
Abstract
Reliable prediction and diagnosis of concussion is important for its effective clinical management. Previous model-based studies largely employ peak responses from a single element in a pre-selected anatomical region of interest (ROI) and utilize a single training dataset for injury prediction. A more systematic and rigorous approach is necessary to scrutinize the entire white matter (WM) ROIs as well as ROI-constrained neural tracts. To this end, we evaluated injury prediction performances of the 50 deep WM regions using predictor variables based on strains obtained from simulating the 58 reconstructed American National Football League head impacts. To objectively evaluate performance, repeated random subsampling was employed to split the impacts into independent training and testing datasets (39 and 19 cases, respectively, with 100 trials). Univariate logistic regressions were conducted based on training datasets to compute the area under the receiver operating characteristic curve (AUC), while accuracy, sensitivity, and specificity were reported based on testing datasets. Two tract-wise injury susceptibilities were identified as the best overall via pair-wise permutation test. They had comparable AUC, accuracy, and sensitivity, with the highest values occurring in superior longitudinal fasciculus (SLF; 0.867–0.879, 84.4–85.2, and 84.1–84.6%, respectively). Using metrics based on WM fiber strain, the most vulnerable ROIs included genu of corpus callosum, cerebral peduncle, and uncinate fasciculus, while genu and main body of corpus callosum, and SLF were among the most vulnerable tracts. Even for one un-concussed athlete, injury susceptibility of the cingulum (hippocampus) right was elevated. These findings highlight the unique injury discriminatory potentials of computational models and may provide important insight into how best to incorporate WM structural anisotropy for investigation of brain injury.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Traumatic brain injury (TBI) is a leading cause of morbidity and mortality in the USA (CDC 2015). Of the estimated 1.7–3.8 million individuals suffering from TBI each year in the USA alone, 75% are classified as mild traumatic brain injury (mTBI) or concussion (CDC 2015). The public awareness of concussion and its adverse cognitive and neurodegenerative consequences is growing (NRC 2014). A reliable prediction and diagnosis of TBI, including concussion, is important for effective management of this prevailing neurological disorder.
Despite decades of active research, the biomechanical mechanisms behind TBI remain elusive. Historically, efforts have been focused on characterizing head impact kinematics, using linear and/or rotational acceleration peak magnitudes and their variants. However, as head rotation is considered the main mechanism for mTBI and that angular velocity, as opposed to acceleration, is more predictive of strains (Zhao and Ji 2016), recent metrics have explicitly incorporated peak angular velocity [e.g., Rotational Injury Criterion (RIC) and Power Rotational Head Injury Criterion (PRHIC) (Kimpara and Iwamoto 2012), and Brain Injury Criterion (BrIC) (Takhounts et al. 2013)].
Unfortunately, these empirically derived kinematic metrics do not directly inform brain mechanical responses that are thought to initiate injury (King et al. 2003). In part, this may explain why no consensus has been reached on the most appropriate metric for injury prediction. To estimate impact-induced responses, finite element (FE) models of the human head are important tools (Yang et al. 2011). Model-estimated brain responses have been shown to be more effective in predicting injury than kinematic metrics alone (Zhang et al. 2004; Marjoux et al. 2008; Takhounts et al. 2008; Giordano and Kleiven 2014a; Hernandez et al. 2014). In addition, FE models also enable correlating tissue deformation with specific concussion symptomatic measures (Viano et al. 2005), which is not feasible for kinematic metrics.
Improving the models’ injury predictive power is an ongoing, constant process. Sophisticated head models continue to emerge with more anatomical details (Mao et al. 2013), representing subject-specific anatomies (Ji et al. 2015), and characterizing anisotropic material properties of the white matter (WM) (Sahoo et al. 2014; Giordano and Kleiven 2014b). Lately, there are also efforts to integrate information from neuroimages (Fahlstedt et al. 2015; Miller et al. 2016), e.g., WM structural anisotropy (Wright and Ramesh 2012; Garimella and Kraft 2016), into biomechanical modeling for injury analysis. This aligns well with in vitro studies that suggest strain component along axonal longitudinal direction responsible for axonal injury (Cullen and LaPlaca 2006). Initial evidence indicates that WM fiber orientation-dependent strain (termed “fiber strain”, “axonal strain”, or “tract-oriented strain”) improves injury prediction performance relative to its isotropic counterpart, maximum principal strain (Chatelin et al. 2011; Wright et al. 2013; Giordano and Kleiven 2014a; Sullivan et al. 2014; Ji et al. 2015).
Regardless of the injury predictor variables employed, previous model-based studies typically utilize responses from a single pre-selected anatomical region of interest (ROI) to assess injury risk. The commonly used CSDM (cumulative strain damage measure) relies on maximum principal strain of the entire brain (Bandak and Eppinger 1994), while a variant is defined on generic ROIs (Weaver et al. 2012). Other generic or more targeted ROIs, including the corpus callosum, midbrain, and brainstem, are also common choices. For example, the maximum shear stress in the brainstem was found to correlate the strongest with the occurrence of mTBI when analyzing 24 NFL head impacts [9 concussions vs. 15 non-injury cases; (Zhang et al. 2004)]. Using an expanded dataset (58 impacts; 25 concussions vs. 33 non-injury cases), Kleiven studied 8 tissue injury predictors in 6 brain regions. He found that strain in the gray matter and \(\hbox {CSDM}_{0.1}\) (using a strain threshold of 0.1) in the WM had the highest accuracy in concussion classification (Kleiven 2007). After incorporating WM material property anisotropy, however, their model indicated that instead, peak axonal strain within the brainstem had the highest predictive power (Giordano and Kleiven 2014a).
These conflicting reports on the “best” predictor variable and brain ROI to achieve the most accurate injury prediction highlight current challenges in studying the biomechanical mechanisms of TBI. Conceptually, model-based brain injury prediction is analogous to an ill-posed optimization problem—using model-estimated responses to “fit” the given binary injury data. First, errors in the “input variables”, head impact kinematics, seem unavoidable. Laboratory-reconstructed NFL head impacts had a reported error of up to 11% for impact velocity and a maximum error of 25% for resultant angular acceleration (Newman et al. 2005). However, error magnitude in angular velocity was not available, even though it is considered as the primary injury mechanism (Takhounts et al. 2013) and more predictive of strains (Kleiven 2006; Zhao and Ji 2015, 2016). For on-field head impacts, temporally validated rotational acceleration/velocity profiles appear yet to be developed (Beckwith et al. 2012; Allison et al. 2014).
Second, the “optimizer”, FE models of the human head, could vary substantially due to uncertainties in model assumptions [material properties of the brain in particular (Chatelin et al. 2010)]. The lack of high-quality experimental data especially in live humans under injury-causing impacts (Hardy et al. 2001, 2007; Sabet et al. 2008) also precludes sufficient, reliable model validations (Yang et al. 2011). Consequently, even “validated” head models could produce substantially discordant brain responses under identical head impacts (Ji et al. 2014a).
Third, and equally importantly, the “objective function” is also under-defined. Well-documented and accepted brain injury cases including both impact kinematics and clinical injury diagnoses are lacking and are subject to errors. Thus, a single “training dataset” has been used in previous studies to evaluate injury prediction performance. A separate “testing dataset” is not widely available to enable an independent performance verification, even though this is considered necessary and important (Anderson et al. 2007; Sullivan et al. 2014). Taken together, these uncertainties and lack of high-quality, well-accepted injury data could yield multiple “local minima” in the optimization, which may explain previous conflicting observations based on the same injury dataset. However, they may not necessarily correspond to the true “global minimum”.
