
Abstract
We provide new information on Yunnan snub-nosed monkey (Rhinopithecus bieti) behavioral ecology, contributing to future conservation efforts within the Laojun Mountain National Park. Habitat evaluation procedures are used to quantify the value of land as a habitat for a species. We analyzed environmental variables hypothesized to influence habitat suitability for Yunnan snub-nosed monkeys, and mapped the distribution of suitable habitats across the study area and adjacent areas. Spatial analysis with GPS data was conducted to investigate home-range change of these monkeys. Predictor variables were generated using ArcMap and R programming language. We prepared 34 environmental variables at 30-m spatial resolution. Maxent was used to analyze environmental variables that contributed to suitability. Using satellite remote sensing and GIS, we modeled the distribution of suitable habitat for Yunnan snub-nosed monkeys in the Jinsichang area of the Laojun Mountains in China. This study did not describe the frequency or intensity of habitat use. Habitat suitability was affected by several variables, the most influential, as determined by permutation importance, being mean diurnal temperature range (31.6%), precipitation during the wettest quarter of the year (30.4%), average annual precipitation (17%), normalized difference vegetation index (5%), wetness (4.6%), and aspect (4.5%). This habitat suitability model provides information about the current distribution of Yunnan snub-nosed monkeys, which is important for appropriate implementation of conservation actions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
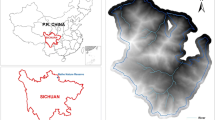
Yunnan snub-nosed monkeys (Rhinopithecus bieti) are endemic to the People’s Republic of China (hereafter, China) and are distributed throughout northwestern Yunnan Province, China and southeastern Tibet (Fig. 1). They are confined to a narrow habitat in the Yun Range between the Yangtze River to the east and the Mekong River to the west, with a population of ~ 2000 individuals (Ren 2009; Liedigk et al. 2012). Yunnan snub-nosed monkeys are one of the most endangered Old World primates (Long and Kirkpatrick 1994), and their habitat is restricted to elevations between 3000 and 4000 m and areas dominated by alpine, temperate forests (Long et al. 1995). Yunnan snub-nosed monkeys have been studied across the Yun Range in areas such as the Baima Snow Mountains, Lasha Mountains, and Yunlong Mountains. Collectively, these regions have approximately 100–300 individuals (Cui et al. 2006; Huang et al. 2012; Li et al. 2003). Laojun Mountain National Park (NP), which is located within the Yun Range, is estimated to have approximately 150–200 individuals, but this area has not been well studied. Laojun Mountain NP is a complex landscape with undulating terrain, deep gorges, sheer peaks, and diverse flora and fauna (Li et al. 2009).

Free-ranging Yunnan snub-nosed monkey at Laojun Mountain National Park
As an endangered species on the IUCN Red List, Yunnan snub-nosed monkeys (also called black snub-nosed monkeys by the IUCN) face major threats from hunting and habitat loss (Bleisch and Richardson 2008). Their dispersal has been limited by the Mekong, Salween, and Yangtze Rivers since at least the early Miocene (Liu et al. 2009; Liedigk et al. 2012; Ma et al. 2014). These landscape features created natural habitat fragmentation that has been exacerbated by more recent anthropogenic disturbances such as poaching, logging, livestock grazing, and collection of forest products for subsistence (Li et al. 2009; Bleisch and Richardson 2008; Hallet and Molnar 2001). The outlook for Yunnan snub-nosed monkeys is not very optimistic in view of the anthropogenic interference and natural features of the landscape they inhabit (Jablonski 1998; Wu 1993; Li et al. 2009). These factors also make tracking and data collection on this species difficult.
From 2004 to present, The Nature Conservancy (TNC) in China has led increased research efforts to create more effective conservation strategies for Yunnan snub-nosed monkeys. Increased knowledge of the species’ behavioral ecology and ranging patterns is central to this effort. On-the-ground efforts in Laojun Mountain NP have contributed a large amount of data on the presence of monkeys in the region and some information on the extent of anthropogenic disturbance.
Species distribution models (SDMs), usually based on statistical models, facilitate inferences about species and habitat distributions across many spatial and temporal scales (Franklin 2009; Fourcade et al. 2014). SDMs have recently become increasingly popular among researchers for evaluating relationships between species and their environments across a variety of scales and geographic areas (Cayuela et al. 2009). These models use species occurrence and environmental variables to derive spatially explicit predictions of distributions or habitat suitability (Franklin 2009), and to inform and execute conservation strategies (Hernandez et al. 2006).
In our study, we used SDMs to gain insight into the influence of extreme environmental conditions (e.g., low temperatures and high altitudes) on the presence of Yunnan snub-nosed monkeys in Laojun Mountain NP. The use of extreme environments by Yunnan snub-nosed monkeys has been documented, but detailed information remains sparse. For example, Kirkpatrick et al. (1998) and Long and Kirkpatrick (1994) suggested that these monkeys are found at high altitudes (3000–4300 m) because of the presence of lichens, a key food resource. It was also noted that Yunnan snub-nosed monkeys prefer areas where they can access direct sunlight for warmth (Xiang et al. 2007; Gittins 1982). Additionally, Liu and Zhao (2004) documented the use of mild east- and southeast-facing slopes for protection from strong winds. We aimed to present a relatively comprehensive habitat suitability map of Yunnan snub-nosed monkeys in Laojun Mountain in a more systematic and quantitative way.
