
Abstract
Effects of deployment of miniaturised transmitters and loggers have been studied mainly in diving seabirds such as penguins, and less so in flying seabirds. However, some studies of albatrosses and petrels recorded extended trip durations and elevated rates of nest desertion following device attachment, especially if transmitter loads exceeded 3 % of adult mass. Studies have usually compared performance parameters such as trip duration, meal mass, breeding success or rate of return in the next season between birds with devices and controls. We here examined the effects of geolocator loggers (Global Location Sensing, (GLS)) on thin-billed prions Pachyptila belcheri (130 g), by comparing performance parameters and additionally eco-physiological parameters. GLS weighed ca. 1 % of the body mass, and were fixed on leg rings, which may influence the flight efficiency by creating an asymmetric load. We found no differences in the performance parameters, either in the season of attachment or the season following recovery. Similar stable isotope ratios in adult blood and feather samples further indicated that the foraging ecology was not influenced. However, after 1 year of logger deployment, adults differed in their hormonal response to stress: while baseline corticosterone levels were not influenced, corticosterone levels in response to handling were elevated. Moreover, increased heterophil/lymphocyte ratios and a decreased tail growth in winter suggest that carrying the GLS was energetically costly, and adults adapted physiologically to the higher work load, while keeping up a normal breeding performance.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The development of miniature data loggers has opened a way to study the foraging ecology and movements of animals, which formerly were impossible to follow, such as medium to small pelagic seabirds during the winter migration period. However, the additional weight and volume of external devices may increase the cost of flight, thereby reducing foraging efficiency, especially for such small species. Thus, we need to test for potential changes in behaviour resulting from carrying external devices (Barron et al. 2010; Vandenabeele et al. 2011, 2012).
Because most petrel species regularly perform long-distance trips lasting several days or even weeks and often covering thousands of kilometres, these birds are particularly prone to the effects of reduced flight efficiency. In fact, researchers studying parent–offspring conflict in petrels have often aimed at changing the parental behaviour by experimentally adding weight or reducing the wing area of adults. For example, Leach’s storm petrel Oceanodroma leucorhoa delivered less food to their offspring in response to an increased cost of flight by reducing wing span (Mauck and Grubb 1995). No difference was found in nutritional condition between treatment and control parents (Mauck and Grubb 1995), which is in line with life-history theory predicting that parents in long-lived species should prioritise their own survival (i.e. future reproduction) over that of the chick (i.e. current reproduction) if resources are limited. Similarly, Antarctic Prions Pachyptila desolata that had cost of foraging increased experimentally (by adding mass to adults) kept their body mass stable, while increasing the duration of long foraging trips during chick feeding (Weimerskirch et al. 1999). Likewise, Cory’s shearwaters Calonectris diomedea carrying experimental loads increased foraging trip lengths during incubation (Navarro et al. 2008), and thin-billed prions Pachyptila belcheri increased the frequency of short trips during chick provisioning in response to removal of some flight feathers (Weimerskirch et al. 1995). However, the experimental reduction of foraging efficiency in these studies was relatively large. The 45-g loads added in Cory’s shearwaters are equivalent to 6 % increase in body mass and hence, wing loading (Navarro et al. 2008), and 10 g added in Antarctic Prions are equivalent to 7 % increase in body mass. With logging devices, the common rule is to stay below 3 % of body mass (Phillips et al. 2003), and thus, minimise the impact. However, some studies have found effects even with lighter logging devices (e.g. Adams et al. 2009).
While researchers have traditionally measured changes in body mass and trip duration (e.g. references above), physiological responses have received less attention (e.g. Navarro et al. 2008). However, changes in the physiology underlie the observed behavioural responses and are a key to their understanding. For example, increased effort can lead to an increase in the levels of corticosterone in plasma (Silverin 1982; Kitaysky et al. 1999), a hormone that is directly involved in the regulation of foraging behaviour (e.g. Kitaysky et al. 2001a; Angelier et al. 2007). Furthermore, the allocation of resources to other body functions might be influenced during increased effort.