Given these challenges, potentially there could be data “over-fitting” concerns when attempting to pinpoint a specific injury predictor and ROI for the best performance, especially when using responses from a single element [e.g., peak maximum strain regardless of the location in a given ROI; (Zhang et al. 2004; Giordano and Kleiven 2014a)]. Therefore, there is a need for more systematic and objective evaluation and comparison of the injury predictors’ performances. Here, we employed a repeated random subsampling technique to train and optimize injury predictors while reporting their performances using independent testing datasets. This is a popular cross-validation technique (Arlot and Celisse 2010), but does not appear to have been applied in model-based TBI studies to date. Further, we analyzed injury prediction performances and vulnerabilities of the entire deep WM ROIs as well as ROI-constrained neural tracts from whole-brain tractography (Zhao et al. 2016). Instead of relying on peak responses from a single element from a predefined ROI, we used data sampling across all of the deep WM ROIs/neural tracts. Similarly, regional vulnerabilities were also evaluated via data sampling of predictor responses across all of the simulated head impacts.
These injury analyses significantly extended previous model-based TBI studies to formulate a more systematic and rigorous approach for evaluation of injury prediction performance. A generic head model was also established with the directly associated neuroimages and whole-brain tractography, which was a critical stepping stone toward better integration of neuroimaging and TBI biomechanics studies in the future. Therefore, findings from this work may provide important insight into how best to predict injury for improved mitigation and clinical management.
2 Methods
2.1 Image registration and geometrical transformation
Essential to this study was to integrate structural neuroimaging into biomechanical modeling. This required transforming all image volumes, their corresponding geometrical entities, and the head FE model into a common coordinate system. For convenience, we have chosen the coordinate system of the Worcester Head Injury Model [WHIM; formerly known as the Dartmouth Head Injury Model or DHIM; (Ji et al. 2015; Zhao et al. 2016)] as a common reference. The WHIM was created based on high-resolution T1-weighted MRI (at an isotropic resolution of \(\hbox {1 mm}^{3}\)) of an individual. It had a resolution of \(3.3\pm 0.79\hbox { mm}\) for the brain. Diffusion tensor imaging (DTI) of the same individual provided WM fiber orientations at discrete voxels (at an isotropic resolution of \(\hbox {2 mm}^{3}\)) and real-valued fiber sampling points of the whole-brain tractography (at a resolution of 1 mm). In comparison, the standard ICBM-DTI-81 WM atlas (Mori et al. 2008) averaged from a group of 81 healthy adults served as the WM ROI anatomical constraints. This atlas is provided within another standard anatomical template (ICBM-152). Figure 1 schematically illustrates how these image volumes were registered to transform their corresponding geometrical entities into the WHIM common coordinate system. A rigid registration was performed between the T1-weighted MRI and DTI anatomical image \((b=0)\) of the same individual. In contrast, a non-linear registration using the FNIRT tool in FSL was applied to transform the ICBM-DTI-81 atlas into the WHIM space (Andersson et al. 2007).

Schematic illustration of image volume registrations to transform all geometrical entities into the common reference coordinate system of the Worcester Head Injury Model (WHIM)
2.2 The Worcester Head Injury Model
Details of the Worcester Head Injury Model (WHIM; Fig. 2), including model creation, mesh quality, assignment of material properties and boundary conditions, and validation performances have been reported extensively in recent publications (Ji et al. 2015; Ji and Zhao 2015; Zhao et al. 2016; Zhao and Ji 2016). Therefore, they are not repeated here. Importantly, the WHIM has achieved an overall “good” to “excellent” validation [as assessed by correlation score based on Normalized Integral Square Error (Donnelly et al. 1983; Kimpara et al. 2006)] at the low (\(\sim \)250–300\(\hbox { rad}/\hbox {s}^{2}\) for a live human volunteer), mid (\(\sim \)1.9–2.3\(\hbox { krad}/\hbox {s}^{2}\) for cadaveric impact tests C755-T2 and C383-T1), and high (\(\sim \)11.9\(\hbox { krad}/\hbox {s}^{2}\) for cadaveric test C393-T4) levels of head angular acceleration magnitudes provided important confidence of the fidelity in WHIM-estimated brain responses.

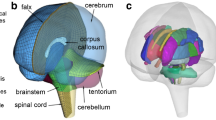
The exterior features (a) and intracranial components (b) of the WHIM (formerly known as the Dartmouth Head Injury Model or DHIM), along with eight representative WM ROIs (c) and four corresponding neural tracts from the whole-brain tractography (showing a 10% random subset; d)
2.3 Simulation of the NFL reconstructed head impacts
We used the 58 reconstructed NFL head impacts as model inputs (Newman et al. 2000), which included 25 concussions and 33 non-injury cases. Details of video recording analysis (Pellman et al. 2003) and the procedures of head impact reconstruction (Newman et al. 2000, 2005) were previously reported. Briefly, all head impact accelerations were collected at 10 kHz following the SAE J211 protocol. The acceleration profiles were pre-processed according to the CFC 1000 requirements (Newman et al. 2000). Identical to previous studies (Newman et al. 2000; Kleiven 2007), all acceleration profiles were filtered using the CFC 180 low-pass filter. The resulting time histories of the linear and angular accelerations were prescribed to the WHIM head center of gravity (CG) to induce brain mechanical responses.
2.4 Voxel- and tract-wise WM fiber strains
Whole-brain tractography was generated using the DTI of the same individual selected to develop the baseline WHIM, as previously reported (Zhao et al. 2016). This led to \({\sim }35\hbox { k}\) fibers and \({\sim }3.3\) million sampling points in total. Using the transformed ICBM-DTI-81 atlas (Fig. 1) as anatomical constraints, the 50 deep WM ROIs (see Table 4 in “Appendix 1”) and their corresponding neural tracts were identified within the WHIM [illustrated in Fig. 2c, d; (Zhao et al. 2016)].
For each impact simulated, element-wise maximum principal strain and strain tensor were extracted at every time step during the entire impact simulation (temporal resolution of 1 ms). Fiber strain at each WM voxel or fiber sampling point was calculated, at every time step, using the corresponding fiber orientation and strain tensor of the nearest element (Ji et al. 2015). For all strains, their peak values across the entire impact simulation, regardless of the time of occurrence, were used. They were denoted as \(\varepsilon _p \) and \(\varepsilon _n \) for maximum principal strain and fiber strain, respectively. Due to the large number of fibers for each WM neural tract, a 10% random subset was utilized for improved computational efficiency. This led to an average of \(183\pm 42\) (range of 88–307) fibers for a given WM neural tract. The down-sampling did not significantly alter strain or injury susceptibility measures (Zhao et al. 2016).
2.5 Injury susceptibility measures in the deep WM neural tracts
A tract-wise injury susceptibility index \((\varphi _{\mathrm{tract}} )\) was established for a given neural tract as the fraction of “injured” WM fibers (i.e., \(\varepsilon _n \) greater than a threshold, determined in Sect. 2.8, regardless of the occurrence location along the given fiber):
However, a potential weakness of this, perhaps over-simplified, definition was that it did not differentiate the relative likelihood of injury to a given fiber according to the number of sampling points exposed to high strains (e.g., a fiber with either one or 100% of the sampling points “injured” would be treated equally). To characterize this “confidence” in injury likelihood, a weighting factor, w, was devised:
Applying w to the binary injury status (0 and 1 for “uninjured” and “injured”, respectively) of each fiber led to the following enhanced tract-wise injury susceptibility index:
Further, a sampling point-based susceptibility index was also established to describe the fraction of “injured” fiber sampling points:
Essentially, \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) and \(\varphi _{\mathrm{tract}}^\mathrm{point}\) extended the concept of CSDM, which was originally developed for the entire brain, to individual WM neutral tracts (Zhao et al. 2016). The former further accounted for the distribution of “injured” sampling points among fibers.
2.6 Injury predictor variables
While tissue strain is considered as a primary variable to assess injury risk and severity, no consensus exists how best to describe the tissue strain status. Common choices include the maximum strain magnitude from a single element in a particular region, regardless of the time of occurrence or location [i.e., peak strain (Zhang et al. 2004; Giordano and Kleiven 2014a)], or a dichotomous variant describing the percentage of tissue volume experiencing large strains [e.g., CSDM (Takhounts et al. 2008) or Pop90 (Sullivan et al. 2014)]. In this study, we evaluated the performances of peak strain, regional average strain, and the dichotomous variants based on either \(\varepsilon _p \) or \(\varepsilon _n \). Specifically, a total of nine strain-based injury predictor variables were evaluated, as summarized in Table 1. All strains were evaluated on MR voxel locations by interpolating from neighboring FE elements, except for tract-wise injury susceptibilities for which \(\varepsilon _n \) was evaluated at higher-resolution fiber sampling points. For clarity, here we referred to an injury predictor as the predictor variable obtained from a given ROI or neural tract.