In this study, we analyzed environmental variables hypothesized to influence habitat suitability (for example, higher elevations, steep slopes, warm temperatures, low moisture, and high sun exposure, shelter from strong wind) and mapped the distribution of suitable habitat across the study area and adjacent regions. We recorded both direct and indirect evidence of Yunnan snub-nose monkey presence. To evaluate how the spatial distribution of biophysical variables relates to the species’ distribution in our study site, we analyzed these data together with medium-resolution remote sensing data and SDMs. To test our hypotheses on habitat suitability, we used Maxent to construct SDMs to identify environmental and anthropogenic variables that have the greatest influence on the monkeys. A predictive SDM could help improve the design and execution of a conservation plan for Yunnan snub-nosed monkeys (Hernandez et al. 2006).
Methods
Study site
The Jinsichang study area is located within Laojun Mountain NP, approximately 80 km northwest of Lijiang, China. The Laojun Mountains within the NP are an upland karst formation that spans over 1085 km2 and are part of the Three Parallel Rivers World Heritage Site (Huadong 2013; Policy Research Office 2006). Laojun Mountain NP is a biodiversity hotspot, with more than 1800 widely distributed biological species, including 51 nationally rare and endangered species (Fang et al. 2004; Wu 2006; Li et al. 2007; ROPGYP 2010), and constitutes an important portion of the distribution range of Yunnan snub-nosed monkeys (Long and Kirkpatrick 1994). Laojun Mountain NP is a temperate region with pronounced seasonality (Grueter et al. 2009). The rainy season lasts from early summer to early autumn, with the most intense rainfall occurring between the end of July and the middle of August. Snow usually covers the ground from November to April.
The Jinsichang (26°53′N, 99°37′E) study area is the main territory for a band consisting of multiple unit groups of free-ranging Yunnan snub-nosed monkeys in Laojun Mountain NP (Fig. 2), which covers approximately 320 km2 of rich natural resources and primeval forest. The altitude within the study area ranges from 2800 to 4000 m. The four main forest types along this altitude gradient are: (1) dark coniferous forest between 3300 and 3900 m; (2) mixed coniferous and broad leaf forest between 3200 and 3300 m; (3) deciduous broad leaf forest between 3000 and 3200 m; and (4) azaleas and patches of dense bamboo (Fargesia spp.) forest between 2800 and 3600 m. The most common trees are firs (Abies spp.), larches (Larix potaninii var. macrocarpa), and azaleas (Rhododendron spp.), with some interspersed evergreen oaks (Quercus spp.), Chinese cherries (Betula alnoides), and sorbus trees (Sorbus spp.) (Yang 2000). Of these forest types, the Laojun Mountains are mostly composed of cold temperate coniferous forests dominated by Abies georgei and Rhododendron rubiginosum var. ptilostylum (Li et al. 2010); however, vegetation does vary with elevation.

Location of the Jinsichang study area in Laojun Mountain National Park
Presence data collection
We collected 891 occurrence records from on-the-ground field work. We used continuous data from 2010 to 2014 and some fragmented data in 2016 and 2017. Monkeys were tracked daily and data were recorded opportunistically using handheld global positioning system (GPS) devices and camera traps (Fig. 3). Free-ranging Yunnan snub-nosed monkeys are difficult to observe in Jinsichang due to the rugged terrain and because the monkeys are unhabituated to human observers. Occurrence data therefore included direct observations as well as indirect indicators of presence, including feces, feeding remains, and other traces found during on-the-ground surveys. Because absence data were not recorded, this study assessed habitat suitability based on direct and indirect observations (i.e., occurrences) and a suite of environmental variables. This study does not describe the frequency nor intensity of habitat use.

Yunnan snub-nosed monkeys at Jinsichang study area by camera trap
We used spatial filtering to account for potential bias in data collection. Using R, a grid (30-m resolution) was placed over a map of the study site. One occurrence point from each grid cell was randomly selected and retained for modeling; this reduced the occurrence points used in the final model to 664.
Predictor variables
Predictor (environmental) variables were generated using ArcMap 10.5 (ESRI 2011) and R. We initially prepared 34 environmental variables at 30-m spatial resolution. We selected predictor variables based on previous studies that addressed primate habitat suitability (Cayuela et al. 2009; Fitzgerald et al. 2018; Graham et al. 2011; Reiss et al. 2011; Théry 2001; Zhang et al. 2012) and to best address the hypothesized factors that influence Yunnan snub-nose monkey behavior. Climate variables (n = 19) were downloaded from the WorldClim database and resampled to 30-m spatial resolution (WorldClim-Global Climate Data 2018). In ArcMap, the Geomorphometry and Gradient Metrics Toolbox (Baig et al. 2014; Evans et al. 2014) was used to calculate variables that describe landscape structure (n = 11) based on a digital elevation model from The Advanced Spaceborne Thermal Emission and Reflection Radiometer (ASTER). Vegetation-specific variables (n = 4) were calculated from a Landsat 8 satellite image acquired on 13 December 2016. For details on each variable, refer to Table 1.
