
Abstract
According to the viability indicator hypothesis, ornaments of many bird species honestly signal individual quality and health status. In this study, we investigated the health- and stress-indicating capacity of both feather- and skin-based ornamental traits in the grey partridge (Perdix perdix), a vulnerable avian species of the European agricultural landscape. In 50 free-living males, we assessed individual health status by examining the differential and absolute leukocyte counts, absolute erythrocyte count, complement activity, and pro-inflammatory immune responsiveness to phytohaemagglutinin (PHA). We found that males expressing extensive melanin-pigmented breast ornaments developed stronger skin inflammation after PHA stimulation than males with limited ornaments, suggesting that these are high-quality individuals that are able to mount a robust immune response. Furthermore, we found that the UV signal of the carotenoid-pigmented eye patch ornament is negatively related to the absolute immature erythrocyte count and the peripheral blood heterophil/lymphocyte (H/L) ratio, indicating that males with a slow pace of life and healthier males express skin ornaments with higher UV reflectance. Ornamental colouration in grey partridge males may, therefore, serve as a reliable signal of condition (in terms of resource balance) and health (in terms of stress resistance) to females.
Zusammenfassung
Melanin- und karotinoidbasierte Ornamentation hängt bei freilebenden Rebhühnern ( Perdix perdix ) von der individuellen Körperkondition ab.
Gemäß der Handicap-Hypothese dienen die Ornamente vieler Vogelarten als ehrliches Signal für individuelle Qualität und Gesundheitszustand. In dieser Studie untersuchten wir die gesundheits- und stressanzeigende Funktion von ornamentalen Merkmalen sowohl des Gefieders als auch der Hautpartien bei Rebhühnern (Perdix perdix), einer gefährdeten Vogelart europäischer Agrarlandschaften. Wir bewerteten den individuellen Gesundheitszustand von 50 freilebenden Männchen, indem wir die differenziellen und absoluten Leukozytenzahlen, die absoluten Erythrozytenzahlen, die Aktivität des Komplementsystems und die proinflammatorische Immunantwort auf Phytohämagglutinin (PHA) betrachteten. Wir stellten fest, dass Männchen, welche ausgedehnte melaninpigmentierte Brustornamente trugen, nach PHA-Stimulation eine stärkere entzündliche Hautreaktion zeigten, als Männchen mit begrenzten Ornamenten, was nahelegt, dass es sich hier um Individuen hoher Qualität handelt, die eine starke Immunantwort aktivieren können. Weiterhin fanden wir heraus, dass das UV-Signal des durch Karotinoidpigmente gefärbten Augenflecks in negativem Verhältnis zur absoluten Anzahl unreifer Erythrozyten und dem peripheren H/L-Verhältnis (Heterophile Granulozyten/Lymphozyten) steht, was andeutet, dass Männchen mit ruhigem Lebenswandel und besserer Gesundheit Hautornamente mit höherer UV-Reflexion ausbilden. Die Schmuckfärbung männlicher Rebhühner kann daher den Weibchen als zuverlässiges Signal für Körperkondition (im Hinblick auf die Ressourcenbilanz) und Gesundheit (bezüglich der Stressresistenz) dienen.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Birds express a broad range of conspicuous colourful ornaments. Several hypotheses have been proposed to explain their signalling function and the physiological mechanism underlying their expression (McGraw 2006a, b; Vinkler et al. 2011). According to the viability indicator hypothesis, these ornaments honestly signal individual quality and health (Andersson 1994). It has been shown that females of several avian species use such ornamental signals to assess male quality, preferring males that exhibit brighter colouration (Hill 1990; Safran et al. 2005).
The most common visual traits in birds are carotenoid-based (yellow-orange-red) and melanin-based (brown-grey-black) ornaments. These traits are produced through different physiological mechanisms and have thus been frequently hypothesised to possess different functions (Griffith et al. 2006). Carotenoid-based ornaments are believed to be more environmentally dependent, since they are obtained from less abundant food resources and may be involved in frequent trade-offs given their immunomodulatory and antioxidant functions. Compared to carotenoid-based ornaments, the expression of melanin ornaments has been shown to be under tighter genetic control (McGraw 2006a, b; Ducrest et al. 2008). Therefore, carotenoid-based traits have traditionally been viewed as more reliable indicators of actual body condition and health than melanin-based ornaments (Badyev and Hill 2000; McGraw and Hill 2000; Senar et al. 2003). However, recent studies have proposed that melanin-based ornaments are also frequently related to individual condition, similarly to carotenoid-based traits (Griffith et al. 2006; Guindre-Parker and Love 2014). In addition, it has been shown that melanin-pigmented traits can also be associated with immune function (e.g., Gangoso et al. 2011; Jacquin et al. 2011).
Colourful ornaments in birds also have significant UV reflectance. Most attention has been paid to the UV-component signalling function of carotenoid-based ornaments such as the grouse comb (Mougeot et al. 2005), mallard bill (Peters et al. 2004), and budgerigar plumage (Griggio et al. 2010). Several studies have suggested a positive correlation of UV reflectance with parasite resistance (Doucet and Montgomerie 2003) and pro-inflammatory immune responsiveness (Griggio et al. 2010). For example, blue tit females with a high UV signal in their feather ornamentation exhibited low baseline corticosterone levels indicating low physiological stress. Thus, it was hypothesised that females with low baseline corticosterone can invest more in feather growth and quality (Henderson et al. 2013). Nevertheless, associations between UV-based signals and other aspects of individual quality (e.g., humoral immunity or haematological traits other than the frequently used H/L ratio) have never been investigated.
