
Abstract
Tropical forests and the biodiversity within them are rapidly declining in the face of increasing human populations. Resource management and conservation of endangered species requires an understanding of how species perceive and respond to their environments. Species distribution modeling (SDM) is an appropriate tool for identifying conservation areas of concern and importance. In this study, SDM was used to identify areas of suitable chimpanzee (Pan troglodytes verus) habitat within the Greater Nimba Landscape, Guinea, West Africa. This location was ideal for investigating the effects of landscape structure on habitat suitability due to the topographic variation of the landscape and the Critically Endangered status of the Western chimpanzee. Additionally, this is the only mountainous, long-term chimpanzee study site and little is known about the effects of topography on chimpanzee behavior. Suitable habitat was predicted based on the location of direct and indirect signs of chimpanzee presence and the spatial distribution of 12 biophysical variables within the study area. Model performance was assessed by examining the area under the curve. The overall predictive performance of the model was 0.721. The variables most influencing habitat suitability were the normalized difference vegetation index (37.8%), elevation (27.3%), hierarchical slope position (11.5%), surface brightness (6.6%), and distance to rivers (5.4%). The final model highlighted the isolation and fragmentation of chimpanzee habitat within the Greater Nimba Landscape. Understanding the factors influencing chimpanzee habitat suitability, specifically the biophysical variables considered in this study, will greatly contribute to conservation efforts by providing quantitative habitat information and improving survey efficiency.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In 1995, the IUCN (International Union for Conservation of Nature) placed the Pan troglodytes taxon in the red list category of Endangered species. This listing included all four subspecies of chimpanzees: Western (P.t. verus), Central (P.t. troglodytes), Eastern (P. t. schweinfurthii), and Nigeria-Cameroon chimpanzee (P.t. ellioti). Since 1995, populations have continued to decline (Humle et al. 2016a). The current estimate of the total population size of chimpanzees is approximately 200,000 individuals. This estimate indicates a 66% decline over a 30-year span (Kormos et al. 2003). Of the four subspecies, the western chimpanzee is the only subspecies listed as Critically Endangered (Humle et al. 2016a). Since 1990, the population size of western chimpanzees has declined approximately 80% (Kühl et al. 2017). The principal threats to western chimpanzees are habitat loss and/or degradation, hunting, and disease (Humle et al. 2016b).
Chimpanzees in Guinea are the largest remaining population of the western subspecies (Humle et al. 2016a; Kormos et al. 2003). In recognition of the negative effects of habitat destruction and loss of biodiversity, protected areas (PAs) have historically been established in Guinea. There are four PAs in Guinea: Massif du Ziama Strict Nature Reserve, Badiar National Park, Haut Niger National Park, and the Mount Nimba Strict Nature Reserve. An estimated 5–20% of the chimpanzee population in Guinea resides in these areas. The rest lives outside of PAs (Kormos et al. 2003).
Protected areas are impacted by human encroachment and neighboring land-use changes. From 2000 to 2012, the Mount Nimba Strict Nature Reserve (Mt. Nimba SNR hereafter) lost 1.5 km2 of forest within its boundary (approximately 1% of the reserve area) and 21.7 km2 within a 10-km buffer zone around the reserve (Allan et al. 2017). Laurance et al. (2012) found that changes both within and outside PAs influence ecosystem health. For example, changes in the landscape structure of areas surrounding PAs may increase area isolation and edge effects (Laurance et al. 2012). Increasing isolation of chimpanzee communities leads to reductions in gene flow, threatening healthy, viable populations (Morin et al. Morin et al. 1994). Moreover, competition for land and resources leads to increases in human–chimpanzee interaction and conflict (Hockings et al. 2015; McLennan and Hill 2012). For example, in Bossou, Guinea, where there are few forested areas, the chimpanzees rely heavily on cultivars (cassava, papaya, and bananas), terrestrial herbaceous vegetation (Zingeberaceae and Marantaceae families), and oil palm (Elaeis guineensis) during periods of fruit scarcity (Humle 2011). Many of these alternative food sources are in human settlements, so increased reliance accelerates human–chimpanzee conflict (Humle 2011). A decrease in forested areas due to human encroachment will result in an increase in the interaction of humans and chimpanzees.
To effectively protect chimpanzees and their habitats, it is important to understand how chimpanzees respond to their environment, so conservation efforts can focus on areas of highest importance for their long-term survival. Identifying the environmental factors that influence chimpanzee habitat selection is a critical component of developing effective conservation plans (Rushton et al. 2004). Species distribution models (SDMs) (also referred to as habitat suitability models, habitat models, ecological niche models, environmental niche models, etc.) are an informative way to evaluate the importance of environmental variables related to species distribution (Franklin 2009). Species distribution modeling provides a means for mapping chimpanzee habitat. The results of the modeling exercise can be used to guide reserve design, habitat management, and conservation planning. Species distribution models estimate conditions suitable for species survival by examining the relationships between species’ occurrence and associated environmental conditions.
Here, we use direct and indirect evidence of chimpanzee occurrences from fieldwork, medium-resolution remote sensing data, and SDMs to evaluate how the spatial distribution of biophysical variables relates to the distribution of the Seringbara chimpanzee communities in the Mt. Nimba SNR. This modeling effort allows us to test the hypothesis that vegetation is one of the most important factors influencing the occurrence of great apes (Jantz et al. 2016; Junker et al. 2012; Koops 2011; Koops et al. 2012a, b; Torres et al. 2010), specifically within the Seringbara chimpanzee communities. We will also explore the importance of other biophysical variables as they relate to the probability of Seringbara chimpanzee occurrence and compare our modeling results with relevant conservation efforts in the region. To do so, we will quantify and map the spatial distribution of biophysical variables within the study area using remotely sensed images, analyze the importance of each biophysical variable in modeling suitable chimpanzee habitat, and use SDMs to identify areas most suitable for chimpanzees within the Greater Nimba Landscape.
Methods
Study site
The Mt. Nimba SNR is a UNESCO World Heritage Site in Danger (World Heritage Committee 2017). The Mt. Nimba SNR encompasses most of the Nimba Mountain range in Guinea and parts of Côte d’Ivoire on the southeastern side of the mountain range. Covering approximately 175 km2, the reserve is dominated by wet, evergreen forests with diverse topographical features including rocky peaks, rough cliffs, bare granite, steep river valleys, high-altitude savannahs, and rounded hilltops (Guillaumet and Adjanohoun 1971; Koops 2011; Kormos et al. 2003). The reserve is home to a variety of flora and fauna, including the critically endangered endemic Mt. Nimba viviparous toad (Nimbaphrynoides occidentalis) and the Critically Endangered western chimpanzee (P.t. verus) (World Heritage Committee 2017).
