
Abstract
Tracking devices are used in a broad range of species for a broad range of questions, but their potential effects on study species are debated. Outcomes of earlier studies on effects are equivocal: some studies find negative effects on behaviour and life history traits, while others do not. Contrasting results might be due to low sample sizes, temporal scale (no repetition of the study over multiple years) and a limited range of response variables considered. We investigated effects of geolocators on a range of response variables: body condition, physiological states, reproductive performance and, ultimately, annual apparent survival for a medium-sized Palaearctic-African long-distance migrant, the Eurasian Hoopoe Upupa epops, for the combined study period (2009–2014) and for individual years. We investigated response variables 1 year after deployment of the geolocator and found no differences in body condition, physiological states and several components of reproductive performance between individuals with and without geolocators when data were combined. Also, apparent annual survival did not differ between geolocator and control birds. We did, however, find effects in some years possibly related to environmental stochasticity or chance events due to lower sample sizes. We argue that results of studies on the effects of tracking devices should be interpreted and generalized with great caution and suggest that future studies on the effects of tracking devices are conducted over multiple years. Future studies should also apply capture–recapture models to estimate survival, rather than focus solely on return rates.
Zusammenfassung
Keine nachweisbaren Effekte von Leichtgewicht-Geodatenloggern bei einem Langstrecken ziehenden paläarktisch-afrikanischen Zugvogel Technische Instrumente zur individuellen Verfolgung von Tieren werden für viele Arten und verschiedene Verwendungszwecke benutzt. Ob diese jedoch einen Effekt auf das Individuum haben, wird vielfach diskutiert. Ältere Studien zeigen unterschiedliche Resultate auf: manche Studien fanden negative Effekte auf das Verhalten und populationsdynamische Faktoren, während andere Studien keine Effekte nachweisen konnten. Diese kontroversen Resultate entstehen womöglich wegen zu niedrigen Stichprobengrößen, der Zeitspanne (keine mehrjährigen Studien) und weil nur einzelne Faktoren untersucht worden sind. In unserer Studie haben wir die möglichen Effekte von Geodatenloggern auf eine Vielzahl von Variablen untersucht: Körperlicher Zustand, physiologischer Zustand, Reproduktionsleistung und jährliche lokale Überlebensrate. Dies wurde an einem Paläarktisch-Afrikanischen Langstreckenzieher mittlerer Größe, dem Wiedehopf (Upupa epops), untersucht, für die gesamte Dauer unserer Studie (2009–2014) sowie für einzelne Jahre. Wir untersuchten Effekte auf diese Variablen ein Jahr nach dem Einsatz der Geodatenlogger und konnten keine Effekte auf die Körperkondition, den physiologischen Zustand und verschiedene Aspekte der Reproduktionsleistung für die kombinierten Daten über alle Jahre nachweisen. Ebenfalls war die jährliche lokale Überlebensrate nicht unterschiedlich zwischen Vögeln mit und ohne Geodatenlogger. Bei Betrachtung der einzelnen Jahre fanden wir aber in manchen Jahren doch gewisse Effekte, welche möglicherweise mit Umweltstochastizität oder zu kleiner Stichprobengröße zusammenhängen. Wir raten zu Vorsicht bei Interpretation und Verallgemeinerung von Ergebnissen von Studien zu Auswirkungen von Ortungsgeräten und empfehlen, dass zukünftige Studien über mehrere Jahre laufen sollten. Zusätzlich empfehlen wir die Verwendung von „Fang-Wiederfang“-Modellen, um damit jährliche Überlebensraten zu untersuchen anstatt nur auf Rückkehrraten zu fokussieren.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Following individual animals through time and across space usually requires the use of tracking devices, which either actively transmit or archive data, e.g., radio transmitters, geolocators (light archiving loggers) or GPS loggers (Bridge et al. 2011). Despite miniaturization of such devices over the last decades, tracking devices can affect the condition and behavior of individuals carrying them and, ultimately, their fitness. The additional weight and drag may increase energy expenditure (Bowlin et al. 2010; Pennycuick et al. 2012; Vandenabeele et al. 2012), while friction of the device can cause injuries (Peniche et al. 2011; Rodríguez et al. 2009).