The grey partridge (Perdix perdix) was once a widely distributed galliform species in the European-Asian agricultural landscape. However, its population size has dramatically decreased, mainly during the past 60 years (Kuijper et al. 2009). In many European countries, therefore, partridges are nowadays artificially bred and released to reinforce the local populations (Liukkonen 2006; Vidus-Rosin et al. 2010; Buner et al. 2011; Andersen and Kahlert 2012). Both sexes express two ornamental traits that can be used as honest indicators of individual quality, a carotenoid red skin patch behind the eye (Svobodová et al. 2013) and a dark-brown horseshoe-shaped patch in the breast plumage. In captive grey partridge males, it has been found that the two ornamental components were unrelated to each other, and the directions of their association to the H/L ratio were opposite. While the expression level of the carotenoid-based skin ornament is influenced by the actual condition and health (i.e., negatively associated with the H/L ratio), the size of the melanin-based feather ornament is related to dominance and long-term stress (i.e., positively associated with the H/L ratio; Svobodová et al. 2013). Nevertheless, ornaments are frequently context-dependent (Vergara et al. 2012a), and their function may differ not only among species but also among populations (Griffith et al. 2006; Dunn et al. 2010). Investigation of the signalling function of ornaments in wild grey partridge populations is still needed to allow the appropriate utilisation of these traits in breeding conservation programmes.
The aim of our study was to test whether melanin and carotenoid ornaments are related to health and condition in free-living grey partridge males. As general estimators of condition and health status we used standardised weight, differential and absolute leukocyte counts, the total erythrocyte count, complement activity and the pro-inflammatory immune responsiveness to subcutaneous phytohaemagglutinin (PHA) stimulation. Both these haematological traits (Ots et al. 1998; Campbell and Ellis 2007) and the PHA skin-swelling test (Kennedy and Nager 2006; Bílková et al. 2015) have frequently been used in ecological research as good indicators of condition, immune function, and health status (see Adelman et al. 2014). In addition to the frequently used H/L ratio as an indicator of health and stress (Davis et al. 2008), we also evaluated the immature erythrocyte frequency as a trait positively linked to the rate of haematopoiesis, metabolism, and development (Vinkler et al. 2010b). Changes in immature erythrocyte count are known to be associated with anaemic diseases and alterations in the blood cell formation rate (Campbell and Ellis 2007; Yamato et al. 1996; Belskii et al. 2005; Carleton 2008). Since Vinkler et al. (2010b) showed that levels of peripheral blood immature erythrocytes predict the nestling growth rate in scarlet rosefinches (Erythrina erythrina), we hypothesise that this haematopoiesis-related trait indicates the pace of life. We predicted that higher redness and UV reflectance of the carotenoid-based ornament and a larger melanin breast patch signal better health and condition.
Methods
Field procedures
The study was carried out in a hilly, agricultural landscape (490–619 m a.s.l., 17 km2, near the villages Milešín and Nová Ves, 49°22′58.264″N, 16°12′8.364″E, Czech Republic). The core study area consisted of arable land (79.3 % of area), forest fragments (7.9 %), smaller patches of shrubs scattered in fields (6.0 %), a few villages (3.4 %), uncultivated grassy patches (2 %) and small ponds or streams (1.4 %; detailed information in Rymešová et al. 2013). In total, 50 male partridges were caught during January 2009 and January 2010 (30 in 2009 and 20 in 2010) with drop traps at feeding sites. After being caught, the birds were placed into outdoor aviaries where they remained 1–3 weeks until examination (details on caging in Rymešová et al. 2013). First, approximately 200 µl of blood was taken from the brachial vein of each individual and placed into a glass tube with Natt-Herrick solution (15 µl of blood into 2985 μl of Natt-Herrick solution), a glass tube with Drabkin solution (15 µl of blood into 3750 μl of Drabkin solution) and two or three heparinised microhaematocrit capillaries. A blood smear was prepared from a drop of blood. The blood samples collected into the capillaries were immediately centrifuged (11,000 r.p.m for 5 min), haematocrit levels were recorded (the mean value calculated from 2 to 3 capillaries was used for further analysis; n = 38, blood for haematocrit measurements could not be collected in 12 males) and plasma was stored at −80 °C for complement analysis. Then, the weight (measured by a spring balance, accuracy 0.5 g; Pesola, Baar, Switzerland) and tarsus length (measured by a digital calliper, accuracy 0.01 mm; Kinex, Prague, Czech Republic) were recorded. Two ornamental traits potentially important for grey partridge mate choice (Beani and Dessi-Fulgheri 1995) were examined: the red skin patch behind the eyes and the dark-brown horseshoe-shaped patch in the breast plumage. To assess the area of these ornaments, standard digital images of the breast and both sides of the head were taken of each male with a Perfection V10 scanner (Seiko Epson Corporation, Nagano, Japan). All images were taken in a standardised position in a dark room, with grey and colour standard reference swatches equipped with a ruler (a GC 18 grey card and Q 14 colour and grey chart; Danes-Picta, Prague, Czech Republic). In addition, eye patch colouration was measured by an AvaSpec 2048 spectrometer with an AvaLight-XE light source (Avantes, The Netherlands). The spectrometer was standardised against the darkroom and a WS-2 white standard after each ten individuals. The reflection probe (2 mm diameter) was placed at a perpendicular angle at three points of the ornament on both sides of the head. Then, birds were tagged by aluminium rings with a unique code (Museum Prague).