To reduce the effect of collinearity, we examined the correlation between predictor variables using Pearson’s product moment correlation (r). One variable from each pair of highly correlated variables (|r| > 0.70) was eliminated from the analyses. After accounting for correlation, the original suite of variables (n = 34) was reduced to 13.
Modeling approach
Previous research conducted on rare species has shown that a limited amount of data on species presence can be used to create SDMs if these occurrences are relatively unbiased (Hernandez et al. 2006; Pearson et al. 2007). Maxent 3.3.3 was used to model habitat suitability for Yunnan snub-nosed monkeys (Elith 2000; Phillips et al. 2004). Maxent is a program that facilitates SDM construction and estimates the probability of organismal presence (i.e., habitat suitability) given the interaction between user-defined predictor variables and organismal presence data (Elith et al. 2006; Yesson et al. 2012). Maxent performs well with presence-only data and frequently outperforms other SDM methods (Fourcade et al. 2014). The model was initially restricted to the study area but then projected into adjacent areas within Laojun Mountain NP to better estimate the extent of suitable habitat in the region. For the final model, including this projection, the area under the receiver-operating curve (AUC) was used to evaluate predictive performance. The AUC value measured the probability that a location known to be suitable for monkeys (recorded occurrences) was ranked higher than a randomly chosen background location (Davis and Goadrich 2006). AUC values range from 0 to 1; a value of 0.5 indicates random prediction and 1 indicates perfect predictive performance. Values over 0.70 are considered ecologically relevant (Fitzgerald et al., 2018; Araujo et al. 2005; Pearce and Ferrier 2000; Swets 1988). The results reveal a best-fit model that classifies locations according to habitat suitability. The model was replicated ten times to obtain a model average and standard deviation.
Results
Habitat suitability model
The final model was ecologically relevant, with an average AUC of 0.751 and standard deviation of 0.015. Figure 4 is a map that shows the distribution of suitable habitat for Yunnan snub-nosed monkeys in the Jinsichang study area and adjacent regions. The variables were ranked according to their influence on habitat suitability (i.e., permutation importance), as shown in Table 2. The top five variables were mean diurnal temperature range (31.6%), precipitation in the wettest quarter (30.4%), annual precipitation (17%), normalized difference vegetation index (NDVI) (5%), wetness (4.6%), and aspect (4.5%).

Final habitat suitability model for Yunnan snub-nosed monkeys in the Jinsichang study area and adjacent regions of Laojun Mountain National Park
Environmental variable response curves
The response curves for each of the top six environmental variables represent how habitat suitability (or probability of monkey presence) changes in response to changes in the environmental variables used in the model. To create these curves, each environmental variable was varied while keeping all others at their average sample value.
The environmental variable that contributed most to habitat suitability was mean diurnal temperature range (Fig. 5), which measures daily temperature fluctuation (O’Donnell and Ignizio 2012). Mean diurnal temperature range initially had a positive relationship with habitat suitability, which steadily increased with mean diurnal temperature ranges from 8.9 to 9.5 °C. After peaking at around 9.5 °C, habitat suitability showed a steep decrease as mean diurnal temperature range continued to increase.

Response curve plot generated by Maxent that shows the dependence of predicted habitat suitability on mean diurnal range. The curve shows the mean response of the ten replicate Maxent runs (red) and the standard deviation (blue)
The second most important variable was precipitation during the wettest 3 months of the year (Fig. 6). Habitat suitability was highest between 383 to 393 ml of precipitation. Next, annual precipitation showed peaks in habitat suitability between 840 and 855 ml (Fig. 7). Unlike the sharp rise in suitability seen at lower precipitation values, a gradual decrease occurred as annual precipitation increased beyond approximately 853 ml.

Response curve plot generated by Maxent that shows the dependence of predicted habitat suitability on precipitation in the wettest quarter. The curve shows the mean response of the ten replicate Maxent runs (red) and the standard deviation (blue)

Response curve plot generated by Maxent that shows the dependence of predicted habitat suitability on annual precipitation. The curve shows the mean response of the ten replicate Maxent runs (red) and the standard deviation (blue)
NDVI, a measure of greenness or photosynthetically active vegetation, was the next most important environmental variable (Fig. 8). The relationship between NDVI and habitat suitability was bimodal. The first peak in suitability occurred at low NDVI values, with a second peak occurring in the upper range of NDVI values. It should be noted that the standard deviation at low NDVI values was much greater than at higher values.

