
Abstract
Understorey habitats are optimal ecological structures for natural enemy enhancement in fruit orchards. A large-scale experiment was carried out to establish the effect of resident vegetation cover (VC) on green lacewings as compared to bare soil, the dominant soil management strategy used in Spanish olive orchards. Lacewings were sampled using baited McPhail traps for adults, and suction was used to collect adults and larvae from olive canopies. Additionally, we monitored the presence of the lacewing’s main target pest, olive moth eggs, as well as VC composition and density. McPhail trapping showed higher Chrysopidae abundances in VC plots during two consecutive years even though flowering plants represented 29.7 % of the total. Multivariate analysis identified Chrysoperla carnea s.l. and Pseudomallada prasinus as contributing to differences in abundance. VC slightly increased capture diversity; however, no specific link between any Chrysopidae species and VC was detected. No differences were observed in individuals collected through suction in 2009, which could be attributed to low sampling efficiency. In 2010, when sampling was increased considerably, higher adult and larval abundances were recorded in VC only with respect to C. carnea s.l. A delay was detected between McPhail captures and suction collection peaks. The fact that VC promoted higher abundance detected earlier through trapping, and later on olive canopies through suction, coinciding with P. oleae presence, suggests that resident VC may contribute to a build-up of green lacewing populations moving onto the crop at the time of the pest attack.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Key message
-
Green lacewings are important predators in olive insect communities contributing to the natural suppression of the olive moth.
-
We evaluated the influence of managed resident vegetation cover (VC) on the adult and larval populations of lacewings vs. the widely extended practice of bare soil maintenance.
-
Adult abundance and diversity increased under the influence of VC. Higher abundances translated into higher presence of larvae on trees during the olive moth egg-laying period indicating a potential of VC for pest control.
Introduction
Habitat management of agricultural ecosystems is a set of strategies that can be aimed at increasing the abundance and fitness of native natural enemies by delivering resources at the optimal temporal and spatial scales (Landis et al. 2000; New 2005). Of the different options available, within-field habitats have been demonstrated to play an important role in predator and parasitoid enhancement on agricultural sites (Bianchi et al. 2006; New 2005; Tscharntke et al. 2005). Non-crop vegetation provides refuge, alimentary resources, and alternative prey and also facilitates contact between natural enemies and their target pests. Natural enemies can, in turn, increase pressure on insect pest populations, thus reducing the pesticide input required for their control (Thomson and Hoffmann 2009).
Arboreal crops such as fruits are characterized by a multi-strata structure comprising an understorey habitat and tree canopies (Simon et al. 2010). The understorey stratum, given its proximity to the targeted crop, has considerable potential for habitat manipulation through the introduction of vegetation cover. The cover may be established by allowing wildflowers and grasses to grow or by sowing selected plant species. Its positive contribution to increasing the abundance and diversity of natural enemies on crop canopies has been observed in many studies (Aguilar-Fenollosa and Jacas 2013; Danne et al. 2010; Silva et al. 2010; Thomson and Hoffmann 2009), and this habitat management practice has been seen as a promising option for IPM optimization in fruit orchards (Rieux et al. 1999). However, the effects of understory habitat management are complex and depend on a number of factors such as crop type and vegetation cover composition. Some studies have found that understory habitat management does not affect natural enemy abundance in crops (Bone et al. 2009; Bugg et al. 1991) and even observed some negative effects in relation to pest suppression (Simon et al. 2010).
Green lacewings (Neuroptera: Chrysopidae) are predators of numerous soft-bodied insects, mites, and eggs that are known to play a biological control role by consuming insect pests in almost all agricultural ecosystems (Duelli 2001; Stelzl and Devetak 1999). Chrysopidae have a complex life history. While all larvae are predaceous, adults of most species rely on vegetal substrates for feeding and are believed to be attracted to large patches of flowering plants (Villenave et al. 2006). Although vegetation cover may therefore potentially contribute to increasing this group of predators, few studies have confirmed their positive impact on lacewing densities in the perennial crop canopy (Daane 2001). In southern Spanish olive orchards, green lacewings, mainly those of the carnea group, are known to play a predominant role in the predation of eggs of the carpophagous generation of the olive moth Prays oleae (Bernard) (Campos 2001; Szentkirályi 2001b). Early predatory consumption of these eggs prevents the pest from causing fruit fall, which is the most important effect attributed to this key pest in olive cropping. Some years, predatory efficiency may result in a drastic decrease in the high rate of olive moth attacks on olive fruits (Ramos and Ramos 1990).
Nowadays, the predominant understorey management strategy in Spanish olive orchards consists of maintaining a bare, weed-free soil all year round by using a combination of herbicides and minimum tillage or by using herbicides alone (ESYRCE 2014). Nutrient and soil loss caused by this type of soil management is progressively driving farmers, encouraged by local authorities, to introduce vegetation cover, which is known to be highly effective in preventing severe erosion problems in olive orchards (Gómez et al. 2009). To date, 29.0 % of the total area occupied by olive orchards in Spain has been managed using vegetation cover in the form of resident weedy plants (ESYRCE 2014). In this context, the objective of our study was to determine the impact of this soil management practice on the abundance and diversity of green lacewings and their availability for the biological control of P. oleae.