In the present study, we therefore measured not only performance parameters of birds carrying geolocator loggers (Global Location Sensing, GLS) and control birds, but added the following measures designed to indicate more subtle changes in physiology and behaviour:
-
(1)
Stable isotope signatures of C and N in blood and feathers. Changes in these isotopes are expected if the birds changed their foraging ecology in the weeks prior to sampling (blood) or in the winter (feathers).
-
(2)
Levels of corticosterone in plasma. Changes in this hormone indicate energetically stressful phases.
-
(3)
Leucocyte profiles. Lymphocytes and heterophils belong to different components of the immune system, and relative percentages of these cells are often changed during chronically stressful or energetically demanding phases.
-
(4)
Feather growth measures of chicks and adults, in particular tail feather growth. Reduced feather growth can indicate a reduced availability of resources, due to lower delivery in chicks or due to reduced foraging efficiency in adults.
Materials and methods
Study site and study species
The study was carried out at New Island, Falkland/Malvinas Islands (51°43′S, 61°18′W), in December 2009 and February 2010 (deployment of geolocators), and between December 2010 and March 2011 (deployment of geolocators and measurement of effects).
Thin-billed prions are small (130 g) seabirds breeding on sub-Antarctic islands, chiefly in the Falkland and Kerguelen archipelagos. Thin-billed prions show the typical procellariiform pattern of a single-egg clutch and slow chick development, with an average fledging period of 50 days (Strange 1980). Under good conditions, the chicks accumulate large lipid reserves, attaining peak masses of up to 190 % of adult mass. At the end of nestling development, chicks lose mass and fledge close to adult mass. Variation in water temperatures and upwelling coupled with larger-scale processes (e.g. El Niño/La Niña) may affect the availability of prey, and an analysis of provisioning in relation to ocean climate and prey availability (Quillfeldt et al. 2007) suggested that periods of elevated sea surface temperatures were associated with periods of low food availability for thin-billed prions.
Thin-billed prions are burrow nesters, and we reached chicks and adults in their nest chambers via short access tunnels in the roof of each burrow, capped with removable stone lids. This system facilitated rapid access to chicks, reducing overall disturbance.
Egg and chick measurements
Nests were followed in detail throughout the breeding season 2010–2011. In December 2010, marked nests of GLS and control birds were checked for eggs to determine laying success (Table 1). We followed 31 control nests, 17 nests where adults were equipped with GLS in December 2010 (called “season of deployment”) and 14 nests where adults had been equipped with GLS in the breeding season 2009–2010 (called “season of recovery”). Egg sizes were measured and nests were monitored for hatching chicks. After the guard stage, chicks were weighed daily, using a digital 200-g balance to the nearest 0.1 g, until fledging in late February 2011. Daily measurements enable correct peak mass detection (e.g. Quillfeldt et al. 2007), while regularly handled chicks do not suffer any detectable influence on growth or corticosterone levels (Quillfeldt et al. 2009). Every 3 days, wing and tail lengths were measured with a wing and feather ruler, respectively. Wing and tail growth rates were calculated during the phase of approximately linear growth (20–40 days of age for wings, 30–50 days for tails) in mm per day. As measures of breeding success, we registered laying success (the percentage of known nests containing an egg), hatching success (the percentage of eggs hatching) and fledging success (the percentage of hatched chicks surviving to fledging). We calculated a body condition index using the observed body mass (m), relative to a multi-year mean mass for chicks of each age (mmean), using the formula: BC index = m/mmean* 100 (e.g. Quillfeldt et al. 2009).
Geolocator deployment and retrieval, sampling of adults
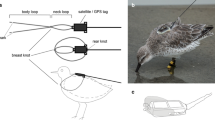
Deployment of loggers (MK10, weighing 1.1 g, developed by British Antarctic Survey, Cambridge, UK, and mounted on Darvic rings fitted to the leg) was carried out in December 2009 (incubation, N = 20), February 2010 (chick feeding, N = 5) and December 2010 (incubation, N = 25). In the breeding season 2009–2010, the 25 adults corresponded to 15 nests (i.e. two adults were equipped in 10 nests, and a single bird in the remaining 5 nests). In December 2010, the 25 adults corresponded to 17 nests (i.e. two adults were equipped in 8 nests, and a single bird in the remaining 9 nests). Of the 25 loggers deployed on the first breeding season, 20 were recovered in December 2010. Five adults with devices were not recaptured. In the case of three adults from two different nests, the nest had been occupied by a different pair. One nest had become too deep and could not be reached, and in one nest, the female was recaptured, but found with a different male. These nests were therefore excluded from the analyses of breeding success.