Response curve plot generated by Maxent that shows the dependence of predicted habitat suitability on normalized difference vegetation index. The curve shows the mean response of the ten replicate Maxent runs (red) and the standard deviation (blue)
Similar to some of the other environmental variables discussed above, there was a unimodal relationship between habitat suitability and wetness, or soil moisture content, which was the next most important variable in the model. There was an initial spike and peak in habitat suitability at wetness values near 0; then, as wetness increased, habitat suitability continuously dropped (Fig. 9).

Response curve plot generated by Maxent that shows the dependence of predicted habitat suitability on wetness. The curve shows the mean response of the ten replicate Maxent runs (red) and the standard deviation (blue)
The final variable of interest here is aspect, which is the direction in which a slope faces. Aspect can be divided into four compass directions based on the primary, cardinal directions: north (315° to 360° and 0° to 45°), east (45° to 135°), south (135° to 225°), and west (225° to 315°) (Fig. 10). Using these quadrants, the response curve for aspect showed that habitat suitability was highest for north-facing slopes and lowest for east- to southeast-facing slopes.

Response curve plot generated by Maxent that shows the dependence of predicted habitat suitability on aspect. The curve shows the mean response of the ten replicate Maxent runs (red) and the standard deviation (blue)
Discussion
In this study, the aim was to use SDM to provide information about the current distribution of Yunnan snub-nosed monkeys that can assist with the conservation of this endangered species. The results indicated that habitat suitability was influenced by several environmental variables. The most influential were mean diurnal temperature range (31.6%), precipitation during the wettest quarter of the year (30.4%), average annual precipitation (17%), normalized difference vegetation index (5%), wetness (4.6%), and aspect (4.5%).
It has been over a century since Grinnell (1917) described local factors that potentially affect species’ distributions, including food, vegetation, soil, breeding, climate, refuge sites, inter-specific interactions, and individual preferences. Until recently, however, our understanding and collection of relevant data remained temporally and spatially limited. With the introduction of and subsequent advances in remote sensing and geographic information systems (GIS), scientists can obtain a wide variety of spatial and temporal data to better understand species distributions and the environmental factors that affect these distributions at multiple scales (Hirzel and Le Lay 2008; Grinnell 1917). In parallel with these technological advances and increases in publicly available data, there has been a surge in the use of predictive modeling (e.g., habitat suitability modeling or species distribution modeling) to understand past, current, and future outlooks for species (Hirzel et al. 2006; Hirzel and Le Lay 2008). The predictive performances of several modeling techniques have been extensively examined to identify the optimum ways to map species distributions under different constraints (Brotons et al. 2004; Hirzel et al. 2006; Ferraz et al. 2012).
Using satellite remote sensing and GIS, we modeled the distribution of suitable habitat for Yunnan snub-nosed monkeys in the Jinsichang area of the Laojun Mountains in China. In addition, we examined some environmental variables hypothesized to influence habitat suitability. Species distribution modeling provides information on the distribution of suitable habitat for species of interest, important for the effective implementation of conservation strategies (Cayuela et al. 2009; Ferraz et al. 2012; Fourcade et al. 2014). In this study, the final model of habitat suitability was ecologically useful, with an average model fit of 0.751. The variables that contributed most to habitat suitability were mean diurnal temperature range, precipitation during the wettest quarter of the year, average annual precipitation, NDVI, wetness, and aspect. Our final model is a spatially explicit representation of habitat suitability for Yunnan snub-nosed monkeys that can easily be visualized and discussed with conservation practitioners that work in Laojun Mountain NP. For example, practitioners on the ground may realize that there are areas identified as highly suitable in the final model, but that these areas are actually devoid of Yunnan snub-nosed monkeys. This may result in refocusing or changing research and conservation efforts to better understand such areas and the reasons monkey are absent. Conversely, there may be highly suitable areas where species data are lacking or completely absent. We hope our model can be used to direct future research and efforts to widen our understanding of this species’ range and distribution.
Of the top six variables that had the greatest influence on habitat suitability, mean diurnal temperature range ranked highest. Mean diurnal temperature range is a measure of temperature fluctuation. Laojun Mountain NP is a highly seasonal temperate area with clear differences between summer and winter (Baoping et al. 2009; Liu and Zhao 2004; Tsuji et al. 2013). Based on our observations and research by Xiao et al. (2003), snow usually covers the ground from November to April, (up to 6 months) which may contribute to a large diurnal temperature range and few monkey occurrences above 3800 m. Habitat suitability was highest in areas where daily temperature fluctuated between only 9 °C and 9.5 °C throughout the year; beyond this, habitat suitability dropped rapidly. This finding indicates that Yunnan snub-nosed monkeys may have a specific tolerance range for temperature fluctuations that plays a significant role in shaping their distribution.
