
Abstract
The giant mottled eel Anguilla marmorata Quoy & Gaimard is an important aquaculture candidate in eel farming industry. The high economic value of the species leads to intensive cultivation affecting its welfare under farm condition. Since colour environment affects the welfare of confined fishes, basic information on colour vision is necessary. By means of a behavioural experiment of A. marmorata elvers in the laboratory, we determined colour vision under natural daylight. The elvers were subjected to classical conditioning to associate a reward feed with a blue or a green stimulus placed amid seven shades of grey. The elvers learned this visual task after 30 or 36 trials and thus the ability of colour discrimination was confirmed. Since the elvers have the retinal tapetum, it may be suggested that colour vision in dim light enables crepuscular feeding by the A. marmorata elvers, clearly an advantage in aquaculture.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The tropical giant mottled eel Anguilla marmorata Quoy and Gaimard is the second largest of the anguillid eels and a highly valued aquatic product of major economic importance for the Chinese market. The Chinese market for eel aquaculture was originally dominated by the Japanese eel Anguilla japonica. However, to meet the global demand for consumption, the species faced a dramatic decrease in the wild population due to over fishing, and other factors such as habitat destruction and climate change (Casselman 2003; Dekker 2003; Tatsukawa 2003). Due to this reason, the International Union for Conservation of Nature listed A. japonica as endangered species, thus causing the rise in the market price for the species (Dekker 2004; Han et al. 2009; Jacoby and Gollock 2014). Henceforth, A. marmorata became the new emerging eel species for aquaculture, mostly concentrated in Hainan, Guangdong and Fujian provinces (Luo et al. 2013).
In Taiwan, the glass eels of A. marmorata are largely imported from the Philippines and cultivated in outdoor soil ponds, concrete ponds and indoor ponds (Huang et al. 2016). At present, eel aquaculture is completely dependent on wild catch, while the artificial propagation of the species has not yet succeeded. According to Huang et al. (2016), the average survival is low, 20−30%, and improvements in farming technology and management are required. Unlike the temperate eel species, the historical data on the biology and physiology of the tropical eels are still very scarce. Research studies are therefore important to enhance wild eel stocks and continue their commercial use for human consumption, with the main goal of course, to protect and retrieve the welfare of the wild eel population (Arai 2014).
Exposure of confined fish to poor conditions may compromise their welfare and production (Migaud et al. 2007). Many factors affect the production of A. marmorata such as stocking density, feed quality, temperature, water quality, pond or tank design (Cheng 2011; Huang et al. 2016; Wang et al. 2016). Colour of tanks and light wavelength range might also affect feeding, growth, survival and stress response as noted in many other fish species (reviewed in Kawamura et al. 2015). The American eel Anguilla rostrata and the European eel Anguilla anguilla are reported to have two types of single cone photoreceptors with peak spectra at around 440 nm (blue) and 525 nm (green) (Gordon et al. 1978; Damjanović et al. 2005; Cottrill et al. 2009) indicating possession of colour vision. On the other hand, A. marmorata is reported to have a single type of cone with a λmax at 508 nm and rod with a λmax at 489 nm during yellow phase predicting colour blindness (Wang et al. 2014). Colour vision has been claimed or speculated in fishes based on the types of photoreceptors, but only behavioural studies can show that a given visual system is capable of this ability (Douglas and Hawryshyn 1990).
The tapeta lucida, retinal tapeta and choroidal tapeta, are found in fishes of wide taxa and visually recognized as eye shine (Somiya 1980). The retinal tapetum is a specialized reflecting surface that increases the number of times that low-intensity light can pass through the visual cells, and thus confers greater scotopic sensitivity to the eye (Nicol 1981). However, according to Munk (2010), the retinal tapetum of mesopelagic fish Lestidiops affinis is present in the pure-single cone region apposing to the tip of each cone outer segment. This suggests that the retinal tapetum functions to increase the photopic vision sensitivity thereby colour vision sensitivity.