2.7 Repeated random subsampling and logistic regression
The 58 NFL head impact cases were randomly split into training (39 cases, or approximately two-thirds) and testing (the remaining 19 cases) datasets. This process was repeated 100 times (considered sufficiently large) (Arlot and Celisse 2010). For each training dataset in a random subsampling trial, a standard univariate logistic regression was performed against the binary injury statuses (0 and 1 for “uninjured” and “injured”, respectively) for each injury predictor. A receiver operating characteristic (ROC) curve was generated to export the area under the curve (AUC). The logistic regression model was then applied to the corresponding non-overlapping testing dataset for injury prediction. The resulting numbers of true positives (TP), true negatives (TN), false positives (FP), and false negatives (FN) were used to calculate performance measures such as the accuracy, sensitivity, and specificity according to the following equations:
For each random trial, this produced four 9-by-50 matrices encoding the AUC, accuracy, sensitivity, and specificity, respectively (9 predictor variables and 50 WM ROIs/neural tracts). Combining all of the 100 random trials led to four corresponding 9-by-50-by-100 matrices, which were used for subsequent evaluation.
2.8 Optimal \(\varepsilon _\mathrm{thresh}\) for injury susceptibility indices
The dichotomous injury susceptibility indices (Table 1) depended on a strain threshold. A definitive \(\varepsilon _\mathrm{thresh}\) for the human brain has not been established. An in vivo study suggests a wide range [0.09–0.47 to induce impairment (Bain and Meaney 2000)]. Thresholds from other model-based studies also varied depending on the region (e.g., 0.21 in the corpus callosum (Kleiven 2007); 0.26 (Kleiven 2007) or 0.19 (Zhang et al. 2004) in the gray matter) or type of strain [e.g., 0.21 in maximum principal strain (Kleiven 2007) or 0.07 in axonal strain in the corpus callosum (Giordano and Kleiven 2014a)]. As even “validated” head injury models can produce substantially discordant strains under identical head impact conditions (Ji et al. 2014a), these thresholds were not directly applicable here.
Instead, the following method was adopted within the repeated random subsampling framework, which was an extension to that applied to determine the threshold for CSDM (Giordano and Kleiven 2014a). Specifically, for each training dataset and a given injury susceptibility index, a range of candidate thresholds (\(\varepsilon _\mathrm{thresh}\); 21 unique values within a range of 0.05–0.25 at a step size of 0.01, based on the previous studies) were enumerated to define the susceptibility index for a given WM ROI/neural tract. A logistic regression analysis was then conducted, from which a Wald \(\chi ^{2}\) test was performed. A significant relationship was said to exist between the injury risk and the predictor when the p value was less than 0.05, with a lower value indicating a more significant relationship. Next, for each WM ROI/neural tract, tied rank values were assigned to score \(\varepsilon _\mathrm{thresh}\) in the order of their corresponding \(\chi ^{2}\) test p values (Wilcoxon 1946), where a lower value led to a smaller rank value. At each \(\varepsilon _\mathrm{thresh} \) value, the average rank value across all WM ROIs/neural tracts was used to represent its overall performance. The optimal \(\varepsilon _\mathrm{thresh}\) corresponding to the smallest rank value was then identified (Table 2). Essentially, this was to minimize the overall p value, or to maximize the significance of risk-response relationship, for the group of ROIs/tracts. This process is illustrated in Fig. 3 for a typical training dataset. Finally, an average optimal \(\varepsilon _\mathrm{thresh}\) among the trials was obtained for each injury susceptibility index, which was subsequently used in a separate round of repeated random subsampling to evaluate injury prediction performances. The three \(\varepsilon _n\)-based metrics, \(\varphi _{\mathrm{ROI}}^{\varepsilon _{n}}\), \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \), and \(\varphi _{\mathrm{tract}}^\mathrm{point}\), had an optimal \(\varepsilon _\mathrm{thresh}\) of 0.09–0.10, which was consistent with the lower bound of a conservative injury threshold of 0.09 established from an in vivo optical nerve stretching experiment (Bain and Meaney 2000).

a Illustration of the ranking approach to identify the optimal \(\varepsilon _\mathrm{thresh}\) based on the training dataset in a typical round of random subsampling trial. For each WM ROI/neural tract, a tied rank was assigned for each \(\varepsilon _\mathrm{thresh}\) according to the Wald \(\chi ^{2}\) test p values. For each \(\varepsilon _\mathrm{thresh} \), an average rank value was obtained, as shown. b Average rank values as a function of \(\varepsilon _\mathrm{thresh} \). For each injury susceptibility measure, the one that yielded the smallest (i.e., the best) average rank value was chosen as the optimal \(\varepsilon _\mathrm{thresh} \) (Table 2)
2.9 Relative injury prediction performance among predictor variables
For each predictor variable, its performance measures in each WM ROI/neural tract were averaged across the 100 subsampling trials. This led to a 9-by-50 matrix of average values for each performance measure. To assess the relative performances among these predictor variables, we performed a pair-wise one-sided permutation test (see “Appendix 2”) based on the average AUC, accuracy, sensitivity, and specificity (\(9\times 8=72\) pairs for each performance metric) sampled across all of the 50 deep WM ROIs/neural tracts. The predictor variables were then ranked by the number of times that a given candidate was found to have a significantly larger average value than others (as determined by the permutation test p values).

Summary of AUCs averaged from 100 random subsampling trials based on the training datasets, shown as a \(9\times 50\) intensity-encoded image. Each row corresponds to an injury predictor variable while each column represents a WM ROI/neural tract. The average AUC value for each injury predictor variable, regardless of the region, is also shown, along with the average and maximum standard deviations of the AUC samples across the 100 trials. Five of the nine injury predictor variables achieved the largest AUCs (averaged across trials) in the SLF-R (arrow)

Summary of average accuracy measures based on the testing datasets. Four of the nine injury predictor variables achieved the best accuracy score in the CGC-R and SLF-R (four for each; arrows)

Summary of average sensitivity measures based on the testing datasets. Three of the nine injury predictor variables achieved the best sensitivity score in the SLF-R (arrow)

Summary of average specificity based on the testing datasets. Five of the nine injury predictor variables achieved the best specificity score in the CGC-R (arrow)

Pair-wise performance comparisons between the injury predictor variables in terms of AUC (a), accuracy (b), sensitivity (c), and specificity (d). Each square in a row represents whether the performance measure was significantly larger (dark gray; otherwise, white) than that in a column (self-comparisons along the diagonal excluded)
2.10 Relative vulnerability among deep WM ROIs and neural tracts
Identifying the most vulnerable WM ROIs or neural tracts may be of clinical significance to potentially relate impacts to specific brain functional alteration as well as for potential targeted therapeutic treatment in the future. Here, we defined the relative vulnerabilities among the 50 ROIs/neural tracts as the frequency that each region experienced responses larger than others. Similarly, for each injury predictor variable (Table 1), we employed one-sided permutation test based on paired-sample t-statistics, using the response values of the ROIs/neural tracts sampled across the 58 simulated head impacts. A total of \(50\times 49 = 2450\) pairs of permutation test were conducted for a given injury predictor variable. Finally, their relative vulnerabilities were ranked.