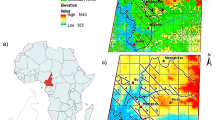
The study site (N07.634°, W08.425°), spanning 30 km2, is located on the Guinean side of the Nimba Mountains within the Mt. Nimba SNR (Fig. 1). The site is largely composed of primary tropical forests, but as the terrain becomes steeper, it transitions to a mosaic of terrestrial herbaceous vegetation, montane forest, and high-altitude grasslands (Koops 2011). The elevation ranges from 595 to 1511 m. The climate is characterized by a rainy season from February to October and a dry season lasting from November to February (Koops et al. 2012a, b, Koops et al. 2013). The site is adjacent to the small village of Seringbara, located about 6 km from Bossou at the foot of the Nimba Mountains (Koops 2011). Bossou is home to a community of chimpanzees (currently seven individuals) that have been the focus of research for over 30 years by the Kyoto University Primate Research Institute (KUPRI) (Matsuzawa and Humle 2011). The Mt. Nimba SNR and Bossou are separated by savannah that few chimpanzees traverse (Matsuzawa et al. 2011b). This study focuses on at least two communities of chimpanzees within the Mt. Nimba SNR, known as the Seringbara communities (Koops unpublished data). The Seringbara communities have been the focus of habituation efforts since 2003 (Koops 2011) and intermittent ecological studies and surveys since 1996 (Matsuzawa and Yamakoshi 1996; Humle and Matsuzawa 2001, 2004). The chimpanzees remain mostly unhabituated to humans (Koops 2011; Koops and Matsuzawa 2006; Matsuzawa et al. 2011a).

Location of the Seringbara study area on the Guinean side of the Mt. Nimba SNR in West Africa
Occurrence data
Between January 2012 and April 2014, a team of research assistants and local field assistants collected data on chimpanzee behavior at the Seringbara study site on the Guinean side of the Mt. Nimba SNR. Research teams maintained a nearly constant presence in the forest during this period, only missing data collection for 1–2 days a month. Field days focused on tracking and directly observing chimpanzees to obtain data on ranging, grouping, diet, nest building, and tool use. Direct observations of wild chimpanzees can be difficult, especially when communities are not fully habituated, such as the Seringbara communities. For this reason, nests, fecal samples, ant dipping sites, and feeding traces (i.e., wadges) were considered indirect indicators of chimpanzee presence and included as occurrence points along with direct chimpanzee sightings.
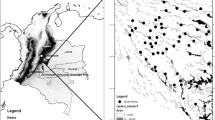
Direct and indirect evidence of chimpanzee presences were recorded using handheld global positioning system (GPS) devices during daily tracking of the chimpanzees. Sampling effort within the study area was comprehensive, as we covered the whole study area when searching for chimpanzees by splitting into teams and exhaustively surveying the study area using opportunistic sampling. In total, 1385 occurrence points were recorded. Occurrence points were not evenly distributed throughout the study area due to the behavior of the chimpanzees and perhaps also due to sampling bias. In a study comparing the different methods commonly used to correct for sampling bias, Fourcade et al. (2014) found that systematic spatial filtering consistently outperformed most other methods regardless of the species or type of bias. Systematic spatial filtering uses a grid of a user-defined cell size and randomly keeps one occurrence point per cell. We used R 3.3.2 (Supplementary Appendix A) to place a grid (30-m resolution) over the study area and randomly select one occurrence point from each grid cell. The total occurrence points were filtered and reduced to N = 947 for use in the final model (Fig. 2). Filtering to include only one occurrence point per cell did not influence our results because this study does not address the frequency nor magnitude of use by chimpanzees. Absence data were not available for this study. In addition, we chose to combine direct and various indirect types of evidence of occurrence for modeling, because (1) we wanted a robust sample size and classifying occurrences into behavior categories would drastically decrease sample size for the model and (2) it is unclear the behavior category for which we would attribute the feces occurrences, given that they can be found at feeding locations, along movement routes, at resting spots, as well as other locations of use (Supplementary Appendix A Table 1).

Location of chimpanzee occurrence points used in the model (N = 947)
Predictor variables
Raster layers of predictor variables (Table 1) dealing with landscape structure and land cover, herein referred to as biophysical variables, were prepared at a 30-m spatial resolution. An initial set of 17 biophysical variables (Table 1) was assessed, as detailed below, before being narrowed down to 12 variables in the final model. Minimum distance, supervised classification of a Landsat 8 image, obtained during the study period (December 26, 2013), was developed in ENVI 5.0.2 to delineate five land-cover types: dense forest, mixed forest, bare ground, village, and savannah (Supplementary Appendix B). These five classes were chosen based on expert knowledge of the region after analyzing the spectral groupings of the supervised classification. Dense forests consist of mostly primary, undisturbed forest comprised of tree species such as Parinari excelsa, Parkia bicolor, Antiaris africana, and Aningeria altissima. Mixed forests are mostly secondary, disturbed forests with less dense vegetation and less canopy cover. Tree species common in mixed forests include Musanga cecropioides, Elaeis guineensis, and Uapaca sp. Bare ground includes cleared areas, sparsely vegetated grasslands, and bare rock. Savannah consists of very dense, tall grass areas lacking trees. The village class includes buildings, huts, and other anthropogenic structures interspersed with bare ground. The minimum distance land-cover classification procedure performed well (overall accuracy of 90.78%) in distinguishing between macrohabitats, such as savannah and forest, but was not able to distinguish microhabitats, such as vegetation types, at the spatial resolution of the image (30 m) (Supplementary Appendix B). Because chimpanzees have sophisticated mental mapping capabilities (Ban et al. 2014; Boesch and Boesch 1984; Normand et al. 2009; Normand and Boesch 2009) and are able to perceive their surroundings at the level of individual trees and forest patches, vegetation indices were calculated to capture differences at microscales (Pintea et al. 2003; Torres et al. 2010). Landsat 8 imagery from six different dates within the data collection period was used to derive an average normalized difference vegetation index (NDVI) raster. NDVI is an indication of relative biomass (i.e., healthy, photosynthetically active vegetation) within each raster cell and can range from − 1 (water or bare ground) to 1 (healthy, dense vegetation). It is calculated from the near-infrared and red bands of a satellite image ((NIR – R)/(NIR + R)) (Campbell and Wynne 2011). In addition, we captured microhabitat characteristics within the study area using a tasseled cap transformation of the original Landsat 8 image. This process transforms the original spectral data into a new coordinate system with four orthogonal axes (Campbell and Wynne 2011). Each of these axes carries specific information that can be interpreted as (1) soil and surface brightness (brightness), (2) photosynthetically active vegetation (greenness), (3) soil moisture (wetness), and (4) atmospheric noise (Crist and Cicone 1984).
