12(4):495. https://doi.org/10.3390/atmos12040495
Eriş A, Sivritepe N, Sivritepe HÖ (1997) A review of studies on Stress physiology of some fruits and vegetables. Uludağ Agr Fac J (1997) 13: 203-214
Eriş, A. (1982). Research on determining the chiling requirements and cold hardinessof some grape varieties grown under Ankara conditions. Ankara University, Faculty of Agriculture Publications, (856). (in Turkish).
Ershadi A, Karimi R, Mahdei KN (2016) Freezing tolerance and its relationship with soluble carbohydrates, proline and water content in 12 grapevine cultivars. Acta Physiol Plant 38:2. https://doi.org/10.1007/s11738-015-2021-6
Fennell A (2004) Freezing tolerance and injury in grapevines. J Crop Improv 10(1-2):201–235. https://doi.org/10.1300/J411v10n01_09
Frank D, Reichstein M, Bahn M, Thonicke K, Frank D, Mahecha MD, Zscheischler J (2015) Effects of climate extremes on the terrestrial carbon cycle: concepts, processes and potential future impacts. Glob Chang Biol 21(8):2861–2880. https://doi.org/10.1111/gcb.12916
Hamman RA, Renquist AR, Hughes HG (1990) Pruning effects on cold hardiness and water content during deacclimation of Merlot bud and cane tissues. Am J Enol Vitic 41(3): 251-260. https://doi.org/10.5344/ajev.1990.41.3.251
Howell GS, Shaulis N (1980) Factors influencing within-vine variation in the cold resistance of cane and primary bud tissues. Am J Enol Vitic 31(2):158–161. https://doi.org/10.5344/ajev.1980.31.2.158
Işık H (2002) Research on low temperature sensitivity and post-damage productivity characteristics of some table grapes. In: Türkiye 5th Viticulture and Winemaking Symposium. 89–96. 5–9 October, Nevşehir (in Turkish)
Kalkan NN, Karadoğan B, Değirmenci Karataş D (2011) Investigation of the effects of severe winter cold in 2007–2008 on the ‘Karaerik’ grape cultivar, Urfa. In: Türkiye 6th National Horticultural Congress. 04–08 October 2011-Urfa, pp 163–167 (in Turkish)
Kalkan NN, Kaya Ö, Karadoğan B, Köse C (2017) Determination of cold damage and lipid peroxidation level on winter buds of ‘Karaerik’ (Vitis vinifera L.) grape cultivar with different stem heights. Alinteri J Agric Sci 32(1):11–17. https://doi.org/10.28955/alinterizbd.297798
Kara Z, Sabır A, ve Göksu N (2005) Levels of exposure to cold of primary and secondary buds in some table grape cultvars and the effects of wrapping with paper. In: VI. Turkish Viticulture Symposium, pp 19–23 (in Turkish)
Karataş H, Özdemir G, Filizay M, Değirmenci D (2008) Effects of 2006–2007 winter cold damage on grape varieties grown in Diyarbakır (central) conditions. In: National Viticulture-Winemaking Symposium and Exhibition, pp 441–447 (in Turkish)
Karimi R (2020) Cold hardiness evaluation of 20 commercial table grape (Vitis vinifera L.) cultivars. Int J Fruit Sci 20(3):433–450. https://doi.org/10.1080/15538362.2019.1651242
Kaya Ö (2020) Bud death and its relationship with lateral shoot, water content and soluble carbohydrates in four grapevine cultivars following winter cold. Erwerbs-Obstbau 62(Suppl 1):43–50. https://doi.org/10.1007/s10341-020-00495-w
Kaya Ö, Köse C (2018) Effects of cold damage on grapevine. Yyu J Agr Sci 28(2):241–253. https://doi.org/10.29133/yyutbd.405252
Köse C, Güleryüz M (2009) Frost damage in dormant buds of ‘Karaerik’ grapevine grown at Üzümlü province of Erzincan during the winter of 2007–2008. Ataturk Univ J Fac Agric 40(1):55–60
Köse B, Ateș S, Çelik H (2014) Determination of late spring frost affections on some grape varieties grown in Samsun. Turk J Agric Res 1:162–169. https://doi.org/10.19159/tutad.91587
Küpe M, Köse C (2019) Determination of cold damage in field and laboratory conditions in dormant buds of Karaerik grape cultivar. Atatürk Univ Agric Fac Journal 50(2): 115-121 (in Turkish). https://doi.org/10.17097/ataunizfd.522485
Lang GA (1987) Dormancy: a new universal terminology. HortScience 25:817–820. https://doi.org/10.21273/HORTSCI.22.5.817
Matti GB, Nieces FP, Pisani PL, Rinaldelli E, Vignozzi N (1989) Investigations on shoots aristing from secondary buds and their productive behavior of several tuscan grapevine varieties. ATTI Accademia Italiana Della Vite e del Vino
Mirás-Avalos JM, Araujo ES (2021) Optimization of vineyard water management: Challenges, strategies, and perspectives. Water 13(6):746. https://doi.org/10.3390/w13060746
Petitpierre B, Arnold C, Phelps LN, Guisan A (2023) A tale of three vines: current and future threats to wild Eurasian grapevine by vineyards and invasive rootstocks. Divers Distrib 29(12):1594–1608. https://doi.org/10.1111/ddi.13780
Poni S, Sabbatini P, Palliotti A (2022) Facing spring frost damage in grapevine: recent developments and the role of delayed winter pruning—a review. Am J Enol Vitic 73(4):211–226. https://doi.org/10.5344/ajev.2022.22011
Pratt C (1979) Shoot and bud development during the prebloom period of Vitis. Vitis‑J Grapevine Res 18(1):1–5
Rapacz M (2002) Cold-deacclimation of oilseed rape (brassica napus var. oleifera) in response to fluctuating temperatures and photoperiod. Ann Bot 89(5):543–549. https://doi.org/10.1093/aob/mcf090
Rende M, Köse C, Kaya Ö (2018) An assessment of the relation between cold-hardiness and biochemical contents of winter buds of grapevine cv.’Karaerik’in acclimation-hardening-deacclimation phases. Mitteilungen Klosterneuburg, Rebe und Wein, Obstbau und Früchteverwertung 68(2): 67-81.
