
Abstract
Among territorial birds, the number of conspecific neighbours sets the social context of communication. There have been many investigations of vocal behaviour and its important role in territory defense and mate attraction in birds; however, the effect of the density of conspecific neighbours on avian communication has received little attention. In this study, we reviewed the literature on the influence of local density on vocal communication in birds, exploring how the number of conspecific neighbours influences avian vocal behaviour, by conducting a literature review following the guidelines of the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) in two databases: Web of Science and Scopus. We also included additional articles from a more flexible search in Google Scholar. We found 19 studies of birds that included data on vocal behaviour in relation to the number of conspecific neighbours. The majority of these studies (14 of 19) showed that local density influences avian vocal behaviour. Overall, these studies revealed an effect of local density of conspecifics on song rate (expressed as song rate, syllable rate, solo song rate, or singing rate). Other vocal behaviours that were shown to vary with local density included song-type switching, song duration, peak frequency, and song variation. Most studies focused on temperate-zone species where males are the predominant singers. Our results reveal a bias towards the study of male songs in the context of local density, with very few studies on female song; this corresponds with a historical geographical focus on temperate birds. We present experimental design suggestions for future investigations, including predictions for males and females of tropical and temperate species. Finally, we argue that more research on this topic is needed, and that investigations of vocal communication will benefit from further study of the influence of density on avian vocal behaviour.
Zusammenfassung
Verhaltenskonsequenzen von Artgenossen-Nachbarn: Eine systematische Literaturübersicht über die Auswirkungen der lokalen Dichte auf die vokale Kommunikation von Vögeln. In territorialen Vögeln setzt die Anzahl konspezifischer Artgenossen in der Nachbarschaft dessen Gesangsverhalten fest. Obwohl es diesbezüglich viele Untersuchungen im Zusammenhang mit Revierverteidigung und Partnerattraktion bei Vögeln gibt, wurde der Einfluss der Dichte von konspezifischen Artgenossen auf den Vogelgesang wenig beachtet. Diese Studie untersuchte die Literatur über den Einfluss der lokalen Dichte auf das Gesangsverhalten von Vögeln und den Einfluss der Anzahl konspezifischer Arten in der Nachbarschaft auf deren Kommunikation. Die Literaturstudie wurde nach den Richtlinien des Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) in zwei Datenbanken durchgeführt: Web of Science und Scopus. Wir haben auch zusätzliche Artikel aus einer flexibleren Suche in Google Scholar eingefügt. Wir fanden neunzehn Studien über das Gesangsverhalten von Vögeln im Zusammenhang mit der Anzahl von lokalen konspezifischen Artgenossen. Die meisten Studien (14 von 19) zeigten, dass die lokale Dichte die Gesangsrate (ausgedrückt als Gesangsrate, Silbenrate, Sologesangsrate oder Singrate) von Vögeln beeinflusst. Andere Gesangsverhalten, die mit der lokalen Dichte variierten, umfassten den Wechsel von Gesangstypen, die Gesangsdauer, die Höchstfrequenz und die Gesangsvariation. Die meisten Studien konzentrierten sich auf Arten der gemäßigten Zone, wo überwiegend Männchen singen. Daraus resultierend ergaben unsere Ergebnisse eine Bevorzugung der Untersuchung des Gesangsverhaltens männlicher Vögel im Kontext der lokalen Dichte durch den geografischen Fokus wissenschaftlicher Artikel auf Vögel der gemäßigten Zonen. Mit dieser Studie stellen wir zukünftige Untersuchungsvorschläge, einschließlich Vorhersagen für Männchen und Weibchen tropischer und gemäßigter Vogelarten. Weitere Forschung zu diesem Thema ist notwendig, da Studien zu dem Gesangsverhalten von Vögeln von weiteren Untersuchungen zu dem Einfluss der lokalen Dichte auf den Vogelgesang profitieren würden.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The density of conspecific individuals affects the social behaviour of diverse animal taxa in a variety of contexts. For territorial species, territories define the spatial relationships of nearby individuals, and signals used to defend territorial resources are public information that is shared among neighbours. This is true for many species of songbird, who defend territories from conspecific animals and rely heavily on acoustic signals for social and sexual communication (Todt and Naguib 2000; Catchpole and Slater 2003; Naguib 2005). Many avian acoustic signals transmit over long distances and whenever birds’ acoustic signals travel beyond their territory boundaries, it is worthwhile to consider the interactions between more than two individuals as a communication network (McGregor 2005). Communication networks in songbirds have received considerable attention, since bird territories are often smaller than the transmission range of bird vocalizations, and these ideas have expanded our understanding of the effects of nearby conspecific animals on the vocal behaviour of birds (Burt and Vehrencamp 2005; Foote et al. 2010; Snijders and Naguib 2017). Although the social context of avian vocal communication has been recognized as a factor that affects individual social behaviour, the influence of the density of neighbours on songbird vocal behaviour has received scant attention.
Conspecific neighbours constitute the social environment for territorial songbirds when they sing to defend territories and attract mates. It is well known that females assess males based on features of their vocal behaviour (Searcy and Andersson 1986), including traits such as repertoire size (Franco and Slabbekoorn 2009), vocal performance (Ballentine et al. 2004; Janicke et al. 2008; Podos et al. 2009), singing rate (Alatalo et al. 1990; Nolan and Hill 2004), and performance in vocal interactions with other males (Mennill et al. 2002; Naguib 2005). At the same time, it is well known that both males and females use songs to defend territorial limits (Nowicki et al. 1998; Langmore 2000; Logue 2005; Mennill 2006) where salient traits include song matching (Beecher et al. 2000; Vehrencamp 2001), song rate (Cooney and Cockburn 1995; Searcy and Beecher 2009; Searcy et al. 2014), and performance in vocal interactions assessed through eavesdropping (Mennill et al. 2004; Peake et al. 2002). When the number of conspecific birds in close proximity changes, both the inter-sexual and intra-sexual dynamics of vocal behaviour may change. Therefore, we must develop a deeper understanding of the importance of local density in driving variation in acoustic communication in birds.
