
Abstract
Temporal variation in physical activity is mainly determined by the day–night cycle. While this may be true for diurnal species whose vision at night is often poor, the situation might be more complex in nocturnal animals as many such species can see both in the dark and in the daylight. We examined in Barn Owl (Tyto alba) nestlings whether temporal variation of behavioural activities and sleep is shaped by parental feeding visits occurring during the first part of the night and the extent to which they also occur during daylight hours. We measured several behaviours in 280 individuals from 90 broods recorded in 4 years. Parental feeding visits progressively declined in frequency from the beginning to the end of the night, and a number of offspring behaviours followed the same pattern of activity (feeding, vocalization and self-preening). Surprisingly, nestlings were awake not only at sunset, but also at sunrise. Several behaviours (locomotion, wing flapping and sibling interactions, such as pecking and allopreening among nestlings) showed peaks of activity at sunset and sunrise, suggesting that they were performed for other reasons than to interact with parents. Allopreening was performed more often during the day than at night. We conclude that although adult Barn Owls are nocturnal, nestlings display a complex temporal pattern of activity that is governed not only by feeding but also by other unknown factors.
Zusammenfassung
Nächtliche, tägliche und bimodale Muster der Lokomotion, der Interaktionen mit Geschwistern und des Schlafs bei Schleiereulennestlingen
Der zeitliche Verlauf der körperlichen Aktivität wird hauptsächlich vom Tag-Nacht-Rhythmus bestimmt. Besonders tagaktive Arten sind oft nicht in der Lage nachts zu sehen, während nachtaktive Arten sowohl am Tag als auch nachts sehen können. Wir untersuchten an Schleiereulennestlingen (Tyto alba), ob zeitliche Variation von Verhalten und Gehirnaktivität von den elterlichen Besuchen in der ersten Hälfte der Nacht abhängen und ob Aktivitäten während des Tages ausgeführt werden. Wir bestimmten Verhaltensweisen von 280 Individuen aus 90 Bruten über vier Jahre. Elterliche Fütterungsbesuche nahmen über die Nacht stetig in der Frequenz ab, und einige der Verhaltensweisen der Nestlinge folgten diesem Muster (Fressen, Rufen und Putzen). Erstaunlicherweise waren Nestlinge nicht nur bei Sonnenuntergang wach, sondern auch bei Sonnenaufgang. Einige Verhaltensweisen (Lokomotion, Flügel schlagen und Interaktionen mit Geschwistern wie Picken und gegenseitiges Putzen) zeigten hohe Aktivität bei Sonnenuntergang und –aufgang, was darauf hindeutet, dass sie aus anderen Gründen ausgeführt wurden als mit den Eltern zu interagieren. Gegenseitiges Putzen wurde tagsüber öfter durchgeführt als nachts. Zusammenfassend ist zu sagen, dass Schleiereulen zwar nachtaktiv sind, Nestlinge aber ein komplexes Aktivitätsmuster zeigen, das durch die elterliche Fütterung vorgegeben ist, aber auch durch weitere, noch unbekannte Faktoren bestimmt ist.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Specific behaviours should be expressed at the appropriate time of the day or night to maximize the associated benefits while minimizing the costs. An animal’s activity pattern can be shaped by environmental factors, such as food availability, temperature and light intensity, or by the social environment (Vasquez 1996; Elvert et al. 1999; Mrosovsky 1999; Boydston et al. 2001). For example, in teleost fishes, rain improves nocturnal foraging efficiency, which in turn induces them to switch their activity period from diurnal to nocturnal (Payne et al. 2012). Predation risk can also limit the activity of prey species to the periods with the lowest risk of predation. Three-toed sloths (Bradypus variegatus) exposed to nocturnal predators preferentially sleep at night, whereas the pygmy sloth (Bradypus pygmaeus) living on an island without any predator shows no preference for sleeping during the night or day (Voirin et al. 2014).
Although numerous studies demonstrate the influence of environmental and species-specific adaptation on activity pattern in adults (Hut and Beersma 2011), few studies have examined the activity and sleep–wakefulness patterns during development. Studies from a wide range of taxa are required to determine the major underlying factors that influence temporal variation in sleep–wakefulness states and locomotor activity (e.g. Scriba et al. 2013b). Nocturnal species are interesting because their visual systems allow for active behaviour during both the day and night (Hall and Ross 2007; Hall 2008). Consequently, nocturnal animals may be more flexible in their timing of activity than diurnal species (Hoogenboom et al. 1984; Levy et al. 2007; Daan et al. 2011).
We have been studying whether Barn Owl (Tyto alba) nestlings are primarily active at night, like their parents (Erkert 1969; Scriba et al. 2013a), or throughout both the day and night. Although nocturnal provisioning by the parents might favour nocturnal activity in nestlings, the storage of food in the nest may free nestlings to also feed during the daylight hours (Roulin 2004). Furthermore, nestlings experience various environmental and social influences, such as environmental disturbances during the day and interactions with siblings.