Materials and methods
Study site
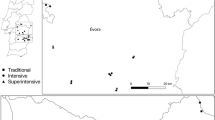
The experiment was conducted in a 235 ha commercial olive extension (37°17′46.7″N, 3°46′28.7″W) situated in a large, homogeneous, intensively farmed olive-growing area 20 km northwest of the city of Granada in southern Spain. The climate is typically Mediterranean, with a local annual mean temperature of 16.2 °C and an average annual rainfall of 438.3 mm (average of data for 2001–2010). The whole extension, managed according to integrated growing regulations for fertilization, soil management, weed control, and plant protection, was planted with 90-year-old trees exclusively of the ‘Picual’ cultivar, grown 10 × 10 m apart (at a density of 110–130 trees per ha) without irrigation. The trees were pruned every other year. The surrounding landscape consisted of olive plantations of a similar structure except for a field of commercial wheat located near a stream on the eastern edge at a minimum distance of 615 m from the experimental plots. The deep, well-drained, Calcaric Calcisol soil is, according to the IUSS Working Group WRB (WRB 2006), typical of olive orchards in southern Spain.
Soil management involved maintaining weed cover situated in the alleys between tree lines growing in 1.5-m-wide strips perpendicular to the slope direction. No mechanical or herbicidal control was applied to this cover, which was allowed to progressively senesce during the month of June. The strips of bare soil (BS) under the canopies were maintained by means of 4 L ha−1 oxyfluorfen (24 %) chemical herbicide in a 400 L ha−1 solution for preemergence treatment in late winter, and glyphosate (36 %) whenever further patch postemergence applications were required. Additionally, chopped pruning residues were used to create inert intercrop cover strips, running along the direction of the slope and perpendicular to the weed cover strips. Thus, no weeds were present on the inert cover strips. The experimental area had been under the same soil management system for over 10 years. During the study, no fertilizers, insecticides, or other pest control methods were employed.
Two different soil treatments were compared during the experiment: (1) maintenance of long-established resident cover during the usual period, hereafter referred to as vegetation cover (VC), and (2) a treatment without vegetation cover between the rows of trees, hereafter referred to as bare soil (BS). During the years 2009 and 2010, the VC zones were managed as usual. The BS zones were chemically mowed, as described previously, to remove weeds from the entire soil surface (both under the tree canopy and between the rows of trees). BS treatment covered an area of 54.4 ha divided into a northern and a southern zone by a gravel road alongside a gully running in an east–west direction.
Chrysopidae
Lacewing adults in both VC and BS were monitored using McPhail traps, which are widely employed for lacewing sampling in olive groves (Corrales and Campos 2004; Szentkirályi 2001b). The traps were baited with an aqueous solution of 5 % diammonium phosphate as an attractant plus 2 % Borax® for insect conservation and were suspended facing north at a height of between 1.5 and 2.0 m on the inner side of the trees. Forty McPhail traps were displayed in plots, each containing five traps. Traps within each plot were placed at least 20 m apart. Plots were replicated up to four times (20 traps) per treatment and zone and were always placed on north-facing slopes and positioned at least 150 m apart within the same zone and 750 m apart between the northern and southern zones. Captures were examined biweekly from late March to September in 2009 and 2010, and their contents were filtered using a nylon mesh to obtain the adult lacewings.
In addition, canopy suction samples were obtained using an insect aspirator (Modified CDC Backpack Aspirator Model 1412, John W. Hock Co., Gainsville, FL, USA) to determine larval and adult abundance associated with olive trees. Square plots consisting of 16 trees (4 × 4) were sampled according to the method described by Porcel et al. (2011). In 2009, three plots were sampled per treatment in May, June, and September, two in the northern zone and one in the southern zone (a total of 96 trees). Suction plots were placed at least 100 m apart within each zone and 500 m apart in the area between the zones and were always north facing. Due to an inadequate level of lacewing captures obtained in 2009, in 2010 the number of suction samples was increased to six plots per treatment and per zone (192 trees) and were obtained every 10 days between March and September. On arrival from the field, all samples were frozen for insect conservation purposes. In the laboratory, the lacewing larvae and adult in the suction samples were separated from other insects and vegetal material. The adults captured in McPhail traps were also separated from the rest of the insects. All the individuals were refrozen for later identification under a stereomicroscope (Stemi SV8; Zeiss, Oberkochen, Germany) following instructions provided by the taxonomic specialist Dr. Víctor Monserrat (Complutense University, Madrid). Adults were identified up to species level and larvae to genus level. Dr. Víctor Monserrat also clarified issues regarding the identification of uncommon species.
Prey presence
The presence of P. oleae eggs laid on the olive fruits was monitored biweekly on three consecutive sampling occasions; the first sampling was carried out when the fruit was formed and was receptive to P. oleae oviposition. Two olive fruits were selected from a random selection of 10 olive twigs (20 olives per tree). Plots, consisting of five trees (100 fruits) and replicated four times per treatment and per zone (400 fruits), were at least 20 m apart within each zone and 750 m apart in the area between the northern and southern zones. The eggs were counted under a stereoscope and expressed in eggs per tree.
Vegetation cover
The plant density and species composition of VC were estimated by measuring 20 m linear transects along the VC strips. Measurements were carried out annually in 2009 and 2010 at peak standing VC in early June. In 2009, 15 transects were conducted in the southern zone and 24 in the northern zone. In 2010, 18 transects were conducted in the southern zone and 13 in the northern zone. The point-intercept method was used for each transect by driving 20 sharpened pins (1 m long, 5 mm ϕ) into the ground 1 m apart, making sure that the pin protruded above the vegetation. At each position, the number of times the pin touched each plant species was recorded. VC density was calculated as the number of total plant contacts divided by the total number of points measured.