The second most important variable was total precipitation during the wettest 3 months of the year. The rainy season in Laojun Mountain NP lasts from July to early October. The most intense rainfall usually occurs between the end of July and the middle of August, when downpours can last several days; heavy rain and snow can impede monkey group movements and influence their ranging patterns (Baoping et al. 2009; Liu and Zhao 2004). Yunnan snub-nosed monkeys prefer areas with precipitation between 390 and 395 ml during the wettest months of the year, suggesting a specific tolerance or preference for places that receive moderate precipitation. This pattern was also seen in the next most important environmental variable—total annual precipitation. Although annual precipitation and precipitation of the wettest quarter are different measures, they both showed that these monkeys prefer moderate precipitation levels.
Additionally, the amount of photosynthetically active green vegetation (NDVI) in an area strongly influenced habitat suitability. The NDVI response curve was bimodal, indicating that Yunnan snub-nosed monkeys preferred areas with relatively low and high amounts of green vegetation, respectively. The monkeys are therefore selecting areas with specific vegetation types.
Yunnan snub-nosed monkeys are semi-arboreal, large-bodied, and more terrestrial than most other colobines (Long and Kirkpatrick 1994). They have been observed spending approximately 80.5% of their time in trees and 19.5% of their time on the ground (Ren et al. 2000; Li 2007). Areas with lush green vegetation appeared to be preferred for feeding and resting (pers. obs.). For example, Prunus conadenia, Sorbus spp., Acanthopanax evodiaefolius, and Padus brachypoda, all imported food resources (Li et al. 2011), are found in high NDVI areas. In contrast, areas with lower amounts of green vegetation possibly provided easier travel routes and food resources that met different nutritional needs of the monkeys. For example, many of the mountain ridges—part of the Danxia landform, where vegetation cover is generally very sparse—are natural corridors for monkeys to travel along (pers. obs.). Therefore, NDVI is an important index that provides insight into the behavioral ecology of these monkeys.
Wetness is a measure of soil moisture content that we predicted would influence Yunnan snub-nosed monkey behavior and ranging. In the study area, regions with low soil moisture content (i.e., low wetness values), such as mountain slopes and ridges, yielded high habitat suitability for Yunnan snub-nosed monkeys. Previous studies revealed that the monkeys preferred sunlit and warmer areas (Gittins 1982). Valley bottoms and areas near rivers, with high soil moisture content or wetness values, tend to receive less sunlight (pers. obs.), and therefore may also be less preferred by the monkeys. Even though valleys are potential sources of drinking water for monkeys, this is rarely observed. The monkeys may have other means to obtain water, such as eating snow (Matsuzawa 2017).
Finally, aspect—the direction that a slope faces—also played a large role in determining habitat suitability, with north-facing slopes having the highest suitability. Aspect plays a role in how much solar radiation and rain a slope receives. Both of these factors influence the composition and structure of the vegetation. For example, if prevailing winds travel east to west across a mountainous region, west-facing leeward slopes will experience a rain shadow effect in which rain is mostly deposited on the east-facing slopes (Tian et al. 2001; Böhner and Antonić 2009). The Yun Range runs north–south in northwestern Yunnan, between the Lancang River (Mekong) to the west and the Jinsha River (Yangtze) to the east. As a part of Yun Range, Laojun Mountain NP is affected by annual southwest monsoons from the Indian Ocean Bay of Bengal (Zhang 1998). These monsoons cause a rain shadow effect on most north-facing leeward slopes. Based on this observation, Usnea longissima and Bryora himalayana (lichen species), which represent 50.6% of the annual diet (Li et al. 2000) of Yunnan snub-nosed monkeys, grow vigorously on the leeward slopes. Moreover, the monkeys avoid strong wind, choosing mid-slope areas and areas in the shadow of ridges that provide protection from strong winds (Liu and Zhao 2004). Our results further confirm that Yunnan snub-nosed monkeys prefer areas that provide protection from intense winds and rains, such as north-facing, leeward slopes in Laojun Mountain NP.
The Yunnan snub-nosed monkey population is gradually increasing, but this monkey is still listed as endangered. A previous population study estimated that there is a sufficiently high ecological capacity in Laojun Mountain NP for these monkeys to survive and thrive (Li et al. 2009). This species forms highly cohesive groups based on multiple one-male units, and large groups require an area large enough to meet their nutritional requirements. Thus, habitat fragmentation and reduction may have strongly detrimental effects. Fragmentation by both natural terrain features and human activity may be an important determinant of regional species richness (Kirkpatrick et al. 1998; Dias and Strier 2003). Moreover, with increasing development of the local economy, ecotourism and ever-expanding anthropogenic activities may aggravate fragmentation. We recommend urgent consideration of the following: (1) strengthening protection and management of all areas with high habitat suitability, (2) promoting connectivity between suitable habitat patches, and (3) continuing surveillance and analyses for more robust conservation-related decision-making.