We thus designed a study in which we recorded a number of behaviours in different individuals monitored in 2007, 2011, 2012 and 2014. These behaviours included the timing of nestling cerebral wakefulness and sleep, parental food provisioning, nestling feeding, preening of themselves and of their siblings (i.e. allopreening), locomotion, wing flapping, vocalization and competitive behaviour (pecking). We assessed these activities in the field and also in the laboratory in the absence of the parents. We predicted that some activities, such as feeding and vocal competition for food, would follow the nocturnal timing of the parental visits. This nocturnal pattern may however shift to bimodality if there is some benefit in performing activities at different times of day (e.g., exposure to different cues, for improved sensory development or having many bouts of sleep for cognitive development). As a consequence, nestlings might be highly active shortly after sunset and around sunrise. In rats, the rhythmic clock gene expression important for a circadian rhythm develops gradually during the first weeks after birth (Vallone et al. 2007). In young quail, a circadian rhythm for feeding activity first occurs with 4–6 weeks of age (Formanek et al. 2011). These findings raise the interesting possibility that nestlings might not yet display a circadian rhythm in various behaviours at this stage of their development as they live in a non-rhythmic or slightly rhythmic environment (e.g. Kouba et al. 2014). Digestive constraints might also lead to cyclic behaviour. When the owlet’s stomach is full, it may be best to invest in sleep-related functions. In particular, sleep is an important state for learning, memory, the immune system, energy homeostasis and brain maintenance (Vyazovskiy et al. 2008; Markwald et al. 2013; Rasch and Born 2013; Xie et al. 2013; Opp and Krueger 2015), and sleep deprivation negatively impacts physiology and cognition (Van Dongen et al. 2003). Hence, nestlings are likely to be awake only during some parts of the nights. Superimposed on this expected nocturnal or bimodal activity pattern, nestlings may also have an endogenous need for sleep at regular short intervals, since developing animals spend more time asleep than adults (e.g. Roffwarg et al. 1966). Additionally, interactions between siblings are important before the night, as nestlings show reciprocation between allopreening taking place during the day and food-sharing at night (Roulin et al. 2016). Nestling Barn Owls vocally negotiate the priority of access to the impending food, a process that takes time and hence is already expressed during the day (Dreiss et al. 2016a). The use of prosocial behaviour allows animals to reduce the cost of competition, but this activity is commonly time consuming (Lewis et al. 2007), implying that such behaviours may have to be expressed not only at night but also during the day. Furthermore, nestlings can feed on stored food at any time of the day or night to optimize energy intake (Roulin 2004). Hence, some behaviour directed towards siblings, such as allopreening and pecking, may show a more even temporal distribution. Answering all of these questions is a huge undertaking, and a first approach to tackle this research agenda is to perform a descriptive study that describes the temporal pattern of activity in many behaviours. This was the goal of the study reported here.
Methods
Study system
We studied a population of Barn Owls breeding in nest boxes (62 × 56 × 37 cm) fixed to the external wall of barns in western Switzerland (46°49′N, 06°56′E). Hence, nestlings were exposed to the light–dark cycle through the nest box entrance hole (13 × 20 cm), and they could hear sounds in the environment from a variety of external sources, including humans. We also studied nestlings in the laboratory in similarly sized, ventilated nest boxes with a weak light source (LEDs, 1 W), switched on during natural daytime. Nestlings in the laboratory were therefore also exposed to the light–dark cycle, as in nature. We recorded behaviour and electroencephalograms (EEG) in nestlings at about 4 weeks of age when they were able to thermoregulate and eat without maternal help (we regularly visited nests to determine hatching dates of all siblings after measuring wing length a few days after hatching; Roulin 2004). At this time, the parents were not sleeping in the nest box, but somewhere close by during the daytime. For each behaviour, we calculated the mean hourly values 4 h before and after sunset and sunrise, respectively; specific details including sample sizes and behavioural definitions are provided in the following sections for each of the different aspects of this study. We recorded nestlings between May and October, during which time day length varies by about 4 h. Therefore, we examined locomotion, sleep and behavioural pattern in relation to sunset and sunrise rather than the exact time of the day or night. For each 1-h interval around sunset and sunrise (up to 4 h before and after these specific time points), we recorded behaviours and cerebral activity to evaluate temporal variation in relation to natural light conditions to accommodate seasonal changes in night length. We chose this methodology, because we predicted that nestling behaviour would be sensitive to the day–night cycle. Indeed, Barn Owl parents bring the first prey item of the night at around 2230 hours in June but at 1830 hours in October (personal observation).