In December 2010, adults were captured in the nest during daytime while incubating. Adults that had been carrying a GLS over the winter 2010 (N = 20) were compared to adults without GLS in that period (“controls,” N = 23). They were weighed with a digital 200-g balance to the nearest 0.1 g, and measured using callipers (tarsus, bill), wing ruler (wing length) and feather ruler (tail length, as the protruding length of the central tail feathers). To calculate adult body condition, we corrected body mass for a measure of body size derived from a principal component analysis (PCA) with varimax rotation of measures of structural size (wing length, tarsus length, bill length, bill width and bill height). This procedure was justified as indicated by the Kaiser–Meyer–Olkin measure of sampling adequacy of 0.6 and Bartlett’s test of sphericity revealing significance (χ2 = 21, df = 10, P = 0.019). The PCA extracted one factor: PC1 explained 38 % of the total variation and was positively correlated to all five measures of body size.
Blood samples were collected within 2 min after capture of the adult by hand, by puncture from the brachial vein in heparinised capillaries. All samples were taken during daytime (0800 to 1800 h local time). Blood smears were made directly in the field, and whole blood samples (max. 0.5 ml) were kept on ice. Blood cells were separated from plasma by centrifugation within 1 h. Blood cells were oven dried at 37 °C for stable isotope analyses, while plasma was frozen at −20 °C. One capillary of blood (0.06 ml) was used to determine haematocrit values, using a haematocrit centrifuge. Although more adults were captured and measured, only 23 control adults with blood samples were included into the statistical analyses, to achieve a balanced data set. Some adults had insufficient blood volumes for haematocrit and corticosterone analyses, and only 22 control adults were included into the stable isotope analyses; therefore, the sample sizes are lower in these analyses (18–23 per group, Table 2). For stable isotope analyses, we also sampled a small section from the inner vane of primary feathers P1 and P6, as well as undertail coverts (UTC).
Stable isotope analyses
Carbon and nitrogen isotope analyses were carried out at Scottish Universities Environmental Research Centre as described previously (e.g. Quillfeldt et al. 2008) on 0.65- to 0.7-mg aliquots, weighed into tin cups, using dried red blood cells and small fragments of feathers. Carbon and nitrogen isotope ratios were measured simultaneously by continuous-flow isotope ratio mass spectrometry (CF-IRMS) using a Costech Elemental Analyser (EA) linked to a Thermo Finnigan Delta Plus XP Mass Spectrometer. Two laboratory standards were analysed for every 10 unknown samples, allowing any instrument drift over a typical 14-h run to be corrected. Stable isotope ratios were expressed in δ notation as parts per thousand (‰) deviation from the international standards V-Pee Dee Belemnite (carbon) and AIR (nitrogen). Based on internal standards (tryptophan), the analytical precision (±1 SD) was estimated as ±0.18 and ±0.17 ‰ for δ15N and δ13C, respectively.
Hormone measurements
Hormone analyses were carried out at the Veterinary University of Vienna. Plasma was extracted using diethyl ether. After extraction, the vials were kept overnight at −20 °C, and on the next day, the organic phase was decanted into a new vial and evaporated in a stream of nitrogen. The extract was re-dissolved in assay buffer and the concentration of corticosterone was measured using enzyme immunoassay (Palme and Möstl 1997).
Differential leucocyte counts
In the laboratory, blood smears were fixed with methanol for 1 min and stained using the Giemsa technique. The differential count included relative percentages of lymphocytes (L), heterophils (H), monocytes, basophils and eosinophils, which were identified according to the criteria of Hawkey and Dennett (1989). A total of 100 leucocytes were counted per slide. Using the percentages of heterophils and lymphocytes, the H/L ratio was determined for each sample. The number of fields until reaching the target of 100 leucocytes was registered, and the number of erythrocytes was counted in three representative fields, and used to calculate the total leucocyte number per 10,000 erythrocytes.