In conclusion, our analyses have limitations that should be addressed in future studies. One limitation of our study is the lack of data on anthropogenic activities in Laojun Mountain NP. Although we collected data on activities such as grazing, logging, poaching, and forest product collection, we did not do so systematically throughout the park. Thus, it is possible that anthropogenic activities influence snub-nosed monkey ranging and behavior, but our study was not able to factor such variables into the model. Future studies should investigate this further. Additionally, distribution of food resources is an environmental variable that we did not include, because previously data were not collected systematically throughout the study area (see Li et al. 2009; Yang 2000). Future studies can improve upon our model with greater consideration of food resource-related variables, incorporation of information on anthropogenic activity, and overall larger data sets.
Change history
23 November 2019
The correct name of the last author should be ‘‘Tetsuro Matsuzawa’’, and not ‘‘Tetsuro Matsuzwa’’ as given in the original publication of the article.
References
Araujo MB, Pearson RG, Thuillers W, Erhard M (2005) Validation of species-climate impact models under climate change. Glob Change Biol 11:1504–1513
Baig MHA, Zhang L, Shuai T, Tong Q (2014) Derivation of a tasseled cap transformation based on Landsat 8 at-satellite reflectance. Remote Sens Lett 5:423–431
Baoping R, Ming L, Yongcheng L, Fuwen W (2009) Influence of day length, ambient temperature, and seasonality on daily travel distance in the Yunnan snub-nosed monkey at Jinsichang, Yunnan, China. Am J Primatol 71:233–241
Bleisch W, Richardson M (2008) Rhinopithecus bieti. The IUCN Red List of Threatened Species 2008:e.T19597A8986243. http://doi.org/10.2305/IUCN.UK.2008.RLTS.T19597A8986243.en. Downloaded on 15 Sept 2019
Böhner J, Antonić O (2009) Land-surface parameters specific to topo-climatology. Dev Soil Sci 33:195–226
Brotons L, Thuiller W, Araújo MB, Hirzel AH (2004) Presence-absence versus presence-only modelling methods for predicting bird habitat suitability. Ecography 27:437–448
Cayuela L, Golicher DJ, Newton AC, Kolb M, De Alburquerque FS, Arets EJMM, Alkemade JRM, Pérez A (2009) Species distribution modeling in the tropics: problems, potentialities, and the role of biological data for effective species conservation. Trop Conserv Sci 2:319–352
Cui LW, Quan RC, Xiao W (2006) Sleeping sites of black-and-white snub-nosed monkeys (Rhinopithecus bieti) at Baima Snow Mountain, China. J Zool 270:192–198
Davis J, Goadrich M (2006) The relationship between precision-recall and ROC curves. In: Proceedings of the 23rd international conference on machine learning (pp. 233–240). ACM
Dias LG, Strier KB (2003) Effects of group size on ranging patterns in Brachyteles arachnoides hypoxanthus. Int J Primatol 24:209–221
Elith J (2000) Quantitative methods for modeling species habitat: comparative performance and an application to Australian plants. In: Ferson S, Burgman MA (eds) Quantitative methods in conservation biology. Springer, New York, pp 39–58
Elith J, Graham CH, Anderson RP, Dudik M, Ferrier S, Guisan A, Hijmans RJ, Huettmann F, Leathwick JR, Lehmann A, Li J, Lohmann LG, Loiselle BA, Manion G, Moritz C, Nakumura M, Nakazawa Y, Overton JMM, Peterson AT, Pillipes SJ, Richardson K, Scachetti-Pereira R, Schapire RE, Soberón J, Williams S, Wisz MS, Zimmermann NE (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
ESRI (2011) ArcGIS desktop: release 10.5. Environmental Systems Research Institute, Redlands
Evans JS, Oakleaf J, Cushman SA, Theobald D (2014) An ArcGIS toolbox for surface gradient and geomorphometric modeling, version 2.0-0. Laramie, WY. http://evansmurphy.wix.com/evansspatial
Fang J, Shen Z, Tang Z, Wang Z (2004) The protocol for the survey plan for plant species diversity of China’s mountains. Biodivers Sci 12:5–9
Ferraz KMPMB, Ferraz SFB, Paula RC, Beisiegel B, Breitenmoser C (2012) Species distribution modeling for conservation purposes. Nat Conserv 10:214–220
Fitzgerald M, Coulson R, Lawing AM, Matsuzawa T, Koops K (2018) Modeling habitat suitability for chimpanzees (Pan troglodytes verus) in the Greater Nimba Landscape, Guinea, West Africa. Primates 59:361–375
Fourcade Y, Engler JO, Rödder D, Secondi J (2014) Mapping species distributions with MAXENT using a geographically biased sample of presence data: a performance assessment of methods for correcting sampling bias. PLoS One 9(5):e97122
Franklin J (2009) Mapping species distributions: spatial inference and prediction. Cambridge University Press, Cambridge