Although the historical focus of bird song research has been on males, female song has received more attention in recent years (Odom et al. 2014; Riebel et al. 2019). There is a growing effort to document female song across the globe and to understand its function in different ecological and social settings (Wilkins et al. 2020; Austin et al. 2021). Females sing and perform duets commonly in tropical regions and south-temperate regions (Tobias et al. 2016; Odom and Benedict 2018), and the presence of female song is recognized as the ancestral state of songbirds (Odom et al. 2014), even though female song is comparatively rare in temperate regions (Jetz et al. 2012). Consequently, the historical underrepresentation of studies of female song corresponds to a temperate-zone bias that hampers our understanding of bird behaviour in the globe’s most biodiverse regions (Stutchbury and Morton 2022). From the growing number of studies on female song, as well as male–female duets, we recognize the primary functions of female song are pair bond maintenance, mate attraction, and territory defence, corresponding to parallel functions in male song (Austin et al. 2021). For both well-studied male songbirds and less-studied female songbirds, the influence of the number of conspecific neighbours on acoustic signals remains an open area of investigation.
Density, in ecological terms, is the number of individuals per unit area (Mayor and Schaefer 2005). Density can be studied at many different scales, ranging from the number of individuals in a population distributed across the entire area they inhabit (i.e., “population density”; Connor et al. 2000; Gaston and Matter 2002), to a subset of a population at a smaller scale of adjacent neighbours (i.e., “local density”; Connor et al. 2000; Sharma et al. 2024). For territorial songbirds, local density is often quantified by counting the number of territorial males or territorial pairs in a relatively small area (Falls 1981; Christman 1984). Behavioural ecologists have focused on local density in studies of the effects of conspecific neighbours of territorial individuals, both in temperate species (Hiebert et al. 1989; Moiron et al 2015; Cain and Langmore 2016) and tropical species (Osmun and Mennill 2011; Demko and Mennill 2018; Stutchbury and Morton 2022). We focus on local density in this investigation and use the word “density” to mean the number of conspecific neighbours (e.g., number of singing males, number of nest boxes occupied by singing males or singing pairs, number of family groups, or number of pairs) influencing the singing behaviour of nearby territorial animals.
In this study, our goal is to review the literature on vocal communication in territorial birds to better understand the effects of conspecific local density on avian vocal behaviour. Specifically, we were motivated to survey the published literature on the influence of the number of neighbours on singing behaviour of birds by focusing on both: (1) male and female birds; (2) tropical and temperate regions; and (3) descriptive and experimental studies. We aimed to summarize the effects of local density on vocal behaviours in birds and propose a path towards future studies that could enhance our understanding of the relationship between local density and avian vocal behaviour.
Literature search methodology
We conducted a systematic review following the guidelines recommended by the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA; Moher et al. 2009). On July 12, 2022, we performed two searches in Web of Science (304 records) and Scopus (184 records) using specific keywords with no date restriction. The search terms in “Topic” were: avian, bird*, density, densities, behaviour*, behavior*, neighbour*, neighbor*, and singing. (The wildcard symbol “*” included in the search any letters after the indicated word; e.g., bird* will include both “bird” and “birds”). Given our focus on birds, we excluded the studies of density and social behaviour in humans (term excluded: human*). The Boolean operators used were “AND” and “OR” for the search in “Topic” and “NOT” for the excluded term in “Topic”. After the search, we removed duplicates from both databases, giving rise to 306 records in total.
The first criterion to include an article in our study was that the title contained the words: “density” or “densities”, and “behaviour*” or “behavior*”. We also considered an article if the title contained the terms “number of conspecifics” or “conspecifics”. After applying the title criterion, we maintained 65 records. The second criterion to include an article in our study was that the authors indicated in the Abstract that they explored the effect of density of conspecific birds on vocal behaviour or on any non-vocal behaviour, which reduced the number of records to 29. For example, we did not include studies focused on estimating density of conspecific animals from autonomous recording units (e.g., Dawson and Efford 2009, Perez-Granados and Traba 2021), or theoretical modeling papers using density of the population and population-averaged signaling effort (Halupka 2017). Finally, only 13 articles specifically indicated in the abstract that local density or the number of neighbours was investigated as a variable affecting vocal behaviours of the focal species. Halupka (2014) was excluded after reading the methods and finding that the results describe a complex variable (specifically: Euclidean distance between spectra of synchronously sampled time series), which is not common in the literature of vocal behaviours of songbirds. Therefore, 12 published articles met our search criteria.
We then performed an additional search in a more flexible database, Google Scholar. In this database, we used the same keywords as in our Web of Science and Scopus searches. We added two keywords: “song*” OR “passeriform*” resulting in approximately 840 articles, and we excluded the word “-intrusion” resulting in approximately 285 articles. We read the title and abstracts of these articles, and found 13 articles that focused on local density of conspecifics and song behaviour. After reading the entire manuscripts, we excluded 6 articles that did not measure the density of conspecifics, or that used playback to elicit vocal responses (Barrero et al. 2023), or compared song features between two different species with different densities (Opaev and Shishkina 2021), or measured population size but did not account for the number of animals per unit area and the spatial relationship between neighbours (Valderrama et al. 2013). Therefore, our additional search added seven articles (19 in total when combined with the original 12 articles) exploring the effect of local density on avian vocal behaviours.
We present our review of this literature by focusing first on studies that looked at song rate or dawn song rate showing an effect caused by a change of the number of conspecifics, and then the studies showing no effect. We present the studies based on publication year starting with the most recent. We draw attention to studies that explored the relationship between local density on song rate, and then the studies that considered additional vocal behaviours beyond song rate.
Local density effects on song rate
We found that 14 of the 19 published studies revealed a relationship between number of conspecific neighbours and the singing behaviour of birds. The species in these 14 studies represented a cross-section of passerine bird species, including oscine and sub-oscine species, including a swallow, a flycatcher, a kingbird, a cardenal, two antbirds (sub-oscines), a widespread wren, a tropical wren, a bush warbler, a warbler, a robin, a sparrow weaver, a sparrow, and a bunting (Table 1). In 9 of the 14 studies testing the effect of local density of conspecific neighbours on song, the researchers focused on song rate using both experimental and correlational approaches (Table 1). The other five studies focused on the following vocal behaviours: song-type switching, song duration, peak frequency, daily song rate, and song-type variation (Table 1).
In an observational study of White-browed Sparrow Weavers (Plocepasser mahali) in South Africa, males produced longer solo songs in an area with higher local density when compared with individuals with fewer neighbours, and solo song rate and duet and chorus rate increased with the number of neighbours (Voigt et al. 2021). Sparrow weavers are colonial breeders where social groups include a dominant male, a dominant female, and their offspring. Only dominant males sing solo songs; duets are performed by a pair (dominant male and dominant female), and choruses are performed by the pair with members of the group. Duets and choruses were analyzed together revealing an effect of local density. The main explanation for higher solo song rates was that males in low-density areas have more food and more time to sing. Higher duet and chorus rates were explained by the increase in territorial interactions with neighbouring groups in areas with higher density at territorial boundaries when compared with individuals with fewer neighbours.