Studies of the Seringbara chimpanzees (Koops 2011; Koops et al. 2007, 2012a, b, 2013, 2015), as well as other non-human primates (Clee et al. 2015; Gregory et al. 2014; Hickey et al. 2013; Plumptre et al. 2010; Serckx et al. 2016; Torres et al. 2010; Wich et al. 2012), indicate that climate, vegetation, and anthropogenic factors may play a significant role in identifying suitable habitat. In particular, the dietary preferences of Seringbara chimpanzees indicate that the availability of fruit affects their ranging patterns (Koops 2011; Koops et al. 2013). Many of the tree species producing fruit utilized by the chimpanzees occur in primary forests and at elevations higher than 800 m (e.g., Parinari excelsa) (Koops 2011). Moreover, the Seringbara chimpanzees prefer to nest at locations with lower humidity (Koops et al. 2012a, b). For example, they tend to nest at higher altitudes (above 1000 m) where relative humidity is low and avoided nesting in areas of high humidity (below 800 m) (Koops et al. 2012a, b). Therefore, the other biophysical variables included in the initial model were chosen for their ability to serve as proxies for these (i.e., climate, vegetation, and anthropogenic) factors (Franklin 2009).
The following variables were generated using ArcMap 10.2.2 (ESRI 2011) and R 3.3.2 (R Core Team 2005) and derived from a digital elevation model (DEM) from the Advanced Spaceborne Thermal Emission and Reflection Radiometer (ASTER) Global Digital Elevation Model Version 2 (NASA 2009): elevation, slope, aspect, topographic position index, roughness, integrated moisture index, heat load index, landform curvature, compound topographic index surface relief, and hierarchical slope position (Table 1). The R script for calculating hierarchical slope position is found in Supplementary Appendix C. The distance to rivers variable was generated in ArcMap 10.2.2 using a shapefile of rivers within the Greater Nimba Landscape and calculating the Euclidean distance of each 30-m2 cell from the nearest permanent river or stream.
We examined correlation between variables to reduce the effect of collinearity on interpreting Maxent results (Dormann et al. 2013; Kumar et al. 2014; Rödder et al. 2013). Correlation was calculated using Pearson’s product moment correlation (r). For a set of highly correlated variables (|r| > 0.7), the variable with the highest predictive power (training gain), in the preliminary model using all 17 biophysical variables, was retained (Dormann et al. 2013; Estes et al. 2010; Hickey et al. 2013).
Modeling technique
To map suitable chimpanzee habitat and analyze biophysical variables contributing to suitability, we used Maxent 3.3.3 software based on the maximum entropy framework (Phillips et al. 2004). Maxent estimates relative probability of species presence given data on occurrence and user-selected predictor variables (Franklin 2009; Phillips et al. 2006). Maxent performs well with presence-only data and frequently outperforms other SDM methods (Elith et al. 2006, 2011; Phillips and Dudík 2008; Wilson et al. 2013). The result is a best-fit model classifying locations in the study area according to probability of presence (0–1, with 1 indicating highest probability of presence). The model’s predictive performance is evaluated using the area under the receiver operating characteristic curve (AUC). AUC was chosen over other evaluation measures because it does not require an arbitrary selection of a threshold (Phillips et al. 2006). For presence-only data, AUC describes the probability that the model scores a presence site higher than a background site (Phillips et al. 2009). An AUC of 1 indicates perfect predictive power and an AUC of 0.5 indicates random prediction. A model with a high AUC, such as 0.70, indicates that there is a greater-than-random chance that a randomly selected presence site will be given a higher value than a randomly selected background site (Elith et al. 2006). Thus, a model with a high AUC has more discriminative power. A k-fold cross-validation procedure was replicated ten times to obtain a mean AUC value for the final model (Dormann et al. 2013; Kumar et al. 2014; Wich et al. 2012). Additionally, Maxent was used to generate response curves showing the relationship between each predictor variable and predicted probability of chimpanzee presence (i.e., predicted habitat suitability). Percent contribution and permutation importance were reported for each variable. Percent contribution is a measure of the amount of explained variance each variable contributes to the model. Permutation importance is a measure of how AUC changes when a variable is removed from the model and it is not sensitive to the order variables are put into the model (Songer et al. 2012; Wilson et al. 2013).
The model was projected beyond the study area to better assess habitat suitability for the Seringbara chimpanzees within the larger landscape. This geographically projected model is hereafter referred to as the final model. The extent, referred to as the Greater Nimba Landscape for this study, includes the majority of the Nimba Mountain range in Guinea, Liberia and Côte d’Ivoire, as well as the regions surrounding a few of the closest villages to the study site and an iron ore mining site (Fig. 3). The total area of the Greater Nimba Landscape is 992 km2. By including these villages, namely Bossou, Seringbara, Nyon, and Zuoguepo, and their near surroundings, the model is better able to capture the landscape heterogeneity of the region and its influence on habitat suitability beyond the protected area. It is important to note a few limitations of projecting, or transferring, a model into a geographic region where data were not collected (Warren and Seifert 2011). One issue in model transferability is the difference in predictor variable ranges between the sampled area and the area into which the model is projected (i.e., Greater Nimba Landscape). If the ranges in the sampled region are narrower, it can cause the response curves to be truncated (Randin et al. 2006). In addition, transferring a model can reduce the model’s predictive ability in the new region (Eger et al. 2016). For this reason, the results from the study should be interpreted carefully while keeping these limitations in mind.

Landsat 8 satellite image of the Greater Nimba Landscape
Since absence data were not available, a maximum of 10,000 background points were randomly generated to represent the availability and range of environmental conditions within the study area (Wilson et al. 2013). A minimum-convex polygon around the occurrence points was created to restrict background point generation to only the area covered while collecting data in the field. This procedure ensures that sampling of background points is restricted to the same region from which occurrence points were collected and helps account for sampling bias (Phillips et al. 2009).
The final Maxent output is a gradient model classifying each pixel according to probability of presence or habitat suitability. In many cases, SDMs are converted to binary models, delineating suitable versus unsuitable habitat, which are used by conservationists and land managers (Fourcade et al. 2014; Escalante et al. 2013). Reclassification to create a binary model requires the identification of a threshold, above which a location is considered suitable for a species (Liu et al. 2005). There is not a single method for threshold selection that is better than all others regardless of the species or study objective (Liu et al. 2005). For this study, we reclassified the final model output to create binary maps of habitat suitability for the Seringbara chimpanzees based on three commonly used threshold selection approaches: minimum training presence, 10 percentile training presence, and equal training sensitivity and specificity (Escalante et al. 2013; Fourcade et al. 2014; Norris 2014; Pearson et al. 2007). The purpose of these binary maps was to visually and quantitatively assess the amount of suitable and unsuitable habitat for the Seringbara chimpanzees in the Greater Nimba Landscape, while also emphasizing the importance of carefully choosing a threshold approach.