Finally, individual pro-inflammatory immune responsiveness was assessed by the PHA skin-swelling test (Smits et al. 1999; Cucco et al. 2006; Vinkler et al. 2010b, 2014). The thickness of the central part of the left wing web (patagium) was measured three times in each male with a pressure-sensitive digital gauge (Mitutoyo 7301, Mitutoyo Corp., Kanagawa, Japan; accuracy 0.01 mm). This site was then injected subcutaneously with 1 mg PHA dissolved in 40 µl PBS (product nos. L8754 and D5652, Sigma-Aldrich, St Louis, MO, USA). Then, the individual was placed into a small cage in a dark room and left in calm until measuring the response. After 6 ± 0.5 h the thickness of the patagium was again measured (three times with the pressure-sensitive gauge; repeatability r = 0.86, n = 48, p < 0.001; Lessels and Boag 1987). All thickness measurements were performed during afternoon hours to minimise time-dependent variation. The PHA-induced swelling response index was later calculated as the average tissue thickness 6 h after the treatment minus the average thickness before the PHA injection (Vinkler et al. 2010b). The research was approved by the Ethical Committee of the University of Life Sciences in Prague and was carried out in accordance with the current laws of the Czech Republic.
Measurements of ornamental traits
Colouration of the eye patch ornament was measured as its reflectance between 300 and 700 nm. Five variables of colour were calculated according to Svobodová et al. (2013) from the spectral measurements: (1) total brightness (sum of the reflectance from 300 to 700 nm), (2) red chroma (reflectance from 600 to 700 nm, in percent, relative to the total brightness), (3) λ Rvis50 (the wavelength, λ, of the point with reflectance at half the distance between the minimum and maximum reflectance in the visible light spectrum), (4) UV chroma (reflectance from 300 to 400 nm, in per cent, relative to total brightness), and (5) λ RUV (wavelength, of the point in which maximal reflectance was reached in the UV light spectrum, 300–400 nm). Given that each ornamental eye patch (on each side of the head) was measured three times, only average values were used for further analysis. Although PC1 and PC2 of these carotenoid-based ornament colour parameters explained 75.0 % of the variance, we did not use PCA scores in subsequent analyses because using this approach would preclude later utilisation of information on the transmittance in particular variables. Since red chroma correlated with total brightness, UV chroma, and λ Rvis50 (see Table S1), only red chroma and λ RUV were used for further analysis. As λ RUV negatively correlated with λ Rvis50 (which positively correlated with red chroma), higher λ RUV may also be interpreted as lower redness of the red patch area (see also Svobodová et al. 2013).
The areas of the eye patch and breast ornaments were measured from digital images using Adobe Photoshop CS3 software version 10.0 (Adobe Systems Inc., San Jose, CA). First, the image scales were equalised according to the rulers. From these standardised images, areas of the melanin- and carotenoid-based ornaments were measured in the software (see Svobodová et al. 2013; for details).
Haematological assays
In all 50 individuals, the differential leukocyte count, absolute leukocyte count (total white blood cell count, TWBC), absolute erythrocyte count (total red blood cell count, TRBC), and immature erythrocyte frequency were evaluated. The air-dried blood smears were stained with Modified Wright-Giemsa Stain (product no. WG128, Sigma–Aldrich) and scanned with an Olympus CX-31 microscope (Olympus, Tokyo, Japan) under 1000× magnification to count the proportion of lymphocytes, heterophils, eosinophils, basophils, monocytes and immature leukocytes from a sample of 110–140 leukocytes per smear. TWBC and TRBC were calculated from the cell count in 100 and 20 big squares (0.2 × 0.2 mm) in a Bürker counting chamber, respectively. The repeatability of the measurement was r = 0.73, n = 10, p = 0.004. The immature erythrocyte frequencies were estimated from five randomly chosen monolayer fields photographed at 100× objective magnification (ca. 500–1000 cells; for more details, see Vinkler et al. 2010b). The repeatability of this estimate was r = 0.74, n = 10, p = 0.003. Finally, the absorbance at 540 nm was measured for all blood samples dissolved in Drabkin solution using a laboratory spectrophotometer. Haemoglobin concentrations were then calculated using calibration with a human haemoglobin standard (product no. H7379, Sigma-Aldrich). Since the proportion of the variance explained by PC1 and PC2 was very low (52.5 %), particular haematological and immunological variables were chosen for subsequent analyses based on results from a correlation matrix. Given the strong correlations between most erythrocyte- and leukocyte-based parameters (Table S2, namely between the TRBC, haematocrit and haemoglobin concentration, and between the H/L ratio and basophil frequency), only the H/L ratio and TRBC were used as stress- and health-related traits and absolute immature erythrocyte count as a metabolism-related trait in further analysis.