The number of times that a given WM ROI had an average response value (from the 58 simulated head impacts) significantly larger than others using injury predictors based on \(\varepsilon _p\): peak \(\varepsilon _p\) (a), average \(\varepsilon _p \) (b), and \(\varphi _{\mathrm{ROI}}^{\varepsilon _p} \) (c). Their respective top five most vulnerable ROIs are highlighted (gray)

The number of times that a given WM ROI had an average response value significantly larger than others using injury predictors based on \(\varepsilon _n \): peak \(\varepsilon _n\) (a), average \(\varepsilon _n\) (b), and \(\varphi _{\mathrm{ROI}}^{\varepsilon _n}\) (c). Their respective top five most vulnerable ROIs are highlighted
3 Data analysis
All head impacts were simulated using Abaqus/Explicit (Version 6.12; Dassault Systèmes, France). For each impact, element-wise peak strains during the entire simulated event were obtained. For injury predictor variables that required a pre-determined threshold, an optimal threshold value was determined. Repeated random subsampling was utilized to assess injury performances. For each performance measure, average values across all of the WM ROIs/neural tracts were used to conduct one-sided permutation tests based on paired-sample t-statistics. For each injury predictor variable, the relative vulnerabilities of the WM ROIs/neural tracts were ranked to identify the top five most vulnerable ones. Injury thresholds at 50% injury probability were computed. Finally, representative distributions of strain and injury susceptibility responses for a pair of striking and struck athletes (non-concussed and concussed, respectively) were also illustrated.
For each head impact (100 ms in duration), the computational cost was \({\sim }120\) min for impact simulation with 8 CPUs and \({\sim }60\) min for response extraction (parallel processing on 12 CPUs). All data analyses were performed with in-house MATLAB programs (R2016a; MathWorks, Natick, MA) on a 12-core Linux machine (Intel Xeon X5560, 2.80 GHz, 126 GB memory).
4 Results
4.1 Injury prediction performances
Figures 4, 5, 6, and 7 summarize the average AUC, accuracy, sensitivity, and specificity for the 9 injury predictor variables across the 50 ROIs/neural tracts. For each predictor variable, the performance consistency among the 100 subsampling trials was assessed in terms of standard deviation (either further averaged across the 50 regions or using the maximum value to represent the “extreme” case). In general, AUC based on training datasets had the highest consistency, which was not surprising as this variable was a more stable measure. Other performance measures based on independent testing datasets were also largely consistent among the subsampling trials, perhaps, with an exception of the original \(\varphi _{\mathrm{tract}} \) in terms of sensitivity and specificity (maximum standard deviation reached 37.3 and 33.6%, respectively; Figs. 6 and 7).
The two tract-wise injury susceptibility indices, \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) and \(\varphi _{\mathrm{tract}}^\mathrm{point} \), consistently achieved the highest AUC, accuracy, and sensitivity (averaged from the 50 regions; Figs. 4, 5, 6, 7). However, without weighting the “confidence” of injury for each fiber within a neural tract, the original \(\varphi _{\mathrm{tract}} \) had the worst average AUC (tied with peak \(\varepsilon _n\); Fig. 4). It was intriguing to observe that 5 of the 9 injury predictor variables achieved the best AUC in SLF-R (superior longitudinal fasciculus right) among the 50 deep WM regions. Similarly, 4 (3) of the 9 metrics had the best accuracy (sensitivity) in the same region. Overall, the same three metrics, \(\varphi _{\mathrm{ROI}}^{\varepsilon _n}\), \(\varphi _{\mathrm{tract}}^\mathrm{enhanced}\) and \(\varphi _{\mathrm{tract}}^\mathrm{point}\), performed the best in AUC, accuracy and sensitivity in SLF-R (Figs. 4, 5, 6). In terms of specificity, however, 5 of the 9 metrics, including \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) and \(\varphi _{\mathrm{tract}}^\mathrm{point} \), performed the best in CGC-R (cingulate gyrus right; Fig. 7). Pair-wise permutation tests based on performances sampled across the 50 regions confirmed that the two tract-wise injury susceptibility indices, \(\varphi _{\mathrm{tract}}^\mathrm{enhanced}\) and \(\varphi _{\mathrm{tract}}^\mathrm{point} \), were the best overall, despite the statistically significant but subtle difference in sensitivity (between themselves; 72.5 vs. 71.7%) and specificity (slightly lower than that for average \(\varepsilon _n\); Fig. 8). For each best performer (i.e., region of the highest values in each row in Figs. 4, 5, 6, 7), its metric value was significantly higher than most of the other remaining regions (at least 90% or 44 out of the other 49), according to permutation tests.
4.2 Injury vulnerability and threshold
The relative vulnerabilities for ROI-based injury predictor variables using \(\varepsilon _p\) and \(\varepsilon _n \) are reported in Figs. 9 and 10, respectively. Results for tract-based predictor variables are given in Fig. 11. For ROI-based variables, CP-R (cerebral peduncle right) consistently ranked within the top five most vulnerable regions (highlighted; Figs. 9, 10). When using \(\varepsilon _n \) for injury prediction, GCC (genu of corpus callosum), besides CP, was also identified within the top five (Fig. 10). When using the tract-wise injury predictors (Fig. 11), however, BCC (body of corpus callosum) was consistently found to experience high vulnerability. Perhaps most interestingly, for the top five most vulnerable neural tracts identified by the two best performing injury predictors, \(\varphi _{\mathrm{tract}}^\mathrm{enhanced}\) and \(\varphi _{\mathrm{tract}}^\mathrm{point}\), four of them overlapped. They were GCC, BCC, SLF-L (superior longitudinal fasciculus left) and ALIC-R (anterior limb of internal capsule right; Fig. 11b, c). Finally, thresholds at the 50% injury probability are summarized for the predictor variables (Table 3).

The number of times that a given WM neural tract had an average response value significantly larger than others using injury predictors established from tract-wise \(\varepsilon _n \): \(\varphi _{\mathrm{tract}} \) (a), \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) (b), and \(\varphi _{\mathrm{tract}}^\mathrm{point} \) (c). Their respective top five most vulnerable neural tracts are highlighted
4.3 Illustration from selected cases
Distributions of \(\varepsilon _p\), voxel- and tract-wise \(\varepsilon _n\) are shown for a pair of striking and struck athletes (non-concussed and concussed, respectively) involved in the same head collision (Case157). For the striking player, large \(\varepsilon _n \) responses occurred in the midbrain region (Fig. 12c). For the concussed player, higher \(\varepsilon _p \) and \(\varepsilon _n \) mostly occurred in the peripheral subcortical areas and the midbrain (Fig. 12b, d). The injury susceptibilities using the two best predictors are also illustrated (Fig. 13). Perhaps as expected, most of the tracts in the struck/concussed athlete experienced injury susceptibilities greater than the respective tract-wise thresholds. While the opposite was true for the striking/un-concussed athlete, CGH-R (cingulum (hippocampus) right) also experienced elevated \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) that exceeded its injury threshold.
5 Discussion
Numerous biomechanical (Yang et al. 2011) and neuroimaging (Bigler and Maxwell 2012; Shenton et al. 2012) studies exist that attempt to elucidate the mechanisms behind traumatic brain injury (TBI). However, their integration remains rather limited. Our study using whole-brain tractography and a well-established WM atlas to analyze brain injury in contact sports is an important extension to previous efforts (Kraft et al. 2012; Wright et al. 2013; Giordano and Kleiven 2014b; Sullivan et al. 2014; Ji et al. 2015; Zhao et al. 2016). Instead of similarly pinpointing a specific injury predictor variable in a given brain ROI for injury prediction, here we systematically analyzed injury prediction performances and vulnerabilities of the entire deep WM ROIs and neural tracts. Further, a repeated random subsampling technique was employed to objectively evaluate prediction performances in order to avoid or minimize “over-fitting” concerns in previous efforts where a single training dataset was used.
5.1 Injury prediction performance
The two tractography-based injury susceptibility indices, \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) and \(\varphi _{\mathrm{tract}}^\mathrm{point} \), consistently had the best overall predictive power, while peak \(\varepsilon _n \) performed the worst in general (Fig. 8). Without weighting the “confidence” of injury in each WM fiber, however, the performance of \(\varphi _{\mathrm{tract}} \) degraded significantly. The two best performers virtually had identical AUC and accuracy. Their responses as sampled across the 58 impacts were highly correlated (Pearson correlation coefficients close to 1.0 for all of the neural tracts; range of 0.989–0.998, \(p<0.0001\)). This suggests certain inherent concordance between the two. This was not surprising, given that the two predictor variables would become identical if a neural tract were to be composed of a single fiber. However, unlike \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \), \(\varphi _{\mathrm{tract}}^\mathrm{point}\) depends on the total number of “injured” sampling points only and is invariant to their distribution among the fibers. Therefore, some subtle, but statistically significant, differences were found in their sensitivity and specificity (Fig. 8c, d). Further investigation is necessary to discern their similarities/differences as well as implications in assessing injury risk.