Results
Correlation analysis
The following variables were highly correlated (|r| > 0.7): TPI and curvature (r = 1), slope and roughness (r = 0.85), NDVI and greenness (r = 0.91), NDVI and wetness (r = 0.74), wetness and greenness (r = 0.74), and LCC and wetness (r = − 0.72) (Supplementary Appendix D). For each highly correlated pair, the variable retained in the test models was chosen because it had the higher permutation importance when an initial model was run using all variables. Thus, the final model was created using only 12 of the original 17 biophysical variables: NDVI, elevation, HSP, brightness, DTR, aspect, HLI, CTI, IMI, roughness, curvature, and relief (Table 1).
Gradient Habitat Suitability Model
The fit of the final chimpanzee habitat suitability model for the Greater Nimba Landscape was 0.721 with a standard deviation of 0.023. Models with AUC values greater than 0.70 are considered to have fair discriminative abilities and are ecologically useful (Araujo et al. 2005; Pearce and Ferrier 2000; Swets 1988). The resulting map from the final model (Fig. 4) highlights areas of highest predicted suitability for chimpanzee habitat. The biophysical variables contributing most to the model, as measured by permutation importance, were NDVI (37.8%), elevation (27.3%), HSP (11.5%), brightness (6.6%), and DTR (5.4%) (Table 2).

Chimpanzee habitat suitability model showing the geographic distribution of suitable chimpanzee habitat throughout the Greater Nimba Landscape. This is a gradient model displaying habitat suitability on a scale from 0 (low suitability) to 1 (high suitability). This figure illustrates the importance of the Mt. Nimba SNR in providing habitat for chimpanzees within the Greater Nimba Landscape
Variable response curves
The spatial distributions for the biophysical variables of highest importance were mapped and displayed above the corresponding response curves (Fig. 5). The response curve for NDVI shows a positive relationship between probability of presence and NDVI, as healthy, photosynthetically active vegetation increases, so does the probability of chimpanzee presence (Fig. 5a). The response curve for elevation shows that probability of presence is highest between 800 and 1200 m (Fig. 5b). The response curve for hierarchical slope position indicates that probability of presence fluctuates in mildly exposed areas (HSP values between 0.3 and 0.65), whereas probability of presence is relatively low in valley bottoms and toe slopes (low HSP values) and is lowest in topographically exposed areas, such as cliff faces and ridges (high HSP values) (Fig. 5c). For brightness, probability of chimpanzee presence peaks at an index value of 0.35 before declining sharply at higher brightness values (Fig. 5d). There is a negative relationship between DTR and probability of presence, with a sharp decline in probability of presence for areas farther than 500 m from a river (Fig. 5e). Response curves and maps for all other biophysical variables used in the final model can be found in Supplementary Appendix E.



Plots of the response curves showing the dependence of probability of presence on a given biophysical variable. Each plot represents a Maxent model using only the corresponding variable. The plots are given for the five biophysical variables with highest permutation importance (percent shown on plot). The plots show the average response (red line) and the standard deviation (blue interval around the average). X-axes show the units of the corresponding variable. Y-axes indicate the logistic output. The maps above each response curve illustrate the spatial distribution of the biophysical variable in the Greater Nimba Landscape
Binary habitat suitability models
The final model was reclassified to create three binary models based on different threshold levels: minimum training presence (0.08), 10 percentile training presence (0.33), and equal training sensitivity and specificity (0.46) (Fig. 6a–c, respectively). Using a threshold allowed the amount of suitable versus unsuitable habitat to be delineated and quantified within the Greater Nimba Landscape (992 km2) (Table 3). For the minimum training presence threshold (0.08), 42% of the landscape was classified as suitable and 58% was classified as unsuitable for the Seringbara chimpanzees. The equal training sensitivity and specificity threshold (0.46) lends itself to a different interpretation of the Greater Nimba Landscape, as only 3% was classified as suitable habitat and 97% was unsuitable. Similarly, the 10% training presence threshold (0.33) delineated 7% of the Greater Nimba Landscape as suitable and 93% as unsuitable.

Final model output showing the distribution of suitable chimpanzee habitat throughout the Greater Nimba Landscape as a series of binary models of three different threshold values: a minimum training presence, b 10% training presence, and c equal training sensitivity and specificity
Discussion
Data on habitat requirements of chimpanzees are needed for effective management and conservation. Constant advancements in technologies, such as remote sensing and GIS, combined with modeling techniques, such as Maxent, allow researchers to assess the influences on habitat suitability for many different species. In this study, we modeled the habitat suitability for the Seringbara chimpanzees in the Greater Nimba Landscape and identified the most important biophysical variables contributing to habitat suitability. The results indicate that NDVI, elevation, hierarchical slope position, brightness, and distance to rivers contributed most to predicted habitat suitability (Table 1).
The most important variable in predicting chimpanzee habitat suitability was NDVI. This index indicates the presence of photosynthetically active vegetation (Campbell and Wynne 2011). The positive relationship between NDVI and probability of occurrence suggests that chimpanzees prefer forested areas with dense, healthy vegetation (Fig. 5a). A study by Koops et al. (2012a, b) showed that the Seringbara chimpanzees prefer larger trees with dense leaf cover in primary forests to build nests. In addition, many of the tree species, utilized by the chimpanzees for feeding, are found predominantly in primary forests (e.g., Parinari excelsa, Parkia bicolor, Antiaris africana, and Aningeria altissima) (Koops 2011). This relation indicates that the habitat suitability model presented here is capturing important biological signals from the Seringbara chimpanzees’ use of the landscape. Similar studies at other locations have also shown that vegetation influenced chimpanzee behavior (Jantz et al. 2016; Torres et al. 2010), as well as great ape behavior in general (Junker et al. 2012).
Elevation was the second most important biophysical variable in predicting habitat suitability for the Seringbara chimpanzees. The relationship between elevation and the probability of Seringbara chimpanzee occurrence is bell shaped (Fig. 5b). Increasing elevation up to 900 m is associated with increasing probability of occurrence. Above 900 m, increasing elevation is associated with decreasing probability of occurrence. Within the Greater Nimba Landscape, elevation serves as a good proxy for climate and vegetation, as well as anthropogenic disturbance. Unfortunately, there is not sufficient data on anthropogenic disturbance for the whole reserve, but based on personal observations, we noticed that many of the villages and cultivated fields surrounding the study site are all located below 700 m. Thus, as elevation increases, so does the distance from anthropogenic disturbance. In addition, the protected status of the Nimba Mountains increases this effect because the mountains are within high-elevation areas. Although protected status does not directly indicate a lack of anthropogenic disturbance, the Mt. Nimba Strict Nature Reserve is remote, hunting pressures tend to decrease with distance from villages, and illegal hunting is targeted at animals other than the Seringbara chimpanzees (pers. obs., Koops and Fitzgerald). Moreover, as elevation increases above 1200 m, the landscape is dominated by high-altitude grasslands (Lamotte 1998), which may not provide ample resources for chimpanzees (Koops 2011). Thus, resulting in the bell-shaped curve of the relationship between elevation and probability of Seringbara chimpanzee occurrence.