Data analysis
Statistical tests were performed with SPSS 11.0.0. Kolmogorov–Smirnov tests were used to test for normality (P > 0.2). Homogeneity of error variances was checked by plotting residuals against predicted values. The requirements for parametric tests were met for all chick parameters. Several adult parameters (CORT (3 min) ng/ml, H/L ratio, eosinophils %, δ13C in all feathers (P1, P6, UTC) and δ15 N in P6 and UTC feathers) were not normally distributed and hence we applied non-parametric tests for these parameters.
Results
Performance parameters
We found no influence of either short-term (same breeding season, i.e. up to 3 months) or long-term (breeding season following 1 year of deployment) deployment on breeding performance (Table 1). The breeding success was not influenced, and chicks were fed equally well as in control nests, as shown by their growth rates. Although the mortality of very young chicks was relatively high, the surviving chicks in the year 2010–2011 achieved high peak body masses, indicating good food availability during the chick-rearing period.
Foraging ecology
According to stable isotope ratios of nitrogen (representing trophic level) and carbon (representing foraging location), no statistically significant differences between control adults and adults carrying GLS were found, either in the breeding season (blood samples) or during moult (feathers). However, δ15N values were slightly lower in GLS birds in all four measurements (blood and four feathers grown at different times), suggesting there might be a small decrease in trophic level in birds carrying GLS although this is not detected by the statistical tests (P = 0.052 for P6, Table 2).
Condition and stress parameters
Adults had similar body condition, haematocrit values and baseline corticosterone values (Table 2). However, corticosterone values in response to handling were elevated in adults carrying GLS, compared to control adults (Table 2). Adults with GLS also showed elevated H/L ratios (Table 2). This was mainly caused by an increase by 10 % in the relative number of heterophil leucocytes, while numbers of lymphocytes and monocytes decreased and numbers of eosinophil leucocytes were strongly depleted (Table 2). Tail feathers of birds carrying GLS were 3 mm shorter than in previous seasons by the same individuals (Table 2).
Discussion
Data from GLS loggers have the potential to revolutionise our knowledge on the distribution of pelagic seabirds during periods of absence from the colony (e.g. Shaffer et al. 2006) and are valuable in a conservation context (e.g. Guildford et al. 2012). However, impacts of logger attachment on the birds and the quality of the data need to be considered (e.g. Igual et al. 2005).
In the present study, we found only mild influences of GLS weighing ca. 1 % of the body mass on thin-billed prions. We found no differences in any performance parameters, either in the season of attachment or the season following recovery (Table 1). Although some parameters like fledging success have previously been shown to have a limited response to provisioning by adult thin-billed prions, other parameters like peak mass and tail growth rate were sensitive to changes in feeding rates (Quillfeldt et al. 2007). Therefore, our data suggest that adults maintained a normal provisioning rate, both while they were carrying the GLS and in the breeding season after recovery.
Stable isotope ratios in adult blood and feather samples did not differ between control adults and adults that had been carrying a GLS for 1 year at the time of sampling. A previous study (Quillfeldt et al. 2008) had found changes among years: during years of low food availability, more depleted isotope values suggested more southerly foraging (based on carbon isotope ratios) and a lower trophic level diet (based on nitrogen isotope values). The similar values of control and GLS birds in the present study, in contrast, indicate that the foraging behaviour may not have changed substantially either in the winter (feather samples) or during incubation (blood samples).
However, ecophysiological measures suggested that carrying the GLS was energetically costly, and adults adapted physiologically to the higher workload, while maintaining a normal breeding performance.
