
Abstract
The new cardinalfish Rhabdamia novaluna sp. nov. (Apogonidae) is described on the basis of 79 specimens collected from the western Pacific Ocean. The new species is most similar to the Indo-West Pacific species Rhabdamia gracilis (Bleeker 1856) in having VI-I, 9 dorsal-fin rays, II, 12 anal-fin rays and 13 pectoral-fin rays, and lacking blackish blotches on the opercle and anterior portion of the body. However, the former can be distinguished from R. gracilis by having 26–31 (modally 28–29) developed gill rakers, 27–31 (modally 28) total gill rakers (including rudiments), body depth at second dorsal-fin origin 25.5–33.1% of SL (mean 30.2%) and a subdermal black blotch or spot on the lower caudal peduncle. Three nominal species, Apogonichthys nudus Regan 1905, Apogonichthys mentalis Evermann and Seale 1907 and Rhabdamia clupeiformis Weber 1909, are regarded as junior synonyms of R. gracilis. A key to species of Rhabdamia is provided.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The apogonid genus Rhabdamia Weber 1909 (Apogonidae) is characterized by VI-I, 9 or VII-I, 10–11 dorsal-fin rays, II, 10–13 anal-fin rays, a forked caudal fin, two supraneurals and five hypurals (first to fourth hypurals firmly fused, fifth hypural free) (Fraser 1972; Mabuchi et al. 2014; Yoshida and Motomura 2018).
During a revisionary study of Rhabdamia, 79 unidentified specimens from the western Pacific Ocean were located in museum collections and collected during field surveys by the authors. Although similar to Rhabdamia gracilis (Bleeker 1856) in having VI-I, 9 dorsal-fin rays, II, 12 anal-fin rays and 13 pectoral-fin rays, and lacking 3–6 reddish-brown to blackish blotches on the opercle and anterior portion of the body, the specimens differed in having a higher number of gill rakers, greater body depth and a subdermal black blotch or spot on the lower caudal peduncle. They are described herein as a new species of Rhabdamia and compared in detail with the Indo-West Pacific species R. gracilis, including an assessment of the synonymies of the latter. A key to species of Rhabdamia is also provided.
Materials and methods
Counts and measurements followed Yoshida and Motomura (2016), with the following additions: body depth at the first dorsal-fin origin was measured between the first dorsal-fin spine and pelvic-fin spine origins, that at the second dorsal-fin origin between the origins of the first spine of the second dorsal fin and first anal-fin spine. Measurements were made to the nearest 0.1 mm with needlepoint calipers under a dissecting microscope. Standard and head lengths are abbreviated as SL and HL, respectively. Curatorial procedures for the newly collected specimen followed Motomura and Ishikawa (2013). Osteological characters, including vertebral counts, were observed from radiographs of specimens of Rhabdamia gracilis which have been listed in Yoshida and Motomura (2018), with the following additions: 12 specimens of Rhabdamia novaluna sp. nov. (KPM-NI 18687–18696, 10 specimens; URM-P 4726, 2 specimens) and one specimen of R. gracilis (USNM 55905, 52.2 mm SL). The formula for the configuration of supraneural bones, anterior neural spines and anterior dorsal-fin pterygiophores follows Ahlstrom et al. (1976). Institutional codes follow Sabaj (2016).
Key to the species of Rhabdamia
1a. First dorsal fin with 7 spines; second dorsal fin with one spine and 11 soft rays; anal fin with 2 spines and 11 soft rays; pectoral fin rays 16………R. nigrimentum (Red Sea)
1b. First dorsal fin with 6 spines; second dorsal fin with one spine and 9 soft rays; anal fin with 2 spines and 12 soft rays; pectoral fin rays 13……………………………………2
2a. Three to six reddish-brown to blackish blotches on opercle and anterior portion of body……………………………………………………………………….R. spilota (Indo-West Pacific)
2b. Blackish blotches absent on opercle and anterior portion of body…………………………………..……..3
3a. Body at second dorsal-fin origin relatively deep, its depth 25.5–33.1% of SL (mean 30.2%); high numbers of developed gill rakers 26–31 (mode 28–29) and total gill rakers (including rudiments) 27–31 (28); a subdermal black blotch or spot on the lower caudal peduncle………………………………………………R. novaluna sp. nov. (western Pacific Ocean)
3b. Body at second dorsal-fin origin relatively shallow, its depth 19.2–30.6% of SL (mean 25.7%); low numbers of developed gill rakers 22–28 (mode 24) and total gill rakers (including rudiments) 23–28 (25); no subdermal black blotch or spot on the lower caudal peduncle…………………………………………………………R. gracilis (Indo-West Pacific)
Rhabdamia novaluna sp. nov.