Sleep and locomotor activity in the field in 2011
The Barn Owl, like other birds and mammals, exhibits two sleep states, namely REM (rapid eye movement) and non-REM sleep, which alternate in short intervals and can be recorded with EEG (Rattenborg et al. 2011). Between May and October 2011 we recorded the brain activity of nestlings continuously for about 5 days using minimally invasive subcutaneous electrodes. This was done in the field in 29 broods involving 31 male and 35 female nestlings, aged 27–48 days [mean ± standard deviation (SD) 38.2 ± 0.6 days]. Using Somnologica software (Medcare, Embla Systems Europe, Amsterdam, The Netherlands), a single person analysed the last 24-h period of the recordings. Only one 24 h-period was analysed in order to exclude the periods during which owlets habituated to the recording device. The EEG signals were scored for wakefulness, REM and non-REM sleep in 4-s epochs, and epochs containing more than one state were scored according to the predominant state. Wakefulness was characterized by low-amplitude, high-frequency EEG activity, but it was often also accompanied by movement artefacts characterized by high frequency and amplitude activity when the birds were engaged in active behaviour. Non-REM sleep consisted of low-frequency, high-amplitude EEG activity. REM sleep was characterized by low-amplitude, fast-frequency activity, accompanied by head dropping or swaying of the body detected via accelerometer recordings (for more details, see Scriba et al. 2013a, b; 2014). We calculated the fraction of time per hour spent awake, in non-REM and REM sleep, respectively. The recording equipment, which weighed at most 2.1% of the bird’s body weight, did not seem to have an adverse effect on the nestlings, as all of the birds fledged, and recruitment into the breeding population in the following year was actually higher in those Barn Owl nestlings which had their sleep recorded (26.3%) than in those that had not (19.0%; Scriba et al. 2013a, b; 2014). Furthermore, we installed the device on the nestlings during daylight hours, and they usually went asleep a few minutes after we completed the procedure, demonstrating that our method allowed us to record brain activities without artefacts. We found that the temporal variation in REM and non-REM sleep was very similar (Electronic Supplementary Material Fig. 1), because REM sleep mostly takes place after a bout of non-REM sleep; hence we report here only temporal variation in wakefulness (we nevertheless separately examined whether REM and non-REM sleep are more often performed at day than night). We did not examine how the number and duration of wakefulness–sleep bouts vary over a 24-h period, because this is closely associated with the percentage of time spent asleep. However, we did examine whether the number and duration of wakefulness and REM and non-REM sleep bouts differ between the light and dark phase.
To record locomotor activity, a neurologger (Neurologger 2A; Vyssotski et al. 2009, http://www.vyssotski.ch/neurologger2) equipped with an accelerometer to monitor locomotor activity at 200 Hz was placed on the head of the nestling (Scriba et al. 2013a; Anisimov et al. 2014). The logger electronics was complemented with a board carrying a three-dimensional (3D) accelerometer chip (LIS302DLH; STMicroelectronics Inc., Calamba, Laguna, Philippines) and communication microcontroller PIC18LF13K22 (Microchip Technology Inc., Chandler, AZ). Acceleration in each direction was measured in the range ±2 g and stored as an 8-bit value. Acceleration data were analysed in R (version 2.15.2; R Core Team 2012) using custom written scripts. To estimate the amount of body movement, the absolute value of the 3D acceleration vector was computed, and the variance of this vector was taken as an estimate of locomotor activity. We obtained activity data for 64 of the 66 nestlings for the same period of time during which we recorded their EEG.
Feeding, allopreening, pecking and wing flapping in the field in 2007
Between May and September 2007, we artificially reduced 21 broods to three nestlings to better score nestling behaviour of 49 individuals for which we successfully obtained data from the video footage. The other nestlings were kept in ventilated boxes (with food) at some distance from the nests. The older nestlings of these three-chick broods were on average 42 days old (range 35–50 days), the middle-born individuals 38 (range 28–47) days and the juniors 34 (range 21–44) days. We used two infrared-sensitive video cameras per nest box to film under dark conditions from 1900 to 0700 hours. All of the video recordings were analysed by the same observer, who was blind to the study question. For each hour, this observer counted the number of times nestlings were pecking and allopreening each other. Pecking is an agonistic behaviour characterized by one individual hitting another one with its beak, while allopreening involves one individual using its beak to preen its sibling. The observer also scored for each hour the number of times nestlings were flapping their wings. Finally, the observer recorded the time when parents were bringing food to the nest and when nestlings were eating.
Vocal negotiation in the field in 2014
Barn Owl nestlings communicate vocally during the night while parents are foraging with the purpose to negotiate which individual will have priority access to the next delivered prey item (Roulin 2001). Between June and August 2014 we installed microphones inside of ten nest boxes containing on average 5.7 (range 4–9) nestlings with a mean age of 36.9 (range 20–45) days. We counted the so-called “negotiation calls” of the entire broods (calls could not be assigned to specific individuals) using Matlab (R2012b 8.0.0.783; MathWorks, Natick, MA). For each 1-h interval around sunset and sunrise, we divided the total number of calls by the number of nestlings. Calls were recorded from 2000 or 2200 hours to 0400 or 0700 hours during the dark period, explaining why we could not always count calls up to 4 h before sunset and 4 h after sunrise.
Locomotor activity, self-preening, allopreening, vocal negotiation and feeding in the laboratory in 2012
Between May and August 2012, we brought 116 nestlings (average age 30.6 days; range 16–39 days) from 30 broods to nest boxes in the laboratory. These boxes were soundproofed so that nestlings placed in one box could not hear other individuals placed in neighbouring boxes. From 1400 hours to 1300 hours the following day, we recorded their behaviour using video cameras (TVCCD-150SET; Monacor International GmbH and Co. KG, Bremen, Germany) and microphones (MC930; Beyerdynamic GmbH and Co. KG, Heilbronn, Germany). Each nestling was recorded alone for one 23-h period and with another sibling during the next 23-h period (or the other way round, with the order randomized). Nestlings are known to produce negotiation calls also when they are alone in the nest box.
We recorded the number of negotiation calls per nestling in the same way as described in section "Vocal negotiation in the field in 2014". Locomotor and feeding behaviours were assessed in a randomly chosen subsample of 44 nestlings from 13 broods, and self-preening and allopreening were assessed in 79 randomly chosen nestlings from 20 broods and placed alone (to measure self-preening) or in pairs (to measure self-preening and allopreening). We considered a subsample because measuring these behaviours during a 23-h period cannot be done automatically but manually, which is very time-intense. Nestlings were fed ad libitum at 1400 hours so that we could monitor the time when they consumed food. Video images of boxes were divided in 5 × 4 squares to measure locomotor activity. The video recording was split in 1-min episodes, and nestlings were considered to be active if they moved from one square to another. Allopreening was measured as explained in section "Feeding, allopreening, pecking and wing flapping in the field in 2007". An individual was considered to be self-preening when it touched its feathers with its bill or scratched its body with its feet. We measured the amount of time nestlings were self-preening and, for each hour, we calculated the proportion of time nestlings were self-preening.