The regional average \(\varepsilon _p \) and \(\varepsilon _n \) within a given ROI consistently outperformed their peak counterparts found in a single element, regardless of its location, for all of the performance measures (Fig. 8). The regional averages were also slightly (but significantly) better in AUC than their dichotomized counterparts, \(\varphi _{\mathrm{ROI}}^{\varepsilon _p } \) and \(\varphi _{\mathrm{ROI}}^{\varepsilon _n}\); however, their comparisons in other performance measures were inconclusive. In addition, \(\varphi _{\mathrm{ROI}}^{\varepsilon _p } \) and \(\varphi _{\mathrm{ROI}}^{\varepsilon _n } \) did not differ in performance between themselves, except that the latter had a larger average AUC than the former based on training datasets (Fig. 8a). Regardless, these tract- and ROI-wise findings indicated that on a group-basis, fiber orientation-dependent strain along WM neural tracts may be more effective in injury prediction than the ROI-based counterparts.

Brain strain distributions for a pair of striking/struck athletes involved in the same head collision. a, b Peak \(\varepsilon _p \) resampled on a coronal plane; c, d Peak \(\varepsilon _n\) in a coronal MR image; and e, f peak \(\varepsilon _n\) along a 10% subset of the whole-brain tractography. For the resampled strain map, only regions corresponding to the brain parenchyma are shown—other regions such as falx and tentorium appear as empty space. All responses are peak values during the entire impact, regardless of the time of occurrence [vs. a time-frozen snapshot (Kleiven 2007)]. The peak magnitudes of resultant angular acceleration and velocity were \(3807 \hbox { rad}/\hbox {s}^{2}\) and 25 rad/s for the striking athlete and were \(7083 \hbox { rad}/\hbox {s}^{2}\) and 38 rad/s for the struck player, respectively
The most injury discriminative WM ROIs/neural tracts were also observed. Note that because of head symmetry relative to the mid-sagittal plane (WHIM, WM atlas, and largely for whole-brain tractography as well), the ROI/tract results should be interpreted bilaterally. For example, findings regarding SLF-R would be equally applicable to its contralateral counterpart, SLF-L [e.g., considering mirrored head impacts (Zhao and Ji 2015)].
SLF appeared to be one of the most injury discriminative neural tracts across a number of \(\varepsilon _n \)-based predictor variables that consistently achieved the best AUC, accuracy, and sensitivity. For example, the two best performers, \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) and \(\varphi _{\mathrm{tract}}^\mathrm{point} \) in this region, had an accuracy and sensitivity of 84.4–85.2 and 84.1–84.6%, respectively, and the corresponding specificity score was only slightly less than CGC (85.4–86.3 vs. 86.2–87.5%). Interestingly, the same neural tract, SLF, was also among the most vulnerable ones, both biomechanically (Fig. 11) and in neuroimaging ((Bigler and Maxwell 2012; Gardner et al. 2012); further see below).
5.2 Injury vulnerability
The relative vulnerabilities depended on the injury predictor variables used. For ROI-based predictor variables, CP (cerebral peduncle, near the brainstem and midbrain) was consistently found to be among the most vulnerable ROIs, regardless of whether \(\varepsilon _p \) or \(\varepsilon _n \) was used (Figs. 9 and 10). This agreed well with typical reports of vulnerability in this region (Zhang et al. 2004; Viano et al. 2005). Large \(\varepsilon _p \) and \(\varepsilon _n\) occurred near this area even for the striking/uninjured athlete (Fig. 9a, c; similarly for the other). Regardless, the regional average responses and susceptibility measures yielded more consistent findings than when using peak values. For example, three of the top five most vulnerable ROIs were identical [CP-R, FX/ST-R (fornix and stria terminalis), and IFO-R (inferior fronto-occipital fasciculus)] when using average \(\varepsilon _p \) and \(\varphi _{\mathrm{ROI}}^{\varepsilon _p }\) as predictor variables. In comparison, average \(\varepsilon _n \) and \(\varphi _{\mathrm{ROI}}^{\varepsilon _n}\) identified the same top five most vulnerable ROIs [GCC, CP-L/R and UNC-L/R (uncinated fasciculus)]. These ROI-wise observations agreed well with a previous study utilizing a subset of the NFL impact cases, where fornix, midbrain, and corpus callosum were found to experience the largest strains (Viano et al. 2005).
For the two best injury predictors based on neural tracts, the top four most vulnerable ones were identical (GCC, BCC, ALIC-R and SLF-L; Fig. 11). In general, this agreed well with neuroimaging studies in the context of sports-related concussion. Certain neural tracts, including GCC (genu of the corpus callosum), SLF, UNC, the inferior longitudinal fasciculus, internal capsule, among others, are known to be more susceptible to mTBI (Kraus et al. 2007; Niogi et al. 2008; Bigler and Maxwell 2012). For example, using tract-based spatial statistics (TBSS), significant increase in mean diffusivity (MD) was observed in the SLF for concussed collegiate athletes playing football ((Cubon et al. 2011); significant changes in DTI parameters are considered indications of damages to the underlying WM). An increased fractional anisotropy (FA) was also found in the GCC and BCC in injured student athletes (Zhang et al. 2010a), while a decreased FA and increased apparent diffusion coefficient (ADC) were found in internal capsule based on a cohort of 81 professional male boxers and 12 male controls (Chappell et al. 2006). In general, these results also agreed with other neuroimaging findings using data from traffic accidents (Messé et al. 2011; Xiong et al. 2014), falls, and assaults (Messé et al. 2011).
Even for un-concussed players, \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) in CGH-R (cingulum (hippocampus) right) was found to have exceeded the corresponding injury threshold (Fig. 12). This appeared to agree with the notion that athletes even without a clinically diagnosed concussion could still experience significant changes in neuroimaging (Talavage et al. 2014; Bazarian et al. 2014) or cognitive alteration (McAllister et al. 2012, 2014), presumably a result from local tissue deformation.
Taken together, our biomechanical investigation, especially for results based on \(\varepsilon _n \) (Figs. 10, 11), appeared to reinforce reports from various neuroimaging studies that have identified the corpus callosum, SLF, UNC, and internal capsule as more frequently injured in mTBI patients (Bigler and Maxwell 2012). Undoubtedly, further studies and more independent models (further see below) are necessary to verify the concordance between biomechanical and neuroimaging findings on a same sizable population in the future. Nevertheless, collectively, these findings do seem to provide a multi-faceted insight into the mechanisms of mTBI.

Illustration of injury susceptibilities using \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) (top) and \(\varphi _{\mathrm{tract}}^\mathrm{point}\) (bottom) for the same pair of athletes. Stars/dashed lines represent tract-wise/average injury thresholds at a 50% probability for concussion. Potentially “injured” tracts (i.e., susceptibilities exceeding the corresponding tract-wise thresholds) are highlighted in gray. Even for the un-concussed athlete, \(\varphi _{\mathrm{tract}}^\mathrm{enhanced}\) in CGH-R still exceeded its tract-wise threshold (arrow)
5.3 Comparison with previous findings
The corpus callosum ranked among the most vulnerable regions for \(\varepsilon _n\)-based predictors. However, it did not have the best injury predictive power. For example, AUC of 0.812 and 0.809 for \(\varphi _{\mathrm{tract}}^\mathrm{enhanced} \) and \(\varphi _{\mathrm{tract}}^\mathrm{point}\) in the main body, respectively, lower than that for SLF-R: 0.879 and 0.867, respectively; Fig. 4). Further, peak \(\varepsilon _n \) in the main body of corpus callous achieved relatively poor AUC, accuracy, sensitivity, specificity scores (0.685, 62.3, 43.9, and 79.0%). In contrast, peak \(\varepsilon _n \) in this region was one of the “best” injury predictors for the latest KTH model that reported an AUC of 0.9488 (Giordano and Kleiven 2014a). This was notably higher than those achieved in our study. Unfortunately, injury prediction performances based on independent testing datasets were not available in that study to enable a direct, more objective comparison.