Since the retinal tapetum has not been reported in Anguillidae, we determined the presence of the retinal tapetum histologically. The present study, for the first time, demonstrated colour vision of the tropical A. marmorata determined behaviourally and the presence of the retinal tapetum in the retina of A. marmorata.
Materials and methods
Elvers and husbandry. The Anguilla marmorata elvers were imported from a local fish supplier in Indonesia with permission by the Fisheries Department of Malaysia. The species was ascertained based on the morphological characteristics of the tooth pattern from the upper jaw (Tesch 2003). They were maintained in two 80 L glass aquaria equipped with a 100 L water filtration unit. Each aquarium was covered with black cloths to avoid stressors from the surroundings. The elvers were fed three times daily with marine sinking feed (Cargill Feed Sdn. Bhd., Kuala Lumpur, Malaysia).
Colour discrimination conditioning trials. The classical conditioning test was conducted. von Frisch’s (1950) “grey-card experiment” with bees is a convincing demonstration of colour vision (Kelber et al. 2003). An animal learns to associate a reward with a colour, and then chooses between this colour and many shades of grey. If all grey shades are discriminable from the colour, the animal is not relying on achromatic cues but on chromatic cues. The methodology was adapted to the present study.
The colour discrimination conditioning trials were carried out under natural daylight inside a roofed fish hatchery of the Universiti Malaysia Sabah with two groups of elvers in two glass test aquaria (100 cm × 45 cm, filled to a depth of 40 cm) with continuous aeration and recirculating water through a biological filter. Light and water variables were measured twice daily, in the morning and afternoon. The experiments were always performed in shady conditions at from 0800 to 1700. The illuminance ranged from 70 to 150 lx (measured with a Light meter 401036, Extech Instruments, USA). Water temperature ranged 26.5–27.4 °C; dissolved oxygen 7.5−8.2 mg/L; and pH 7.4−8.4 (Hanna Instruments, HI 9828, Washington, USA).
Fish learn colour discrimination faster in groups than singly (Beyer 1976), and A. rostrata and A. anguilla have visual spectral sensitivity with peaks at around 440 nm (blue) and 525 nm (green) wavelengths (Gordon et al. 1978; Damjanović et al. 2005). Thus, the conditioning experiment trained groups of 10 A. marmorata elvers (mean total length 13.89 ± 0.49 cm SD; mean body weight 3.00 ± 0.22 g) to associate a feed reward with either a blue or green feed well amid seven other wells in different shades of grey under natural daylight. Two paint palettes with eight wells were used in the tests. One palette had one well-painted blue (Nippon Q-Lac oil blue) and the other palette had one well-painted green (Nippon Q-Lac oil green) as the reward colours. The other seven wells painted different shades of grey were left empty of feed (Fig. 1). A blue well was used in the test aquarium A and the other palette with a green well was used in the test aquarium B. An artificial feed (moist marine pellet, Cargill Sdn. Bhd., Malaysia) was formed into dough with stocking water from the rearing aquarium and 10 g of it was placed on the blue and green wells as a feed reward.

White palettes with a blue- or green-painted well among seven wells painted different shades of grey. The blue and green wells contained a feed pellet as reward during the conditioning trials and no reward during the colour vision tests of Anguilla marmorata elvers
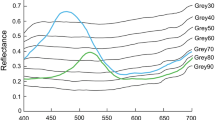
Shades of grey were produced by mixing white paint (Nippon Q-Lac oil white) and black paint (Nippon Q-Lac oil black) at seven different ratios: 30, 40, 50, 60, 70, 80 and 90% black), referred hereafter as Grey 30, Grey 40, Grey 50, Grey 60, Grey 70, Grey 80 and Grey 90, and applied in the arrangement as shown in Fig. 1. The spectra of light reflected from the blue, green and grey paints were recorded with a spectroradiometer (HSR-8100, Maki Manufacturing Co., Ltd., Hamamatsu, Japan) in the wavelength range of 400−700 nm (Fig. 2). Since behavioural response of fish to coloured objects changes according to types of artificial light (Kawamura et al. 2017a), the spectral measurements were performed at 1130 and 1700 (one hour before the sunset) under natural daylight inside the roofed fish hatchery without artificial light source. While the light intensity level was lower at 1700 than at 1130, there was no notable difference in spectral pattern between the spectra.