Even though it is commonly acknowledged that effects can occur, evidence is equivocal for many investigated traits such as body condition (Adams et al. 2009; Barron et al. 2010), physiological state (Barron et al. 2010; Quillfeldt et al. 2012), components of reproductive performance like onset of breeding (Arlt et al. 2013; Quillfeldt et al. 2012) or fledgling mass (Ackerman et al. 2004; Quillfeldt et al. 2012), and also survival, usually investigated in the form of return rates (Arlt et al. 2013; Barron et al. 2010; Costantini and Møller 2013; Gómez et al. 2014; Phillips et al. 2003; Rodríguez et al. 2009; Scandolara et al. 2014; Townsend et al. 2012). Because these traits may be differently affected by the tracking device, it is difficult to draw general conclusions on device effects when only one or a few traits are studied. For instance, when solely reporting the effects on return rates or onset of the reproductive season, one might well miss the effects on other fitness-related traits such as stress levels. Different tracking devices might cause different effects due to their divergent type of attachment (leg-loop harness, wing-loop harness, elastic versus rigid loops, fixed to bands, glued to feathers), shape (with antenna, light stalk, without any appendices) and duration of deployment (few hours up to several years).
Negative effects of tracking devices might be particularly apparent in migrating animals carrying a device for at least 1 year. Migrants face multiple challenging phases during their annual cycle, during which optimal energy management is crucial for successful reproduction and to survive. A tracking device that is increasingly being used is a lightweight geolocator. These register light intensity by which positions can be estimated and are usually carried for approximately 1 year. A meta-analysis on the effects of geolocators on birds showed survival to be negatively affected for tagged birds (Costantini and Møller 2013). However, the effects could be statistically supported only when combining all data; most individual studies had insufficient sample sizes to draw conclusive results. Furthermore, none of these studies considered more than three seasons, although the magnitude of effects may vary from year to year due to environmental variation (Bro et al. 1999; Hupp et al. 2006; Pietz et al. 1993).
The equivocal effects of geolocators can result from many reasons, such as size and mode of attachment of the device, behavior of the study species and the weight of the devices relative to the weight of the study species. Moreover, equivocal effects might result from low sample sizes, the temporal scale of the study (no repetition over multiple years) and a restrictive range of the studied traits. To date, a comprehensive study on the effects of carrying a geolocator over an entire year on a wide range of traits over multiple years is still lacking.
To fill this gap, we aimed to identify the effects of geolocators on several life-history traits of a Palaearctic-African migratory bird, the Eurasian Hoopoe (Upupa epops), over a 7-year period. We compared body condition, physiological states, several components of reproductive performance and survival between geolocator and control birds. We investigated whether the effects were present over the whole study period combined and for each year separately.
Methods
Study population and data acquisition
We studied geolocator effects in a population of Hoopoes breeding in nestboxes in the Valais, south-western Switzerland (46°14′N, 7°22′E). The study area is about 62 km2 in area and harbors ~550 nestboxes (Arlettaz et al. 2010). Hoopoes feed on large soil invertebrates and birds from this population are long-distance Palaearctic-African migrants, which spend the non-breeding season in the Sahelian belt south of the Sahara (Bächler et al. 2010). They typically return to the breeding grounds from the beginning of April, start breeding in late April and the last nestlings fledge in early August. Most Hoopoes in our study population produce only one clutch a year, but about one-third have two or more clutches (Hoffmann et al. 2015). Birds were caught in or at the nestboxes which were checked every 10–14 days from mid-April until mid-July each year.