A comparative study of the effects of noise on vocal behaviour in urban and rural populations of Northern Cardinals (Cardinalis cardinalis) in the United States (Narango and Rodewald 2016), explored variation with conspecific density on changes in vocal behaviours. Higher density of conspecific cardinals was associated with behavioural differences including both temporal and frequency features of songs during aggressive encounters (Table 1). Urban populations had higher densities than rural populations, and syllable rate of cardinal songs was best explained by high densities of conspecific animals in urban environments. Therefore, variation in density associated with an urban environment appears to contribute to changes in temporal features of cardinal songs.
An opportunistic study of local density in Dusky Flycatchers (Empidonax oberholseri) in Canada revealed an effect of the number of conspecific neighbours on dawn singing and song rate (Stehelin and Lein 2014). The authors measured the dawn chorus and the singing rate of 13 flycatchers before a series of three storms, as well as the 6 individuals that remained after the storms. They reported a lack of dawn chorus during the post-storm period and a dramatic reduction in singing rate throughout the day (Table 1). Additionally, the authors simulated intrusions to the territory for the post-storm males finding an increase in vocalization at dawn, and this response supported a key prediction of the social dynamics hypothesis (the hypothesis that male singing behaviour mediates social relationships between neighbors through interactive communication, as explained below), given that singing was influenced by the presence of other males.
The effect of neighbours on dawn singing and song rate on the Brownish-flanked Bush Warbler (Cettia fortipes) in China has also been explored in the context of the social dynamics hypothesis (Xia et al. 2014). This study hinged on a comparison of the song rate for 22 males before and after sunrise, comparing the number of neighbours for birds in two categories: males with 0 or 1 neighbours versus males with 2–4 neighbours. Bush Warblers sang earlier and with higher singing rates when they had more neighbouring males, supporting the idea that pre-dawn song plays a role in social interactions between males.
Two sympatric tropical sub-oscine songbirds, Yellow-breasted Warbling-Antbirds (Hypocnemis subflava) and Peruvian Warbling-Antbirds (H. peruviana) in Peru showcase variation in song rate associated with the behaviour of conspecific neighbours (Tobias et al. 2011). Colour-banded males of both species were observed during 10-min focal observations and minimum convex polygons were drawn around the birds’ locations to model the spatial locations of neighbours. For Yellow-breasted Warbling-antbirds, 8 pairs had a single neighbour and 11 pairs had two neighbours; for Peruvian Warbling-Antbirds, 8 pairs had one or two neighbours, whereas 12 pairs had three or four neighbours (Table 1). In both species, song rate increased with number of neighbours (Tobias et al. 2011) demonstrating that these complex acoustic ornaments should be understood in the context of social selection and communication between neighbours.
A study of Eastern Kingbirds (Tyrannus tyrannus) in the United States explored the singing rate during the dawn chorus under the premise that song performance is mediated by the presence of females, males, or both (Sexton et al. 2007). Local density of neighbours was specifically defined as the inverse distance to the nearest neighbour. The study showed that dawn song onset was earlier (i.e., males began singing earlier) at closer distances to the nearest neighbour and that song rate increased as the number of neighbours increased. The influence of local density on song rate was suggested to reflect the likelihood of extra-pair copulations, which occur immediately before dawn in this species (Sexton et al. 2007).
A removal experiment designed to investigate the function of male song during the dawn chorus and during the day showed that male Chipping Sparrows (Spizella passerina) reduced the dawn chorus and daily singing rate when neighbours were removed (Liu 2004). Nine clusters of sparrows were subject to removals, where all neighbours of a focal bird were removed, later to be released following 2 days of song recording. During removals, the remaining male reduced his dawn chorus from 17–36 min to 1–12 min, and singing rate during the day also decreased. There were also changes in non-vocal behaviours; before the removals, males sang their dawn chorus around the territory boundaries and on the ground, but during removals six males sang from the center of the territory, two males sang at higher perches, and three males simply did not sing at all. With an especially compelling experimental design, this study reveals that both dawn chorus and daytime singing behaviour, including song rate and song position, are heavily influenced by the number of conspecific neighbours.
Finally, in Corn Buntings (Miliaria calandra), researchers evaluated the singing activity of males in relation to the number of conspecific neighbours in a population in Poland (Olinkiewicz and Osiejuk 2003). Buntings were censused through population level surveys every 10 days, and birds showed lower song rates when they had two or three neighbours compared to one neighbour. The explanation for this result was that males with more neighbours trade-off singing time to actively defend the territories against neighbours, and to better listen to their neighbours.
Local density effects on other vocal behaviours
In three additional correlational studies (Owen and Mennill 2021; Goretskaia 2004; Galeotti et al. 1997), song rate showed no relationship with the number of conspecifics, but other song features did reveal an effect of density (Table 1). In contrast, two experimental studies and one correlational study looked at the effect of local density of conspecific neighbours while not considering song rate as a response variable, but instead focusing on dawn song onset, spectrographic features of songs (Ripmeester et al. 2010; Stuart et al. 2019; Grabarczyk et al. 2020), and temporal features of songs (Ripmeester et al. 2010). We describe these six investigations below.
Duetting neotropical Rufous-and-white Wrens (Thryophilus rufalbus) in Costa Rica showed that conspecific local density influenced the song-type switching of females (Owen and Mennill 2021). Females showed higher song-type switching in cases where they had two neighbouring pairs compared with only one neighbouring pair. This result arose from analyses of a long-term dataset, spanning 17 years and 45 pairs of territorial birds, comparing vocal behaviour of females and males (solo song rate, duet rate, repertoire use, and song-type switching rate). The explanation for higher song-type switching performed by females, when the focal pair had 1 neighbour in comparison to 2 neighbours, was the increased territorial defense against conspecifics. The authors suggested the need to test hypotheses in females and males separately and avoid assuming that both sexes are influenced in the same way by external factors.
A study of Willow Warblers (Phylloscopus trochilus) in Russia explored the influence of social context on song structure and singing behaviour (Goretskaia 2004). The author mapped the territories of nesting males in two locations, one with high density and one with low density, and evaluated multiple aspects of vocal behaviour including song rate, notes in the initial phrases of songs, and number of double songs (Table 1). Males in the high-density population performed more double songs and sang more notes in the initial phrase of songs. Further, song rate was higher in the high-density population, although the difference was not statistically significant. This study demonstrated a link between song structure and density, supporting the hypothesis that song versatility and population density are related (Kroodsma 1999).