a Spectra of background light measured with a spectroradiometer at 1130 (small a) and at 1700 (small b), and b light reflected spectra from blue and green paints and seven shades of grey. All spectra were measured in natural light
Conditioning sessions started after a day of acclimation and continued for 9 days in the test aquarium A and 10 days in the test aquarium B. Before the first morning trial, about 0.5 g of feed dough was given to the elvers to prevent the negative effect of hunger on learning (Kawamura et al. 2002). Each day, one conditioning session with 6 trials was done to each group of elvers at 1 h intervals. The dough was consumed within 1 min. After 1 min, the palettes were taken out, washed in running tap water, air-dried, and prepared for the next trial. In each subsequent trial, position of the palettes was rearranged randomly to ensure that the elvers did not associate the reward feed with any particular position but rather the well colour.
Non-reward test trials. After each day’s conditioning session (6 trials; from 6th to 11th day for green well training, from 5th to 10th day for blue well training), the palette was replaced with an another palette which had never been used in conditioning test trials, and a non-reward test trial was conducted for 5 min, where the elvers exposed to the palette without the feed reward in the colour well and video recorded with a digital camera (IXUS 160, Canon, Tokyo, Japan). The number of visits by the elvers to each well was later counted, inevitably including the repeated visits of the same as individual identification was difficult. The elvers exhibited following behavioural responses to the wells: (i) stopped swimming and placed the head in a well for 1 to 4 s, (ii) stopped swimming and rubbed the well surface with the mouth. These behaviours were considered as conditioned responses and defined as vising well behaviour. When these behaviours were exhibited in two or more wells, all visits were counted.
Histological examination of the retinal tapetum. Three light adapted elvers were killed with an overdose of MS222 and preserved in Bouin’s solution. For histological observation of the retinae, the eyes of fixed samples were dehydrated in a graded series of ethanol, embedded in paraffin, cut into 6 μm-thick sections, and stained with haematoxylin and eosin for examination of the retina under a light microscope (ECLIPSE 80i, Nikon). The presence of the retinal tapetum was ascertained by illuminating histological preparations with oblique surface light from a light guide (LS-JHA, Sumita Optical Glass, Inc., Saitama, Japan) under a light microscope. The retinal tapetum shines under surface lighting due to birefringence (Kawamura et al. 2016).
Statistical analysis. The frequencies of the visit by the elvers in the non-reward test trials were statistically analysed by the χ2 test to assess the goodness of fit between a set of observed values and those expected theoretically, i.e., based on the hypotheses that the probability of choice for each well is 1/8. The significance level was set at α = 0.05. The mean frequencies of the visit by the elvers in the six non-reward tests trials were calculated for eight wells and the 95% confidence interval of each mean frequency was calculated.
Results
Non-reward test. During the conditioning trials, the elvers slowly approached the feeding palette and randomly swam around the wells. They then detected the feed reward on the conditioning colour wells and started to feed on the dough vigorously in mass. Both of the palettes were emptied within 1 min. Empty feeding palettes were immersed back again after complete session of conditioning trials. In the early sessions, the elvers were observed swimming randomly around the wells in search for the food reward provided earlier. The elvers slowly showed learning behaviour towards colour discrimination when they significantly swam towards the green or blue conditioning colour well rather than the grey shades.
During the non-reward test trials, the elvers exhibited the conditioned behaviour, stopped swimming and rubbed the well surface with the mouth, only in the conditioned colour well, green or blue. In the 1st trial, the elvers showed significantly biased visit frequency to the conditioned colour well, χ2 = 32.4 (P < 0.01) for green well, χ2 = 38.05 (P < 0.01) for blue well (Table 1), indicating that the elvers have learnt the conditioning task after 36 (green) or 30 (blue) conditioning trials. There was no increasing tendency in the visiting frequency to the correct colour well until the last non-reward test trial. The mean frequency of visit to each colour well and its 95% confidence interval was significantly higher for the conditioned colour well, green or blue (Table 1), indicating that the elvers learnt to discriminate green or blue from various shades of grey.