Statistical analysis
Total abundance and species diversity of Chrysopidae adults captured in McPhail traps were analyzed using a generalized additive mixed model (GAMM). Additive models are non-parametric extended versions of linear models that can process a mix of parametric and non-parametric variables (Yee and Mitchell 1991) allowing for non-linear relationships between the response variable and all or some of the explanatory variables. The relationship between these variables is thus modeled using non-parametric smoothing curves (Zuur et al. 2009). Additive modeling is a useful tool for modeling abundance trends as a non-linear function of time (Fewster et al. 2000). We used the GAMM to model the temporal variability of abundance and diversity. Smooth functions were constructed using a penalized regression spline approach, and the amount of smoothing was automatically estimated by cross-validation (Wood 2006). During data exploration (Zuur et al. 2010), the autocorrelation function (ACF) plots for regularly spaced time series revealed the existence of serial dependence of observations over time. To account for this temporal dependence, we took advantage of the GAMM’s ability to deal with fixed and random effects and added an auto-regressive model of order 1, AR(1), to the GAMM as a random effect to allow for the temporal correlation. Akaike’s Information Criteria (AIC) were used to verify whether the inclusion of the dependence structure improved the models by comparing their fit with and without the correlation structure. AIC provide an effective technique for selecting the best approximating model for data analysis (Burnham and Anderson 2002). Two GAMMs were constructed with abundance and diversity as response variables. Species diversity was calculated using Shannon’s index for captures per McPhail trap and sampling date. This index takes into account species richness and evenness. In relation to the design of the experiment, to achieve the highest level of homogeneity, we selected an area which, though quite large, operates under similar agronomical and environmental conditions. However, during the preliminary data analysis stage, we noticed spatial heterogeneity between data from the plots in the northern and southern zones. “Zone” was therefore included as a variable in the models in order to improve fit. Thus, both models used treatment (VC–BS), zone (north–south), and year (2009–2010) as categorical variables and sampling date as a smooth function. To construct the models, we used the time series data from the first sampling date in late March until VC was completely dry in the month of July. Chrysopidae abundance count data were modeled using a Poisson distribution and the log link function. Overdispersion was corrected using a quasi-poisson model. A Gaussian distribution was selected for the species diversity GAMM using an identity link function. Homoscedasticity and normality assumptions were verified using graphical representations of the model residuals versus fitted values and the residuals histogram, respectively (Zuur et al. 2007).
Principal response curves (PRCs) were used after univariate analysis in order to study the temporal effect of VC on the Chrysopidae community and to identify the individual contributions of each species to differences between treatments. PRC is a multivariate technique for assessing species communities and is an appropriate tool for studying the effect of treatments referenced to a control over time. PRCs are based on partial Redundancy Analysis (pRDA), a more restricted version of Principal Component Analysis (PCA) (Van den Brink and Braak 1999). This type of analysis only extracts information from the variance defined by the explanatory variable (VC–BS) and time variable (sampling date) tested and focuses on the difference between the species composition of the treatment and the control at the corresponding time (Moser et al. 2007). The analysis is presented graphically in a diagram that plots the canonical coefficient (cdt) relative to the control on the y axis against time on the x axis. The same graph plots species weight (b k), representing the importance of each species for the response given in the diagram. High b k values indicate that the response of the species is likely to follow the pattern in the PRC diagram; low b k values of between −0.5 and 0.5 indicate a weak response, while high negative values show a reverse trend from that in the PRC diagram. Separate PRC analyses were carried out for each year (2009 and 2010) and covered the whole sampling period (March to September). Monte Carlo permutation tests were conducted to test the significance of the y axis.
For suction samples, each statistical sampling unit consisted of the sum of all Chrysopidae collected from the square plots containing 16 trees. Adult suction data from 2009 were analyzed using a generalized linear model (GLM) with a negative binomial distribution error due to overdispersion and the log link function. Treatment and the two sampling dates were tested as categorical variables. Zone could not be included as no replications were carried out in the southern zone. The collection of larvae in 2009 was insufficient to construct an adequate model and was therefore not tested statistically. 2010 suction data for adult and larval abundance were analyzed using GAMMs as described for McPhail trap captures. The AR-1 model was added as temporal dependence was also detected. Biodiversity was not analyzed due to the limited number of the different species collected through suction during VC presence. Adult and larva changes in the community structure were analyzed using PRC multivariate analysis as described above.
The number of eggs deposited by P. oleae on olive fruits was analyzed using a generalized linear mixed model (GLMM) with a log link and a negative binomial error distribution due to overdispersion. The model used treatment, zone, and year as categorical variables and sampling date modeled as a random effect. VC density was compared between zones and years using two-way analysis of variance (ANOVA).
All the statistical analyses were performed with R software 2.1.2 using packages mgcv, glmmadmb, and vegan.