Stress physiology
After 1 year of logger deployment, adults differed in their hormonal response to a stressor, as they had elevated corticosterone levels in response to handling. The acute stress response is thought to be adaptive, especially through facilitation of escape behaviour in response to attacks by predators or inclement weather (e.g. Wingfield et al. 1998). Therefore, researchers generally believe that the preferred state of the stress response should contain a combination of low baseline, fast increase and rapid induction of negative feedback (e.g. Breuner et al. 2008). In contrast to this belief, however, the only two studies that have so far examined the relationship between the stress response and survival in wild vertebrates directly (reviewed in Breuner et al. 2008) found the opposite: populations of Galapagos marine iguanas with greater corticosterone reactivity had lower chances of survival (Romero and Wikelski 2001) and the magnitude of the stress response at the chick age was negatively related to survival and recruitment in white stork chicks (Blas et al. 2007). Further studies also suggest that increased stress responsiveness is usually associated with poorer condition. For example, increased adrenocortical responses to acute stress have been found in chicks fed a reduced diet (Kitaysky et al. 2001b), and in adult birds in poorer condition (Hood et al. 1998; Breuner and Hahn 2003; Pereyra and Wingfield 2003). In these adults, self-maintenance behaviours will be activated more quickly when energetic reserves are low. In consequence, the adults carrying geolocators in our study were probably more likely to abandon breeding in adverse conditions such as low food availability.
In addition to the hormonal response to stress, chronic stress may also elicit responses of the immune system (e.g. Ruiz et al. 2002), in particular by a higher proportion of circulating heterophils (H) and low proportion of circulating lymphocytes (L), leading to elevated H/L ratios. In the present study, adults carrying GLS for 1 year showed increased H/L ratios. The lower number of monocytes, the decrease in eosinophils and the similar total leucocyte numbers of adults carrying GLS all indicate that this elevation was due to energetic demands, and not to infection (Davis et al. 2008).
Leucocyte profiles (relative numbers of white blood cell types) have been used by a number of ecological studies to assess immune function and stress in wild birds (e.g. Figuerola et al. 1999; Ruiz et al. 2002; Davis et al. 2004; Gladbach et al. 2010; Dehnhard et al. 2011). Because immune function competes for the resources that can be allocated to other activities, there is a trade-off between the immune system and general condition in free-living birds (e.g. Møller and Petrie 2002). Leucocytes comprise the main element of the immune system, both in the innate and the acquired arm of the immune system. Innate immunity works predominantly by means of phagocytosis. Monocytes and three different types of granulocytes (heterophils, eosinophils and basophils) form the first line of cellular defence against invading microbial pathogens. Heterophils are the most frequent type of granulocyte. Acquired immunity is more pathogen specific and works by means of cell-mediated and humoral responses by the lymphocytes.
Elevated H/L ratios have been found to be related to a range of stressors and energetically demanding phases, such as long-distance migration (Owen and Moore 2006), injuries (Vleck et al. 2000), parasitic infection (Lobato et al. 2005), in chicks during times of low food availability (Hoi-Leitner et al. 2001; Lobato et al. 2005; Hylton et al. 2006), in birds attending experimentally increased broods (Illmonen et al. 2003; Suorsa et al. 2004) and adverse environmental conditions (Plischke et al. 2010). The consequences of increased H/L ratios, however, are not well known. The reduced number of lymphocytes is often interpreted as a sign of immunosuppression, because only the lymphocytes possess the attributes of diversity, specificity, memory and self versus non-self recognition, while all other cells play accessory roles in adaptive immunity, serving to activate lymphocytes, increasing the effectiveness of antigen clearance by phagocytosis or secreting immune effector molecules. However, it has also been pointed out (e.g. Davis et al. 2008 and references therein) that a reduced number of circulating lymphocytes does not result from large-scale destruction of these usually long-lived cells, but rather from changes in the distribution of lymphocytes. In response to glucocorticoids, lymphocytes migrate from the blood into other tissues such as lymph nodes, spleen, bone marrow and skin, where they might be needed during the stress response. As leucocyte profiles do not indicate the number of heterophils or lymphocytes that are stored in other body compartments, H/L ratios provide no direct information regarding immunocompetence, to our knowledge. Nevertheless, H/L ratios have been shown to have predictive power, for example, for winter survival in breeding birds (Kilgas et al. 2006), recruitment into the adult population (Lobato et al. 2005) and susceptibility to infection (Al-Murrani et al. 2002).