Temporal variation in the frequency of allopreening was very similar in 2012 in the laboratory as in 2007 in the wild (data not shown). Therefore, for each year we standardized the data [(value − mean)/SD] and combined the 2 years of data to calculate the mean standardized frequency of allopreening for each hour in relation to sunset and sunrise.
Statistical analyses
The time of sunset and sunrise had been obtained from the Astronomical Almanac by the H.M. Nautical Almanac Office in the UK and the U.S. Naval Observatory. We calculated the frequency of each behaviour in relation to time to sunset (i.e. 4, 3, 2 or 1 h before sunset; during the hour comprising sunset; and 1, 2, 3 and 4 h after sunset). A similar procedure was applied to sunrise. We used this approach (i.e. calculate mean hourly values) and standardized the data [i.e. (value − mean)/SD] to compare all behaviours between each other and in relation to sunset and sunrise. The data were normally distributed, and hence we used parametric paired t tests to determine if behaviours differed between light and dark periods, using average values per brood. Note that, with the exception for sleep and locomotor activity in nature, we did not record behaviours during the entire dark–light period. We therefore compared the mean values of light and dark periods during the studied period only. We performed non-parametric Spearman’s correlations when samples sizes were small. All P values are two-tailed, and P < 0.05 was considered to be significant. Statistical analyses were performed with the JMP software version 11.0. Using the Hartigan’s dip test statistic for unimodality (package ‘diptest’; Maechler 2012; version 0.75-74, in R), we tested if the distribution of each behaviour (using the recorded 16-h periods) departs from unimodality.
Results
Comparison between diurnal and nocturnal behaviour
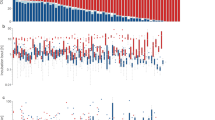
Parental feeding visits, nestling feeding, calling, locomotion and self-preening behaviours were significantly more often performed at night than during the day. Cerebral wakefulness, wing flapping, pecking and REM sleep were observed as often at night as during the daylight hours. Only allopreening in the field and non-REM sleep were significantly more frequent during the day than at night (Fig. 1).

Extent to which behaviour and cerebral activity are nocturnal vs. diurnal in Barn Owl (Tyto alba) nestlings. Bars represent brood average values for the daylight and dark periods divided by the daylight and dark average, respectively, to obtain a percentage. *, **, *** Difference (according to paired t test on mean values per brood) is significant at P < 0.05, P < 0.001 and P < 0.0001, respectively. n.s. Non-significant
Temporal variation in nocturnal behaviours
In the field, feeding activities were synchronized with the parental visits (parents brought food to the nest). There was a strong correlation between the mean hourly parental feeding visits and the number of prey items offspring consumed at night (Spearman’s correlation r s = 0.88, n = 9 night periods of 1 h, P = 0.002; Fig. 2). Parental feeding visits and offspring feeding mainly started 1 hour after sunset (Fig. 2) and progressively declined throughout the night (parents: r s = −0.69, n = 9, P = 0.038; offspring: r s = −0.83, n = 9, P = 0.005). A similar pattern of nestling feeding activity was observed in the laboratory, although it was significant only in the situation when nestlings were placed alone in a nest box for 23 h (r s = −0.67, n = 9, P = 0.049; Fig. 2) but not when placed in a nest box in pairs (r s = −0.12, n = 9, P = 0.77). Not surprisingly, the temporal pattern of nestling vocal negotiation followed the same trend, with high vocal activity at the beginning of the night followed by a progressive decline over the night (r s = −0.83, n = 9, P = 0.005; Fig. 3). Finally, self-preening activities followed a similar temporal pattern at night (r s = −0.93, n = 9, P = 0.002; Fig. 4).

Parental feeding visits and nestling feeding activities in the Barn Owl. Data points are mean standardized hourly values [whiskers standard error (SE)] according to time to sunset and time after sunrise as indicated by 0 (i.e. up to 4 h before and 4 h after each of these time points, respectively). Shaded area dark period. Data on feeding in nestlings were collected in 21 three-chick broods in the field in 2007 and in the laboratory in 44 nestlings from 13 broods in 2012. The data were standardized for each year, and mean values are presented. Data on parental food provisioning in the field were collected in the same 21 three-chick broods in 2007 (note that we fed the nestlings in the laboratory therefore cannot provide these data for the laboratory)

Vocal negotiation in Barn Owl nestlings. Data points are mean standardized hourly number of calls per nestling (whiskers SE) according to sunrise and sunset as indicated by 0 (i.e. up to 4 h before and 4 h after each of these time points, respectively). Shaded area dark period. Negotiation calls were recorded in the field in 2014 in 10 broods and in the laboratory in 2012 in 116 nestlings from 30 broods placed alone (‘solo’) or in pairs (‘duo’) in a nest box during a 23-h period (for each individual a mean value of the two recorded values—solo and duo—was calculated). Values obtained in the field and in the laboratory were separately standardized before being averaged and shown in this figure. Different symbols (squares, circle, triangles) indicate that values are significantly different from each other using paired t test analyses

Self-preening in Barn Owl nestlings in the laboratory. Data points are mean hourly values (whiskers SE) of self-preening events according to sunrise and sunset as indicated by 0 (i.e. up to 4 h before and 4 h after each of these time points, respectively). Shaded area dark period. Self-preening was recorded in the laboratory in 2012 in 79 nestlings placed in pairs or alone in the nest box. For each individual, a mean value was calculated
Do behaviours show a bimodal pattern of nocturnal activity?