Presence of the retinal tapetum. Fig. 3 shows two photomicrographs of the same histological section taken under conventional transmitted light through a condenser lens and illumination with oblique surface light from a light guide. The shining pigment epithelium layer shows the presence of the retinal tapetum.

Microphotographs of a cross section of light-adapted retina of an elver of Anguilla marmorata. a Under conventional lighting through a condenser lens, single cone and rod ellipsoids are indicated by white arrowheads, respectively; b under surface lighting. PE pigment epithelium layer; C single cone; R rod
Discussion
This study demonstrated that the Anguilla marmorata elvers learned to discriminate blue or green from different shades of grey, indicating the possession of colour vision. This result is inconsistent with the finding by Wang et al. (2014) who conducted microspectrophotometry (MSP) of A. marmorata and reported a green sensitive single cone type, which was supported by their phylogenetic analysis of opsin genes, and concluded that this eel is colour-blind. According to Phillips et al. (2015), MSP is difficult to perform on fish retina with the thick pigment epithelium in close contact to the cone cell outer segments, causing damage to these parts of the cell when the pigment is removed. Bowmaker et al. (2008) reported two types, blue or green sensitive, of single cones based on MSP experiment of the European eel A. anguilla and noted that blue-sensitive cones were rarely recorded. Byzov et al. (1998) examined the retinae of A. anguilla electrophysiologically and reported that yellow-sensitive and green-sensitive cones coexist in the retina indicating capacity of colour vision in this eel. This study was supported by Damjanović et al. (2005) who examined the retinae of A. anguilla by electrophysiology and MSP.
Based on the results of present study we conclude that A. marmorata elvers have colour vision. As mentioned above, while colour vision has been claimed or speculated in fishes based on the types of photoreceptors, only behavioural studies can show a capable achievement of the given visual system (Douglas and Hawryshyn 1990). However, this conclusion begs the question, “How is colour vision important in this nocturnal eel?” Anguilla marmorata is reportedly nocturnal in the wild (Setiyanto et al. 2003). But the difference between day and night activity is not substantial (Tesch 2003). Salazar (1997) reported the diurnal activity pattern of yellow phase A. marmorata in three clear streams in Moorea, French Polynesia; the number of specimens observed peaked in the late afternoon between 1630 and 1700 and secondary peak in the morning between 0830 and 0930 in the streams.
Of interest is that A. marmorata has the retinal tapetum. Since the retinal tapetum surrounds the outer segments of cones in light-adapted retina and moves in association with the cones (Awaiwanont et al. 2001; Kawamura et al. 2016), it can enable to increase the sensitivity of cones in dim light. The larvae of African catfish Clarias gariepinus can see colour down to 0.01 lx (similar to star-light), due to the presence of retinal tapetum (Kawamura et al. 2017b). Thus, the elvers might be capable of colour vision in dim light. However, this speculation of the vision in dim light needs to be behaviourally demonstrated.
Anguilla marmorata has two type of photoreceptors, rods and single cones, and lacks double cones (Wang et al. 2014). Colour vision with single cones is also reported for other fish and several reptiles. The retina of the kyusen wrasse Halichoeres poecilopterus lacks the double cone (Kawamura and Tamura 1973). Phillips et al. (2015) reported two single cone opsin genes in this fish: short-wavelength sensitive and long-wavelength opsin genes. Sillman et al. (1992) did MSP study of the retina of the ball python Python regius and reported two distinct types of single cone; a stout single cone which contains a visual pigment with λmax at 552 nm, and a slender single cone with λmax at 360 nm. By analyzing the visual pigments of two henophidian snakes, Xenopeltis unicolor and Python regius which lack double cones, Davies et al. (2009) showed that both species express two cone opsins, a short-wavelength-sensitive (λmax = 361 nm) pigment and a long-wavelength-sensitive (λmax = 550 nm) pigment, providing the potential for dichromatic colour vision. Birds colour vision has been behaviourally determined and it is mediated by four types of single cone sensitive to ultraviolet, short- median- or long-wavelength light (Osorio et al. 1999). Thus, colour vision mediated by single cones is common.