Feather growth
During the moult in winter, the tail feathers of birds carrying GLS were regrown 3 mm shorter than in previous seasons by the same individuals. Environmental conditions and, in particular, food availability have been shown to influence the length, mass and symmetry of feathers (Murphy et al. 1988; Grubb 1989; Swaddle and Witter 1994; Stratford and Stouffer 2001). In adult birds, moulting requires a large amount of energy and nutrients. For example, feathers produced in a complete moult account for over 20 % of the total lean dry body mass of passerines (Lindström et al. 1993; Jenni and Winkler 1994). The shorter tails in the birds carrying GLS suggest that these birds were handicapped by the weight or asymmetry of the loads, and could invest less into feather regrowth during moult. However, the fact that tail length was only marginally shorter in comparison with control birds suggests that this effect was not detrimental.
Conclusions
Previous studies of the impact of loggers on albatrosses and petrels have indicated that the mass of the device as a proportion of adult body weight should be below 3 % to avoid adverse effects (Phillips et al. 2003; see also Vandenabeele et al. 2012 for differences among bird species). The present study suggests that thin-billed prions were able to care for their offspring, survive the winter and return for breeding despite the handicap of the geolocator, which was well below the critical point at ~1 % of body mass. Nevertheless, the increased workload was reflected in ecophysiological measurements, although these changes did not result in decreased performance. However, it has to be kept in mind that the environmental conditions were favourable during the period of study and that under poorer conditions the response of the birds might be different. The elevated stress response, in particular, suggests an increased readiness to abandon breeding.
References
Adams J, Scott D, McKechnie S, Blackwell G, Shaffer SA, Moller H (2009) Effects of geolocation archival tags on reproduction and adult body mass of sooty shearwaters (Puffinus griseus). New Zealand J Zool 36:355–366
Al-Murrani WK, Al-Rawi IK, Raof NM (2002) Genetic resistance to Salmonella typhimurium in two lines of chickens selected as resistant and sensitive on the basis of heterophil/lymphocyte ratio. Br Poult Sci 43:501–507
Angelier F, Clement-Chastel C, Gabrielsen GW, Chastel O (2007) Corticosterone and time-activity Black-legged budget: an experiment with kittiwakes. Horm Behav 52:482–491
Barron DG, Brawn JD, Weatherhead PJ (2010) Meta-analysis of transmitter effects on avian behaviour and ecology. Methods Ecol Evol 1:180–187
Blas J, Bortolotti GR, Tella JL, Baos R, Marchant TA (2007) Stress response during development predicts fitness in a wild, long lived vertebrate. Proc Natl Acad Sci USA 104:8880–8884
Breuner CW, Hahn TP (2003) Integrating stress physiology, environmental change, and behavior in free-living sparrows. Horm Behav 43:115–123
Breuner CW, Patterson SH, Hahn TP (2008) In search of relationships between the acute adrenocortical response and fitness. Gen Comp Endocrinol 157:288–295
Davis AK, Cook KC, Altizer S (2004) Leukocyte profiles in wild House Finches with and without mycoplasmal conjunctivitis, a recently emerged bacterial disease. EcoHealth 1:362–373
Davis AK, Maney DL, Maerz JC (2008) The use of leukocyte profiles to measure stress in vertebrates: a review for ecologists. Funct Ecol 22:760–772
Dehnhard N, Poisbleau M, Demongin L, Quillfeldt P (2011) Do leucocyte profiles reflect temporal and sexual variation in body condition over the breeding cycle in southern rockhopper penguins? J Ornithol 152:759–768
Figuerola J, Munoz E, Gutierrez R, Ferrer D (1999) Blood parasites, leucocytes and plumage brightness in the Cirl Bunting, Emberiza cirlus. Funct Ecol 13:594–601