According to the Hartigan’s dip test statistic, allopreening, pecking, wing flapping, feeding, self-preening, activity and wakefulness were weakly bimodally distributed (HDS, P values between 0.06 and 0.09), whereas parental visits and calls followed an unimodal distribution (P > 0.13).
In the field, nestlings slept mainly during the daylight hours and in the middle of the night until sunrise when there was a sudden and strong peak of cerebral wakefulness (filled symbols in Fig. 5a). Another, less pronounced peak of wakefulness took place during the first 3 h after sunset (Fig. 5a). Locomotor activity showed a similar temporal pattern when measured in the same individuals (Fig. 5a) and in another set of individuals in the laboratory (Fig. 5b). Although the mean hourly values of the frequency of nestlings flapping their wings, pecking each other or allopreening were not significantly different from each other (paired t tests, P > 0.05), there were two peaks of maximal activity at sunset and sunrise, respectively (Figs. 6, 7).

Sleep-wakefulness and locomotor activity in Barn Owl nestlings in nature recorded in 2011 (a) and in the laboratory recorded in 2012 (b). Data points are mean hourly standardized values (whiskers SE) according to sunrise and sunset as indicated by 0 (i.e. up to 4 h before and 4 h after each of these time points, respectively). Shaded area dark period. a Data were collected on 66 nestlings from 29 broods in the field in 2011. b Locomotor activity was recorded in the laboratory in 44 nestlings in 2012. Different symbols (triangles, diamonds, squares and circles) indicate whether within individuals the amount of time spent awake or physically active was sequentially significant according to paired t test analyses

Wing flapping and pecking in Barn Owl nestlings in nature in relation to sunrise and sunset. Data points on wing flapping (open diamonds) and pecking (filled circles) are given as mean standardized hourly values (whiskers SE) according to sunrise and sunset as indicated by 0 (i.e. up to 4 h before and 4 h after each of these time points, respectively). Shaded area dark period. Mean hourly values were not significantly different from each other according to the paired t test. Data are from 21 three-chick broods in 2007

Allopreening in Barn Owl nestlings. Data points (whiskers SE) are given as mean hourly values according to sunrise and sunset as indicated by 0 (i.e. up to 4 h before and 4 h after each of these time points, respectively). Shaded area dark period. Data were collected in the field in 21 three-chick broods in 2007 and in the laboratory in 20 two-chick broods in 2012. Because the pattern of temporal variation was globally similar in 2007 as in 2012, we standardized the data collected in each year and then calculated a mean overall value. Mean hourly values were not significantly different from each other according to the paired t tests
Discussion
Studying temporal variation in locomotor activity and sleep in young birds is particularly interesting in the context of examining whether activity is mainly governed by feeding or whether it is also sensitive to social interactions among family members. When sibling competition is mainly dictated by size-related dominance, social interactions may be relatively simple and take place mainly when parents bring food to the nest. In systems where young siblings can behave altruistically or cooperatively to obtain food, interactions may be more complex because prosocial interactions may take time and involve multiple behaviours, such as allopreening, allofeeding, pecking and vocal negotiation. For these reasons, the temporal pattern of behaviours may not always follow the pattern of feeding activities, and hence individuals may not show circadian rhythms, but rather complex temporal patterns of activity. This is an interesting aspect to study, but, to the best of our knowledge, we are not aware of any other similar study performed in other animals, thus preventing us from comparing our findings with other systems.
The adult Barn Owl is mainly nocturnal (Scriba et al. 2013b), and in our study parents were not sleeping in their nest during daylight hours but at some distance from the nest. However, our results show that the temporal variation in nestling behaviour is more complex. Nestlings are not strictly nocturnal, with some behaviour being displayed as often during the daylight hours as at night (wing flapping, pecking; Fig. 1) and some behaviour (locomotor activity, wing flapping, pecking and allopreening; Figs. 5, 6 and 7) showing a bimodal pattern of activity with a first peak at sunset and a second peak at sunrise. Our recordings revealed that allopreening was slightly more frequent during the day (Fig. 1), at least when measured in the field. The activity patterns show a second peak in nestling activity at sunrise that did not coincide with feeding activities, which were low at this time (Fig. 2).