A study of Barn Swallows (Hirundo rustica) in Italy revealed that males sing shorter songs at higher pitches in a context of more conspecific competitors (Galeotti et al. 1997). The number of conspecific neighbouring swallows varied from zero to five individuals, and seven different song features were measured. The results demonstrated an effect of number of nearby males on two features: song length (songs were longer with fewer nearby males) and peak frequency (songs showed higher peak frequency with more nearby males). This study concluded that longer and more varied song evolved to enhance female preference in a highly competitive scenario associated with increases in the number of conspecific rival males.
The first of two studies of House Wrens (Troglodytes aedon) in the United States explored the influence of biotic and environmental factors on the onset of dawn song, with reference to the importance of human-generated light and noise pollution (Stuart et al. 2019). Both the birds’ nesting stage and number of neighbours (as well as temperature) had an effect on song onset. The number of neighbours was experimentally manipulated by creating groups of nest boxes; males were experimentally subjected to situations where they had zero-to-four neighbouring males. House Wren males began singing earlier relative to civil twilight as the number of breeding males increased within 100 m of their nest boxes. The explanation for this result was higher competition in areas occupied by more pairs. The authors highlighted that in House Wrens, territory defense is important for dawn song onset but is not the sole influencing factor, given that lone males still produce songs.
In a second study of the same population of House Wrens, the number of neighbours and female fertile status affected the frequency structure of male song. Males increased the peak frequency of songs and reduced the duration of songs as the number of neighbours increased up to 4 neighbours (Grabarczyk et al. 2020). Through measurements of spectral and temporal features of songs (song duration, minimum frequency, maximum frequency, peak frequency, and bandwidth), the authors tested the hypothesis that social environment influences song structure. Local density of neighbours was experimentally controlled by creating 18 clusters with six nest boxes each, separated by 150 m. The peak frequency of song increased with number of neighbours (specifically when comparing males with no neighbours within 150 m to males that had one or more neighbours within 150 m); males with neighbours sang shorter songs compared to males without neighbours, revealing an effect of local density on song structure. A decrease in song duration could reduce the chances of song overlapping with the song of other territorial neighbours, which may contribute to fewer aggressive interactions between neighbours, since song overlap has been suggested to serve as an aggressive behaviour (Naguib and Mennill 2010).
Finally, a study of ambient noise and density in urban and forest populations of European Blackbirds (Turdus merula) in the Netherlands, explored whether two habitats vary acoustically and in terms of territory density and social interactions (Ripmeester et al. 2010). Acoustic features of song duration were measured in two components of the song (the motif and the twitter) in a higher-density city population and a lower-density forest population. The twitter portion of male song increased with territory density, and intra-song pause duration showed a negative relationship with territory density (Table 1). Territory density was associated with changes in temporal features of songs.
Studies showing no effect of local density on vocal behaviours
We found five studies that reported no effect of local density on singing behaviour, across several unrelated songbirds: a wren, a warbler, a tit, a blackbird, and a community of temperate birds (Table 1). Four studies were conducted in North America and one was conducted in Japan; three were correlational studies and two were removal experiments.
In Canyon Wrens (Catherpes mexicanus) in the United States, female song rate did not show regional or local variation in response to variation in local density (Dargis et al. 2021). This study followed a correlational approach to test for vocal responses of females to changes in density of conspecific animals, using nearest-neighbour distances to quantify local density. Two study populations varied in the number of individuals in Colorado (low density) and Arizona (high density). Additionally, a playback experiment simulating territorial intrusions was performed in Arizona to test for female vocal responses. In both the comparison of low-density and high-density populations, and in the playback experiment in the high-density population, female song was not shown to change in response to variation in the local density of conspecific animals. This study stands out as the only investigation revealed in our literature search that focused exclusively on female singing behaviour.
A multi-species study of a sub-alpine valley community in the United States, explored vocal individuality of seven bird species: Green-tailed Towhee (Pipilo chlorurus), Lazuli Bunting (Passerina amoena), Lincoln’s Sparrow (Melospiza lincolnii), Mountain White-crowned Sparrow (Zonotrichia leucophrys oriantha), American Robin (Turdus migratorius), House Wren (Troglodytes aedon), and Warbling Vireo (Vireo gilvus) (Blumstein et al. 2012). The authors measured relative density from point counts for each species, and extracted multiple measurements of frequency and temporal features of songs of 10 individuals per species, as well as calculating temporal features of songs (number of syllables/song duration; Table 1). These birds revealed no evidence that vocal individuality in song varied with density of breeding conspecifics, suggesting that breeding bird density is not a driver of the evolution of individually distinctive vocalizations.
A removal experiment testing the effect of local density on Black-throated Blue Warblers (Setophaga caerulescens) in the United States explored the relationship between density and reproduction and social behaviour (Sillett et al. 2004). Countersinging was measured during male–male interactions, both before and after removal. Removal yielded no effect on singing behaviour of males in adjacent territories. The authors argued the lack of effect was due to the timing of the pair removals that occurred when the territories were already established.
In a correlational study across 22 urban parks in Japan, researchers tested the effect of noise and density of conspecifics on the vocal features of the songs of Great Tits (Parus major; Hamao et al. 2011). The authors measured diverse spectro-temporal features across 22 males, with one male randomly selected from each urban park (Table 1), and estimated density as the number for males in a 25 m radius around the focal male. There was a marginal effect of density of conspecifics on the minimum frequency and the number of phrases in songs; but, the effect appeared only after controlling statistically the multiple regressions. The authors did not found a clear effect of density of conspecifics on Great Tits' vocal features, but they highlight that more research on the effects of noise on birdsong should consider both noise and the influence of density on song structure.
Finally, a temporary territorial removal study on Red-winged Blackbirds (Agelaius phoeniceus) in Canada assessed variation in song rate of territorial males during male–male interactions in relation to the number of neighbours (Shutler and Weatherhead 1991). After the temporary removal of neighbours, the song rate of focal males did not change, nor did it change when the unoccupied territories became occupied by floater males. Focal males had zero-to-three or more neighbours and showed no variation in song rate. The authors suggested that song provides basic information that a territory is occupied, but does not appear to vary with local density.