Results
McPhail traps
A total of 3461 adult Chrysopidae were captured in McPhail traps during the whole sampling period. Nine species from five genera were captured in 2009 and 12 species from six genera in 2010 (Online Resource 2). In both years, Chrysoperla carnea s.l. and Pseudomallada prasinus (Burmeister) were the most abundant species, followed by Pseudomallada flavifrons (Brauer) and Rexa lordina Navàs. The other species accounted for 4.1 % of the total captures in 2009 and 1.8 % in 2010, including the exceptionally rare species Suarius walshingami Navàs (Canard et al. 2007). During the presence of VC, captures totaled 2124 individuals and were significantly more abundant in VC plots, with 3.50 ± 0.25 (mean ± SE) compared to BS plots, with 2.36 ± 0.18 (Table 1). Capture rates in VC plots were higher on almost all the sampling dates (Fig. 1); the differences were more accentuated in late May of 2009 and early June of 2010, coinciding with the modeled flight peaks. The number of captures was also significantly lower in plots situated in the northern zone (2.03 ± 0.15) than in those in the southern zone (3.83 ± 0.26). No difference in abundance was observed between years, as the year variable was not significant in the model.

Mean Chrysopidae adults captured per McPhail trap (±SE) in VC and BS plots during the presence of the VC (line plot) and mean number of P. oleae eggs per 20 fruits collected per tree in VC and BS plots (horizontal bar chart) in the years a 2009 and b 2010
The Shannon index was also significantly higher in VC plots than in BS plots (Table 1), with values of 0.48 ± 0.03 and 0.40 ± 0.03 (mean ± SE), respectively. Capture diversity was higher on almost all the sampling dates in 2009 (Fig. 2a) and during the month of June in 2010 (Fig. 2b). Chrysopidae diversity peaks were detected in May 2009 and June 2010, which roughly coincide with maximum abundance levels in both these years (Fig. 1). Zone and year were not found to influence diversity (GAMM, P > 0.05).

Shannon diversity index per McPhail trap (mean ± SE) in VC and BS plots during the presence of the VC in a 2009 and b 2010
PRC analysis of McPhail captures revealed species composition deviance in the VC treatment as compared with BS showing a similar seasonal pattern in both 2009 and 2010 (Fig. 3). Important positive deviations were observed during the presence of VC in May and June. From the end of June onwards, low or negative deviances were observed until the month of September. The species C. carnea s.l. and P. prasinus were the major contributors to the differences detected in the years 2009 and 2010 (Fig. 3). The species P. flavifrons and Pseudomallada picteti (McLachlan), captured in relatively high numbers during VC presence, were not observed to contribute to the variations caused by the treatment. Such was the case for R. lordina, captured only in May and early June, which was also abundant in BS plots. Pseudomallada subcubitalis (McLachlan), Cunctochrysa baetica (Hölzel), Chrysopa viridiana Schneider, Chrysopa formosa Brauer, Chrysoperla mediterranea (Hölzel), and S. walshingami were observed almost exclusively from early July until the end of the sampling period (mainly outside the VC season) and therefore did not affect the deviances obtained. Small numbers of the species Pseudomallada venosus (Rambur) and Pseudomallada genei (Rambur) were captured (Online Resource 2). All the species listed were captured on at least one occasion under both VC and BS conditions.

PRC diagrams showing the effect of the treatment (VC) relative to the control (BS) on Chrysopidae species composition of McPhail captures per trap throughout the whole sampling period in the years a 2009 and b 2010. The BS control is represented by zero deviance on the y axis. The numerical scale on the left y axis applies to both the canonical coefficients (c dt), represented in the line plot and the species weight (b k) indicated for each species in the right y axis. The x axis of both models was significant (Monte Carlo permutation test, P < 0.01)
Suction samples
In 2009, 67 individual adult Chrysopidae were obtained by suction on the two dates sampled. A mean ± SE of 5.66 ± 1.33 individuals was obtained in VC and 5.50 ± 2.71 individuals in BS plots, with no difference being observed between the treatments (GLM, t = 0.03, P = 0.975) or between sampling dates (GLM, t = 1.10, P = 0.302). A total of 16 larvae were recorded in 2009, distributed evenly between VC and BS plots (eight individuals from each). Only three larvae were observed in May, which increased to 13 in June. A statistical comparison of the data was not carried out due to the small number of individuals observed. All the adults collected in 2009 belonged to the species C. carnea s.l. and the larvae to the genus Chrysoperla.
In 2010, due to the increased sampling effort described above, a total of 527 adults from five species and five genera were captured between March and September (Online Resource 2). As was also the case for McPhail traps, C. carnea s.l. and P. prasinus were the predominant species collected by suction, although a smaller number of P. prasinus were obtained as compared to trap captures. The number of larvae sampled also increased sharply compared to 2009, with 513 individuals being recorded. Two genera, Chrysoperla and Pseudomallada, accounted for 49.9 and 50.1 % of total larvae, respectively. A total of 236 adults and 176 larvae were collected during the VC presence period. Adults were sampled from olive trees in considerable numbers during June (Fig. 4a) and peaked in mid-June, which is later than the capture maximum registered for trapping (Fig. 1). Adult abundance was significantly higher in VC plots (Table 1) which averaged 2.47 ± 0.73 (mean ± SE) compared to 1.11 ± 0.36 in BS plots. VC plots showed higher abundance values on all the sampling dates during June (Fig. 4a). Significantly fewer adults were obtained in the northern zone (0.78 ± 0.17) as compared to the southern zone (2.78 ± 0.34) (Table 1). Larvae displayed a bimodal pattern and peaked twice (in mid-April and mid-June) (Fig. 4b). The highest abundance registered in June coincided with the presence of adults in olive canopies (Fig. 4a). As with the adults, larval abundance in VC plots (1.68 ± 0.32, mean ± SE) was significantly higher than in BS plots (0.98 ± 0.22) during the VC presence period (Table 1). No zone effect was detected for larval abundance despite the lower values observed in the northern zone (1.21 ± 0.23) as compared to the southern zone (1.45 ± 0.32) (GAMM, t = 1.36, P = 0.175).