Bimodal pattern
Nestlings behaved differently at different times of the 24-h period. As expected, nestlings were physically active around sunset—but, rather surprisingly, they were as active around sunrise. These two peaks in activity apply, for example, to locomotion, wing flapping and to sibling interactions (pecking and allopreening). We can only speculate on the potential reasons why nestlings are active around sunrise. Bimodal activity patterns are common in animals and can be a strategy to reduce daily energy expenditure (Erkert and Kappeler 2004). Alternatively, nestlings may move and flap their wings to warm up in the cold early morning. Movements performed at sunset may also correspond to maintenance behaviours, which are less important at night when the owlets compete for food. Wing flapping is an important maintenance behaviour as nestlings need to adjust their body mass to reach aerodynamically appropriate wing loadings at fledging (Wright et al. 2006). Accordingly, in our study the propensity to flap wings increased with age, as also observed in Tengmalm’s Owls (Aegolius funereus) (Kouba et al. 2014). Additionally, nestlings might be active at sunrise to find a place in their nest where they will rest and huddle during the day (Dreiss et al. 2016b). Because huddling implies close contact between siblings, they may engage in diverse social interactions, including pecking and allopreening, the latter behaviour being known to reduce social stress and conflicts (Lewis et al. 2007; Fraser et al. 2008; Fraser and Bugnyar 2011).
These temporal patterns of activity indicate that in nestlings, at least in the range of age considered in the present study (16–50 days), nocturnality has not yet been established. A similar result was found in Tengmalm’s Owl nestlings which were spending time at the nest box entrance not only during night when the parents came with food, but also during daylight (Kouba et al. 2014). This daytime behaviour might occur as a preparation for fledging, as we found that older nestlings spent more time at the entrance or in the alert state if a predator was trying to enter the nest. Additionally, the cost for being active at any time of day or night might be low for the nestlings. From the data obtained in this study, we cannot draw any conclusion on circadian rhythmicity; for this, recordings of the behavioral pattern are needed over several 24-h periods.
Daylight activities
During the daylight hours (excluding the period around sunset) nestlings were mostly physically inactive and engaged in more and longer sleep bouts. Nonetheless, nestlings did not sleep continuously during the light phase, and the duration of time spent awake was the same during the day and night. It is possible that the nestlings are disturbed by human activity during the day since Barn Owls breed in nest boxes placed in barns. This interpretation is consistent with the finding that nestlings are physiologically more stressed, as measured by blood circulating corticosterone, when located closer to inhabited houses and livestock (Almasi et al. 2015). Alternatively, some behaviours may be realized during the day for adaptive reasons, which would explain the time spent awake in the day. During the light period nestlings sometimes eat prey remains (Roulin 2004), as also observed in the present study (Fig. 2). The duration of digestion might prevent individuals from ingesting the daily food intake within a short time interval at night, since the egestion of a pellet takes about 6.5–10 h after feeding (Smith and Richmond 1972), which might explain why nestlings sometimes feed during the day. Furthermore, social interactions, such as allopreening and pecking, occurred during the light period, especially before sunset. Nestlings may anticipate parental visits with prosocial (allopreening) and competitive (pecking) behaviours, to induce siblings to later share food. As we recently showed, reciprocation does occur between food-sharing and allopreening in owl nestlings. An individual that preens its sibling during daylight hours is more likely to be later fed by this sibling. These behaviours may reduce the level of sibling competition (Roulin et al. 2016).
Locomotor and vocal activities resume around sunset, before the first parental feeding visit. Nestlings may be active before the sun goes down to position themselves closer to the nest box entrance where the likelihood of being fed by parents is higher (Dreiss et al. 2013a, b; Kouba et al. 2014). Siblings vocally negotiate priority access to the impending food items. In the Barn Owl, parents bring a single non-divisible food item per visit that is consumed by a single offspring. Each individual eats three to four items per night, and there is a pronounced asymmetry in food need between the differently aged siblings. Indeed, as soon as one individual has consumed a food item, it is much less hungry than its siblings. To reduce the level of competition, siblings communicate vocally, a process referred to as “sibling negotiation”. Because such a process takes a long time (Dreiss et al. 2015), nestlings start to vocally negotiate long before the first parental feeding visit explaining, in part, why they can be active during the daylight hours.
Ethics statement
All experiments were performed under the legal authorization of the Veterinary Office (Vaud Canton, Switzerland). Nestlings recorded in the laboratory were not physiologically stressed, as shown by the absence of a rise in baseline corticosterone levels compared to the situation prevailing under natural, undisturbed conditions (Dreiss et al. 2010). Keeping owlets at the university for a few days did not negatively affect their body condition, since mean body mass and survival at fledgling did not differ between experimental nestlings and nestlings remaining in their nest during all rearing period (Dreiss et al. 2013a, b). The minimally invasive EEG recording method did not appear to have any long-term adverse effects on the Barn Owl nestlings, as all nestlings in which the brain activity had been recorded fledged, and the number of owls coming back for breeding in the following year was even higher in owls in which we recorded sleep (26.3%), than in those without sleep recording (19.0%). Also the owls habituated to the electrodes and data logger quickly within a few hours and behaved normally afterwards (as confirmed by video recordings), and nestlings were sleeping as early as 5–10 min after being placed back into the nest box after electrode placement.