Complement activity
Complement activity was measured using the bioluminescence-based method described in Atosuo et al. (2013). The light emission produced by living cells of a bioluminescent strain of Escherichia coli K12 (pEGFPluxABCDEamp) was measured by a luminometer (LM01-T, Immunotech, Prague, Czech Republic) at 37 °C. Total complement activity of 25 µl blood plasma was determined against 65,000 cells/well (total volume 50 µl) in duplicates. Diminishment of the light signal was positively correlated with the decreasing viability of bacteria. Heat-inactivated serum was used as a control. Relative results of complement activity were computed from kinetic curves as the time difference between the final time of the measurement (2 h) and the time needed to kill 50 % of bacterial cells.
Statistical analyses
The relationships among condition, ornaments, and health status were analysed using linear regression models (LMs) in which the red chroma, λ RUV, area of the carotenoid ornament, and area of the melanin ornament were the response variables. For each response variable a separate LM model was created. The intensity of the inflammatory response after the PHA stimulation, H/L ratio, absolute erythrocyte count, absolute immature erythrocyte count, year, and their meaningful two-way interactions (i.e., those between the year and both haematological and immunological parameters, respectively) were included as explanatory variables. Since there was a weak correlation between the standardised weight and the H/L ratio (r s = −0.37, p = 0.009), the standardised weight was included as a covariate in all models. The significance (set to p < 0.05) of particular terms in the models was calculated based on the change in deviance between the full and reduced (null) models. To achieve the best minimal adequate model (MAM—the model with the lowest parsimony and all variables being significant), all non-significant terms were removed using a backward stepwise procedure, with the statistic reported for each corresponding to the step when they were removed from the model (Crawley 2002). All analyses were performed in the software R. 3.0.3 (R Development Core team 2008).
Results
Grey partridge males showed substantial variation in most health-related parameters measured. Table S3 gives statistics on all condition-dependent traits assessed (including haematological, immunological, and ornamental traits). Interestingly, we found a significant negative relationship between the absolute immature erythrocyte count and complement activity (r s = −0.43, p = 0.017), possibly indicating that metabolic rate is linked to baseline humoral innate immunity investments in male partridges. There was no significant correlation between any of the selected haematological parameters and the magnitude of the PHA-induced skin swelling (in all cases p > 0.198).
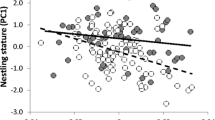
In grey partridge males, the area of the melanin-pigmented breast ornament was positively related to the increase in wing-web thickness 6 h after the PHA injection (LM: estimate ± SE = 855.4 ± 257.4, F 1,48 = 11.05, p = 0.002; Fig. 1). Although the carotenoid pigmentation of the eye patch showed high variation in all parameters of colour measured (Table S3), the two most prominent traits, i.e., the patch area and red chroma, remained unrelated to any of the health-related traits examined (Table S4). An effect of year was found in the MAM model explaining λ RUV (LM: F 1,47 = 10.03, p = 0.003), suggesting inconsistency in λ RUV over time. Therefore, for this response variable, we analysed the data from each year separately. In 2009, λ RUV was negatively related to the H/L ratio (LM: estimate ± SE = −1.695 ± 0.685, F 1,27 = 6.13, p = 0.020, Fig. 2a) and absolute immature erythrocyte count (LM: estimate ± SE = −81.024 ± 38.430, F 1,27 = 4.49, p = 0.043, Fig. 2b). Although in 2010 these relationships were non-significant because of the low sample size, they notably remained in the same direction as in 2009, and the effect of the H/L ratio on λ RUV was only marginally non-significant (LM: estimate ± SE = −1.162 ± 0.525, F 1,18 = 3.52, p = 0.077; Table S5).

Associations between the increase of wing-web thickness (tissue swelling) 6 h after the PHA application and size of the melanin ornament in grey partridge males (n = 50). Dashed lines are 95 % confidence interval

Associations between a the H/L ratio and wavelength of maximal UV reflectance (λ RUV) of the carotenoid ornament, b frequency of immature erythrocytes and λ RUV of the carotenoid ornament in grey partridge males in 2009 (n = 30). Dashed lines are 95 % confidence interval
Discussion
According to the viability indicator hypothesis, individual quality in many bird species is honestly signalled by ornaments. Here we show that in the grey partridge, male individual health and immune responsiveness are signalled through both feather-based and skin-based ornamental traits. Males expressing an extensively melanin-pigmented breast ornament responded with greater intensity to the PHA pro-inflammatory stimulation than males with limited ornamentation, suggesting that these were high-quality individuals that were able to mount a robust immune response. Our results also suggest that healthier males (low H/L ratio) and males with a presumably slow pace of life (low immature erythrocyte count) express skin ornaments with higher UV reflectance. Based on the negative association between complement activity and frequency of immature erythrocytes, we assume that immune function and pace of life are linked in this species, with possible effects on investments into ornamental colouration. Grey partridge ornamental colouration in males may, therefore, serve as a reliable signal of condition and health to females.