(New English name: New-moon Cardinalfish; new Japanese name: Shingetsu-sukashi-tenjikudai) (Figs. 1, 2, 3, 4; Tables 1, 3–4)

Preserved holotype of Rhabdamia novaluna (KPM-NI 18695, 43.5 mm SL, Tokashiku Bay, Tokashiki-jima island, Okinawa, Japan)

Fresh paratype of Rhabdamia novaluna (KAUM–I. 106135, 29.6 mm SL, Ishigaki-jima island, Okinawa, Japan)

Caudal peduncle of preserved specimens of Rhabdamia novaluna. a holotype (KPM-NI 18695, 43.5 mm SL, Japan); b paratype (KAUM–I. 2425, 36.0 mm SL, Australia)

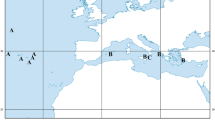
Distributional records of Rhabdamia novaluna, based on specimens examined in this study
Rhabdamia gracilis (not of Bleeker): Ida and Moyer 1974: 117, fig. 5K (Okinawa-jima and Iriomote-jima islands, Okinawa, Japan); Hayashi and Kishimoto 1983: 39, fig. 48 (Iriomote-jima island, Okinawa, Japan); Hayashi 1993: 662, unnumbered fig. on bottom (southward Wakayama Prefecture, Japan); Masuda and Kobayashi 1996: 134, figs. 3–4 (Kerama and Yaeyama Islands, Okinawa, Japan); Hayashi 1997: 305, unnumbered figs. on bottom (Kerama Islands and Iriomote-jima island, Okinawa, Japan); Yoshigou and Nakamura 2002: 104 (Iriomote-jima island, Okinawa, Japan); Yoshigou et al. 2005: 18 (Okinawa-jima island, Okinawa, Japan); Randall 2005: 213, unnumbered fig. on bottom (Great Barrier Reef, Australia); Senou et al. 2006: 74 (Ie-jima island, Okinawa, Japan); Senou et al. 2007: 54 (Miyako Group, Okinawa, Japan); Watai et al. 2009: 124 (Tokashiki-jima island, Okinawa, Japan); Yoshida et al. 2010: 56, figs. 40A, B (Yaku-shima island, Kagoshima, Japan); Allen and Erdmann 2012: 411, unnumbered fig. on middle (Bali, Indonesia).
Rhabdamia sp.: Yoshida and Motomura (2018): 182 (considered to be an undescribed species).
Holotype. KPM-NI 18695, 43.5 mm SL, Tokashiku Bay, Tokashiki-jima island, Kerama Islands, Okinawa, 5–7 m depth, J. Kaiga, 14 March 2006.