Chrysopidae a adults and b larvae collected per plot through suction sampling in VC and BS plots during the presence of the VC (line plot) and mean number of P. oleae eggs per 20 fruits collected per tree in VC and BS plots (horizontal bar chart) in the year 2010
PRC analysis of adults showed positive deviations from the BS treatment during June (Fig. 5a) when the highest abundance rates and greatest differences in total adult abundance between treatments were registered (Fig. 4a). Small or negative deviations were obtained in July, August, and September. C. carnea s.l. was the only species to contribute to the deviations observed, as only 10 P. prasinus individuals (4.23 % of the total) were collected in the olive canopy during VC presence. The other species appeared between July and the end of the sampling period. PRC analysis of larvae showed sustained positive deviations from the beginning of June until the end of July, beyond the VC presence period (Fig. 5b). Again, only larvae of the genus Chrysoperla contributed to the difference observed as they were much more abundant than Pseudomallada larvae (22.1 %) from March to late June (Online Resource 2).

PRC diagrams showing the effect of the treatment (VC) relative to the control (BS) on Chrysopidae a adult and b larval species composition collected per plot through suction sampling throughout the sampling period in the year 2010. The BS control is represented by zero deviance on the y axis. The numerical scale on the left y axis applies to both the canonical coefficients (c dt), represented in the line plot, and the species weight (b k) indicated for each species in the right y axis. The x axis of both models was significant (Monte Carlo permutation test, P < 0.01)
Prey
As olive fruit becomes receptive to P. oleae antophagous generation during mid-June, in both 2009 and 2010, P. oleae egg samples were taken between mid-June and mid-July on three occasions (Fig. 1). The number of eggs recorded on the two sampling dates in June 2009 was similar. However, no eggs were present in July (Fig. 1a). In 2010, as in the previous year, eggs were present during the second half of June, although, unlike in 2009, oviposition, though declining, continued into mid-July (Fig. 1b). The number of olive moth eggs was higher in VC plots in 2009 (Fig. 1a), which was not the case in 2010 (Fig. 1b). Overall, no differences were detected (GLMM, z = 1.13, P = 0.258) possibly due to the large deviance observed. The P. oleae attack rate was higher in 2010 compared to 2009 as year was the only significant variable in the model (GLMM, z = 2.64, P = 0.006). No difference was observed between zones (GLMM, z = 0.80, P = 0.426).
Chrysopid adults caught in McPhail traps started to build up and reached their maximum levels before the onset of olive moth oviposition in both 2009 and 2010 (Fig. 1). As for their presence in the olive canopy recorded through suction sampling in 2010, maximum adult abundance showed temporal correspondence with the onset and maximum presence of P. oleae eggs (Fig. 4a). The peak in the larvae population detected on olive trees in June also coincided with prey presence (Fig. 4b).
Vegetation cover
The Poaceae species Bromus madritensis L. Lolium rigidum Gaudin and Hordeum leporinum Link dominated the indigenous VC (Online Resource 3). Flowering plants represented 29.7 % of the total recorded, with Anacyclus clavatus Reichenb (Asteraceae) and Medicago minima (L.) (Fabaceae) being the most abundant. Total VC density reached values of 1.52 ± 0.11 (mean ± SE) in 2009 and 1.63 ± 0.09 in 2010, with no statistical difference being observed between years (ANOVA, F = 0.33, df = 2, P = 0.567). Also, no difference in VC density was observed between zones (ANOVA, F = 1.22, df = 2, P = 0.273), with values of 1.49 ± 0.09 recorded in the northern zone and 1.65 ± 0.12 in the southern zone.
Discussion
The presence of long-term established resident VC in the experimental olive-growing area resulted in a higher abundance of Chrysopidae adults measured through two different methodologies. This increased abundance of adults translated into a higher larval population in VC plots, coinciding with the olive moth egg-laying period, even though prey density did not differ between treatments. The importance of natural vegetation patches for lacewing concentrations and conservation has been reviewed by Szentkirályi (2001a), who identified valuable ecological structures such as hedgerows, flower strips, and ground cover. Despite the important role played by natural vegetation in green lacewing diversity and conservation, little evidence of the function of ground cover in increasing lacewing presence on the targeted crop has been found in orchards. Smith et al. (1996) registered 3–5 times higher Chrysoperla rufilabris (Burmeister) abundance in the month of July in pecan trees with legume cover as compared to those with grass cover. Similarly, in apple trees, Wyss (1995, 1996) recorded larger numbers of C. carnea s.l. individuals associated with weedy strips compared to rye grass cover for all three sampling methods employed. Although these findings are consistent with the results presented in our study, similar studies have not observed this impact of cover. In vineyards, neither grass nor saltbush VC influenced the presence of lacewings on vine canopies, even though positive results were observed for predators (Danne et al. 2010); in apple orchards, a mix of selected plants failed to increase green and brown lacewing abundance on tree canopies (Bone et al. 2009).