To investigate geolocator effects, we studied several traits both for the whole study period combined as well as for each individual year. We investigated traits that might indicate short-term effects [body condition, physiological states (baseline corticosterone, stress response), components of reproductive performance (territory occupancy, onset of breeding, brood success, number of fledglings, fledgling weight, average food biomass delivered to the nest per hour and per visit] of carrying a geolocator, and survival as an indication of a long-term effect. For a definition of each response variable, see Table 2b (below).
Data collection
Between 2008 and 2013, 328 breeding birds were equipped with geolocators of type SOI-GDL1 (Bächler et al. 2010) of which 81 were recaptured between 2009 and 2014. These geolocators have a light stalk of 10 mm positioned under an angle of 45° on the geolocator and weigh on average 1.32 g including a leg-loop harness, which corresponds to ~1.9 % of the body mass of a Hoopoe. As a control group for birds that were recaptured with a geolocator after 1 year, we used 273 individuals that never carried a geolocator (see Table 1 for the annual sample size).
Birds were caught at least 5 days after their young had hatched and were given a unique combination of a metal ring and three color rings. Following this procedure, about 85–90 % of our study population was marked and measured each year. We only used data of the first capture or of the first time a bird was equipped with a geolocator; we excluded repeated tracks in all analyses except the capture–recapture analysis. We did this to exclude the possibility that birds compensate their behavior or physiology for carrying a geolocator over longer deployments, which would lead to an unfair comparison. To obtain a measure of body condition, we recorded mass to the nearest 0.1 g and measured maximum chord wing length to the nearest 0.1 mm, and calculated their ratio. Data were obtained between 2009 and 2014.
Corticosterone as a measure of physiological state was sampled during the reproductive seasons 2010–2013 by taking blood from 164 breeding birds (117 control and 47 equipped birds; Table 1) when they were feeding their nestlings. We obtained measurements of baseline and stress response following the method described in Schmid et al. (2013): all included baseline samples were taken within 3 min after capture; which is within the time limit after which baseline corticosterone might be affected (Romero and Reed 2005). Only individuals for which we had data for both corticosterone levels were included.
To investigate the different components of reproductive performance, occupied nestboxes were visited every 3 days to obtain accurate data on numbers of eggs, hatchlings and fledglings, and on breeding phenology. Nestlings were ringed at an age of c.18 days and were weighed to the nearest 0.1 g. All these data were available for the reproductive seasons 2009–2014.
Data on parental care in terms of delivered prey biomass were collected in just 1 year (2012) for 45 pairs. We analyzed prey biomass delivered per hour and per visit when nestlings were about 2 weeks old. These data were acquired using video recordings with estimated weights for each prey size/species (for more details, see Guillod 2013).
Analysis of body condition, physiological states and reproductive performance
Explanatory variables are listed in Table 2a and definitions of response variables in Table 2b. Additionally, models on body condition and physiological states included breeding phase: the time between onset of breeding and capture. The model on physiological states furthermore included age, capture time, the time between capture and the first blood sample when baseline corticosterone was measured (delta 1, minimum 0.33, mean 1.45 and maximum 4 min), and the time between capture and the second blood sample when the stress response was analyzed (in minutes, delta 2; see Table 2 for a complete description of variables used). The model on parental care (Table 2a) additionally included the age of nestlings, frequency of visits, and territory (as a random factor) as explanatory variables.
To test for geolocator effects we used generalized linear mixed effect models with year as a random effect to study effects over the whole study period and generalized linear models to investigate each year separately. The analyses were conducted with R 3.1.0 (R Core Team, Vienna, 2014) using the function lmer and glm. The best model for each dependent variable was selected using a step-wise backwards procedure with the package lmerTest (Kuznetsova et al. 2015). To illustrate each model’s results, we made posterior predictions for geolocator and control individuals using parametric bootstrapping of 1000 simulations with the package arm.