References
Almasi B, Béziers P, Roulin A, Jenni L (2015) Agricultural land-use and human presence around breeding sites increase stress-hormone levels and decrease body mass in barn owl nestlings. Oecologia 179(1):89–101
Anisimov VN, Herbst JA, Abramchuk AN, Latanov AV, Hahnloser RH, Vyssotski AL (2014) Reconstruction of vocal interactions in a group of small songbirds. Nat Methods 11(11):1135–1137
Boydston EE, Morelli TL, Holekamp KE (2001) Sex differences in territorial behavior exhibited by the spotted hyena (Hyaenidae, Crocuta crocuta). Ethol 107(5):369–385
Core Team R (2012) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Daan S, Spoelstra K, Albrecht U, Schmutz I, Daan M, Daan B, Riensk F, Poletaeva I, Dell’Omo G, Vyssotski A, Lipp H-P (2011) Lab mice in the field: unexpected activity patterns and effects of a dysfunctional mPer2 allele. J Biol Rhythms 26(2):118–129. doi:10.1177/0748730410397645
Dreiss AN, Henry I, Ruppli CA, Almasi B, Roulin A (2010) Darker eumelanic barn owls better withstand food depletion through resistance to food deprivation and lower appetite. Oecologia 164:65–71
Dreiss AN, Calcagno M, Van den Brink V, Laurent A, Almasi B, Jenni L, Roulin A (2013a) The vigilance components of begging and sibling competition. J Avian Biol 44(4):359–368
Dreiss AN, Ruppli CA, Oberli F, Antoniazza S, Henry I, Roulin A (2013b) Barn owls do not interrupt their siblings. Anim Behav 86:119–126
Dreiss AN, Ruppli CA, Faller C, Roulin A (2015) Social rules govern vocal competition in the barn owl. Anim Behav 102:95–107
Dreiss AN, Gaime F, Delarbre A, Moroni L, Lenarth M, Roulin A (2016a) Vocal communication regulates sibling competition over food stock. Behav Ecol Sociobiol 70:927–937
Dreiss AN, Séchaud R, Béziers P, Villain N, Genoud M, Almasi B, Jenni L, Roulin A (2016b) Social huddling and physiological thermoregulation are related to melanism in the nocturnal barn owl. Oecologia 180(2):371–381
Elvert R, Kronfeld N, Dayan T, Haim A, Zisapel N, Heldmaier G (1999) Telemetric field studies of body temperature and activity rhythms of Acomys russatus and A. cahirinus in the Judean Desert of Israel. Oecologia 119(4):484–492
Erkert HG (1969) Die Bedeutung des Lichtsinnes für Aktivität und Raumorientierung der Schleiereule (Tyto alba guttata Brehm). Z Vergl Physiol 64(1):37–70
Erkert HG, Kappeler PM (2004) Arrived in the light: diel and seasonal activity patterns in wild Verreaux’s sifakas (Propithecus v. verreauxi; Primates: Indriidae). Behav Ecol Sociobiol 57(2):174–186
Formanek L, Richard-Yris MA, Houdelier C, Lumineau S (2011) Rhythmic birds show a better social integration than arrhythmic birds. Chronobiol Internat 28(1):48–57
Fraser ON, Bugnyar T (2011) Ravens reconcile after aggressive conflicts with valuable partners. PLoS One 6(3):e18118. doi:10.1371/journal.pone.0018118
Fraser ON, Stahl D, Aureli F (2008) Stress reduction through consolation in chimpanzees. Proc Nat Acad Sci USA105(25):8557–8562
Hall MI (2008) Comparative analysis of the size and shape of the lizard eye. J Zool 111:62–75. doi:10.1016/j.zool.2007.04.003
Hall MI, Ross CF (2007) Eye shape and activity pattern in birds. J Zool 271:437–444. doi:10.1111/j.1469-7998.2006.00227.x
Hoogenboom I, Daan S, Dallinga H, Schoenmakers M (1984) Seasonal change in daily timing of behaviour in the common vole Microtus arvalis. Oecologia 61:18–31
Hut RA, Beersma DG (2011) Evolution of time-keeping mechanisms: early emergence and adaptation to photoperiod. Phil Trans R Soc B 366(1574):2141–2154
Kouba M, Bartoš L, Zárybnická M (2014) Perching of Tengmalm’s Owl (Aegolius funereus) nestlings at the nest box entrance: effect of time of the day, age, wing length and body weight. PLoS One 9(5):e97504. doi:10.1371/journal.pone.0097504
Levy O, Dayan T, Kronfeld-Schor N (2007) The relationship between the golden spiny mouse circadian system and its diurnal activity: an experimental field enclosures and laboratory study. Chronobiol Int 24:599–613
Lewis S, Roberts G, Harris MP, Prigmore C, Wanless S (2007) Fitness increases with partner and neighbour allopreening. Biol Lett 3(4):386–389
Maechler M (2012) Diptest: Hartigan’s dip test statistic for unimodality—corrected code. R package version 0.75-74. http://CRAN.R-project.org/package=diptest. Accessed 24 Sept 2016
Markwald RR, Melanson EL, Smith MR, Higgins J, Perreault L, Eckel RH, Wright KP (2013) Impact of insufficient sleep on total daily energy expenditure, food intake, and weight gain. Proc Nat Acad Sci USA 110(14):5695–5700
Mrosovsky N (1999) Further experiments on the relationship between the period of circadian rhythms and locomotor activity levels in hamsters. Physiol Behav 66(5):797–801
Opp MR, Krueger JM (2015) Sleep and immunity: a growing field with clinical impact. Brain Behav Immun 47:1
Payne NL, van der Meulen DE, Gannon R, Semmens JM, Suthers IM, Gray CA, Taylor MD (2012) Rain reverses diel activity rhythms in an estuarine teleost. Proc R Soc B 280:20122363. doi:10.1098/rspb.2012.2363