The HSP (a measure of topographic exposure) was the next most important variable in predicting chimpanzee presence. Topographic exposure is the degree to which a location is surrounded by high relief terrain. A high HSP value indicates that a location is not surrounded by areas of higher relief (i.e., exposed), such as a cliff face or ridge top. A low value indicates that the landscape is surrounded by high relief terrain (i.e., not exposed), such as valley bottoms and toe slopes. The relationship between topographic exposure and the probability of Seringbara chimpanzee occurrence is generally negative, where an increase in topographic exposure is associated with a decrease in the probability of occurrence (Fig. 5c). Thus, Seringbara chimpanzees are more likely to occur in less-exposed areas, such as mild slopes, not surrounded by high relief terrain. Exposure can serve as a proxy for temperature and vegetation similar to the other important biophysical features, but it might also relate to the ease of movement through an area. Non-human primates have been found to distinguish between topographic features when traveling. For example, Gregory et al. (2014) found that bearded saki monkeys use ridge tops and slopes near ridges, because it may reduce the energetic cost of travel and/or serve a function in route-based mental mapping. This behavior is yet to be explored for chimpanzees in the Greater Nimba Landscape. Future studies examining the role of topography in chimpanzee movement would contribute greatly to our understanding of their perception and utilization of the landscape.
Another important variable in predicting chimpanzee habitat suitability was the tasseled cap brightness index. As brightness values increase, it indicates an increase in open canopy and an increase in bare ground (Campbell and Wynne 2011; Cohen and Goward 2004; Cohen et al. 1995). Cohen et al. (1995) showed that closed forest stands tend to have moderate brightness values. Previous studies from other chimpanzee research sites indicate that mature, closed forests are preferred by chimpanzees (Torres et al. 2010). Thus, the results from this study, showing highest probability of presence at moderate brightness values support previous findings. Nevertheless, caution must be taken when interpreting brightness values, because this index is responsive to topographic variation in addition to forest condition (Cohen and Goward 2004). For example, in our study site, some of the high savannah areas have very low brightness values despite very minimal canopy cover (Fig. 5d). Other very similarly vegetated savannah regions have much higher brightness values. Thus, the low brightness value in some high savannah areas might be explained by the steepness of the terrain and the incidence angle of the radar from the satellite collecting the image (Cohen et al. 1995).
Habitat suitability is also affected by the proximity of an area to the nearest river. As distance increases, the probability of chimpanzee presence decreases. This biophysical variable may serve as a proxy for vegetation (Hickey et al. 2013; Koops 2011). In evaluating the distribution of the variable distance to river throughout the Greater Nimba Landscape (Fig. 5d), many of the places that are more than 500 m from rivers are in the high savannah areas of the Nimba Mountains or in areas outside of the Mt. Nimba SNR, where the terrain is slightly flatter and rivers are more dispersed. Riverine areas may also provide food resources not available elsewhere in the landscape (Koops pers. comm.).
The final habitat suitability model illustrates the isolation of high suitability areas within the Greater Nimba Landscape. The areas of highest predicted habitat suitability for the Seringbara chimpanzees are located almost entirely within the Nimba mountain range. This is highlighted in the binary classification of the habitat suitability map into areas of suitable and not suitable habitat based on various threshold values (Fig. 6). A comparison of the three binary models also highlights the importance of carefully selecting a threshold value. In this study, the amount of suitable habitat within the landscape ranged for 3–42% (Table 3). This variation in amount of suitable habitat based on threshold values may result in very different conservation strategies and threshold selection should be carefully considered based on local knowledge, research, and specific conservation goals. Although binary models can be arbitrary and over simplify the landscape for behaviorally flexible and dynamic species that may not perceive the landscape in binary terms, the ability to identify suitable versus unsuitable habitat is useful for conservation practitioners (Escalante et al. 2013; Ferrer-Sánchez and Rodríguez-Estrella 2016; Liu et al. 2005). For example, Torres et al. (2010) delineated suitable from unsuitable chimpanzee habitat to assess changes in habitat suitability over time as well as temporal changes in the most important ecogeographical factors influencing chimpanzee habitat in Guinea-Bissau. Their results provide a basis for practitioners to adapt their strategies based on past changes as well as forecasted changes to chimpanzee habitat suitability.
Additionally, within the Nimba mountain range, high suitability areas are fragmented by terrain features such as high ridgelines and anthropogenic disturbances, such as the iron-ore mining concession in the NE region of the Nimba mountain range (Fig. 4). Thus, not only are the Seringbara chimpanzees isolated from other chimpanzee communities outside the Mt. Nimba SNR, they are at risk of becoming isolated from other communities within the Mt. Nimba SNR. Isolation and fragmentation of suitable habitat hampers gene flow between groups and can lead to further decline in chimpanzee populations in the region.
Maintaining viable, healthy chimpanzee populations requires movement between communities, thus the creation of corridors is one solution to restoring connectivity. One of the current efforts in the Greater Nimba Landscape is the Green Corridor Project. This project was established in 1997 with the aim of connecting chimpanzee populations in Bossou with those in the Nimba Mountains by planting trees species utilized by chimpanzees in the savannah between the sites (Matsuzawa et al. 2011b). Despite difficulties with fires, the Green Corridor Project has made and continues to make progress. One sign of this progress was the video recording of two male chimpanzees traveling into the corridor and the use of the corridor by monkeys (“The Green Corridor Project” 2017). The project is ongoing and technologies such as remote sensing (e.g., use of unmanned aerial vehicles and satellite imagery) and modeling may prove useful for monitoring and expanding the corridor. In addition, as the vegetation in the corridor matures, its NDVI value will increase. NDVI was the most important biophysical variable in our model and increasing NDVI was related to increasing probability of occurrence. Our modeling effort supports the hypothesis that the corridor will increasingly provide more suitable habitat for chimpanzees as the vegetation within the corridor matures. Future plans for new corridors might additionally consider locations with low topographic exposure that are near rivers.