Recapture probabilities and annual survival
We used a capture–recapture model to estimate survival, taking into account detection probability and the potential loss of a geolocator. We used a multi-event model framework (Pradel 2009), which links field observations to biological states of individuals. We defined the following states: “Alive with geolocator” (G+), “Alive without geolocator” (G−) and “Dead” (D). However, we handled birds at recapture and equipped some of them with a geolocator, thereby modifying their states in a deterministic way. To account for this, we added a dummy capture occasion after the real capture occasion and therefore expanded the biological states with “Alive and equipped with geolocator” (S), “Alive and not equipped” (R), “Alive with geolocator but outside the study population” (AE+) and “Alive without geolocator but outside the study population” (AE−). At each capture, birds could be in states G+, G− or D. At the following dummy occasion (t′ + 1), captured birds were assigned state S or R, while non-captured birds were assigned state AE+ or AE− (see Appendix A for a complete description of the model). This model allowed us to estimate loss rate (L) and survival probability (φ) while accounting for imperfect detection (P) of birds and deterministic changes of individual states.
We investigated effects of year, sex and geolocator on survival and recapture probabilities, and effects of year and sex on geolocator loss probability: L y×sex, Φ y×sex, Ρ sex×geo with y, sex, and geo the effects of year, sex and geolocator, respectively.
To investigate whether the structure of the model was appropriate and included major sources of heterogeneity, we performed a goodness-of-fit of the capture–recapture model using the program U-CARE (Choquet et al. 2009a). The model selection followed a step-down approach (Lebreton et al. 1992), starting with the initial model and sequentially fitting models with constrained parameterization for detection, loss and survival probabilities. Model selection relied on Akaike’s information criterion (Burnham and Anderson 2002). Model selection and parameter estimations were performed using the program E-SURGE (Choquet et al. 2009b).
Results
Body condition, physiological states and components of reproductive performance
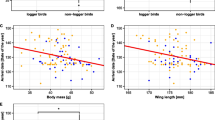
Neither body condition, physiological states nor any component of reproductive performance were affected by carrying a geolocator when data of all years were pooled (Fig. 1; Appendix B, Tables S1–S3). When geolocator was added to the final model, effect sizes were very small (Table S4).

Summary of model predictions using 1000 simulations for response variables studied over multiple years. Points represent control and triangles geolocator Eurasian Hoopoes (Upupa epops), respectively. Median and 95 % spread of the data are indicated. The line indicates the median for all years combined, the gray area the 95 % distribution. For more information, see text and Appendix B, Tables S1–S3; for estimated sizes of geolocator effects, see Appendix B Table S5
We found differences in several traits between geolocator and control birds in individual study years, but, contrary to our expectations, these were mostly positive for geolocator birds (Fig. 1): (1) their body condition was higher in 2012 (Table S1); (2) they tended to occupy better territories in 2009 (Table S3 A); (3) their onset of breeding was earlier in 2011 (~2 weeks) and in 2012 (~5 days) (Table S3 B); and (4) the number of fledglings of successful first broods was higher in 2011 and 2014 (~1 more fledgling; Table S3 D). We only found negative effects for baseline corticosterone, which was slightly elevated for geolocator birds in 2012 (Table S2 A).
Annual survival and recapture probabilities
The goodness-of-fit test of the capture–recapture model was not significant (χ 2 = 18.78, df = 26, P = 0.85) indicating an appropriate structure of our initial model.
Based on AIC, the most supported model included sex, a temporal trend in survival and constant parameters for loss and detection probabilities (model 15, Table 3), but a geolocator effect was not retained. The best model including a geolocator effect on apparent survival was considerably worse (model 10, Table 3).
The recapture probability was very high (0.88 ± SE 0.04), suggesting most marked birds were detected and captured each year. The annual loss rate was high with about one-quarter of geolocators lost (0.28 ± SE 0.04). The mean annual apparent survival probability over the whole study period was 0.35 ± 0.02 (from model 13, best model without time variation). Survival of geolocator individuals was lower compared to survival of control individuals (0.33 ± 0.04 vs. 0.36 ± 0.02, from model 10), but confidence intervals of both estimates overlapped for all years (0.27–0.41 vs. 0.31–0.41; Fig. 2).