Immune function has repeatedly been shown to be condition-dependent (Hõrak et al. 2000; Rodríguez et al. 2014). This is because mounting an immune response may be costly in terms of energy as well as resources (Martin et al. 2003; Sears et al. 2011; Moreno-Rueda 2010). The costs of an immune response are especially high when triggering an inflammatory response (Ashley et al. 2012). Since the PHA skin-swelling test commonly used in birds is a test of pro-inflammatory immune responsiveness (Vinkler et al. 2010a, 2014), individuals in poor condition often tend to develop a weak immune response to PHA treatment (Mougeot and Redpath 2004). Various studies have reported that melanin-pigmented feather-based ornaments are related to body condition (Gladbach et al. 2011), immune function (Galván and Alonso-Alvarez 2008; Jacquin et al. 2011), parasite loads (Fitze and Richner 2002), and oxidative stress (Galván and Alonso-Alvarez 2008). Our results also show that the area of breast melanin in male Grey Partridges is positively linked to the magnitude of the increase in wing-web thickness after a PHA injection, indicating stronger inflammatory responses in high-quality birds. Given that no contradicting association (see Bílková et al. 2015) between haematological traits and the immune responsiveness to PHA has been found, this suggests that free-living partridges with large melanin breast ornaments may be in better condition, investing more into their immune defence. These findings are consistent, for example, with those from feral pigeons (Columba livia), where darker “melanic” individuals had greater skin-swelling responses to PHA than paler ones (Jacquin et al. 2011). Furthermore, this is also congruent with the results of a manipulative experiment in the grey partridge reported by Beani and Dessi-Fulgheri (1995). These authors found that males with larger melanin feather-based ornaments were more frequently involved in antagonistic interactions during the mating period (Beani and Dessi-Fulgheri 1995). Due to frequent involvement in fights being associated with an increased risk of injuries, these males may be predicted to invest more into self-defending pro-inflammatory immune responsiveness.
We found a negative link between standardised weight and the H/L ratio. The H/L ratio is frequently used in the field of avian research as a peripheral blood indicator of inflammation (Buchanan et al. 2003; Bílková et al. 2015), disease (Ots and Hõrak 1998; Garvin et al. 2003), and long-term physiological stress (Davis et al. 2008). Hence, our data suggest that birds in poor condition (low standardised weight) were more stressed. This could have been influenced, for instance, by their susceptibility to infectious diseases, and thus an elevated H/L ratio may be also associated with survival probability (Lobato et al. 2005; Parejo and Silva 2009).
Carotenoid-based colouration is widely acknowledged as a health signal in many avian species (Badyaev and Young 2004). This holds true for both feather ornaments (McGraw and Hill 2000; Hõrak et al. 2004) and living tissues such as the ramphotheca of the bill or bare skin (Pérez-Rodríguez and Viñuela 2008; Mougeot et al. 2010). In our previous experiments, we have shown that redness of the grey partridge eye patch is caused by carotenoid pigmentation (Svobodová et al. 2013), similarly to several other galliform species expressing red-coloured skin-based ornamentation (e.g., Egeland et al. 1993; Mougeot et al. 2007). In captive grey partridge males, we found that the birds with paler carotenoid-based ornament had higher H/L ratios, supporting the relationship between the eye patch signalling and health (Svobodová et al. 2013). In the present study using free-ranging birds, we did not find any direct association between redness (red chroma) and any individual physiological state parameter measured. Nevertheless, our results show that the UV signal of the carotenoid eye patch ornament is negatively related to the peripheral blood H/L ratio and absolute immature erythrocyte count. While an increased H/L ratio is indicative of increased stress and impaired health (Ots et al. 1998; Davis et al. 2008), the absolute immature erythrocyte count is positively linked to the rate of haematopoiesis (Campbell and Ellis 2007; Yamato et al. 1996; Belskii et al. 2005; Carleton 2008), developmental rate (Vinkler et al. 2010b), and presumably also with metabolism and pace of life. Therefore, our results suggest that males with lower UV signalling lived likely faster (reflected by higher immature erythrocyte levels released into circulation) and were exposed to more stress (decreased health associated with an increased H/L ratio). This is consistent with the results reported earlier by Mougeot et al. (2005) showing that impaired health (measured as elevated nematode parasite loads) is negatively related to UV reflectance of the red comb in the closely related red grouse (Lagopus lagopus). The association between the complement activity and absolute immature erythrocyte count also suggests that the immune function and pace of life are linked in this species. This is consistent with the allocation trade-off theory, which suggests that increased resource investments into growth and development may negatively affect health-promoting traits such as immune function (van der Most et al. 2011). Both the immune function and rate of growth and development may then affect ornament development (Vinkler et al. 2011). However, we found a significant relationship among the UV reflectance of the eye patch, immature erythrocyte frequency, and the H/L ratio only in the first year of our study, which supports the idea of context dependency of the signalling (Vergara et al. 2012a, b).
To conclude, this study presents evidence showing that the extent of melanin feather-based ornamentation is a reliable predictor of pro-inflammatory immune responsiveness in free-living grey partridge males. Females choosing males with large melanin ornaments may thus obtain high-quality males in prime condition (in terms of resource balancing). The skin-based red eye patch, on the other hand, appears to signal health (in terms of stress resistance). Considering that ornament function may differ not only among species but also among populations (Griffith et al. 2006; Dunn et al. 2010) and that this signalling may be context-dependent (Vergara et al. 2012a, b), there is little surprise that slightly different health-related signals are emitted in birds from a natural population than in the captive birds investigated earlier (Svobodová et al. 2013). This is likely due to completely different environmental conditions in captivity (no predation, low stress from parasites, pathogens, and starvation) and in the wild. Our findings are thus potentially valuable for conservation efforts in this species.