Synthesis of the effects of local density on vocal behaviour
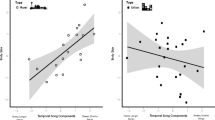
Our review of the primary literature, following a standardized guideline for systematic reviews, and supplemented by an additional search, reveals that the number of conspecific neighbours has been considered in 19 studies across the passeriform songbirds, and that there have been only a few explicit tests of this idea. We found 19 investigations that compared vocal behaviour in relation to variation in the number of conspecific birds (summarized in Table 1). The majority—74% of 19 studies—presented evidence that some facet of bird singing behaviour shows a relationship with the number of neighbours. Song rate was the most common vocal response that changed with local density, and this pattern held true in both correlational and experimental investigations (Liu 2004; Sexton et al. 2007; Xia et al. 2014; Stehelin and Lein 2014; Voigt et al. 2021). The main pattern observed considering temperate and tropical studies is that males often show different singing behaviours in relation to local density (Fig. 1). By contrast, females did not show a vocal response to variation in local density in the temperate zone (Dargis et al. 2021; Fig. 1A), but did show a response in the tropics (a duetting species studied by Owen and Mennill 2021, and two duetting antbird species Tobias et al. 2011; Fig. 1B).

Illustration of the prediction of the effect of higher local conspecific density on male and female vocal behaviours in temperate versus tropical regions. In temperate regions (left), we expect a higher response in the intensity of vocal behaviours when the density of local conspecific neighbours is higher, but no change for females, since they do not participate as intensely in vocal behaviours associated with territory defence. In tropical regions (right), we expect a similar change in the intensity of vocal behaviours at higher local density of conspecifics for both males and females
Two studies included in our review tested the social dynamics hypothesis to explain the variation of song rate during the dawn chorus. This hypothesis states that male vocal behaviour during the dawn chorus mediates social relationships between territorial neighbours through interactive communication (Staicer et al. 1996). One prediction of this hypothesis is that a higher number of nearby conspecific individuals will increase the pressure to interact during the dawn chorus. In both studies that considered this hypothesis (Stehelin and Lein 2014; Xia et al. 2014), an increase in the number of neighbours resulted in an increase in singing rate, supporting the idea that interactions are mediated by social factors during the dawn chorus. Among many singing behaviours that can be measured in studies of the social dynamics hypothesis (e.g., frequency matching and overlapping, Foote et al. 2008), song rate was shown to vary with the number of neighbours (Stehelin and Lein 2014; Xia et al. 2014). In future investigations, we recommend that investigators test the social dynamics hypothesis by measuring or controlling for the number of conspecific neighbours, possibly through temporary removal of neighbours to focal individuals (as in Liu 2004) or through increasing the number of neighbours experimentally with playback simulating male and female song (Fig. 2). These experimental designs could expand beyond a test of the aforementioned hypothesis to elucidate the function of dawn chorus and daytime singing behaviour while controlling for the social environment, expanding our knowledge of the effects of the number of conspecific neighbours on bird singing behaviour. We note that removal experiments may yield interesting insights on the effect of the number of neighbours on avian vocal signals, yet they must be conducted carefully, because removal experiments pose ethical challenges (particularly for the removed birds), and they create an unusual situation where territorial dynamics are altered at the same time that local density is manipulated.

Schematic representation of three different methodological approaches to test the effect of local density of conspecific neighbours on avian vocal behaviour. For each approach, we show a pair of birds with a dotted line representing their territory (in black), as well as neighbouring birds and their territories (in grey). The first approach involves comparison of behaviour between birds across natural differences in density. The second approach involves experimental removal of birds as a tool for decreasing density (represented by the grey “X” symbols). The third approach involves experimental addition of birds simulated through playback (represented by black speaker symbols) as a tool for increasing apparent density
Only 2 of the 19 studies considered variation in female vocal behaviour in response to variation in local density in an explicit test. Both of these investigations tested female song responses to an increase in local density using a correlational approach (Dargis et al. 2021; Owen and Mennill 2021). In the tropical species (Rufous-and-White Wrens), females showed increased song-type switching at higher densities. In contrast, in the temperate species (Canyon Wrens), female vocal behaviour did not vary with local density. The study of antbirds combined the data for males and females as the females mostly participate in duets (Tobias et al. 2011), although the results supported the prediction of higher vocal responses in females as conspecific density increases (Fig. 1B). The limited evidence on the function of female song in songbirds, especially in tropical species, is an opportunity to explore whether vocal behaviour varies with local density both in males and females, and should be an area of focus in future investigations, both within the wrens (where female song, as well as male–female duets, are commonplace) and beyond the wrens as well.
It is paramount to consider the different pressures that influence territoriality in temperate and tropical regions while testing the effect of local density on vocal behaviour. It is well known that males in temperate regions sing to defend resources that allow them to attract breeding partners during a relatively short breeding season (Searcy 1992; Nowicki and Searcy 2005) and females use song output and song features to evaluate potential mates (Reid et al. 2004; Riebel 2009). Therefore, we anticipate a stronger response to variation in local density for males compared to females in the temperate zone (Fig. 1A). In the tropics, by contrast, it is common that males and females defend year-round territories (Stutchbury and Morton 2001, 2022) and both sexes sing (Slater and Mann 2004; Odom 2014; Riebel et al. 2019). Across examinations of the hypotheses for why females sing, there is support for territorial defense in some cases when the male is not on the territory as well as in polygynous systems (Langmore 2000; Slater and Mann 2004). Species that perform duets in the tropics add additional cases where females sing, in the context of joint female–male signals (i.e., duets) which are important in defending territories and aiding in maintaining contact between pair members (Langmore 1998; Mennill et al. 2006; Mennill and Vehrencamp 2008; Tobias et al. 2011). If female song plays a primary role in mate attraction and pair bond maintenance, we would expect lower song output when the number of neighbours is lower. However, if female song is important in territory defence (as it is in males), then we should anticipate similar responses to males in terms of the number of neighbours (Fig. 1B), since a greater number of neighbours will result in a higher risk of agonistic interactions and potential reduction in territory size. Consequently, this pattern would be especially anticipated in tropical regions where female song is a common trait (Fig. 1B).
Conclusions and a call for future studies
Based on our systematic review of the literature published to date, we found evidence that the number of conspecific neighbours is indeed a factor that affects singing behaviour of songbirds, in spite of the fact that this variable social feature has seldom been studied. One focus of the research on this topic has centered on the social dynamics hypothesis explaining the dawn chorus in songbirds; the emerging pattern reveals that males adjust their song rate depending on the number of neighbours. All studies argued that more neighbours necessitate more vocal interactions and therefore elicit an increased song rate. Beyond song rate, our review also showed that other vocal behaviours can reflect an increased number of neighbours, including song-type switching, song duration, peak frequency, and song variation.