Point estimates and 95 % confidence intervals (from Model 7) of survival probabilities of Hoopoes with (points) and without geolocator (triangles)
Discussion
We found no effect of geolocators on any of the investigated traits when data of all years were pooled: body condition, physiological state, reproductive performance and survival did not differ between geolocator and control birds. This is in contrast to many earlier studies (mentioned in Costantini and Møller 2013), yet most of those focussed solely on the effects of a few traits that were assessed during just 1 year with relatively low sample sizes. When we analyzed data for each year separately, some traits did differ between geolocator and control birds.
Effects of geolocator on life-history traits for the combined study period
A prominent reason why the geolocators had essentially no effect on different traits of Hoopoes could be the relative low weight of the devices we used (~2 % of body mass). However, it has been shown that even for weights below 2 %, the effects of a device might be profound in some species. For example, the effects are more pronounced in seabirds where devices would have to be streamlined both in the air as well as underwater (Vandenabeele et al. 2012). Moreover, aerial foragers were especially negatively affected by carrying a geolocator which might relate to the extra drag of the device (Costantini and Møller 2013). Hoopoes are landbirds that mainly forage on larger soil invertebrates and thus might be less affected by carrying a device. Lastly, Hoopoes are not known to make long, non-stop flights, but rather have multiple stopovers along the way (Bächler et al. 2010).
The glucocorticoid hormone corticosterone orchestrates many physiological processes, including energy mobilization (Bonier et al. 2011) and reproduction (Schmid et al. 2013), and it also affects behavior (Coppens et al. 2010). The physiological role of corticosterone is quite distinct depending on whether it circulates at low baseline or at high acute stress-induced levels. At low baseline levels, corticosterone is involved in maintaining physiological homeostasis in everyday life (Romero 2004). In response to stressful events, however, corticosterone is released in high amounts into the blood and helps to mobilize energy and to redirect the behavior to self-maintenance (Sapolsky et al. 2000). While a short-term release of corticosterone is considered beneficial in allowing individuals to overcome threatening situation, chronically elevated levels can entail negative long-term effects (Sapolsky et al. 2000). If geolocators were to evoke stress in the Hoopoes, for instance by the higher weight they have to carry or by hampering their flight performance, we would expect an increased level of circulating baseline corticosterone and/or a reduced release of corticosterone in response to handling. However, we found no effects on baseline corticosterone and stress response. This might be related to the low relative weight of the geolocators (<2 % of the body weight) or because, over the course of 1 year, the birds got used to the geolocator and thus stress levels returned to normal. Other studies, however, have shown increases in both baseline corticosterone and stress response after 1 year of deployment (Elliott et al. 2012; Quillfeldt et al. 2012), though a very recent study that investigated corticosterone in feathers for two aerial feeding migrants also did not find any differences (Fairhurst et al. 2015).
Even though their condition and physiological states might be the same, there could still be differences in reproductive performance, but we found no effects of geolocator on any component of reproductive performance, not even on finer-scale traits such as fledgling mass and food provisioning rates, contrasting with the majority of the literature (Ackerman et al. 2004; Adams et al. 2009; Barron et al. 2010; Costantini and Møller 2013). For example, we found no difference in the onset of breeding, an important trait that is directly linked to the quality of the territory occupied and thus indirectly to reproductive success: birds that arrive later in the breeding grounds are forced to occupy lower quality territories and therefore would have a lower reproductive success (Tschumi et al. 2014).