Gittins SP (1982) Feeding and ranging in the agile gibbon. Folia Primatol 38:39–71
Graham CH, Loiselle BA, Velásquez-Tibatá J, Cuesta F (2011) Species distribution modeling and the challenge of predicting future distributions. Climate change and biodiversity in the Tropical Andes, Inter-American Institute for Global Change Research (IAI) and Scientific Committee on Problems of the Environment. Sao José dos Campos, Brazil, 295–310
Grinnell J (1917) Field tests of theories concerning distributional control. Am Nat 51:115–128
Grueter CC, Li D, Ren B, Wei F, Xiang Z, van Schaik CP (2009) Fallback foods of temperate-living primates: a case study on snub-nosed monkeys. Am J Phys Anthropol 140:700–715
Hallet B, Molnar P (2001) Distorted drainage basins as markers of crustal strain east of the Himalaya. J Geophys Res Solid Earth 106(B7):13697–13709
Hernandez PA, Graham CH, Master LL, Albert DL (2006) The effect of sample size and species characteristics on performance of different species distribution modeling methods. Ecography 29:773–785
Hirzel AH, Le Lay G (2008) Habitat suitability modelling and niche theory. J Appl Ecol 45:1372–1381
Hirzel AH, Le Lay G, Helfer V, Randin C, Guisan A (2006) Evaluating the ability of habitat suitability models to predict species presences. Ecol Model 199:142–152
Huadong G (2013) Three parallel rivers of Yunnan Protected Areas. In: Atlas of remote sensing for world heritage: China (pp. 288–293). Springer, Berlin
Huang ZP, Cui LW, Scott MB, Wang SJ, Xiao W (2012) Seasonality of reproduction of wild black-and-white snub-nosed monkeys (Rhinopithecus bieti) at Mt. Lasha, Yunnan, China. Primates 53:237–245
Jablonski NG (1998) The natural history of the doucs and snub-nosed monkeys. World Scientific Publishing, Singapore
Kirkpatrick RC, Long YC, Zhong T, Xiao L (1998) Social organization and range use in the Yunnan snub-nosed monkey Rhinopithecus bieti. Int J Primatol 19:13–51
Li Y (2007) Terrestriality and tree stratum use in a group of Sichuan snub-nosed monkeys. Primates 48:197–207
Li BG, Chen C, Ji WH, Ren BP (2000) Seasonal home range changes of the Sichuan snub-nosed monkey (Rhinopithecus roxellana) in the Qinling mountains of China. Folia Primatol 71:375–386
Li BG, Jia ZY, Pan RL, Ren BP (2003) Changes in distribution of the snub-nosed monkey in China. In: Marsh LK (ed) Primates in fragments: ecology and conservation. Kluwer, New York, pp 29–51
Li GQ, Yang YM, Xiao W (2007) A study on vegetation types of Rhinopithecus bieti habitat. J West China For Sci 36:95–98
Li L, Yu S, Ren B, Li M, Wu R, Long Y (2009) A study on the carrying capacity of the available habitat for the Rhinopithecus bieti, population at Mt. Laojun in Yunnan, China. Environ Sci Pollut Res 16:474–478
Li X, Long Y, Long C (2010) Study on the habitat of Rhinopithecus bieti at Jinsichang, Laojun Mountains, Yunnan province. Life Sc Res 14:44–49
Li DY, Ren BP, He XM, Hu G, Li BG, Li M (2011) Diet of Rhinopithecus bieti at Xiangguqing in Baimaxueshan National Nature Reserve. Acta Theriol Sinica 31:338–346
Liedigk R, Yang M, Jablonski NG, Momberg F, Geissmann T, Lwin N, Yongcheng L (2012) Evolutionary history of the odd-nosed monkeys and the phylogenetic position of the newly described Myanmar snub-nosed monkey Rhinopithecus strykeri. PLoS One 7(5):e37418
Liu ZH, Zhao QK (2004) Sleeping sites of Rhinopithecus bieti at Mt. Fuhe, Yunnan. Primates 45:241–248
Liu Z, Ren B, Wu R, Zhao L, Hao Y, Wang B, Li M (2009) The effect of landscape features on population genetic structure in Yunnan snub-nosed monkeys (Rhinopithecus bieti) implies an anthropogenic genetic discontinuity. Mol Ecol 18:3831–3846
Long Y, Kirkpatrick CR (1994) Report on the distribution, population, and ecology of the Yunnan snub-nosed monkey (Rhinopithecus bieti). Primates 35:241–250
Long YC, Zhong T, Xiao L (1995) Conservation strategy for the Yunnan snub-nosed monkey. In: Xia WP, Zhang YZ (eds) Primate research and conservation. China Forestry Publishing House, Beijing, pp 157–164
Ma C, Huang ZP, Zhao XF, Zhang LX, Sun WM, Scott MB, Wang XW, Cui LW (2014) Distribution and conservation status of Rhinopithecus strykeri in China. Primates 55:377–382
Matsuzawa T (2017) Monkeys and mountains in Yunnan, China. Primates 58:379–383
O’Donnell MS, Ignizio DA (2012) Bioclimatic predictors for supporting ecological applications in the conterminous United States. US Geol Surv Data Ser 691:1–10
Pearce J, Ferrier S (2000) Evaluating the predictive performance of habitat models developed using logistic regression. Ecol Model 133:225–245