Nevertheless, these findings, once again, highlight differences among models. A more systematic and objective comparison of brain responses across models may be valuable to gain further confidence in model-based injury studies. This is particularly true given the challenges for reliable, high-quality model validations at present (Yang et al. 2011) and the fact that significant differences exist even among “validated” models (Ji et al. 2014a). The approach presented here (random subsampling and evaluation of the entire deep WM ROIs and neural tracts via a standard WM atlas) may provide a common framework for future model comparisons [vs. simply peak responses in generic regions (Ji et al. 2014a)]. With more independent models analyzing an identical injury dataset using the same approach, the observed concordance between biomechanical and neuroimaging findings may be further reinforced.
5.4 Limitations
Limitations of WHIM on the use of isotropic, homogeneous (vs. anisotropic, heterogeneous) material properties of the brain, and resolution mismatch between FE elements and DTI voxels have been discussed (Ji et al. 2015; Zhao et al. 2016). They are not repeated here. Extensive discussions also exist on errors in the impact kinematics (Newman et al. 2005), the resulting uncertainties in model results, and implications in injury prediction such as under-sampled non-injury cases that could have biased injury thresholds (Zhang et al. 2004; Kleiven 2007; Giordano and Kleiven 2014a). These random kinematic errors likely could significantly influence results on an individual basis. This will be the subject of a future “sensitivity” evaluation. However, they are unlikely to significantly alter the group-wise results presented here such as the relative discriminative power among the injury predictor variables (as indicated by the consistency among random trials; Figs. 4, 5, 6, 7) or vulnerabilities among the ROIs/tracts. Importantly, a model-based TBI study is analogous to an ill-posed optimization problem—uncertainties and potential errors will likely persist in virtually all aspects involved. A systems or integrated approach is important to ultimately elucidate the mechanisms behind TBI (Zhao et al. 2016). When additional injury datasets are available, further independent evaluations would reveal whether the findings in this study based on the limited NFL dataset could be generalized to other at-risk populations.
There are several other limitations. First, we have used neuroimaging and whole-brain tractography from one individual to study the NFL athletes on a group-wise basis. Individual variability could not be assessed because their neuroimages were unavailable. Despite this limitation, it must be recognized that our generic head model was a critical stepping stone toward more individualized integration of neuroimaging with TBI biomechanics studies in the future. The model-image mismatch was similar to a recent study that used 2D head models (vs. 3D here) developed from images of a normal individual to study injury of a reconstructed accident in ice-hockey of a different subject (Wright et al. 2013). Another study correlated simulated strain patterns from a generic model with injury findings from individual neuroimages for three reconstructed bicycle accidents, where model and images differed in size and shape (Fahlstedt et al. 2015). An image-atlas-based model represented “averaged” neuroimages but not specific individuals (Miller et al. 2016). Further work is necessary to understand the implications of using generic vs. individualized model and/or neuroimages in injury characterization; however, this is beyond the scope of this study. Nevertheless, importantly, our generic model with the associated neuroimages and whole-brain tractography may provide a valuable tool to enable better integration of neuroimaging and TBI biomechanics studies in the future, especially given that most 50th percentile head models do not yet have detailed neuroimages directly associated with (Zhang et al. 2004; Kleiven 2007; Kimpara and Iwamoto 2012; Takhounts et al. 2013; Giordano and Kleiven 2014a; Sahoo et al. 2016).
Second, a more complete WM atlas also exists [containing as many as 130 ROIs (Zhang et al. 2010b)], which was employed before (Wright et al. 2013). Here, we chose to focus on the deep WM ROIs/neural tracts because certain regions deep in the brain appear more susceptible to mTBI based on neuroimaging findings (Kraus et al. 2007; Bigler and Maxwell 2012). In addition, there is inconsistency in representing the brain-skull boundary conditions in FE head models [e.g., frictional sliding interface (Kleiven 2007) vs. direct nodal sharing via a layer of soft CSF in between the brain and skull (Takhounts et al. 2003; Ji et al. 2015)]. Thus, there could be greater response uncertainty in cortical and subcortical regions. Although using ROIs/tracts only in the deep WM did not necessarily eliminate the concern, it was a reasonable compromise, at least at present, to enable our study along this line of research.
Finally, we have used a univariate logistic regression in each WM region (ROI/neural tract) independently to assess injury risk. As concussion is diffuse in nature, a binary brain injury may well have involved damages to multiple (vs. a single) WM ROIs and/or neural tracts (Fig. 13b, d). It is reasonable to assume that combining responses from all of these regions could improve injury prediction performance. In this case, responses from each WM region (ROI or tract) could serve as unique “features” to enable more sophisticated analysis techniques—multi-variate logistic regression or machine learning [e.g., support vector machine (Hernandez et al. 2014)]—for more effective injury classification. In addition, strain rate (Cullen and LaPlaca 2006), the combination of strain and strain rate (King et al. 2003), inter-regional differences in injury tolerance (Elkin and Morrison 2007), as well as “sustained maximum principal strain” (Fijalkowski et al. 2009) that considers the duration of above-threshold strains (vs. peak strains alone in this study) could also be incorporated. These will be the subjects of future investigations.
6 Conclusion
Using WHIM to simulate brain strain responses in NFL head collisions, we found that two injury susceptibility indices based on fiber strain along WM neural tracts had the best overall performance. SLF (superior longitudinal fasciculus) appeared to be among the most injury discriminative neural tracts (e.g., AUC and accuracy up to 0.879 and 85.2%, using training and testing datasets, respectively, based on tract-wise injury susceptibilities at an optimal strain threshold of 0.10). It was also among the top most vulnerable ones, along with corpus callosum. These findings highlight the unique injury discriminatory potentials of computational models and may provide important insight into how best to incorporate WM structural anisotropy for investigation of brain injury. Future studies include (1) applying multi-variate analysis techniques to classify injury, while accounting for inter-regional differences in tolerance; (2) investigating the significance of neuroimaging inter-subject variability and accuracy of neural tracts on brain injury risk; and (3) assessing the generalizability of the findings to other at-risk populations using additional, independent injury datasets.