Survival was often found to be negatively affected for geolocator birds (Costantini and Møller 2013). In almost all studies, the effect of geolocators on survival was assessed based on return rates. However, return rates are the product of the probabilities to survive and to be recaptured. Consequently, return rates can only be used as an index of survival, if recapture probabilities for individuals with a geolocator and for control individuals are identical. Since researchers often target catching birds with geolocators to retrieve the data, resighting and thus recapture probabilities of geolocator birds are likely to be elevated. Moreover, if a study is restricted to breeding individuals, recapture probability is affected by the probability to breed, which itself might be affected by the geolocator. Consequently, return rates cannot be used to evaluate the effects of geolocators on survival, because they depend on strong assumptions that remain untested. Using a capture–recapture framework, we found no difference in recapture probability between geolocator and control birds for our study population, suggesting an equal effort to detect both groups of individuals. Furthermore, the high recapture rate in our study, due to a systematic search for birds in the study area, allowed us to accurately estimate demographic parameters. By also taking into account geolocator loss, we are thus confident in our estimates of survival and our finding that carrying a geolocator did not induce a negative effect on annual survival.
Interannual variation in geolocator effects
Although we found no differences between geolocator and control birds for the combined study period, some traits differed in specific years. These differences can most likely be explained by low sample sizes in some years, most particularly in 2012 when we only had 5 control versus 12 geolocator individuals, and for which we found geolocator birds to be in better body condition and to start breeding earlier. In later years, smaller numbers of control individuals were available, because the largest part of the population were equipped with a geolocator. Therefore, the pool of control birds that never had a geolocator got smaller.
Another explanation for the differences could also be related to interannual variations in environmental conditions, be it on the nonbreeding grounds, along the migration route or upon arrival back in the breeding grounds. For instance, body condition of Montserat Orioles (Icterus oberi) was lower when pre-breeding rainfall was low (Oppel et al. 2013) and reproductive success in shrubsteppe sparrows was greater in years with wetter compared to drier conditions in the breeding grounds (Rotenberry and Wiens 1991). Carrying a device might be an additional constraint in such years with harsher conditions (Pietz et al. 1993; Bro et al. 1999) affecting traits negatively, while under ‘normal’ environmental circumstances a device might exhibit no effect.
Conclusion
It is clear that tracking devices allow sampling very valuable data for fundamental and conservation-oriented research, but results should be interpreted with caution. Besides animal welfare issues, inference based on such data can be compromised if the tracking device affects the behavior or performance of the individuals or because they induce a selection towards strong individuals. Here, we found no effect of geolocators for a European population of medium-sized near-passerine birds, the Eurasian Hoopoe. Even though effects of devices are likely linked to the relative weight of the device and the individual’s behavior and are thus species-specific, such results could be similar for other medium-sized ground-foraging migrants, such as Rollers (Coracias garrulus) or birds where the geolocators have a similar relative weight (<2 %).
Furthermore, our study shows that results can only be considered robust when looking at several years and at a range of life-history traits. We thus recommend investigating effects of tracking devices over several years to include a range of particular environmental conditions and to focus on a broad array of variables. Finally, using a capture–recapture model as proposed here may be a better way to investigate demographic parameters than solely reporting return rates.