References
Adelman JS, Ardia DR, Schat KA (2014) Ecoimmunology. In: Schat KA, Kaspers B, Kaiser P (eds) Avian immunology. Academic Press, San Diego, pp 391–411
Andersen LW, Kahlert J (2012) Genetic indications of translocated and stocked grey partridges (Perdix perdix): does the indigenous Danish grey partridge still exist? Biol J Lin Soc 105:694–710
Andersson M (1994) Sexual selection. Princeton Univ Press, Princeton
Ashley NT, Weil ZM, Nelson RJ (2012) Inflammation: mechanisms, costs, and natural variation. Annu Rev Ecol Evol Syst 43:385–406
Atosuo J, Lehtinen J, Vojtek L, Lilius EM (2013) Escherichia coli K-12 (pEGFPluxABCDEamp): a tool for analysis of bacterial killing by antibacterial agents and human complement activities on a real-time basis. Luminescence 28:771–779
Badyaev AV, Young RL (2004) Complexity and integration in sexual ornamentation: an example with carotenoid and melanin plumage pigmentation. J Evol Biol 17:1317–1327
Badyev AV, Hill GE (2000) Evolution of sexual dichromatism: contribution of carotenoid-versus melanin-based coloration. Biol J Lin Soc 69:153–172
Beani L, Dessi-Fulgheri F (1995) Mate choice in grey partridge, Perdix perdix: role of physical and behavioural male traits. Anim Behav 49:347–356
Belskii EA, Lugas’kova NV, Karfidova AA (2005) Reproductive parameters of adult birds and morphophysiological characteristics of chicks in the pied flycatcher (Ficedula hypoleuca Pall.) in technogenically polluted habitats. Russ J Ecol 36:329–335
Bílková B, Vinklerová J, Vinkler M (2015) The relationship between health and cell-mediated immunity measured in ecology: phytohaemagglutinin skin-swelling test mirrors blood cellular composition. J Exp Zool Part A 323:767–777
Buchanan KL, Evans MR, Goldsmith AR (2003) Testosterone, dominance signalling and immunosuppression in the house sparrow, Passer domesticus. Behav Ecol Sociobiol 55:50–59
Buner FD, Browne SJ, Aebischer NJ (2011) Experimental assessment of release methods for the re-establishment of a red-listed galliform, the grey partridge (Perdix perdix). Biol Conserv 144:593–601
Campbell TW, Ellis CK (2007) Avian and exotic animal hematology and cytology. Blackwell Publishing Professional, Ames
Carleton RE (2008) Ectoparasites affect hemoglobin and percentages of immature erythrocytes but not hematocrit in nestling Eastern Bluebirds. Wilson J Ornithol 120:565–568
Crawley MJ (2002) Statistical computing. Wiley, Chichester
Cucco M, Malacarne G, Ottonelli R, Patrone M (2006) Repeatability of cell-mediated and innate immunity, and other fitness-related traits, in the grey partridge. Can J Zool 84:72–79
Davis AK, Maney DL, Maerz JC (2008) The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists. Funct Ecol 22:760–772
Doucet SM, Montgomerie R (2003) Structural plumage colour and parasites in satin bower birds Ptilonorhynchus violaceus: implications for sexual selection. J Avian Biol 34:237–242
Ducrest A, Keller L, Roulin A (2008) Pleiotropy in the melanocortin system, coloration and behavioural syndromes. Trends Ecol Evol 9:502–510
Dunn PO, Garvin JC, Whittingham LA, Freeman-Gallant CR, Hasselquist D (2010) Carotenoid and melanin-based ornaments signal similar aspects of male quality in two populations of the common yellowthroat. Funct Ecol 24:49–158
Egeland ES, Parker H, Liaaen-Jensen S (1993) Carotenoids in combs of capercaillie (Tetrao urogallus) fed defined diets. Poult Sci 72:747–751
Fitze PS, Richner H (2002) Differential effects of a parasite on ornamental structures based on melanins and carotenoids. Behav Ecol 13:401–407
Galván I, Alonso-Alvarez C (2008) An intracellular antioxidant determines the expression of a melanin-based signal in a bird. PLoS One 3:1–7
Gangoso L, Grande JM, Ducrest A-L, Figuerola J, Bortolotti GR, Andre´S JA, Roulin A (2011) MC1R-dependent, melanin-based colour polymorphism is associated with cell-mediated response in the Eleonora’s falcon. J Evol Biol 24:2055–2063
Garvin MC, Homer BL, Greiner EC (2003) Pathogenicity of Haemoproteus danilewskyi, Kruse, 1890, in blue jays (Cyanocitta cristata). J Wildl Dis 39:161–169
Gladbach A, Gladbach D, Quillefeldt P (2011) Male achromatic wing colouration is related to body condition and female reproductive investment in a dichromatic species, the upland goose. J Ethol 29:243–249
Griffith SC, Parker TH, Olson VA (2006) Melanin-versus carotenoid-based sexual signals: is the differences really so black and red? Anim Behav 71:749–763
Griggio M, Zanollo V, Herbert H (2010) UV plumage color is an honest signal of quality in male budgerigars. Ecol Res 25:77–82
Guindre-Parker S, Love OP (2014) Revisiting the condition-dependence of melanin-based plumage. J Avian Biol 45:29–33
Henderson LJ, Heidinger BJ, Evans NP, Arnold KE (2013) Ultraviolet crown coloration in female blue tits predicts reproductive success and baseline corticosterone. Behav Ecol 24:1299–1305