While importance of olfaction in feeding of eels is well accepted (reviewed in Deelder 1984), there are evidences for the involvement of vision in feeding behaviour. Knights (1983) examined feeding behaviours of the A. anguilla elvers in the laboratory and reported that food particles were located visually from a distance and elvers moved in a straight line towards them, and the selection of optimum-sized particles was done by vision. The food searching by vision might be also true for the A. marmorata elvers. Elvers utilize drifting or inundated food items such as insects and terrestrial annelids (Tesch 2003). The precise location of such the food can be done only by vision. According to Romanda et al. (2019), crabs and shrimps were the main items (86%) in the stomach contents of A. marmorata and the feeding habit was more or less selective. The selective carnivorous predation might be done by vision. Thus, the involvement of vision in feeding of elvers is clear.
Tank experiments have shown that the ability of fish larvae and juveniles to acquire food is largely influenced by prey visibility resulting from the contrast with the background environment (Hinshaw 1985; Sandström 1999). Chromatic information is important for object-contour perception and chromatic objects confer greater contrast than achromatic ones against background (Hansen and Gegenfurtner 2017). The presence of the retinal tapetum would increase colour vision sensitivity in dim light and enable crepuscular feeding by the A. marmorata elvers and extends the daily feeding period, clearly an advantage in aquaculture.
As the colours of the farm environment have significant effects on growth and the stress response of aquaculture species (Kawamura et al. 2015), the effects of colour of tank wall and light should be examined. Since the feeding of A. marmorata is more or less selective (Setiyanto et al. 2003), feed colour preference also should be studied. It is noteworthy that the expression of retinal opsins alters with growth stages in A. marmorata (Wang et al. 2014) and A. anguilla (Cottrill et al. 2009), the effect of environmental colour and feed colour preference possibly change with growth stages. All this information would contribute to the better welfare of A. marmorata in farms.
References
Arai T (2014) Do we protect freshwater eels or do we drive them to extinction? SpringerPlus 3:534
Awaiwanont K, Gunarso W, Sameshima M, Hayashi S, Kawamura G (2001) Grouped, stacked rods and tapeta lucida in the retina of Japanese anchovy Engraulis japonicus. Fish Sci 67:804–810
Beyer I (1976) The effect of grouping and isolation on the learning process of a schooling fish (Scardinius erythrophthalmus L.). Behav Ecol Sociobiol 1:245–263
Bowmaker JK, Semo M, Hunt DM, Jeffery G (2008) Eel visual pigments revisited: the fate of retinal cones during metamorphosis. Vis Neurosci 25:249–255
Byzov AL, Damjanovic I, Utina IA, Mickovic B, Gacic Z, Andjus RK (1998) Electrophysiological and spectral properties of second-order retinal neurons in the eel. Comp Biochem Physiol A Mol Integr Physiol 121:197–208
Casselman JM (2003) Dynamics of resources of the American eel, Anguilla rostrata: declining abundance in the 1990s. In: Aida K, Tsukamoto K, Yamauchi K (eds) Eel biology. Springer, Tokyo, pp 255–274
Cheng WT (2011) Development and extension of swamp eel (Anguilla marmorata). Research report of council of agriculture. 99AS-10.3.1-FA-F1(2). NPUST, Pingtung, Taiwan (in Chinese with English abstract)