References
Ackerman JT, Adams J, Takekawa JY et al (2004) Effects of radiotransmitters on the reproductive performance of Cassin’ s auklets. Wildl Soc Bull 32:1229–1241
Adams J, Scott D, McKechnie S et al (2009) Effects of geolocation archival tags on reproduction and adult body mass of sooty shearwaters (Puffinus griseus). N Z J Zool 36:355–366
Arlettaz R, Schaub M, Fournier J, Reichlin TS (2010) From publications to public actions: when conservation biologists bridge the gap between research and implementation. Bioscience 60:835–842
Arlt D, Low M, Pärt T (2013) Effect of geolocators on migration and subsequent breeding performance of a long-distance passerine migrant. PLoS ONE 8:e82316. doi:10.1371/journal.pone.0082316
Bächler E, Hahn S, Schaub M et al (2010) Year-round tracking of small trans-Saharan migrants using light-level geolocators. PLoS ONE 5:e9566. doi:10.1371/journal.pone.0009566
Barron DG, Brawn JD, Weatherhead PJ (2010) Meta-analysis of transmitter effects on avian behaviour and ecology. Methods Ecol Evol 1:180–187. doi:10.1111/j.2041-210X.2010.00013.x
Bonier F, Moore IT, Robertson RJ (2011) The stress of parenthood? Increased glucocorticoids in birds with experimentally enlarged broods. Biol Lett 7:944–946. doi:10.1098/rsbl.2011.0391
Bowlin MS, Henningsson P, Muijres FT et al (2010) The effects of geolocator drag and weight on the flight ranges of small migrants. Methods Ecol Evol 1:398–402
Bridge ES, Thorup K, Bowlin MS et al (2011) Technology on the move: recent and forthcoming innovations for tracking migratory birds. Bioscience 61:689–698. doi:10.1525/bio.2011.61.9.7
Bro E, Clobert J, Reitz F (1999) Effects of radiotransmitters on survival and reproductive success of gray partridge. J Wildl Manag 63:1044–1051
Burnham KP, Anderson DA (2002) Model selection and multimodel inference: a practical information-theoretic approach. Springer, New York
Choquet R, Lebreton J-D, Gimenez O et al (2009a) U-CARE: utilities for performing goodness of fit tests and manipulating capture–recapture data. Ecography (Cop) 32:1071–1074. doi:10.1111/j.1600-0587.2009.05968.x
Choquet R, Rouan L, Pradel R (2009b) Program E-Surge: a software application for fitting multievent models. In: Thomson D, Cooch E, Conroy M (eds) Modeling demographic processes in marked populations. Springer, New York, pp 845–865
Coppens CM, de Boer SF, Koolhaas JM (2010) Coping styles and behavioural flexibility: towards underlying mechanisms. Philos Trans R Soc Lond B 365:4021–4028. doi:10.1098/rstb.2010.0217
Costantini D, Møller AP (2013) A meta-analysis of the effects of geolocator application on birds. Curr Zool 59:697–706
Elliott KH, McFarlane-Tranquilla L, Burke CM et al (2012) Year-long deployments of small geolocators increase corticosterone levels in murres. Mar Ecol Prog Ser 466:1–7
Fairhurst GD, Berzins LL, Bradley DW et al (2015) Assessing costs of carrying geolocators using feather corticosterone in two species of aerial insectivore. R Soc Open Sci 2:150004. doi:10.1098/rsos.150004
Gómez J, Michelson CI, Bradley DW et al (2014) Effects of geolocators on reproductive performance and annual return rates of a migratory songbird. J Ornithol 155:37–44. doi:10.1007/s10336-013-0984-x
Guillod N (2013) Landscape of prey in a hoopoe population: consequences of spatial variation of molecricket availability on reproductive success and conservation strategies. Unpublished thesis, University of Bern, Bern
Hoffmann J, Postma E, Schaub M (2015) Factors influencing double brooding in eurasian hoopoes Upupa epops. Ibis 157:17. doi:10.1111/ibi.12188
Hupp JJW, Pearce JJM, Mulcahy DMD, Miller DA (2006) Effects of abdominally implanted radiotransmitters with percutaneous antennas on migration, reproduction, and survival of Canada geese. J Wildl Manag 70:812–822
Kuznetsova A, Christensen RHB, Bavay C, Brockhoff PB (2015) Automated mixed ANOVA modeling of sensory and consumer data. Food Qual Prefer 40:31–38
Lebreton JD, Burnham KP, Clobert J, Anderson DR (1992) Modeling survival and testing biological hypotheses using marked animals: a unified approach with case studies. Ecol Monogr 62:67–118