This discrepancy could be explained by various factors. In general, lacewing individuals were collected in small and even insignificant numbers as reported by Bone et al. (2009). Indeed, experiments aimed at studying the effects of ground cover and cover crops rarely consider Chrysopidae in isolation from other predators, meaning that sampling efforts and methods are not adapted to Chrysopidae detection requirements. In this regard, Silva et al. (2010), who compared the effect of two types of VC with a BS control in 2003, observed no significant differences between treatments using the beating method. However, using suction sampling (carried out simultaneously), over three times more lacewings were obtained, and important differences in the canopy between VC treatments and the control were observed. In our study, the small size of the sampled population could also explain why no difference in the number of adults collected by suction was observed in 2009 unlike the marked differences recorded in the same year by McPhail trapping.
The size of the experimental plots may also be an important factor. Prasifka et al. (2005) have observed effects of scale on lacewings and have suggested that abundance evaluation experiments carried out on small plots could not be regarded as equivalent to those conducted on a field-size scale. Most of the agriculturally important lacewing species are known to cover large distances due to their constant nomadism (Duelli 2001). Thus, small-scale plots may be affected by rapid lacewing colonization from the immediate surroundings. Certainly, lacewing abundance evaluation must therefore benefit from experiments designed for larger scales, which is consistent with the positive results reported by Smith et al. (1996) using 10 ha experimental plots and those described in our study (plots of over 20 ha).
Another factor that might explain the differences between the studies is the timing of VC senescence. Bone et al. (2009) reported that an increase in natural enemies in the cover crops did not necessarily influence numbers in the apple tree canopy. Non-crop plants may arrest egg-laying females if the prey presence period overlaps that of the crop (McEwen and Ruiz 1994; Szentkirályi 2001a). Interestingly, Smith et al. (1996) found that legume VC improved C. rufilabris presence on pecan trees just after VC senescence during the month of June. Our 2010 results also show that maximum differences between VC and BS treatments for adults and larvae collected directly from the olive canopies by suction occurred in mid-June, coinciding with the decline in VC. Furthermore, the temporal distribution of McPhail adult catches and those collected in suction samples did not coincide in 2010. Abundance levels registered through trapping started to build up and peaked earlier than presence on tree canopies. Traps, though hung on the inner side of the trees, may have attracted green lacewings at that time associated with VC. This would also be in line with the possible movement of Chrysopidae from VC to tree canopies during the VC senescence period, suggesting that the presence of VC may act as a reservoir that increases lacewing activity in the crop after VC disappears. The change in the vegetal substrate in olive groves may also be boosted by the presence of P. oleae eggs, one of the most abundant types of prey in olive trees, as indicated by the extraordinary temporal coincidence observed with adult abundance on trees.
Apart from the effect of VC, unexpected differences in adult abundance were recorded between zones, suggesting the existence of an important unmeasured factor in the experiment. The reason for this is unknown and cannot be explained by the VC heterogeneity or prey availability as shown in our results. The existence of a large forest stand, located about 1 km away and close to the southern zone, may be responsible for increasing Chrysopidae occupancy in this area.
Our results show that VC slightly increased Chrysopidae biodiversity but did not alter species assemblages. VC therefore increased species evenness by reducing C. carnea s.l. dominance in VC plots. It has been observed that lacewing species richness and diversity in agricultural fields are determined by the heterogeneity and quality of adjacent habitats (Szentkirályi 2001a), although, to date, no effect of understorey habitats has been reported. The limited impact of VC further confirms that green lacewing species assemblages in agricultural fields are related to environmental factors on a larger scale. In general, species richness was quite high in relation to that expected for olive groves (Szentkirályi 2001b) which can be attributed to the absence of insecticidal treatment that negatively impacts species richness and diversity (Porcel et al. 2013).
The most abundant species captured in McPhail traps were the most important cause of the differences observed between VC and BS. Both C. carnea s.l. and P. prasinus are field-crop specialists adapted to living in nonspecific patchy environments and can benefit from the presence of VC. R. lordina, which was only observed during the presence of VC, is, on the contrary, highly substrate specific in olive trees (Monserrat 2008), which explains why no impact on this species was observed. The minor contribution of P. flavifrons and P. picteti to the divergence between VC and BS can only be explained by the limited amount of captures compared to the highly similar species P. prasinus. To the best of our knowledge, this is the first time Suarius walshingami has been captured at an agricultural site. In Europe, this exceptionally rare species has hitherto been exclusively collected in southwestern Spain, in xeric environments with natural vegetation (Monserrat 2010). Its presence in this study is in accordance with its known area of distribution and could indicate that it may also thrive in more manipulated environments such as olive orchards.
Surprisingly, suction collected an almost insignificant number of P. prasinus adults on olive canopies during VC presence. This species was not responsible for any of the difference between VC and BS at either the adult or larval stage, which may be indicative of differing behavior patterns compared to C. carnea s.l. Pseudomallada spp. appeared on olive canopies later than C. carnea s.l., suggesting later displacement from VC. The temporal coincidence of C. carnea s.l. and P. oleae indicates a closer prey–predator relationship than that with Pseudomallada spp. Indeed, while C. carnea s.l. predation upon P. oleae has been confirmed by serological tests (Morris et al. 1999), there is no evidence on Pseudomallada spp.’s predatory capacity.