While conservation efforts can use the methods and results from this study and expert knowledge of the region to more effectively and efficiently promote the long-term viability of chimpanzees in the region, these efforts should also recognize the limitations of this study. Since the model was projected into a novel geographic area where data on chimpanzee occurrences were not collected, the response curves may not encompass the full range of variables. In other words, interpretation of how the probability of chimpanzee presence will respond to a predictor variable beyond the range of the collected data is unknown. This is a limitation for many predictive SDM studies, yet there are few generally applicable solutions (Eger et al. 2016; Elith et al. 2010; Peterson et al. 2007; Zurrell et al. 2012). Future research might be able to mitigate this by surveying more areas within the greater landscape so the sampling effort is more representative of the range in predictor variables. Moreover, given that vegetation and proxies for vegetation greatly influence chimpanzee habitat suitability, this model might be improved with data that are better able to capture vegetation characteristics at a higher spatial resolution. Likewise, the model results could be improved by (1) additional surveys in the Greater Nimba Landscape beyond the study area used to create the model, (2) systematic data pertaining to anthropogenic disturbance, and (3) ground truthing of the variables used and results.
Conclusions
In conclusion, this study demonstrates that species distribution modeling is a useful tool for identifying suitable chimpanzee habitat within montane rainforests. More specifically, the results indicate that (1) biophysical variables quantifying the landscape structure within the Greater Nimba Landscape were useful predictors of chimpanzee presence, (2) NDVI, elevation, hierarchical slope position, brightness, and distance to rivers had the greatest influence on habitat suitability for the Seringbara chimpanzees, (3) suitable chimpanzee habitat within the Greater Nimba Landscape is fairly isolated and does not make up a large portion of the landscape, and (4) enforcing the protection of the Mt. Nimba SNR and adjacent areas is vital to supporting chimpanzee populations.
References
Allan JR, Venter O, Maxwell S et al (2017) Recent increases in human pressure and forest loss threaten many Natural World Heritage Sites. Biol Conserv 206:47–55
Araujo MB, Pearson RG, Thuillers W, Erhard M (2005) Validation of species-climate impact models under climate change. Glob Change Biol 11:1504–1513
Baig MHA, Zhang L, Shuai T, Tong Q (2014) Derivation of a tasselled cap transformation based on Landsat 8 at-satellite reflectance. Remote Sens Lett 5:423–431
Ban SD, Boesch C, Janmaat KRL (2014) Taï chimpanzees anticipate revisiting high-valued fruit trees from further distances. Anim Cogn 17:1353–1364
Blaszczynski JS (1997) Landform characterization with geographic information systems. Photogramm Eng Remote Sens 63:183–191
Boesch C, Boesch H (1984) Mental maps in wild chimpanzees: an analysis of hammer transports for nut cracking. Primates 25:160–170
Bolstad P, Lillesand T (1992) Improved classification of forest vegetation in Northern Wisconsin through a rule-based combination of soils, terrains, and Landsat thematic mapper data. For Sci 38:5–20
Campbell JB, Wynne RH (2011) Introduction to remote sensing, 5th edn. The Guilford Press, New York
Clee PRS, Abwe EE, Ambahe RD, Anthony NM, Fotso R, Locatelli S et al (2015) Chimpanzee population structure in Cameroon and Nigeria is associated with habitat variation that may be lost under climate change. BMC Evol Biol 15:1–13
Cohen WB, Goward SN (2004) Landsat’s role in ecological applications of remote sensing. Bioscience 54:535–545
Cohen WB, Spies TA, Fiorella M (1995) Estimating the age and structure of forests in a multi-ownership landscape of western Oregon, U.S.A. Int J Remote Sens 16:721–746
Crist EP, Cicone RC (1984) A Physically-based transformation of thematic mapper data: the TM Tasseled Cap. IEEE Trans Geosci Remote Sens GE 22:256–263
De Reu J, Bourgeois J, Bats M, Zwertvaegher A, Gelorini V, De Smedt P et al (2013) Application of the topographic position index to heterogeneous landscapes. Geomorphology 186:39–49
Dormann CF, Elith J, Bacher S, Buchmann C, Carl G, Carré G et al (2013) Collinearity: a review of methods to deal with it and a simulation study evaluating their performance. Ecography 36:027–046
Eger AM, Curtis JM, Fortin M-J, Côté IM, Guichard F (2016) Transferability and scalability of species distribution models: a test with sedentary marine invertebrates. Can J Fish Aquat Sci 74:766–778
Elith J, Graham CH, Anderson RP, Dudik M, Ferrier S, Guisan A et al (2006) Novel methods improve prediction of species’ distributions from occurrence data. Ecography 29:129–151
Elith J, Kearney M, Phillips S (2010) The art of modelling range-shifting species. Methods Ecol Evol 1:330–342
Elith J, Phillips SJ, Hastie T, Dudík M, Chee YE, Yates CJ (2011) A statistical explanation of MaxEnt for ecologists. Divers Distrib 17:43–57
Escalante T, Rodríguez-Tapia G, Linaje M, Illoldi-Rangel P, González-López R (2013) Identification of areas of endemism from species distribution models: threshold selection and Nearctic mammals. TIP Revista Especializada en Ciencias Químico-Biológicas 16:5–17
ESRI (2011) ArcGIS Desktop: Release 10.2.2. Environmental Systems Research Institute, Redlands
Estes LD, Reillo PR, Mwangi AG, Okin GS, Shugart HH (2010) Remote sensing of structural complexity indices for habitat and species distribution modeling. Remote Sens Environ 114:792–804
Ferrer-Sánchez Y, Rodríguez-Estrella R (2016) How rare species conservation management can be strengthened with the use of ecological niche modelling: the case for endangered endemic Gundlach’s Hawk and Cuban Black-Hawk. Glob Ecol Conserv 5:88–99
Fourcade Y, Engler JO, Rödder D, Secondi J (2014) Mapping species distributions with MAXENT using a geographically biased sample of presence data: a performance assessment of methods for correcting sampling bias. PLoS One 9:1–13
Franklin J (2009) Mapping species distributions: Spatial inference and prediction. Cambridge University Press, United Kingdom