Oppel S, Hilton GM, Allcorn R et al (2013) The effects of rainfall on different components of seasonal fecundity in a tropical forest passerine. Ibis 155:464–475. doi:10.1111/ibi.12052
Peniche G, Vaughan-Higgins R, Carter I et al (2011) Long-term health effects of harness-mounted radio transmitters in red kites (Milvus milvus) in England. Vet Rec 169:311. doi:10.1136/vr.d4600
Pennycuick CJ, Fast PLF, Ballerstädt N, Rattenborg N (2012) The effect of an external transmitter on the drag coefficient of a bird’s body, and hence on migration range, and energy reserves after migration. J Ornithol 153:633–644. doi:10.1007/s10336-011-0781-3
Phillips RA, Xavier JC, Croxall JP (2003) Effects of satellite transmitters on albatrosses and petrels. Auk 120:1082–1090
Pietz PJ, Krapu GL, Greenwood RJ, Lokemoen JT (1993) Effects of harness transmitters on behavior and reproduction of wild mallards. J Wildl Manag, pp 696–703
Pradel R (2009) The stakes of capture-recapture models with state uncertainty. In: Thomson D, Cooch E, Conroy M (eds) Modeling demographic processes in marked populations. Springer, New York, pp 781–795
Quillfeldt P, McGill RAR, Furness RW et al (2012) Impact of miniature geolocation loggers on a small petrel, the thin-billed prion Pachyptila belcheri. Mar Biol 159:1809–1816
Rodríguez A, Negro JJ, Fox JW, Afanasyev V (2009) Effects of geolocator attachments on breeding parameters of lesser kestrels. J Ornithol 80:399–407
Romero LM (2004) Physiological stress in ecology: lessons from biomedical research. Trends Ecol Evol 19:249–255. doi:10.1016/j.tree.2004.03.008
Romero LM, Reed JM (2005) Collecting baseline corticosterone samples in the field: is under 3 min good enough? Comp Biochem Physiol A 140:73–79. doi:10.1016/j.cbpb.2004.11.004
Rotenberry JT, Wiens JA (1991) Weather and reproductive variation in shrubsteppe sparrows: a hierarchical analysis. Ecology 72:1325–1335. doi:10.2307/1941105
Sapolsky RM, Romero ML, Munck A (2000) How do glucocorticoids influence the stress response? Endocr Rev 21:55–89
Scandolara C, Rubolini D, Ambrosini R et al (2014) Impact of miniaturized geolocators on barn swallow Hirundo rustica fitness traits. J Avian Biol 45:1–7
Schmid B, Tam-Dafond L, Jenni-Eiermann S et al (2013) Modulation of the adrenocortical response to acute stress with respect to brood value, reproductive success and survival in the eurasian hoopoe. Oecologia 173:33–44. doi:10.1007/s00442-013-2598-7
Svensson L (1992) Identification guide to European passerines. Pubished by the author, Stockholm
Townsend JM, Rimmer CC, McFarland KP (2012) Radio-transmitters do not affect seasonal mass change or annual survival of wintering bicknell’s thrushes. J Ornithol 83:295–301. doi:10.1111/j.1557-9263.2012.00378.x
Tschumi M, Schaub M, Arlettaz R (2014) Territory occupancy and parental quality as proxies for spatial prioritization of conservation areas. PLoS ONE 9:e97679. doi:10.1371/journal.pone.0097679
Vandenabeele SP, Shepard EL, Grogan A, Wilson RP (2012) When three per cent may not be three per cent; device-equipped seabirds experience variable flight constraints. Mar Biol 159:1–14. doi:10.1007/s00227-011-1784-6
Acknowledgments
We thank the people that conducted the fieldwork, and Nico Guillod and Alain Jacot for the prey delivery data. We thank three anonymous reviewers that helped improve an earlier version of this manuscript. RVW is supported by a Grant of the Swiss National Science Foundation (Nr. 31003A_138354). All applicable guidelines for animal welfare were followed.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by F. Bairlein.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
van Wijk, R.E., Souchay, G., Jenni-Eiermann, S. et al. No detectable effects of lightweight geolocators on a Palaearctic-African long-distance migrant. J Ornithol 157, 255–264 (2016). https://doi.org/10.1007/s10336-015-1274-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-015-1274-6