Green lacewing species recorded in olive orchards are mostly palynoglycophagous (except Chrysopa spp.), indicating that the increase in adult abundance is mainly due to the moderate amount of flowering species present in VC. A number of studies have reported that flowering species have a positive effect on Chrysopidae abundance in VC compared to grass cover (Smith et al. 1996; Song et al. 2010; Wyss 1996). Theoretically, Chrysopidae presence could therefore be increased by the higher densities of flowering plants. Further study is required to clarify this point. For example, Silva et al. (2010) did not observe any improvement in selected plants sown with respect to resident vegetation. On the other hand, naturally occurring VC in olive orchards offers significant advantages. Implementation of this type of habitat management is inexpensive, and disruption is cut to a minimum as sowing/resowing is not required. VC strip width, natural senescence, and warm climate-adapted weed composition minimize yield loss due to competition which is critical for within-crop habitats (Norris and Kogan 2000). These advantages are likely to make growers less reluctant to adopt this habitat management practice and its implementation on a significant scale. Our study also presents evidence that resident VC in olive groves is temporally and spatially adequate to increase these valuable predators as recommended by Landis et al. (2000). VC in its most productive period coincides with the build-up in lacewing populations, and proximity to trees facilitates olive canopy colonization and availability for biological control. However, we would like to point out that the impact of increased larval population on P. oleae egg predation was not recorded in this study. The biological control services provided by these predators may depend on a number of factors other than temporal and spatial adequacy (e.g., intraguild predation and prey switching). Future studies on this system should thus attempt to measure P. oleae egg consumption and prevention of olive fruit damage in relation to habitat management strategies. Despite this fact, resident VC may be regarded as a valuable habitat manipulation resource for controlling P. oleae within the framework of pest management programs.
Author contribution statement
MC and JC obtained the funds. MP, MC, BC, and JC conceived and designed the study. JC provided the experimental setup. MP, BC, and JC executed the experiments. MP analyzed the data and wrote the manuscript. The manuscript was revised and approved by all the authors.
References
Aguilar-Fenollosa E, Jacas J (2013) Effect of ground cover management on Thysanoptera (thrips) in clementine mandarin orchards. J Pest Sci 86:469–481
Bianchi FJJA, Booij CJH, Tscharntke T (2006) Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. P Roy Soc B-Biol Sci 273:1715–1727
Bone NJ, Thomson LJ, Ridland PM, Cole P, Hoffmann AA (2009) Cover crops in Victorian apple orchards: effects on production, natural enemies and pests across a season. Crop Prot 28:675–683
Bugg RL, Dutcher JD, McNeill PJ (1991) Cool-season cover crops in the pecan orchard understory: effects on Coccinellidae (Coleoptera) and pecan aphids (Homoptera: Aphididae). Biol Control 1:8–15
Burnham KP, Anderson DR (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Campos M (2001) Lacewings in Andalusian olive orchards. In: McEwen P, New TR, Whittington AE (eds) Lacewings in the crop environment. Cambridge University Press, Cambridge, pp 492–497
Canard M, Letardi A, Thierry D (2007) The rare Chrysopidae (Neuroptera) of southwestern Europe. Acta Oecol 31:290–298
Corrales N, Campos M (2004) Populations, longevity, mortality and fecundity of Chrysoperla carnea (Neuroptera, Chrysopidae) from olive-orchards with different agricultural management systems. Chemosphere 57:1613–1619
Daane KM (2001) Ecological studies of released lacewings in crops. In: McEwen PK, New TR, Whittington AE (eds) Lacewings in the crop environment. Cambridge University Press, Cambridge, pp 338–350
Danne A, Thomson LJ, Sharley DJ, Penfold CM, Hoffmann AA (2010) Effects of native grass cover crops on beneficial and pest invertebrates in Australian vineyards. Environ Entomol 39:970–978
Duelli P (2001) Lacewings in field crops. In: McEwen P, New TR, Whittington AE (eds) Lacewings in the crop environment. Cambridge University Press, Cambridge, pp 158–171
ESYRCE (2014) Encuesta sobre superficies y rendimientos de cultivos. Ministerio de Medio Ambiente y Medio Rural y Marino, Madrid. http://www.magrama.gob.es/es/estadistica/temas/estadisticas-agrarias/boletin2014_tcm7-384110.pdf. Accessed date on Aug 6 2015
Fewster RM, Buckland ST, Siriwardena GM, Baillie SR, Wilson JD (2000) Analysis of population trends for farmland birds using generalized additive models. Ecology 81:1970–1984
Gómez JA, Guzmán MG, Giráldez JV, Fereres E (2009) The influence of cover crops and tillage on water and sediment yield, and on nutrient, and organic matter losses in an olive orchard on a sandy loam soil. Soil Till Res 106:137–144
Landis DA, Wratten SD, Gurr GM (2000) Habitat management to conserve natural enemies of arthropod pests in agriculture. Annu Rev Entomol 45:175–201
McEwen PK, Ruiz J (1994) Relationship between non-olive vegetation and green lacewing eggs in a Spanish olive orchard. Antenna 18:148–150
Monserrat VJ (2008) Nuevos datos sobre algunas especies de crisópidos (Insecta: neuroptera: Chrysopidae). Heteropterus Rev Entomol 8:171–196 (In Spanish)
Monserrat VJ (2010) Nuevas e interesantes citas de neurópteros en la Península Ibérica (Insecta: Neuroptera). Heteropterus Rev Entomol 10:19–34 (In Spanish)