References
Allison Ma, Kang YS, Bolte JH et al (2014) Validation of a helmet-based system to measure head impact biomechanics in ice hockey. Med Sci Sports Exerc 46:115–123. doi:10.1249/MSS.0b013e3182a32d0d
Anderson AE, Ellis BJ, Weiss Ja (2007) Verification, validation and sensitivity studies in computational biomechanics. Comput Methods Biomech Biomed Eng 10:171–184. doi:10.1080/10255840601160484
Andersson J, Jenkinson M, Smith S (2007) Non-linear registration, aka spatial normalisation. Technical Report TR07JA2, Oxford Centre for Functional Magnetic Resonance Imaging of the Brain, Department of Clinical Neurology, Oxford University, Oxford, UK. http://www.fmrib.ox.ac.uk/analysis/techrep
Arlot S, Celisse A (2010) A survey of cross-validation procedures for model selection *. Stat Surv 4:40–79. doi:10.1214/09-SS054
Bain AC, Meaney DF (2000) Tissue-level thresholds for axonal damage in an experimental model of central nervous system white matter injury. J Biomech Eng 122:615–622. doi:10.1115/1.1324667
Bandak FA, Eppinger RH (1994) A three-dimensional finite element analysis of the human brain under combined rotational and translational accelerations. In: Proceedings, 38th Stapp Car Crash Conference, SAE paper no. 942215, pp 145–163
Bazarian JJ, Zhu T, Zhong J et al (2014) Persistent, long-term cerebral white matter changes after sports-related repetitive head impacts. PLoS One 9:e94734. doi:10.1371/journal.pone.0094734
Beckwith JG, Greenwald RM, Chu JJ (2012) Measuring head kinematics in football: correlation between the head impact telemetry system and hybrid III headform. Ann Biomed Eng 40:237–248. doi:10.1007/s10439-011-0422-2
Bigler ED, Maxwell WL (2012) Neuropathology of mild traumatic brain injury: relationship to neuroimaging findings. Brain Imaging Behav 6:108–136. doi:10.1007/s11682-011-9145-0
CDC (2015) Report to congress on traumatic brain injury in the United States: epidemiology and rehabilitation
Chappell MH, Ulug AM, Zhang L et al (2006) Distribution of microstructural damage in the brains of professional boxers: a diffusion MRI study. J Magn Reson Imaging 24:537–542. doi:10.1002/jmri.20656
Chatelin S, Constantinesco A, Willinger R (2010) Fifty years of brain tissue mechanical testing: from in vitro to in vivo investigations. Biorheology 47:255–276. doi:10.3233/BIR-2010-0576
Chatelin S, Deck C, Renard F et al (2011) Computation of axonal elongation in head trauma finite element simulation. J Mech Behav Biomed Mater 4:1905–1919. doi:10.1016/j.jmbbm.2011.06.007
Cubon VA, Putukian M, Boyer C, Dettwiler A (2011) A diffusion tensor imaging study on the white matter skeleton in individuals with sports-related concussion. J Neurotrauma 28:189–201. doi:10.1089/neu.2010.1430
Cullen DK, LaPlaca MC (2006) Neuronal response to high rate shear deformation depends on heterogeneity of the local strain field. J Neurotrauma 23:1304–1319. doi:10.1089/neu.2006.23.1304
Donnelly BR, Morgan RM, Eppinger RH (1983) Durability, repeatability and reproducibility of the NHTSA side impact dummy. Stapp Car Crash J 27:299–310
Elkin BS, Morrison B (2007) Region-specific tolerance criteria for the living brain. Stapp Car Crash J 51:127–138
Fahlstedt M, Depreitere B, Halldin P et al (2015) Correlation between injury pattern and finite element analysis in biomechanical reconstructions of traumatic brain injuries. J Biomech 48:1331–1335. doi:10.1016/j.jbiomech.2015.02.057
Fijalkowski RJ, Yoganandan N, Zhang J, Pintar FA (2009) A finite element model of region-specific response for mild diffuse brain injury. Stapp Car Crash J 53:193–213
Gardner A, Kay-Lambkin F, Stanwell P et al (2012) A systematic review of diffusion tensor imaging findings in sports-related concussion. J Neurotrauma 29:2521–2538. doi:10.1089/neu.2012.2628
Garimella HT, Kraft RH (2016) Modeling the mechanics of axonal fiber tracts using the embedded finite element method. Int J Numer Method Biomed Eng 02823:26–35. doi:10.1002/cnm.2823
Giordano C, Kleiven S (2014a) Evaluation of axonal strain as a predictor for mild traumatic brain injuries using finite element modeling. Stapp Car Crash J 58:29–61
Giordano C, Kleiven S (2014b) Connecting fractional anisotropy from medical images with mechanical anisotropy of a hyperviscoelastic fibre-reinforced constitutive model for brain tissue. J R Soc Interface 11:1–14
Hardy WN, Foster CD, Mason MJ et al (2001) Investigation of head injury mechanisms using neutral density technology and high-speed biplanar X-ray. Stapp Car Crash J 45:337–368
Hardy WN, Mason MJ, Foster CD et al (2007) A study of the response of the human cadaver head to impact. Stapp Car Crash J 51:17–80
Hernandez F, Wu LC, Yip MC et al (2014) Six degree of freedom measurements of human mild traumatic brain injury. Ann Biomed Eng 43:1918–1934. doi:10.1007/s10439-014-1212-4
Ji S, Ghadyani H, Bolander RP et al (2014a) Parametric comparisons of intracranial mechanical responses from three validated finite element models of the human head. Ann Biomed Eng 42:11–24. doi:10.1007/s10439-013-0907-2
Ji S, Zhao W (2015) A pre-computed brain response atlas for instantaneous strain estimation in contact sports. Ann Biomed Eng 43:1877–1895. doi:10.1007/s10439-014-1193-3
Ji S, Zhao W, Ford JC et al (2015) Group-wise evaluation and comparison of white matter fiber strain and maximum principal strain in sports-related concussion. J Neurotrauma 32:441–454. doi:10.1089/neu.2013.3268
Ji S, Zhao W, Li Z, McAllister TW (2014b) Head impact accelerations for brain strain-related responses in contact sports: a model-based investigation. Biomech Model Mechanobiol 13:1121–1136. doi:10.1007/s10237-014-0562-z
Kimpara H, Iwamoto M (2012) Mild traumatic brain injury predictors based on angular accelerations during impacts. Ann Biomed Eng 40:114–126. doi:10.1007/s10439-011-0414-2
Kimpara H, Nakahira Y, Iwamoto M et al (2006) Investigation of anteroposterior head-neck responses during severe frontal impacts using a brain-spinal cord complex FE model. Stapp Car Crash J 50:509–544
King AI, Yang KH, Zhang L et al (2003) Is head injury caused by linear or angular acceleration? In: IRCOBI Conference. Lisbon, pp 1–12
Kleiven S (2006) Evaluation of head injury criteria using a finite element model validated against experiments on localized brain motion, intracerebral acceleration, and intracranial pressure. Int J Crashworthiness 11:65–79. doi:10.1533/ijcr.2005.0384
Kleiven S (2007) Predictors for traumatic brain injuries evaluated through accident reconstructions. Stapp Car Crash J 51:81–114
Kraft RH, McKee PJ, Dagro AM, Grafton ST (2012) Combining the finite element method with structural connectome-based analysis for modeling neurotrauma: connectome neurotrauma mechanics. PLoS Comput Biol 8:e1002619. doi:10.1371/journal.pcbi.1002619
Kraus MF, Susmaras T, Caughlin BP et al (2007) White matter integrity and cognition in chronic traumatic brain injury?: a diffusion tensor imaging study. Brain 2508–2519. doi:10.1093/brain/awm216
Mao H, Zhang L, Jiang B et al (2013) Development of a finite element human head model partially validated with thirty five experimental cases. J Biomech Eng 135:111002–111015. doi:10.1115/1.4025101
Marjoux D, Baumgartner D, Deck C, Willinger R (2008) Head injury prediction capability of the HIC, HIP, SIMon and ULP criteria. Accid Anal Prev 40:1135–1148. doi:10.1016/j.aap.2007.12.006
McAllister TW, Flashman La, Maerlender a et al (2012) Cognitive effects of one season of head impacts in a cohort of collegiate contact sport athletes. Neurology 78:1777–1784. doi:10.1212/WNL.0b013e3182582fe7
McAllister TW, Ford JC, Flashman LA et al (2014) Effect of head impacts on diffusivity measures in a cohort of collegiate contact sport athletes. Neurology 82:63–69. doi:10.1212/01.wnl.0000438220.16190.42
Messé A, Caplain S, Paradot G et al (2011) Diffusion tensor imaging and white matter lesions at the subacute stage in mild traumatic brain injury with persistent neurobehavioral impairment. Hum Brain Mapp 32:999–1011. doi:10.1002/hbm.21092