Gessler PE, Moore ID, McKenzie NJJ, Ryan PJ (1995) Soil-landscape modelling and spatial prediction of soil attributes. Int J Geogr Inf Syst 9:421–432
Gregory T, Mullett A, Norconk MA (2014) Strategies for navigating large areas: a GIS spatial ecology analysis of the bearded saki monkey, Chiropotes sagulatus, in Suriname. Am J Primatol 76:586–595
Guillaumet J, Adjanohoun E (1971) Le milieu naturelle de la Côte-d’Ivoire. Mémoire ORSTOM 50:157–264
Guisan A, Weiss SB, Weiss AD (1999) GLM versus CCA spatial modeling of plant species distribution. Plant Ecol 143:107–122
Hickey JR, Nackoney J, Nibbelink NP, Blake S, Bonyenge A, Coxe S et al (2013) Human proximity and habitat fragmentation are key drivers of the rangewide bonobo distribution. Biodivers Conserv 22:3085–3104
Hockings KJ, McLennan MR, Carvalho S, Ancrenaz M, Bobe R, Byrne RW et al (2015) Apes in the Anthropocene: flexibility and survival. Trends Ecol Evol 30:215–222
Humle T (2011) Location and Ecology. In: Matsuzawa T, Humle T, Sugiyama Y (eds) The Chimpanzees of Bossou and Nimba. Springer, New York, pp 3–21
Humle T, Matsuzawa T (2001) Behavioural diversity among the wild chimpanzee populations of Bossou and neighbouring areas, Guinea and Cote d’Ivoire, West Africa—a preliminary report. Folia Primatol 72:57–68
Humle T, Matsuzawa T (2004) Oil palm use by adjacent communities of chimpanzees at Bossou and Nimba Mountains, West Africa. Int J Primatol 25:551–581
Humle T, Boesch C, Campbell G, Junker J, Koops K, Kuehl H, Sop T (2016a) Pan troglodytes ssp. verus. The IUCN red list of threatened species: e.T15935A102327574. Accessed 3 May 2017
Humle T, Maisels F, Oates JF, Plumptre A, Williamson EA (2016b) Pan troglodytes. The IUCN red list of threatened species. e.T15933A102326672. Accessed 3 May 2017
Iverson LR, Dale ME, Scott CT, Prasad A (1997) A GIS-derived integrated moisture index to predict forest composition and productivity of Ohio forests (U.S.A.). Landsc Ecol 12:331–348
Jantz SM, Pintea L, Nackoney J, Hansen MC (2016) Landsat ETM + and SRTM data provide near real-time monitoring of chimpanzee (Pan troglodytes) habitats in Africa. Remote Sens 8:427
Junker J, Blake S, Boesch C, Campbell G, Toit L, Duvall C et al (2012) Recent decline in suitable environmental conditions for African great apes. Divers Distrib 18:1077–1091
Koops K (2011) Elementary technology of foraging and shelter in the chimpanzees (Pan troglodytes verus) of the Nimba Mountains, Guinea. Dissertation, University of Cambridge
Koops K, Matsuzawa T (2006) Hand clapping by a chimpanzee in the Nimba mountains, Guinea. West Africa. PAN 13:21
Koops K, Humle T, Sterck EHM, Matsuzawa T (2007) Ground-nesting in the chimpanzees of the Nimba Mountains, Guinea: environmental or social determinants? Am J Primatol 69:407–419
Koops K, McGrew WC, de Vries H, Matsuzawa T (2012a) Nest-Building by Chimpanzees (Pan troglodytes verus) at Seringbara, Nimba Mountains: antipredation, thermoregulation, and antivector hypotheses. Int J Primatol 33:356–380
Koops K, McGrew WC, Matsuzawa T, Knapp L (2012b) Terrestrial nest-building in wild chimpanzees (Pan troglodytes verus): implications for the tree-to-ground sleep transition in early hominins. Am J Phys Anthropol 148:351–361
Koops K, McGrew WC, Matsuzawa T (2013) Ecology of culture: do environmental factors influence foraging tool use in wild chimpanzees (Pan troglodytes verus)? Anim Behav 85:175–185
Koops K, Schöning C, McGrew WC, Matsuzawa T (2015) Chimpanzees prey on army ants at Seringbara, Nimba Mountains, Guinea: predation patterns and tool characteristics. Am J Primatol 77:319–329
Kormos R, Boesch C, Bakarr M, Butynski TM (2003) Status survey and conservation action plan: West African chimpanzees. IUCN, Gland, Switzerland
Kühl HS, Sop T, Williamson EA, Mundry R, Brugière D, Campbell G et al (2017) The critically endangered western chimpanzee declines by 80%. Am J Primatol. https://doi.org/10.1002/ajp.22681
Kumar S, Neven LG, Yee WL (2014) Evaluating correlative and mechanistic niche models for assessing the risk of pest establishment. Ecosphere 5:86
Lamotte M (1998) Le Mont Nimba: réserve de biosphère et site du patrimoine mondial (Guinée et Côte d’Ivoire); initiation à la géomorphologie et à la biogéographie. UNESCO, Paris
Laurance WF, Carolina Useche D, Rendeiro J, Kalka M, Bradshaw CJA, Sloan SP et al (2012) Averting biodiversity collapse in tropical forest protected areas. Nature 489:290–294
Liu C, Berry PM, Dawson TP, Pearson RG (2005) Selecting thresholds of occurrence in the prediction of species distributions. Ecography 28:385–393
Matsuzawa T, Humle T (2011) Bossou: 33 Years. In: Matsuzawa T, Humle T, Sugiyama Y (eds) The Chimpanzees of Bossou and Nimba. Springer, New York, pp 361–370
Matsuzawa T, Yamakoshi G (1996) Comparison of chimpanzee material culture between Bossou and Nimba, West Africa. In: Russon AE, Bard KA, Parker S (eds) Reaching into thought: the minds of the great apes. Cambridge University Press, Cambridge, pp 211–232
Matsuzawa T, Humle T, Sugiyama Y (2011a) The Chimpanzees of Bossou and Nimba. Springer, New York
Matsuzawa T, Ohashi G, Humle T, Granier N, Kourouma M, Soumah AG (2011b) Green. Corridor Project: Planting Trees in the Savanna Between Bossou and Nimba. In: Matsuzawa T, Humle T, Sugiyama Y (eds) The Chimpanzees of Bossou and Nimba. Springer, New York, pp 361–370
McCune B, Keon D (2002) Equations for potential annual direct incident radiation and heat load. J Veg Sci 13:603–606
McLennan MR, Hill CM (2012) Troublesome neighbours: changing attitudes towards chimpanzees (Pan troglodytes) in a human-dominated landscape in Uganda. J Nat Conserv 20:219–227
McNab WH (1989) Terrain shape index: quantifying effect of minor landforms on tree height. For Sci 35:91–104
McNab WH (1993) A topographic index to quantify the effect of mesoscale landform on site productivity. Can J For Res 23:1100–1107
Morin PA, Moore JJ, Chakraborty R, Jin L, Goodall J, Woodruff DS (1994) Kin selection, social structure, gene flow, and the evolution of chimpanzees. Science 265:1193–1201
Murphy MA, Evans JS, Storfer A (2010) Quantifying Bufo boreas connectivity in Yellowstone National Park with landscape genetics. Ecology 91:252–261
NASA JPL (2009) ASTER global digital elevation model V002 [data set]. NASA JPL. https://doi.org/10.5067/ASTER/ASTGTM.002