Hill GE (1990) Female house finches prefer colourful males: sexual selection for condition-dependent trait. Anim Behav 40:563–572
Hõrak P, Ots I, Tegelmann L, Møller AP (2000) Health impact of phytohaemagglutinin-induced immune challenge on great tit (Parus major) nestlings. Can J Zool 78:905–910
Hõrak P, Saks L, Karu U, Ots I, Surai PF, McGraw KJ (2004) How coccidian parasites affect health and appearance of greenfinches. J Anim Ecol 73:935–947
Jacquin L, Lenouvel P, Haussy C, Ducatez S, Gasparini J (2011) Melanin-based coloration is related to parasite intensity and cellular immune response in an urban free living bird: the feral pigeon Columba livia. J Avian Biol 42:11–15
Kennedy MW, Nager RG (2006) The perils and prospects of using phytohaemagglutinin in evolutionary ecology. Trends Ecol Evol 21:653–655
Kuijper DPJ, Oosterveld E, Wymenga E (2009) Decline and potential recovery of the European grey partridge (Perdix perdix) population—a review. Eur J Wildl Res 55:455–463
Lessels CN, Boag PT (1987) Unrepeatable repeatabilities: a common mistake. Auk 104:116–121
Liukkonen T (2006) Finnish native grey partridge (Perdix perdix) population differs clearly in mitochondrial DNA from the farm stock used for releases. Ann Zool Fenn 43:271–279
Lobato E, Moreno J, Merino S, Sanz JJ, Arriero E (2005) Haematological variables are good predictors of recruitment in nestling pied flycatchers (Ficedula hypoleuca). Ecoscience 12:27–34
Martin LB, Scheuerlein A, Wikelski M (2003) Immune activity elevates energy expenditure of house sparrows: a link between direct and indirect costs? Proc R Soc Lond 270:153–158
McGraw KJ (2006a) Mechanics of carotenoid-based coloration. In: Hill GE, MacGraw KJ (eds) Bird coloration. Mechanisms and measurements, vol I. Harvard Univ Press, Cambridge, pp 177–242
McGraw KJ (2006b) Mechanics of melanin-based coloration. In: Hill GE, MacGraw KJ (eds) Bird coloration. Mechanisms and measurements, vol I. Harvard Univ Press, Cambridge, pp 243–294
McGraw KJ, Hill GE (2000) Differential effects of endoparasitism on the expression of carotenoid- and melanin-based ornamental coloration. Proc R Soc Lond 267:1525–1531
Moreno-Rueda G (2010) Experimental test of a trade-off between moult and immune response in house sparrows Passer domesticus. J Evol Biol 23(10):2229–2237
Mougeot F, Redpath SM (2004) Sexual ornamentation relates to immune function in male red grouse Lagopus lagopus scoticus. J Avian Biol 35:425–433
Mougeot F, Redpath SM, Leckie F (2005) Ultra-violet reflectance of male and female red grouse, Lagopus lagopus scoticus: sexual ornaments reflect nematode parasite intensity. J Avian Biol 36:203–209
Mougeot F, Martínez-Padilla J, Pérez-Rodríguez L, Bortolotti GR (2007) Carotenoid-based colouration and ultraviolet reflectance of the sexual ornaments of grouse. Behav Ecol Sociobiol 61:741–751
Mougeot F, Martínez-Padilla J, Blount JD, Pérez-Rodríguez L, Webster LMI, Piertney SB (2010) Oxidative stress and the effect of parasites on a carotenoid-based ornament. J Exp Biol 21:400–407
Ots I, Hõrak P (1998) Health impact of blood parasites in breeding great tits. Oecologia 116:441–448
Ots I, Murumagi A, Hõrak P (1998) Haematological health state indices of reproducing great tits: methodology and sources of natural variation. Funct Ecol 12:700–707
Parejo D, Silva N (2009) Immunity and fitness in a wild population of Eurasian kestrels Falco tinnunculus. Naturwissenschaftern 96:1193–1202
Pérez-Rodríguez L, Viñuela J (2008) Carotenoid-based bill and eye ring coloration as honest signals of condition: an experimental test in the red-legged partridge (Alectoris rufa). Naturwissenschaften 95:821–830
Peters A, Delhey K, Denk AG, Kempenaers B (2004) Trade-offs between immune investment and sexual signalling in male mallards. Am Nat 164:51–59
R Development Core Team (2008) R: a language and environment for statistical computing. Vienna, Austria. Available at http://www.Rproject.org
Rodríguez A, Broggi J, Alcaide M, Negro JJ, Figuerola J (2014) Determinants and short-term physiological consequences of PHA immune response in lesser kestrel nestlings. J Exp Zool 321:376–386
Rymešová D, Tomášek O, Šálek M (2013) Differences in mortality rates, dispersal distances and breeding success of commercially reared and wild grey partridges in the Czech agricultural landscape. Eur J Wildl Res 59:147–158
Safran RJ, Neuman CR, McGraw KJ, Lovette IJ (2005) Dynamic paternity allocation as a function of male plumage color in barn swallows. Science 309:2210–2212
Sears BF, Rohr JR, Allen JE, Martin LB (2011) The economy of inflammation: when is less more? Trends Parasitol 27:382–387
Senar JC, Figuerola J, Domènech J (2003) Plumage coloration and nutritional condition in the great tit Parus major: the roles of carotenoids and melanins differ. Naturwissenschaften 90:234–237