Rasch B, Born J (2013) About sleep’s role in memory. Physiol Rev 93(2):681–766
Rattenborg NC, Martinez-Gonzalez D, Roth TC 2nd, Pravosudov VV (2011) Hippocampal memory consolidation during sleep: a comparison of mammals and birds. Biol Rev 86:658–691
Roffwarg HP, Muzio JN, Dement WC (1966) Ontogenetic development of the human sleep-dream cycle. Science 152:604–619
Roulin A (2001) Food supply differentially affects sibling negotiation and competition in the barn owl (Tyto alba). Behav Ecol Sociobiol 49:514–519
Roulin A (2004) The function of food stores in bird nests: observations and experiments in the Barn Owl Tyto alba. Ardea 92:69–78
Roulin A, Des Monstiers B, Ifrid E, Da Silva A, Genzoni E, Dreiss AN (2016) Reciprocal preening and food sharing in colour-polymorphic nestling Barn Owls. J Evol Biol 29:380–394
Scriba MF, Ducrest AL, Henry I, Vyssotski AL, Rattenborg NC, Roulin A (2013a) Linking melanism to brain development: expression of a melanism-related gene in barn owl feather follicles covaries with sleep ontogeny. Front Zool 10:42. doi:10.1186/1742-9994-10-42
Scriba MF, Harmening WM, Mettke-Hofmann C, Vyssotski AL, Roulin A, Wagner H, Rattenborg NC (2013b) Evaluation of two minimally invasive techniques for electroencephalogram recording in wild or freely behaving animals. J Comp Physiol A 199(3):183–189. doi:10.1007/s00359-012-0779-1
Scriba MF, Rattenborg NC, Dreiss A, Vyssotski AL, Roulin A (2014) Sleep and vigilance linked to melanism in wild Barn Owls. J Evol Biol 27(10):2057–2068. doi:10.1111/jeb.12450
Smith CR, Richmond ME (1972) Factors influencing pellet egestion and gastric pH in the Barn Owl. Wilson Bull 84(2):179–186
Vallone D, Lahiri K, Dickmeis T, Foulkes NS (2007) Start the clock! Circadian rhythms and development. Dev Dynam 236(1):142–155. doi:10.1002/dvdy.20998
Van Dongen HP, Maislin G, Mullington JM, Dinges DF (2003) The cumulative cost of additional wakefulness: dose–response effects on neurobehavioral functions and sleep physiology from chronic sleep restriction and total sleep deprivation. Sleep 26:117–126
Vasquez RA (1996) Patch utilization by three species of Chilean rodents differing in body size and mode of locomotion. Ecol 77(8):2343–2351
Voirin B, Scriba MF, Martinez-Gonzalez D, Vyssotski AL, Wikelski M, Rattenborg NC (2014) Ecology and neurophysiology of sleep in two wild sloth species. Sleep 37(4):753. doi:10.5665/sleep.3584
Vyazovskiy VV, Cirelli C, Pfister-Genskow M, Faraguna U, Tononi G (2008) Molecular and electrophysiological evidence for net synaptic potentiation in wake and depression in sleep. Nat Neurosci 11:200–208
Vyssotski AL, Dell’Omo G, Dell’Ariccia G, Abramchuk AN, Serkov AN, Latanov AV, Loizzo A, Wolfer DP, Lipp HP (2009) EEG responses to visual landmarks in flying pigeons. Curr Biol 19(14):1159–1166
Wright J, Markman S, Denney SM (2006) Facultative adjustment of pre-fledging mass loss by nestling swifts preparing for flight. Proc R Soc B 273:1895–1900. doi:10.1098/rspb.2006.3533
Xie L, Kang H, Xu Q, Chen MJ, Liao Y, Thiyagarajan M, O’Donnell J, Christensen DJ, Nicholson C, Iliff JJ, Takano T (2013) Sleep drives metabolite clearance from the adult brain. Science 342(6156):373–377
Acknowledgements
The research was supported by the Swiss National Science Foundation, the Société Académique Vaudoise and the Max Planck Society. The experiments comply with the current laws of Switzerland.
Author information
Authors and Affiliations
Contributions
MFS, AD, IH, PB, CR, PD collected data. ALV contributed the neurologger. EI, ADS, BDM analysed video recordings. MFS, AD, NCR, AR analyzed data. MFS, NCR, AR wrote the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All experiments were performed under the legal authorization of the Veterinary Office (Vaud Canton, Switzerland). All procedures performed in the studies were in accordance with the ethical standards of the institution at which these studies were conducted.
Additional information
Communicated by F. Bairlein.
Electronic supplementary material
Below is the link to the electronic supplementary material.
10336_2017_1458_MOESM1_ESM.docx
Supplementary Fig. 1 Sleep-wakefulness in the field in relation to sunrise and sunset in Barn Owl nestlings. Mean hourly standardized values (± SE) are given in relation to sunrise and sunset as indicated by 0 (i.e. up to 4 hours before and 4 hours after these time points, respectively). Shaded area Dark period. Proportion of time spent awake (filled triangles), in non-REM sleep (open circles) and in REM sleep (open squares) in 29 broods in 2011 is shown. (DOCX 74 kb)
Rights and permissions
About this article

Cite this article
Scriba, M.F., Dreiss, A.N., Henry, I. et al. Nocturnal, diurnal and bimodal patterns of locomotion, sibling interactions and sleep in nestling Barn Owls. J Ornithol 158, 1001–1012 (2017). https://doi.org/10.1007/s10336-017-1458-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-017-1458-3