Morris TI, Campos M, Kidd NAC, Symondson WOC (1999) What is consuming Prays oleae (Bernard) (Lep.: Yponomeutidae) and when: a serological solution? Crop Prot 18:17–22
Moser T, Rombke J, Schallnass HJ, Van Gestel CAM (2007) The use of the multivariate Principal Response Curve (PRC) for community level analysis: a case study on the effects of carbendazim on enchytraeids in Terrestrial Model Ecosystems (TME). Ecotoxicology 16:573–583
New TR (2005) Invertebrate conservation and agricultural ecosystems. Cambridge University Press, Cambridge
Norris RF, Kogan M (2000) Interactions between weeds, arthropod pests, and their natural enemies in managed ecosystems. Weed Sci 48:94–158
Porcel M, Cotes B, Campos M (2011) Biological and behavioral effects of kaolin particle film on larvae and adults of Chrysoperla carnea (Neuroptera: Chrysopidae). Biol Control 59:98–105
Porcel M, Ruano F, Cotes B, Peña A, Campos M (2013) Agricultural management systems affect the green lacewing community (Neuroptera: Chrysopidae) in olive orchards in southern Spain. Environ Entomol 42:97–106
Prasifka JR, Hellmich RL, Dively GP, Lewis LC (2005) Assessing the effects of pest management on nontarget arthropods: the influence of plot size and isolation. Environ Entomol 34:1181–1192
Ramos P, Ramos JM (1990) Veinte años de observaciones sobre la depredación oófaga en Prays oleae Bern. Granada (España), 1970-1989. Bol Sanidad Veg Plagas 16:119–127 (In Spanish)
Rieux R, Simon S, Defrance H (1999) Role of hedgerows and ground cover management on arthropod populations in pear orchards. Agr Ecosyst Environ 73:119–127
Silva EB, Franco JC, Vasconcelos T, Branco M (2010) Effect of ground cover vegetation on the abundance and diversity of beneficial arthropods in citrus orchards. B Entomol Res 100:489–499
Simon S, Bouvier J-C, Debras J-F, Sauphanor B (2010) Biodiversity and pest management in orchard systems. A review. Agron Sustain Dev 30:139–152
Smith MW, Arnold DC, Eikenbary RD, Rice NR, Shiferaw A, Cheary BS, Carroll BL (1996) Influence of ground cover on beneficial arthropods in pecan. Biol Control 6:164–176
Song B, Wu H, Kong Y, Zhang J, Du Y, Hu J, Yao Y (2010) Effects of intercropping with aromatic plants on the diversity and structure of an arthropod community in a pear orchard. Biocontrol 55:741–751
Stelzl M, Devetak D (1999) Neuroptera in agricultural ecosystems. Agr Ecosyst Environ 74:305–321
Szentkirályi F (2001a) Ecology and habitat relationships. In: McEwen P, New TR, Whittington AE (eds) Lacewings in the crop environment. Cambridge University Press, Cambridge, pp 82–115
Szentkirályi F (2001b) Lacewings in fruit and nut crops. In: McEwen P, New TR, Whittington AE (eds) Lacewings in the crop environment. Cambridge University Press, Cambridge, pp 172–238
Thomson LJ, Hoffmann AA (2009) Vegetation increases the abundance of natural enemies in vineyards. Biol Control 49:259–269
Tscharntke T, Klein AM, Kruess A, Steffan-Dewenter I, Thies C (2005) Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecol Lett 8:857–874
Van den Brink PJ, Braak CJFT (1999) Principal response curves: analysis of time-dependent multivariate responses of biological community to stress. Environ Toxicol Chem 18:138–148
Villenave J, Deutsch B, Lode T, Rat-Morris E (2006) Pollen preference of the Chrysoperla species (Neuroptera: Chrysopidae) occurring in the crop environment in western France. Eur J Entomol 103:771–777
Wood SN (2006) Generalized additive models: An introduction with R. Chapman & Hall/CRC, Boca Raton
World Reference Base IUSS Working Group (ed) (2006) World reference base for soil resources. World soil resources reports 103. 2nd ed. FAO, Rome
Wyss E (1995) The effects of weed strips on aphids and aphidophagous predators in an apple orchard. Entomol Exp Appl 75:43–49
Wyss E (1996) The effects of artificial weed strips on diversity and abundance of the arthropod fauna in a Swiss experimental apple orchard. Agr Ecosyst Environ 60:47–59
Yee TW, Mitchell ND (1991) Generalized additive models in plant ecology. J Veg Sci 2:587–602
Zuur AF, Ieno EN, Smith GM (2007) Analysing ecological data. Springer, New York
Zuur AF, Ieno EN, Walker N, Saveliev AA, Smith GM (2009) Mixed effects models and extensions in ecology with R. Springer, New York
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Method Ecol Evol 1:3–14
Acknowledgments
We wish to thank Dr. Víctor Monserrat for his contribution to Chrysopidae identification and Dr. Daniel Paredes, Rafael Alcalá, María Luisa Fernández Sierra, and Herminia Barroso for their laboratory and field assistance. We would also like to thank the stateowned Empresa Pública para el Desarrollo Agrario y Pesquero, S.A (D.A.P.S.A.) for their cooperation, Michael O’Shea for proof-reading the manuscript, and two reviewers for their useful comments. The study was funded by the Junta de Andalucía (project P07-AGR-02747).
Compliance with Ethical Standards
This study was funded by the Junta de Andalucía (Project P07-AGR-02747). The authors declare that they have no conflict of interest. This article does not contain any studies with human participants or animals performed by any of the authors.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by B. Lavandero.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Porcel, M., Cotes, B., Castro, J. et al. The effect of resident vegetation cover on abundance and diversity of green lacewings (Neuroptera: Chrysopidae) on olive trees. J Pest Sci 90, 195–206 (2017). https://doi.org/10.1007/s10340-016-0748-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10340-016-0748-5