Pearson RG, Raxworthy CJ, Nakamura M, Peterson AT (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeograph 34:102–117
Phillips SJ, Avenue P, Park F (2004) A maximum entropy approach to species distribution modeling. Proceedings of the Twenty-First International Conference on machine learning. ACM, New York, pp 655–662
Policy Research Office of the Peoples’ Government of Yunnan Province et al. (2006) The three parallel rivers. Yunnan Peoples’ Publishing House, Kunming
Reiss H, Cunze S, König K, Neumann H, Kröncke I (2011) Species distribution modelling of marine benthos: a North Sea case study. Marine Ecol Prog Series 442:71–86
Ren B (2009) Influence of day length, ambient temperature, and seasonality on daily travel distance in the Yunnan snub-nosed monkey at Jinsichang, Yunnan, China. Am J Primatol 71:233–241
Ren BP, Li BG, Zhang SY, Li M, Liang B (2000) Preliminary study on the go-to-ground activity by Sichuan golden monkeys Rhinopithecus roxellana. Acta Theriol Sinica 20:79–80
ROPGYP (Research Office of the Peoples Government of Yunnan Province) (2010) Yunnan National Park management office, and the Nature Conservancy, China office. Resource Book of Yunnan National Park Policy and Research. People’s Government of Yunnan, Kunming
Swets KA (1988) Measuring the accuracy of diagnostic systems. Science 240:1285–1293
Théry M (2001) Forest light and its influence on habitat selection. In: Speight MR (ed) Tropical forest canopies: ecology and management. Springer, Dordrecht, pp 251–261
Tian YQ, Davies-Colley RJ, Gong P, Thorrold BW (2001) Estimating solar radiation on slopes of arbitrary aspect. Agric For Meteorol 109:67–74
Tsuji Y, Hanya G, Grueter CC (2013) Feeding strategies of primates in temperate and alpine forests: comparison of Asian macaques and colobines. Primates 54:201–215
WorldClim-Global Climate Data (2018) http://worldclim.org/version2. Accessed Aug. 2018
Wu BQ (1993) Patterns of spatial dispersion, locomotion and foraging behaviour in three groups of the Yunnan snub-nosed langur (Rhinopithecus bieti). Folia Primatol 60:63–71
Wu Z (2006) The areal-types of seed plants and their origin and differentiation. Yunnan Science & Technology Press, Kunming
Xiang ZF, Sheng H, Lin W, Cui LW, Wen X, Tai RCQZ (2007) Distribution, status and conservation of the black-and-white snub-nosed monkey Rhinopithecus bieti in Tibet. Oryx 41:525–531
Xiao W, Ding W, Cui LW, Zhou RL, Zhao QK (2003) Habitat degradation of Rhinopithecus bieti in Yunnan, China. Int J Primatol 24:389–398
Yang SJ (2000) Habitat, diet, range use and social organization of Rhinopithecus bieti at Jinsichang. PhD Dissertation, Kunming Institute of Zoology, Chinese Academy of Sciences
Yesson C, Taylor ML, Tittensor DP, Davies AJ, Guinotte J, Baco A, Black J, Hall-Spencer JM, Rogers AD (2012) Global habitat suitability of cold-water octocorals. J Biogeograph 39:1278–1292
Zhang YG (1998) Several issues concerning vertical climate of the Hengduan Mountains. Resour Sci 20:12–19
Zhang MG, Zhou ZK, Chen WY, Ferry Slik JW, Cannon CH, Raes N (2012) Using species distribution modeling to improve conservation and land use planning of Yunnan, China. Biol Conserv 153:257–264
Acknowledgements
This study was financially supported by MEXT 24000001 and 16H06283, JSPS core-to-core CCSN, and JSPS leading graduate program U04 (PWS) to TM. Support from Kyoto University and The Nature Conservancy is gratefully acknowledged. The authors also acknowledge Lijiang Laojun Mountain Biodiversity Conservation Center, Yulong Forestry Bureau and Lijiang Laojun Mountain National Park Management Bureau Lijiang Laojun Mountain Community Patrol Team, and Yulong Wildlife Protection Association. We thank Mallory Eckstut, PhD, from Edanz Group (http://www.edanzediting.com/ac) for editing a draft of this manuscript.
Author information
Authors and Affiliations
Contributions
LJ, MF, SH, and TM conceived and designed this study. LJ, HL, and XY conducted the data collection. LJ and MF analyzed data and wrote the paper. LJ, MF, HL, YL, XL, SH, and TM reviewed and edited the manuscript. All authors read and approved the manuscript.
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
The original version of this article was revised due to a correction in the last author name in author group.
Electronic supplementary material
Below is the link to the electronic supplementary material.
About this article

Cite this article
Liu, J., Fitzgerald, M., Liao, H. et al. Modeling habitat suitability for Yunnan Snub-nosed monkeys in Laojun Mountain National Park. Primates 61, 277–287 (2020). https://doi.org/10.1007/s10329-019-00767-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-019-00767-4