Santos JA, Fraga H, Malheiro AC, Moutinho-Pereira J, Dinis LT, Correia C, Schultz HR (2020) A review of the potential climate change impacts and adaptation options for European viticulture. Appl Sci 10(9):3092. https://doi.org/10.3390/app10093092
Sivritepe N, Burak M, Yalçın T (2001) Determination of frost resistance in Ata Sarısı, Uslu, Yalova İncisi and Yalova Misketi grape cultivars. J Uludag Univ Agric Fac 15:25–38
Sivritepe N, Burak M, Yalçın T (2005) Determination of frost resistance in Ergin Çekirdeksizi, Yalova Beyazı and Yalova Misketi grape cultivars. In: Türkiye 6th Viticulture Symposium, 19–23 September, Tekirdağ, pp 14–21 (in Turkish)
Stergios BG, Howell GS (1977) Effect of site on cold acclimation and deacclimation of Concord grapevines. Am J Enol Vitic 28(1):43–48. https://doi.org/10.5344/ajev.1974.28.1.43
Topçu N, Sucu S, Cangi R, Yağcı A, Kılıç D (2013) Damages caused by the 2012 winter cold on the Narince grape variety grown in the Tokat region. 8th Selçuk. J Agric Food Sci 27:413–419
Walsh JE, Ballinger TJ, Euskirchen ES, Hanna E, Mard J, Overland JE, Vihma T (2020) Extreme weather and climate events in northern areas: A review. Earth-Sci Rev 209:103324. https://doi.org/10.1016/j.earscirev.2020.103324
Wan N, Yang B, Yin D, Ma T, Fang Y, Sun X (2023) Overwintering covered with soil or avoiding burial of wine grapes under cold stress: Chinese wine industry’s past and future, challenges and opportunities. Stress Bio 3(1):40. https://doi.org/10.1007/s44154-023-00119-9
Wang ZL, Wu D, Hui M, Wang Y, Han X, Cao X, Li H (2022) Screening of cold hardiness-related indexes and establishment of a comprehensive evaluation method for grapevines (V. vinifera). Front Plant Sci, 13: 1014330. https://doi.org/10.3389/fpls.2022.1014330
Webster DE, Ebdon JS (2005) Effects of nitrogen and potassium fertilization on perennial ryegrass cold tolerance during deacclimation in late winter and early spring. HortScience 40(3):842–849. https://doi.org/10.21273/HORTSCI.40.3.842
Wolf TK, Pool RM (1987) Factors affecting exotherm detection in the differential thermal analysis of grapevine dormant buds. J Am Soc Hortic Sci 112(3):520–525. https://doi.org/10.21273/JASHS.112.3.520
Wolfe W (2001) Vine and vineyard management following low temperature injury. In: Proceedings of the ASEV 50th Anniversary Annual Meeting, Seattle, Washington, June 19–23, 2000. Am Soc Enol Vitic ASEV, pp 101–110
Wolpert JA, Howell GS (1985). Cold acclimation of Concord grapevines. I. Variation in cold hardiness within the canopy. Am J Enol Vitic 36(3): 185-188.
Wolpert JA, Howell GS (1986) Cold acclimation of Concord grapevines. III. Relationship between cold hardiness, tissue water content, and maturation. Vitis 25:151–159. https://doi.org/10.5073/vitis.1986.25.151-159
Zabadal TJ, Dami IE, Goffinet MC, Martinson TE, Chien ML (2007) Winter injury to grapevines and methods of protection. Michigan State University Extension, Chien
Zapata C, Deléens E, Chaillou S, Magné C (2004) Partitioning and mobilization of starch and N reserves in grapevine (Vitis vinifera L.). J Plant Physiol 161(9):1031–1040. https://doi.org/10.1016/j.jplph.2003.11.009
Zhao Y, Wang ZX, Yang YM, Liu HS, Shi GL, Ai J (2020) Analysis of the cold tolerance and physiological response differences of amur grape (Vitis amurensis) germplasms during overwintering. Sci Hortic 259:108760. https://doi.org/10.1016/j.scienta.2019.108760
Acknowledgements
This article is produce from the Ms.D thesis submitted by Yahya Uray at Ondokuz Mayıs University’s Faculty of Agriculture and Department of Horticulture.
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of interest
Y. Uray and B. Köse declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Uray, Y., Köse, B. Performing of the Winter Buds and Phloem Tissue in Grape Cultivars After Cold Applications. Applied Fruit Science 66, 997–1008 (2024). https://doi.org/10.1007/s10341-024-01066-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10341-024-01066-z