Gladbach A, Gladbach DJ, Quillfeldt P (2010) Variations in leucocyte profiles and plasma biochemistry are related to different aspects of parental investment in male and female Upland geese Chloephaga picta leucoptera. Comp Biochem Physiol A 156:269–277
Grubb TC Jr (1989) Ptilochronology: feather growth bars as indicators of nutritional status. Auk 106:314–320
Guilford T, Wynn R, McMinn M, Rodríguez A, Fayet A, Maurice L, Jones A, Meier R (2012) Geolocators reveal migration and pre-breeding behaviour of the critically endangered Balearic Shearwater Puffinus mauretanicus. PlosOne 7:e33753
Hawkey CM, Dennet PB (1989) A colour atlas of comparative veterinary haematology. Wolfe, Ipswich
Hoi-Leitner M, Romero-Pujante M, Hoi H, Pavlova A (2001) Food availability and immune capacity in serin (Serinus serinus) nestlings. Behav Ecol Sociobiol 49:333–339
Hood LC, Boersma PD, Wingfield JC (1998) The adrenocortical response to stress in incubating Magellanic penguins (Spheniscus magellanicus). Auk 115:76–84
Hylton RA, Frederick PC, de la Fuente TE, Spalding MG (2006) Effects of nestling health on postfledging survival of wood storks. Condor 108:97–106
Igual JM, Forero MG, Tavecchia G, González-Solis J, Martínez-Abraín A, Hobson KA, Ruiz X, Oro D (2005) Short-term effects of data-loggers on Cory’s shearwater (Calonectris diomedea). Mar Biol 146:619–624
Ilmonen P, Hasselquist D, Langefors Å, Wiehn J (2003) Stress, immunocompetence and leukocyte profiles of pied flycatchers in relation to brood size manipulation. Oecologia 136:148–154
Jenni L, Winkler R (1994) Molt and ageing of European passerines. Academic Press, London
Kilgas P, Tilgar V, Mänd R (2006) Hematological health state indices predict local survival in a small passerine bird, the great tit (Parus major). Physiol Biochem Zool 79:565–572
Kitaysky AS, Wingfield JC, Piatt JF (1999) Dynamics of food availability, body condition and physiological response in breeding black-legged kittiwakes. Funct Ecol 13:577–585
Kitaysky AS, Wingfield JC, Piatt JF (2001a) Corticosterone facilitates begging and affects resource allocation in the black-legged kittiwake. Behav Ecol 12:619–625
Kitaysky AS, Kitaiskaia EV, Wingfield JC, Piatt JF (2001b) Dietary restriction causes chronic elevation of corticosterone and enhances stress response in red-legged kittiwake chicks. J Comp Physiol B 171:701–709
Lindström Å, Visser GH, Daan S (1993) The energetic cost of feather synthesis is proportional to basal metabolic rate. Physiol Zool 66:490–510
Lobato E, Moreno J, Merino S, Sanz JJ, Arriero E (2005) Haematological variables are good predictors of recruitment in nestling pied flycatchers (Ficedula hypoleuca). Ecoscience 12:27–34
Mauck RA, Grubb TC Jr (1995) Petrel parents shunt all experimentally increased reproductive costs to their offspring. Anim Behav 49:999–1008
Møller AP, Petrie M (2002) Condition dependence, multiple sexual signals, and immunocompetence in peacocks. Behav Ecol 13:248–253
Murphy ME, King JR, Lu J (1988) Malnutrition during the postnuptial molt of White-crowned Sparrows: feather growth and quality. Can J Zool 66:1403–1413
Navarro J, Gonzalez-Solis J, Viscor G, Chastel O (2008) Ecophysiological response to an experimental increase of wing loading in a pelagic seabird. J Exp Mar Biol Ecol 358:14–19
Owen JC, Moore FR (2006) Seasonal differences in immunological condition of three species of thrushes. Condor 108:389–398
Palme R, Möstl E (1997) Measurement of cortisol metabolites in faeces of sheep as a parameter of cortisol concentration in blood. Z Saugetierkd Int J Mammal Biol 62(suppl 2):192–197
Pereyra ME, Wingfield JC (2003) Changes in plasma corticosterone and adrenocortical response to stress during the breeding cycle in high altitude flycatchers. Gen Comp Endocrinol 130:222–231
Phillips RA, Xavier JC, Croxall JP (2003) Effects of satellite transmitters on albatrosses and petrels. Auk 120:1082–1090
Plischke A, Quillfeldt P, Lubjuhn T, Merino S, Masello JF (2010) Leucocytes in adult burrowing parrots Cyanoliseus patagonus in the wild: variation between contrasting breeding seasons, gender and condition. J Ornithol 151:347–354