Our review provides a case study of the widely recognized bias towards studies of male song and temperate regions. We found that female vocal behaviour was rarely studied in the context of changes in local density (represented in just 10.5%, 2 of 19, of the studies included in our systematic review), in spite of the fact that female song is a widespread and ancestral feature across songbirds (Odom et al. 2014; Odom and Benedict 2018). Given the limited evidence from the two studies on female song, and based on the distribution of bird species where females sing, we expect that future studies will yield insight into the influence of local density on female song in tropical regions (Fig. 1). We highlight experimental designs to further investigate vocal responses in songbirds mediated by number of neighbours, and predictions considering males and females in temperate and tropical regions (Fig. 2). Experimental removals, and playback manipulations, represent two especially promising approaches for future investigations.
Data availability
The data are included in the manuscript.
References
Alatalo RV, Glynn C, Lundberg A (1990) Singing rate and female attraction in the Pied Flycatcher: an experiment. Anim Behav 39:601–603. https://doi.org/10.1016/S0003-3472(05)80430-0
Austin VI, Dalziell AH, Langmore NE, Welbergen JA (2021) Avian vocalisations: the female perspective. Biol Rev 96:1484–1503. https://doi.org/10.1111/brv.12713
Ballentine B, Hyman J, Nowicki S (2004) Vocal performance influences female response to male bird song: an experimental test. Behav Ecol 15:163–168. https://doi.org/10.1093/beheco/arg090
Barrero A, Gómez-Catasús J, Pérez-Granados C, Bustillo-de la Rosa D, Traba J (2023) Conspecific density and habitat quality drive the defence and vocal behaviour of a territorial passerine. Ibis. https://doi.org/10.1111/ibi.13295
Beecher MD, Campbell SE, Burt JM, Hill CE, Nordby JC (2000) Song-type matching between neighbouring song sparrows. Anim Behav 59:21–27. https://doi.org/10.1006/anbe.1999.1276
Blumstein DT, Mcclain DR, De Jesus C, Alarcón-Nieto G (2012) Breeding bird density does not drive vocal individuality. Curr Zool 58:765–772. https://doi.org/10.1093/czoolo/58.5.765
Burt JM, Vehrencamp SL (2005) Dawn chorus as an interactive communication network. In: McGregor P (ed) Animal communication networks. Cambridge University Press, Cambridge, pp 320–343. https://doi.org/10.1017/CBO9780511610363.019
Cain KE, Langmore NE (2016) Female song and aggression show contrasting relationships to reproductive success when habitat quality differs. Behav Ecol Sociobio 70:1867–1877. https://doi.org/10.1007/s00265-016-2192-1
Catchpole CK, Slater PJ (2003) Bird song: biological themes and variations. Cambridge University Press
Christman SP (1984) Plot mapping: estimating densities of breeding bird territories by combining spot mapping and transect techniques. Condor 86:237–241
Connor EF, Courtney AC, Yoder JM (2000) Individuals–area relationships: the relationship between animal population density and area. Ecology 81:734–748
Cooney R, Cockburn A (1995) Territorial defence is the major function of female song in the superb fairy-wren, Malurus cyaneus. Anim Behav 49:1635–1647. https://doi.org/10.1016/0003-3472(95)90086-1
Dargis L, Benedict L, Najar NA (2021) Female bird song rates do not covary with population density in a North American species. Ethology 127:1042–1052. https://doi.org/10.1111/eth.13227
Dawson DK, Efford MG (2009) Bird population density estimated from acoustic signals. J Appl Ecol 46:1201–1209. https://doi.org/10.1111/j.1365-2664.2009.01731.x
Demko AD, Mennill DJ (2018) Male and female signaling behavior varies seasonally during territorial interactions in a tropical songbird. Behav Ecol Sociobiol 72:1–13. https://doi.org/10.1007/s00265-018-2495-5
Falls JB (1981) Mapping territories with playback: an accurate census method for songbirds. Stud Avian Biol 6:86–91
Foote JR, Fitzsimmons LP, Mennill DJ, Ratcliffe LM (2008) Male chickadees match neighbors interactively at dawn: support for the social dynamics hypothesis. Behav Ecol 19:1192–1199. https://doi.org/10.1093/beheco/arn087
Foote JR, Ratcliffe LM, Mennill DJ, Fitzsimmons LP (2010) Black-capped chickadee dawn choruses are interactive communication networks. Behaviour 147:1219–1248
Franco P, Slabbekoorn H (2009) Repertoire size and composition in great tits: a flexibility test using playbacks. Anim Behav 77:261–269. https://doi.org/10.1016/j.anbehav.2008.09.023
Galeotti P, Saino N, Sacchi R, Møller AP (1997) Song correlates with social context, testosterone and body condition in male barn swallows. Anim Behav 53:687–700. https://doi.org/10.1006/anbe.1996.0304
Gaston KJ, Matter SF (2002) Individuals-area relationships: comment. Ecology 83:288–293. https://doi.org/10.2307/2680139
Goretskaia MI (2004) Song structure and singing behaviour of willow warbler Phylloscopus trochilus acredula in populations of low and high density. Bioacoustics 14:183–195. https://doi.org/10.1080/09524622.2004.9753524
Grabarczyk EE, Vonhof MJ, Gill SA (2020) Social context and noise affect within and between male song adjustments in a common passerine. Behav Ecol 31:1150–1158. https://doi.org/10.1093/beheco/araa066
Halupka K (2014) Spreading information in a network of interacting neighbours. PLoS ONE 9:e102801. https://doi.org/10.1371/journal.pone.0102801
Halupka K (2017) Information flow in a network of dispersed signalers-receivers. Phys A Stat Mech 486:610–617. https://doi.org/10.1016/j.physa.2017.05.068
Hamao S, Watanabe M, Mori Y (2011) Urban noise and male density affect songs in the great tit Parus major. Ethol Ecol Evol 23:111–119. https://doi.org/10.1080/03949370.2011.554881
Hiebert SM, Stoddard PK, Arcese P (1989) Repertoire size, territory acquisition and reproductive success in the song sparrow. Anim Behav 37:266–273. https://doi.org/10.1016/0003-3472(89)90115-2
Janicke T, Hahn S, Ritz MS, Peter HU (2008) Vocal performance reflects individual quality in a nonpasserine. Anim Behav 75:91–98