Cottrill PB, Davies WL, Semo M, Bowmaker JK, Hunt DH, Jeffery G (2009) Developmental dynamics of cone photoreceptors in the eel. BMC Dev Biol 9:71
Damjanović I, Byzov AL, Bowmaker JK, Gačić Z, Utina IA, Maximova EM, Mićković BM, Andjus AK (2005) Photopic vision in eels: evidences of color discrimination. Ann N Y Acad Sci 1048:69–84
Davies WL, Cowing JA, Bowmaker JK, Carvalho LS, Gower DJ, Hunt DM (2009) Shedding light on serpent sight: the visual pigments of henophidian snakes. J Neurosci 29:7519–7525
Deelder CL (1984) Synopsis of biological data on the eel Anguilla anguilla (Linnaeus, 1758). FAO Fisheries Synopsis, no. 80, revision 1. FAO, Rome
Dekker W (2003) Status of the European eel stock and fisheries. In: Aida K, Tsukamoto K, Yamauchi K (eds) Eel biology. Springer, Tokyo, pp 237–254
Dekker W (2004) Slipping through our hand: population dynamics of the European eel. Doctoral Dissertation, University of Amsterdam
Douglas RH, Hawryshyn CW (1990) Behavioral studies of fish vision: an analysis of visual capabilities. In: Douglas RG, Djamgoz MBA (eds) The visual system of fish. Chapman and Hall, London, pp 373–418
Gordon J, Shapley RM, Kaplan E (1978) The eel retina. Receptor classes and spectral mechanisms. J Gen Physiol 71:125–138
Han YS, Tzeng WN, Liao IC (2009) Time series analysis of Taiwanese catch data of Japanese glass eels Anguilla japonica: possible effects of the reproductive cycle and El Niño events. Zool Stud 48:632–639
Hansen T, Gegenfurtner KR (2017) Color contributes to object-contour perception in natural scenes. J Vis 17:1–19
Hinshaw JM (1985) Effects of illumination and prey contrast on survival of larval yellow perch, Perca flavescens. Trans Am Fish Soc 114:540–545
Huang CT, Chiou JT, Khac HT, Hsiao YJ, Chen SC (2016) Improving the management of commercial giant mottled eel Anguilla marmorata aquaculture in Taiwan for improved productivity: a bioeconomic analysis. Fish Sci 82:95−111
Jacoby D, Gollock M (2014) Anguilla japonica. The IUCN Red list of threatened species. Version 2014.3. http://www.iucnredlist.org. Accessed 22 February 2021
Kawamura G, Tamura T (1973) Morphological studies on the retina of two teleosts Scomber tapeinocephalus and Halichoeres poecilopterus. Bull Jpn Soc Sci Fish 39:715–726
Kawamura G, Matsunaga F, Tanaka Y (2002) Color sense of loggerhead turtle Caretta caretta as determined by discrimination conditioning. Nippon Suisan Gakkaishi 68:542–546
Kawamura G, Bagarinao TU, Lim LS (2015) Fish behaviour and aquaculture. In: Mustafa S, Shapawi R (eds) Aquaculture ecosystems: adaptability and sustainability. John Wiley & Sons Limited, West Sussex, UK, pp 68–106
Kawamura G, Bagarinao T, Justin J, Chen CY, Lim LS (2016) Early appearance of the retinal tapetum, cones, and rods in the larvae of the African catfish Clarias gariepinus. Ichthyol Res 63:536–539
Kawamura G, Bagarinao TU, Asmad MFB, Lim LS (2017a) Food colour preference of hatchery-reared juvenile of African catfish Clarias gariepinus. Appl Anim Behav Sci 196:119-122
Kawamura G, Bagarinao T, Hoo PK, Justin J, Lim LS (2017b) Colour discrimination in dim light by the larvae of the African catfish Clarias gariepinus. Ichthyol Res 64:204–211
Kelber A, Vorobyev M, Osorio D (2003) Colour vision in animals—behavioural tests and physiological concepts. Biol Rev 78:81–118
Knights B (1983) Food particle-size preferences and feeding behaviour in warmwater aquaculture of European eel, Anguilla anguilla (L.). Aquaculture 30:173–190