Quillfeldt P, Strange IJ, Masello JF (2007) Sea surface temperatures and behavioural buffering capacity in thin-billed prions P. belcheri: breeding success, provisioning and chick begging. J Avian Biol 38:298–308
Quillfeldt P, McGill RAR, Masello JF, Weiss F, Strange IJ, Brickle P, Furness RW (2008) Stable isotope analysis reveals sexual and environmental variability and individual consistency in foraging of thin-billed prions. Mar Ecol Prog Ser 373:137–148
Quillfeldt P, Poisbleau M, Chastel O, Masello JF (2009) Acute stress hyporesponsive period in nestling thin-billed prions P. belcheri. J Comp Physiol A 195:91–98
Romero LM, Wikelski M (2001) Corticosterone levels predict survival probabilities of Galapagos marine iguanas during El Niño events. Proc Natl Acad Sci USA 98:7366–7370
Ruiz G, Rosenmann M, Novoa FF, Sabat P (2002) Hematological parameters and stress index in rufous-collared sparrows dwelling in urban environments. Condor 104:162–166
Shaffer SA, Tremblay Y, Weimerskirch H, Scott D, Thompson DR, Sagar PM, Moller H, Taylor GA, Foley DG, Block BA, Costa DP (2006) Migratory shearwaters integrate oceanic resources across the Pacific Ocean in an endless summer. PNAS 103:12799–12802
Silverin B (1982) Endocrine correlates of brood size in adult pied flycatchers, Ficedula hypoleuca. Gen Comp Endocrinol 47:18–23
Strange IJ (1980) The thin-billed prion, P. belcheri, at New Island, Falkland Islands. Gerfaut 70:411–445
Stratford JA, Stouffer PC (2001) Reduced feather growth rates of two common birds inhabiting central Amazonian forest fragments. Conserv Biol 15:721–728
Suorsa P, Helle H, Koivunen V, Huhta E, Nikula A, Hakkarainen H (2004) Effects of forest patch size on physiological stress and immunocompetence in an area-sensitive passerine, the Eurasian treecreeper (Certhia familiaris): an experiment. Proc R Soc Lond B 271:435–440
Swaddle JP, Witter MS (1994) Food, feathers and fluctuating asymmetries. Proc R Soc Lond B 255:147–152
Vandenabeele SP, Wilson RP, Grogan A (2011) Tags on seabirds: how seriously are instrument-induced behaviours considered? Anim Welf 20:559–571
Vandenabeele SP, Shepard EL, Grogan A, Wilson RP (2012) When three per cent may not be three per cent: device-equipped seabirds experience variable flight constraints. Mar Biol 159:1–14
Vleck CM, Vertalino N, Vleck D, Bucher TL (2000) Stress, corticosterone, and heterophil to lymphocyte ratios in free-living Adelie Penguins. Condor 102:392–400
Weimerskirch H, Chastel O, Ackermann L (1995) Adjustment of parental effort to manipulated foraging ability in a pelagic seabird, the thin-billed prion P. belcheri. Behav Ecol Sociobiol 36:11–16
Weimerskirch H, Fradet G, Cherel Y (1999) Natural and experimental changes in chick provisioning in a long-lived seabird, the Antarctic prion. J Avian Biol 30:165–174
Acknowledgments
Fieldwork at New Island was supported by the New Island Conservation Trust, Ian, Maria and Georgina Strange, was approved by the Falkland Islands Government (Environmental Planning Office) and funded by grants provided by Deutsche Forschungsgemeinschaft DFG (Qu 148/1ff). We thank Hendrika (Riek) van Noordwijk and Gabriele Schafheitle for help in the field and laboratory, respectively. Funding for the stable isotope work was provided by the Natural Environment Research Council, UK (Grant NE/102237X/1) and carried out at the Life Sciences Mass Spectrometry Facility. We would like to thank Heiko Schmaljohann and Sylvie Vandenabeele for helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by S. Garthe.
Rights and permissions
About this article
Cite this article
Quillfeldt, P., McGill, R.A.R., Furness, R.W. et al. Impact of miniature geolocation loggers on a small petrel, the thin-billed prion Pachyptila belcheri . Mar Biol 159, 1809–1816 (2012). https://doi.org/10.1007/s00227-012-1971-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00227-012-1971-0