Jetz W, Thomas GH, Joy JB, Hartmann K, Mooers AO (2012) The global diversity of birds in space and time. Nature 491:444. https://doi.org/10.1016/j.anbehav.2007.04.007
Kroodsma DE (1999) Making ecological sense of song development by songbirds. In: Hauser MD, Konishi M (eds) The Design of Animal Communication. MIT Press. https://doi.org/10.7551/mitpress/2359.003.0014
Langmore NE (1998) Functions of duet and solo songs of female birds. Trends Ecol Evol 13:136–140. https://doi.org/10.1016/S0169-5347(97)01241-X
Langmore NE (2000) Why female birds sing. In: Espmark Y, Amundsen T, Rosenqvist G (eds) Animal signals: signalling and signal design in animal communication. Tapir Academic Press, Trondheim, pp 317–327
Liu WC (2004) The effect of neighbours and females on dawn and daytime singing behaviours by male chipping sparrows. Anim Behav 68:39–44. https://doi.org/10.1016/j.anbehav.2003.06.022
Logue DM (2005) Cooperative defence in duet singing birds. Cogn Brain, Behav. 10:497–510
Mayor SJ, Schaefer JA (2005) The many faces of population density. Oecologia 145:275–280. https://doi.org/10.1007/s00442-005-0114-4
McGregor PK (2005) Animal communication networks. Cambridge University Press. https://doi.org/10.1017/CBO9780511610363
Mennill DJ (2006) Aggressive responses of male and female rufous-and-white wrens to stereo duet playback. Anim Behav 71:219–226. https://doi.org/10.1016/j.anbehav.2005.05.006
Mennill DJ, Ratcliffe LM (2004) Do male black-capped chickadees eavesdrop on song contests? A multi-speaker playback experiment. Behaviour 141:125–139
Mennill DJ, Vehrencamp SL (2008) Context-dependent functions of avian duets revealed by microphone-array recordings and multi-speaker playback. Curr Biol 18:1314–1319. https://doi.org/10.1016/j.cub.2008.07.073
Mennill DJ, Ratcliffe LM, Boag PT (2002) Female eavesdropping on male song contests in songbirds. Science 296:873–873. https://doi.org/10.1126/science.296.5569.873
Mennill DJ, Burt JM, Fristrup KM, Vehrencamp SL (2006) Accuracy of an acoustic location system for monitoring the position of duetting songbirds in tropical forest. J Acoust Soc Am 119:2832–2839. https://doi.org/10.1121/1.2184988
Moher D, Liberati A, Tetzlaff J, Altman DG, Altman D, Antes G, Atkins D, Barbour V, Barrowman N, Berlin JA, Clark J (2009) Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement (Chinese edition). Chin J Integr Med 7:889–896. https://doi.org/10.1371/journal.pmed.1000097
Moiron M, González-Lagos C, Slabbekoorn H, Sol D (2015) Singing in the city: high song frequencies are no guarantee for urban success in birds. Behav Ecol 26:843–850. https://doi.org/10.1093/beheco/arv026
Naguib M (2005) Singing interactions in songbirds: Implications for social relations. In: McGregor P (ed) Animal communication networks. Cambridge University Press, Cambridge, p 300
Naguib M, Mennill DJ (2010) The signal value of birdsong: empirical evidence suggests song overlapping is a signal. Anim Behav 80:E11–E15. https://doi.org/10.1017/CBO9780511610363.018
Narango DL, Rodewald AD (2016) Urban-associated drivers of song variation along a rural–urban gradient. Behav Ecol 27:608–616. https://doi.org/10.1093/beheco/arv197
Nolan PM, Hill GE (2004) Female choice for song characteristics in the house finch. Anim Behav 67:403–410. https://doi.org/10.1016/j.anbehav.2003.03.018
Nowicki S, Searcy WA (2005) Song and mate choice in birds: how the development of behavior helps us understand function. Auk 122:1–14. https://doi.org/10.1093/auk/122.1.1
Nowicki S, Searcy W, Hughes M (1998) The territory defense function of song in song sparrows: a test with the speaker occupation design. Behaviour 135:615–628. https://doi.org/10.1163/156853998792897888
Odom KJ, Benedict L (2018) A call to document female bird songs: applications for diverse fields. Auk Ornithol Adv 135:314–325. https://doi.org/10.1642/AUK-17-183.1
Odom KJ, Hall ML, Riebel K, Omland KE, Langmore NE (2014) Female song is widespread and ancestral in songbirds. Nat Commun 5:1–6. https://doi.org/10.1038/ncomms4379
Olinkiewicz A, Osiejuk TS (2003) Effect of time of season and neighbours on singing activity in the Corn Bunting Miliaria calandra. Acta Ornithol 38:117–122. https://doi.org/10.3161/068.038.0208
Opaev AS, Shishkina EM (2021) Song amplitude and population density in two sympatric warblers, Phylloscopus schwarzi and P. fuscatus. Bioacoustics 30:272—283. https://doi.org/10.1080/09524622.2020.1720816
Osmun AE, Mennill DJ (2011) Acoustic monitoring reveals congruent patterns of territorial singing behaviour in male and female tropical wrens. Ethology 117:385–394. https://doi.org/10.1111/j.1439-0310.2011.01887.x
Owen KC, Mennill DJ (2021) Singing in a fragmented landscape: wrens in a tropical dry forest show sex differences in the effects of neighbours, time of day, and time of year. J Ornithol 162:881–893. https://doi.org/10.1007/s10336-021-01882-0
Peake TM, Terry AMR, McGregor PK, Dabelsteen T (2002) Do great tits assess rivals by combining direct experience with information gathered by eavesdropping? Proc R Soc B Biol Sci 269:1925–1929. https://doi.org/10.1098/rspb.2002.2112
Pérez-Granados C, Traba J (2021) Estimating bird density using passive acoustic monitoring: a review of methods and suggestions for further research. Ibis 163:765–783. https://doi.org/10.1111/ibi.12944
Podos J, Lahti DC, Moseley DL (2009) Vocal performance and sensorimotor learning in songbirds. Adv Study Behav 40:159–195. https://doi.org/10.1016/S0065-3454(09)40005-6
Reid JM, Arcese P, Cassidy AL, Hiebert SM, Smith JN, Stoddard PK, Marr AB, Keller LF (2004) Song repertoire size predicts initial mating success in male song sparrows, Melospiza melodia. Anim Behav 68:1055–1063. https://doi.org/10.1016/j.anbehav.2004.07.003
Riebel K (2009) Song and female mate choice in zebra finches: a review. Adv Study Behav 40:197–238. https://doi.org/10.1016/S0065-3454(09)40006-8
Riebel K, Odom KJ, Langmore NE, Hall ML (2019) New insights from female bird song: towards an integrated approach to studying male and female communication roles. Biol Lett 15:20190059. https://doi.org/10.1098/rsbl.2019.0059