Miller LE, Urban JE, Stitzel JD (2016) Development and validation of an atlas-based finite element brain model model. Biomech Model. doi:10.1007/s10237-015-0754-1
Mori S, Oishi K, Jiang H et al (2008) Stereotaxic white matter atlas based on diffusion tensor imaging in an ICBM template. Neuroimage 40:570–582. doi:10.1016/j.neuroimage.2007.12.035
Newman J, Shewchenko N, Welbourne E (2000) A proposed new biomechanical head injury assessment function-the maximum power index. Stapp Car Crash J 44:215–247
Newman JA, Beusenberg MC, Shewchenko N et al (2005) Verification of biomechanical methods employed in a comprehensive study of mild traumatic brain injury and the effectiveness of American football helmets. J Biomech 38:1469–1481. doi:10.1016/j.jbiomech.2004.06.025
Niogi SN, Mukherjee P, Ghajar J et al (2008) Extent of microstructural white matter injury in postconcussive syndrome correlates with impaired cognitive reaction time: a 3T diffusion tensor imaging study of mild traumatic brain injury. AJNR Am J Neuroradiol 29:967–973. doi:10.3174/ajnr.A0970
(NRC) I of M (IOM) and NRC (2014) Sports-related concussions in youth: improving the science, changing the culture. Washington, DC
Pellman EJ, Viano DC, Tucker A, Casson IR (2003) Concussion in professional football: location and direction of helmet impacts—part 2. Neurosurgery 53:1328–1341. doi:10.1227/01.NEU.0000093499.20604.21
Rice JA (2006) Mathematical statistics and data analysis, vol 3. Duxbury Advanced, Belmont
Sabet AA, Christoforou E, Zatlin B et al (2008) Deformation of the human brain induced by mild angular head acceleration. J Biomech 41:307–315. doi:10.1016/j.jbiomech.2007.09.016
Sahoo D, Deck C, Willinger R (2014) Development and validation of an advanced anisotropic visco-hyperelastic human brain FE model. J Mech Behav Biomed Mater 33:24–42. doi:10.1016/j.jmbbm.2013.08.022
Sahoo D, Deck C, Willinger R (2016) Brain injury tolerance limit based on computation of axonal strain. Accid Anal Prev 92:53–70. doi:10.1016/j.aap.2016.03.013
Shenton ME, Hamoda HM, Schneiderman JS et al (2012) A review of magnetic resonance imaging and diffusion tensor imaging findings in mild traumatic brain injury. Brain Imaging Behav 6:137–192. doi:10.1007/s11682-012-9156-5
Sullivan S, Eucker SA, Gabrieli D et al (2014) White matter tract-oriented deformation predicts traumatic axonal brain injury and reveals rotational direction-specific vulnerabilities. Biomech Model Mechanobiol. doi:10.1007/s10237-014-0643-z
Takhounts EG, Craig MJ, Moorhouse K et al (2013) Development of brain injury criteria (Br IC). Stapp Car Crash J 57:243–266
Takhounts EG, Ridella SA, Tannous RE et al (2008) Investigation of traumatic brain injuries using the next generation of simulated injury monitor (SIMon) finite element head model. Stapp Car Crash J 52:1–31
Takhounts EGE, Eppinger RRH, Campbell JQ et al (2003) On the development of the SIMon finite element head model. Stapp Car Crash J 47:107–133
Talavage TM, Nauman E, Breedlove EL et al (2014) Functionally-detected cognitive impairment in high school football players without clinically-diagnosed concussion. J Neurotrauma 31:327–338. doi:10.1089/neu.2010.1512
Viano DC, Casson IR, Pellman EJ et al (2005) Concussion in professional football: brain responses by finite element analysis—part 9. doi:10.1227/01.NEU.0000186950.54075.3B
Weaver AA, Danelson KA, Stitzel JD (2012) Modeling brain injury response for rotational velocities of varying directions and magnitudes. Ann Biomed Eng 40:2005–2018. doi:10.1007/s10439-012-0553-0
Wilcoxon F (1946) Individual comparisons of grouped data by ranking methods. J Econ Entomol 39:269. doi:10.2307/3001968
Wright RM, Post A, Hoshizaki B, Ramesh KT (2013) A multiscale computational approach to estimating axonal damage under inertial loading of the head. J Neurotrauma 30:102–118. doi:10.1089/neu.2012.2418
Wright RM, Ramesh KT (2012) An axonal strain injury criterion for traumatic brain injury. Biomech Model Mechanobiol 11:245–260. doi:10.1007/s10237-011-0307-1
Xiong K, Zhu Y, Zhang Y et al (2014) White matter integrity and cognition in mild traumatic brain injury following motor vehicle accident. Brain Res 1591:86–92. doi:10.1016/j.brainres.2014.10.030
Yang K, Mao H, Wagner C et al (2011) Modeling of the brain for injury prevention. In: Bilston LE (ed) Studies in mechanobiology, tissue engineering and biomaterials. Springer-Verlag, Berlin, pp 69–120
Zhang K, Johnson B, Pennell D et al (2010a) Are functional deficits in concussed individuals consistent with white matter structural alterations: combined FMRI and DTI study. Exp Brain Res 204:57–70. doi:10.1007/s00221-010-2294-3
Zhang L, Yang KH, King AI (2004) A proposed injury threshold for mild traumatic brain injury. J Biomech Eng 10(1115/1):1691446. doi:10.1115/1.1691446
Zhang Y, Zhang J, Oishi K et al (2010) Atlas-guided tract reconstruction for automated and comprehensive examination of the white matter anatomy. Neuroimage 52:1289–1301. doi:10.1016/j.neuroimage.2010.05.049
Zhao W, Ji S (2015) Parametric investigation of regional brain strain responses via a pre-computed atlas. IRCOBI Conference. Lyon, pp 208–220
Zhao W, Ji S (2016) Brain strain uncertainty due to shape variation in and simplification of head angular velocity profiles. Biomech Model Mechanobiol. doi:10.1007/s10237-016-0829-7
Zhao W, Ford JC, Flashman LA et al (2016) White matter injury susceptibility via fiber strain evaluation using whole-brain tractography. J Neurotrauma 33:1834–1847. doi:10.1089/neu.2015.4239
Acknowledgements
Funding is provided by the NIH Grants R01 NS092853 and R21 NS088781. The authors are grateful to the National Football League (NFL) Committee on Mild Traumatic Brain Injury (MTBI) and Biokinetics and Associates Ltd. for providing the reconstructed head impact kinematics. The authors thank Dr. Thomas W. McAllister at Indiana University for providing neuroimaging data, Dr. James C. Ford and Dr. Laura A. Flashman at Dartmouth College, and Dr. Richard M. Greenwald and Mr. Jonathan Beckwith at Simbex, LLC, for their help. In addition, they thank Dr. Qiang Liu at Dartmouth College, and Ms. Chiara Giordano at Royal Institute of Technology (KTH) for helpful discussions.
Conflict of interest
The authors declare that they have no competing interests.
Author information
Authors and Affiliations
Corresponding author
Appendices
Appendix 1: Description of deep brain white matter atlas
Appendix 2: One-sided permutation test
We adopted a non-parametric, one-sided permutation test (Rice 2006) to compare injury prediction performances and vulnerabilities of the deep WM ROIs/neural tracts. A paired-sample t-statistic is first calculated according to:
where \(\bar{d}\) is the mean of the difference between two samples, x and y; s is the standard deviation of d; and n is the sample size. The pseudo-algorithm for the permutation test is described below:
- Step 1 :
-
For a given pair of data samples, calculate the baseline paired-sample t-statistic, \(T_0 \), using Eqns. 8 and 9.
- Step 2 :
-
Randomly flip the pairs of values from the two samples to generate two new samples.
- Step 3 :
-
Compute a new paired-sample t-statistic, \(T_{\mathrm{perm}} \), using the newly generated samples from Step 2.
- Step 4 :
-
Repeat Steps 2–3 by \(N_{\mathrm{perm}} \) times (e.g., 100).
- Step 5 :
-
Calculate a probability p value:
$$\begin{aligned} p=\frac{\# (T_{\mathrm{perm}} >T_0 )}{N_{\mathrm{perm}}}, \end{aligned}$$(10)
where “\(\# (T_{\mathrm{perm}} >T_0 )\)” is the number of permutations where \(T_{\mathrm{perm}}\) is found to be greater than \(T_0\). A p value smaller than a given threshold (e.g., 0.05) is considered a strong indication that the given pair has a statistically significant difference.
Rights and permissions
About this article

Cite this article
Zhao, W., Cai, Y., Li, Z. et al. Injury prediction and vulnerability assessment using strain and susceptibility measures of the deep white matter. Biomech Model Mechanobiol 16, 1709–1727 (2017). https://doi.org/10.1007/s10237-017-0915-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10237-017-0915-5