Luo MZ, Guan RZ, Li ZQ, Jin H (2013) The effects of water temperature on the survival, feeding, and growth of the juveniles of Anguilla marmorata and A. bicolor pacifica. Aquaculture 400–401:61–64
Migaud H, Cowan W, Taylor J, Ferguson W (2007) The effect of spectral composition and light intensity on melatonin, stress and retinal damage in post-smalt Atlantic salmon, Salmo salar. Aquaculture 270:390–404
Munk O (2010) Duplex retina in the mesopelagic deep-sea teleost Lesitidiops affinis (Ege, 1930). Acta Zool 70:143–149
Nicol JAC (1981) Tapeta lucida of vertebrates. In: Enock JM, Toby FL Jr (eds) Vertebrate photoreceptor optics. Springer-Verlag, Berlin, pp 401–431
Osorio D, Vorobyev M, Jones CD (1999) Colour vision of domestic chicks. J Exp Biol 202:2951–2959
Phillips GAC, Carleton KL, Marshall NJ (2015) Multiple genetic mechanisms contribute to visuals variation in the Labridae. Mol Biol Evol 33:201–215
Romanda R, Putra DF, Dewiyanti I, Nurfadillah N, Batubara AS, Mustaqim M, Muthmainnah CR, Nur FM, Muchlisin ZA (2019) Feeding habits and length-weight relationship of giant marbled eel Anguilla marmorata in the Brayeun River, Aceh Besar District, Aceh Province, Indonesia. IOP Conf Ser: Earth Environ Sci 348:012035
Salazar CE (1997) Diurnal behaviors of Anguilla marmorata in streams on Moorea, French Polynesia. In: Gump RB (ed) The biology and geomorphology of tropical islands. University of California, Berkeley, pp 38–46
Sandström AL (1999) Visual ecology of fish - a review with special reference to percids. Fiskeriverket Rapport 2:45–80
Setiyanto, Yuwono E, Sulistyo I, Sakardi P, Sulistyo I (2003) Study on feeding behaviour of eels and the larvae occurrence in Segara Anakan. Proceeding of Tropical Eel Fishery Resource. BPPT, Ministry of Research and Technology, Indinesia
Sillman AJ, Carver JK, Loew ER (1992) The photoreceptors and visual pigments in the retina of a boid snake, the ball python (Python regius). J Exp Biol 202:1931–1938
Somiya H (1980) Fishes with eye shine: functional morphology of guanine type tapetum lucidum. Mar Ecol Prog Ser 2:9–26
Tatsukawa K (2003) Eel resources in east Asia. In: Aida K, Tsukamoto K, Yamauchi K (eds) Eel biology. Springer, Tokyo, pp 293–298
Tesch FW (2003) The eel (3rd ed). Blackwell Science Ltd, Oxford, UK
von Frisch K (1950) Bees. Their vision, chemical sense, and language. Cornell University Press, London
Wang FY, Fu WC, Wang IL, Yan HY, Wang TY (2014) The giant mottled eel, Anguilla marmorata, uses blue-shifted rod photoreceptors during upstream migration. PLoS ONE 9(8):e103953
Wang L, Fu WC, Wang X, Yin S (2016) Effects of salinity change on two superoxide dismutases (SODs) in juvenile marbled eel Anguilla marmorata. PeerJ 4:e2149
Acknowledgments
The authors would like to thank all the supporting staffs of the Fish Hatchery of Universiti Malaysia Sabah for their endless support, and providing materials and facilities throughout this study. This study was funded by UMS External Collaboration Research Grant: GKP0002-STWN-2016. All the experimental trials were conducted in accordance with the Researcher’s Guidelines On Code Of Practice For The Care And Use Of Animals For Scientific Purposes Universiti Malaysia Sabah. The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
About this article

Cite this article
Ching, F.F., Joshua, M.S., Kawamura, G. et al. Colour vision in the tropical giant mottled eel Anguilla marmorata as determined by classical conditioning test. Ichthyol Res 69, 90–96 (2022). https://doi.org/10.1007/s10228-021-00816-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10228-021-00816-3