Normand E, Boesch C (2009) Sophisticated Euclidean maps in forest chimpanzees. Anim Behav 77:1195–1201
Normand E, Ban SD, Boesch C (2009) Forest chimpanzees (Pan troglodytes verus) remember the location of numerous fruit trees. Anim Cogn 12:797–807
Norris D (2014) Model thresholds are more important than presence location type: understanding the distribution of lowland tapir (Tapirus terrestris) in a continuous Atlantic forest of southeast Brazil. Trop Conserv Sci 7:529–547
Pearce J, Ferrier S (2000) Evaluating the predictive performance of habitat models developed using logistic regression. Ecol Modell 133:225–245
Pearson RG, Raxworthy CJ, Nakamura M, Townsend Peterson A (2007) Predicting species distributions from small numbers of occurrence records: a test case using cryptic geckos in Madagascar. J Biogeogr 34:102–117
Peterson AT, Papeş M, Eaton M (2007) Transferability and model evaluation in ecological niche modeling: a comparison of GARP and Maxent. Ecography 30:550–560
Phillips SJ, Dudík M (2008) Modeling of species distribution with Maxent: new extensions and a comprehensive evaluation. Ecography 31:161–175
Phillips SJ, Avenue P, Park F (2004) A maximum entropy approach to species distribution modeling. Proceedings of the Twenty-First International Conference on Machine Learning. ACM, New York, pp 655–662
Phillips SJ, Anderson RP, Schapire RE (2006) Maximum entropy modeling of species geographic distributions. Ecol Modell 190:231–259
Phillips SJ, Dudik M, Elith J, Graham CH, Lehmann A, Leathwick J, Ferrier S (2009) Sample selection bias and presence-only distribution models: implications for background and pseudo-absence data. Ecol Appl 19:181–197
Pike RJ, Wilson SE (1971) Elevation-relief ratio, hypsometric integral, and geomorphic area-altitude analysis. Bull. Geological Society of America 82:1079–1084
Pintea L, Bauer ME, Bolstad PV, Pusey A (2003) Matching multiscale remote sensing data to interdisciplinary conservation needs: the case of chimpanzees in Western Tanzania. Pecora 15/l. Satell. Inf. IV/ISPRS Comm. I/FIEOS 2002 Conference Proceedings 12. http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.222.2431&rep=rep1&type=pdf
Plumptre AJ, Rose R, Nangendo G, Williamson EA, Didier K, Hart J et al (2010) Eastern Chimpanzee (Pan troglodytes schweinfurthii): Status Survey and Conservation Action Plan 2010-2020. IUCN, Gland, Switzerland
Randin CF, Dirnböck T, Dullinger S, Zimmermann NE, Zappa M, Guisan A (2006) Are niche-based species distribution models transferable in space? J Biogeogr 33:1689–1703
Riley SJ, DeGloria SD, Elliot R (1999) A terrain ruggedness index that quantifies topographic heterogeneity. Intermt J Sci 5:23–27
Rödder D, Lawing AM, Flecks M, Ahmadzadeh F, Dambach J et al (2013) Evaluating the significance of paleophylogeographic species distribution models in reconstructing quaternary range-shifts of nearctic chelonians. PLoS ONE 8(10):1–19
Rushton S, Ormerod S, Kerby G (2004) New paradigms for modelling species distributions? J Appl Ecol 41:193–200
Serckx A, Huynen MC, Beudels-Jamar RC, Vimond M, Bogaert J, Kühl HS (2016) Bonobo nest site selection and the importance of predictor scales in primate ecology. Am J Primatol 78:1326–1343
Songer M, Delion M, Biggs A, Huang Q (2012) Modeling impacts of climate change on giant panda habitat. Int J Ecol 2012:1–12
Swets KA (1988) Measuring the accuracy of diagnostic systems. Science 240:1285–1293
R Core Team (2005) R: a language and environment for statistical computing. R foundation for statistical computing, Vienna, Austria. https://www.R-project.org/. Accessed 15 Jan 2017
The Green Corridor Project (2017) http://www.greencorridor.info/en/videos/Green-Corridor/. Accessed 3 May 2017
Torres J, Brito JC, Vasconcelos MJ, Catarino L, Gonçalves J, Honrado J (2010) Ensemble models of habitat suitability relate chimpanzee (Pan troglodytes) conservation to forest and landscape dynamics in Western Africa. Biol Conserv 143:416–425
Warren DL, Seifert SN (2011) Ecological niche modeling in Maxent: the importance of model complexity and the performance of model selection criteria. Ecol Appl 21:335–342
Wich SA, Gaveau D, Abram N, Ancrenaz M, Baccini A, Brend S et al (2012) Understanding the impacts of land-use policies on a threatened species: is there a future for the Bornean Orang-utan? PLoS One 7:e49142
Wilson JW, Sexton JO, Todd Jobe R, Haddad NM (2013) The relative contribution of terrain, land cover, and vegetation structure indices to species distribution models. Biol Conserv 164:170–176
World Heritage Committee (2017) Mount Nimba Strict Nature Reserve. http://whc.unesco.org/en/list/155/. Accessed 1 Sep 2017
Zurell D, Elith J, Schröder B (2012) Predicting to new environments: tools for visualizing model behaviour and impacts on mapped distributions. Divers Distrib 18:628–634
Acknowledgements
We thank: Direction Nationale de la Recherche Scientifique (DNRST) and Institut de Recherche Environnementale de Bossou (IREB) in Guinea for research authorization; Seringbara guides, K. Doré, F. Doré, F. Zogbila, N. Doré, D. Zogbila, Y. Zogbila, C. Samy, and N. Gbouomy; research assistants, P. Le Sommer, W. Edwards, J. Caraway, D. Montanari, N. James, I. Vélez del Burgo Guinea, D. Hassler, M. McCann, S. Canington, and G. Mamy for help in the field. Research was supported by grants from Gates Cambridge Trust, Lucie Burgers Foundation for Comparative Behaviour Research (the Netherlands), Homerton College and Newnham College (Cambridge) to K.K., and by the Japan Society for the Promotion of Science (JSPS) Leading Graduate Program‐U04‐ PWS, JSPS core-to-core CCSN, and the Ministry of Education, Culture, Sports, Science and Technology of Japan (MEXT)/JSPS-KAKENHI (#07102010, #12002009, #16002001, #20002001, #24000001, #16H06283) grants to T.M.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Electronic supplementary material
Below is the link to the electronic supplementary material.
About this article

Cite this article
Fitzgerald, M., Coulson, R., Lawing, A.M. et al. Modeling habitat suitability for chimpanzees (Pan troglodytes verus) in the Greater Nimba Landscape, Guinea, West Africa. Primates 59, 361–375 (2018). https://doi.org/10.1007/s10329-018-0657-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10329-018-0657-8