Smits JE, Bortolotti GR, Tella JL (1999) Simplifying the phytohaemagglutinin skin-testing technique in studies of avian immunocompetence. Funct Ecol 13:567–572
Svobodová J, Gabrielová B, Synek P, Marsik P, Vaněk T, Albrecht T, Vinkler M (2013) The health signalling of ornamental traits in the grey partridge (Perdix perdix). J Ornithol 154:717–725
van der Most PJ, de Jong B, Parmentier HK, Verhulst S (2011) Trade-off between growth and immune function: a meta-analysis of selection experiments. Funct Ecol 25:74–80
Vergara P, Martinez-Padilla J, Mougeot F, Leckie F, Redpath SM (2012a) Environmental heterogeneity influences the reliability of secondary sexual traits as condition indicators. J Evol Biol 25:20–28
Vergara P, Martinez-Padilla J, Mougeot F, Leckie F, Redpath SM (2012b) The condition dependence of a secondary sexual trait is stronger under high parasite infection level. Behav Ecol 23:502–511
Vidus-Rosin A, Meriggi A, Pella F, Zaccaroni M (2010) Demographic parameters of reintroduced grey partridges in central Italy and the effect of weather. Eur J Wild Res 56:369–375
Vinkler M, Bainová H, Albrecht T (2010a) Functional analysis of the skin-swelling response to phytohaemagglutinin. Funct Ecol 24:1081–1086
Vinkler M, Schnitzer J, Munclinger P, Votýpka J, Albrecht T (2010b) Haematological health assessment in a passerine with extremely high proportion of basophils in peripheral blood. J Ornithol 151:841–849
Vinkler M, Svobodová J, Maršík P, Albrecht T (2011) Carotenoids and health signalling in animals. In: Yamaguchi M (ed) Carotenoids: properties, effects and diseases. Nova Science Publishers, Hauppauge, pp 189–234
Vinkler M, Svobodová J, Gabrielová B, Bainová H, Bryjová A (2014) Cytokine expression in phytohaemagglutinin-induced skin inflammation in a galliform bird. J Avian Biol 45:43–50
Yamato O, Goto I, Maeda Y (1996) Hemolytic anemia in wild seaducks caused by marine oil pollution. J Wildl Dis 32:381–384
Acknowledgments
We thank M. Šálek, D. Rymešová, and O. Tomášek for their help with the field work. We are very grateful to the members of the Hunting Club Borovina Vidonín, especially to J. Holý, P. Jobánek, and P. Jobánková for providing their facilities. This study was supported by the Czech Science Foundation (Project Nos. P206/08/1281 and P502/12/P179), Internal Grant Agency of CULS (IGA 20144268), and Institutional Research Support (Grant SVV-260 313/2016).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
There was no conflict of interests in this research.
Additional information
Communicated by K. C. Klasing.
Electronic supplementary material
Below is the link to the electronic supplementary material.
10336_2016_1349_MOESM1_ESM.docx
Supplementary Table S1: Correlations between parameters of ornaments in captive grey partridge males (n = 50), Spearman correlation coefficient (rs) above and probability level below (p) the diagonal (rs with p < 0.05 was significant) (DOCX 14 kb)
10336_2016_1349_MOESM2_ESM.docx
Supplementary Table S2: Correlations between haematological and immunological parameters in captive grey partridge males (n = 50), Spearman correlation coefficient (rs) above and probability level below (p) the diagonal (rs with p < 0.05 was significant) (DOCX 16 kb)
10336_2016_1349_MOESM3_ESM.docx
Supplementary Table S3: Summary statistics of the examined immunological traits, condition-related variables, and parameters of ornamentation in captive grey partridge males (n = 50). SD = standard deviation, SE = standard error, CV = coefficient of variation, TWBC = total white blood cell count (i.e. absolute leukocyte count), H/L = heterophil/lymphocyte ratio, TRBC = total red blood cell count (i.e., absolute erythrocyte count) (DOCX 16 kb)
10336_2016_1349_MOESM4_ESM.docx
Supplementary Table S4: Results of linear models where the red chroma, λ RUV, area of red patch, and area of the melanin ornament are response variables. Statistics for particular explanatory variables were found using a backward stepwise procedure. All statistics correspond to the step when they were removed from the model (n = 50, Δdf = 1) (DOCX 13 kb)
10336_2016_1349_MOESM5_ESM.docx
Supplementary Table S5: Results of linear models where λ RUV in 2009 and 2010, respectively, are response variables. Statistics for particular explanatory variables were found using a backward stepwise procedure. All statistics correspond to the step when they were removed from the model (n2009 = 30, n2010 = 20, Δdf = 1) (DOCX 12 kb)
Rights and permissions
About this article

Cite this article
Svobodová, J., Gabrielová, B., Hyršl, P. et al. Melanin and carotenoid ornaments are related to the individual condition in free-living grey partridges (Perdix perdix). J Ornithol 157, 1007–1015 (2016). https://doi.org/10.1007/s10336-016-1349-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-016-1349-z