Ripmeester EA, Kok JS, van Rijssel JC, Slabbekoorn H (2010) Habitat-related birdsong divergence: a multi-level study on the influence of territory density and ambient noise in European blackbirds. Behav Ecol Sociobiol 64:409–418. https://doi.org/10.1007/s00265-009-0857-8
Searcy WA (1992) Song repertoire and mate choice in birds. Am Zool 32:71–80. https://doi.org/10.1093/icb/32.1.71
Searcy WA, Andersson M (1986) Sexual selection and the evolution of song. Annu Rev Ecol Evol Syst 17:507–533. https://doi.org/10.1146/annurev.es.17.110186.002451
Searcy WA, Beecher MD (2009) Song as an aggressive signal in songbirds. Anim Behav 78:1281–1292. https://doi.org/10.1016/j.anbehav.2009.08.011
Searcy WA, Akçay C, Nowicki S, Beecher MD (2014) Aggressive signaling in song sparrows and other songbirds. Adv Study Behav 46:89–125. https://doi.org/10.1016/B978-0-12-800286-5.00003-1
Sexton K, Murphy M, Redmond L, Dolan AC (2007) Dawn song of eastern kingbirds: intrapopulation variability and sociobiological correlates. Behaviour 144:1273–1295. https://doi.org/10.1163/156853907781890922
Sharma SPS, Dobney SL, Norris DR, Doucet SM, Newman AEM, Burant JB, Moran IG, Mueller SD, Spina HA, Woodworth B, Mennill DJ (2024). Intrinsic and extrinsic factors influence territory size and shape in Savannah Sparrows: effects of age, breeding strategy, population density, and number of neighbours. Ornithol in press
Shutler D, Weatherhead PJ (1991) Basal song rate variation in male red-winged blackbirds: sound and fury signifying nothing? Behav Ecol 2:123–132. https://doi.org/10.1093/beheco/2.2.123
Sillett TS, Rodenhouse NL, Holmes RT (2004) Experimentally reducing neighbor density affects reproduction and behavior of a migratory songbird. Ecology 85:2467–2477. https://doi.org/10.1890/03-0272
Slater PJ, Mann NI (2004) Why do the females of many bird species sing in the tropics? J Avian Biol 35:289–294. https://doi.org/10.1111/j.0908-8857.2004.03392.x
Snijders L, Naguib M (2017) Communication in animal social networks: a missing link? In: Naguib M, Podos J, Simmons LW, Barrett L, Healy SD, Zuk M (eds) Advances in the study of behavior. Elsevier Academic Press, pp 297–359. https://doi.org/10.1016/bs.asb.2017.02.004
Staicer CA, Spector DA, Horn AC (1996) The dawn chorus and other diel patterns in acoustic signaling. In: Kroodsma DE, Miller EH (eds) Ecology and evolution of acoustic communication in birds. Cornell University Press, Ithaca, pp 426–453. https://doi.org/10.7591/9781501736957-033
Stehelin TE, Ross M, Lein M (2014) Social stimulation of dawn singing in Dusky Flycatchers: a serendipitous experiment. J Field Ornithol 85:63–74. https://doi.org/10.1111/jofo.12050
Stuart CJ, Grabarczyk EE, Vonhof MJ, Gill SA (2019) Social factors, not anthropogenic noise or artificial light, influence onset of dawn singing in a common songbird. Auk 136:ukz045. https://doi.org/10.1093/auk/ukz045
Stutchbury BJ, Morton ES (2001) Behavioral ecology of tropical birds. Academic Press, San Diego
Stutchbury BJ, Morton ES (2022) Behavioral ecology of tropical birds. Academic Press, San Diego
Tobias JA, Gamarra-Toledo V, García-Olaechea D, Pulgarin PC, Seddon N (2011) Year-round resource defence and the evolution of male and female song in sub-oscine birds: social armaments are mutual ornaments. J Evol Biol 24:2118–2138. https://doi.org/10.1111/j.1420-9101.2011.02345.x
Tobias JA, Sheard C, Seddon N, Meade A, Cotton AJ, Nakagawa S (2016) Territoriality, social bonds, and the evolution of communal signaling in birds. Front Ecol Evol 4:74. https://doi.org/10.3389/fevo.2016.00074
Todt D, Naguib M (2000) Vocal interactions in birds: the use of song as a model in communication. Advances in the study of behavior, vol 29. Academic Press, pp 247–296. https://doi.org/10.1016/S0065-3454(08)60107-2
Valderrama SV, Molles LE, Waas JR (2013) Effects of population size on singing behavior of a rare duetting songbird. Conserv Biol 27:210–218. https://doi.org/10.1111/j.1523-1739.2012.01917.x
Vehrencamp SL (2001) Is song–type matching a conventional signal of aggressive intentions? Proc R Soc B Biol Sci 268:1637–1642. https://doi.org/10.1098/rspb.2001.1714
Voigt C, Leitner S, Gahr M, Ter Maat A (2021) Seasonal and diurnal variation of vocal behaviour in duetting white-browed sparrow weavers. J Ornithol 162:1163–1172. https://doi.org/10.1007/s10336-021-01905-w
Wilkins MR, Odom KJ, Benedict L, Safran RJ (2020) Analysis of female song provides insight into the evolution of sex differences in a widely studied songbird. Anim Behav 168:69–82. https://doi.org/10.1016/j.anbehav.2020.07.018
Xia C, Wei C, Lloyd H, Liu J, Wu Q, Zhang Y (2014) Dawn singing intensity of the Male Brownish-Flanked Bush Warbler: effects of territorial insertions and number of neighbors. Ethology 120:324–330. https://doi.org/10.1111/eth.12205
Acknowledgements
The authors thank Mitacs Canada, and supporting partners Birds Canada and Wildlife Acoustics, for a grant supporting NVS. The authors thank the Guanacaste Dry Forest Conservation Fund for support to NVS. The authors thank NSERC for Discovery Grant support to DJM. The authors thank Kian Lanzenstiel for translating the abstract into German. The authors thank J. Aubin, S. Dobney, and D. Zapata for comments on an early version of this manuscript, and two anonymous reviewers who provided thoughtful feedback that improved the manuscript.
Author information
Authors and Affiliations
Contributions
NVS conceived of the idea, design the study, collected the data, wrote the first version of the manuscript, and edited subsequent versions of the manuscript. DJM conceived of the idea, and wrote and edited multiple versions of the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
We declare no conflict of interest.
Additional information
Communicated by T. S. Osiejuk.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Sánchez, N.V., Mennill, D.J. Behavioural consequences of conspecific neighbours: a systematic literature review of the effects of local density on avian vocal communication. J Ornithol (2024). https://doi.org/10.1007/s10336-024-02188-7
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10336-024-02188-7