Paratypes. 78 specimens, 22.4–49.6 mm SL. JAPAN: KAUM–I. 106135, 29.6 mm SL, Ishigaki-jima island, Yaeyama Islands, Okinawa, 28 August 2017; KPM-NI 18687, 41.7 mm SL, KPM-NI 18688, 45.3 mm SL, KPM-NI 18689, 45.5 mm SL, KPM-NI 18690, 43.4 mm SL, KPM-NI 18691, 42.6 mm SL, KPM-NI 18692, 43.0 mm SL, KPM-NI 18693, 43.9 mm SL, KPM-NI 18694, 42.0 mm SL, KPM-NI 18696, 42.0 mm SL, same data as the holotype; URM-P 4726, 2 specimens, 31.0–33.0 mm SL, Sotobanari-jima island, Yaeyama Islands, Okinawa, 11 September 1982; URM-P 6668, 5, 43.3–47.2 mm SL, Amuro-jima island, Kerama Islands, Okinawa, 8 June 1978; URM-P 6669, 4, 41.9–49.6 mm SL, Kerama Islands, Okinawa, 5 June 1977. MARSHALL ISLANDS: BPBM 18426, 3, 32.0–35.6 mm SL, Parry Island, Enewetak Atoll, 5 m, J. Randall, 11 July 1975; BPBM 29161, 4, 23.1–37.7 mm SL, Japtan Island, Enewetak Atoll, 5–6 m, J. Randall and S. Johnson, 13 October 1982. INDONESIA: BPBM 32234, 3, 25.4–37.6 mm SL, Flores, 08°37′46″S, 122°16′35″E, 32 m, R. Kuiter, 20 September 1987; KAUM–I. 116854, 34.6 mm SL, KAUM–I. 116855, 34.6 mm SL, KAUM–I. 116856, 36.6 mm SL, KAUM–I. 116921, 36.9 mm SL, KAUM–I. 116922, 37.7 mm SL, WAM P.29713-024, 9, 36.0–39.8 mm SL, Flores, 08°29′S, 122°25′E, 5–10 m, G. Allen, 26 April 1988; WAM P. 30513-002, 2, 33.6–35.1 mm SL, Flores, 08°36′S, 122°17′E, 14–20 m, R. Kuiter, 15 May 1992. PAPUA NEW GUINEA: AMS I. 17086-012, 33.3 mm SL, off Madang Harbour, 06°49′S, 145°50′E, B. Collette et al.; BPBM 32517, 7, 22.4–32.4 mm SL, Pig Island, 13 m, J. Randall, 5 November 1987. SOLOMON ISLANDS: BPBM 15623, 9, 26.0–34.1 mm SL, Alite Reef, Malatia Island, 3 m, J. Randall and G. Allen, 25 July 1973. AUSTRALIA: AMS I. 27786-002, 40.0 mm SL, One Tree Island, Queensland, 24°30′S, 152°06′E, T. Trnski; KAUM–I. 2425, 36.0 mm SL; WAM P. 27482-008, 39.9 mm SL, Great Barrier Reef, Queensland, 15°50′S, 145°50′E, 10–23 m, G. Allen, 11 July 1981; WAM P. 29060-011, 3, 34.8–36.2 mm SL, Cartier Island, Western Australia, 12°31′S, 123°23′E, 23–26 m, G. Allen, 20 September 1986. FIJI: CAS 219878, 39.6 mm SL, Kia Island, 27 m, M. David et al.; CAS 223927, 7, 32.9–38.9 mm SL, Kia Island, K. Tang, 29 March 2002.
Diagnosis. A species of Rhabdamia with the following combination of characters: dorsal-fin rays VI-I, 9; anal-fin rays II, 12; pectoral-fin rays 12 or 13 (usually 13); developed gill rakers 26–31 (modally 28–29); total gill rakers (including rudiments) 27–31 (28); body depth at second dorsal-fin origin 25.5–33.1% of SL (mean 30.2%); a black stripe from jaw tips to anterior margin of orbit; black pigmentation lining margin of caudal-fin rays; a subdermal black blotch or spot on lower caudal peduncle; reddish-brown to blackish blotches absent from opercle and anterior portion of body.
Description. Meristics and morphometrics of the type specimens are shown in Table 1. Data for the holotype are presented first, followed by paratype data (if different) in parentheses. Vertebrae 10 + 14. Formula for supraneural bones, anterior neural spines and anterior dorsal pterygiophores 0/0/1/1+1/1/1+1/.
Body oblong, moderately deep, compressed, deepest at second dorsal-fin origin. Dorsal and ventral profiles of head and body convex. Caudal peduncle moderately deep. Head large, compressed. Eye large, round, orbit diameter 3.2 (2.8–3.9) in HL. Mouth moderate in size, oblique, forming angle of ca. 40 degrees to horizontal axis of body. Posterior margin of maxilla slightly concave, extending beyond vertical through anterior margin of eye; supramaxilla absent. Upper-jaw length 2.9 (2.4–2.9) in HL. Lower jaw projecting, its thickened tip fitting into median indentation in upper jaw. No enlarged caniniform teeth on jaws. Villiform teeth in 1–3 rows on both jaws; several tiny teeth on vomer and palatines; teeth absent on ectopterygoid. Anterior nostril with short oval tube, uppermost margin of opening level with dorsal margin of pupil. Posterior nostril oval, opening vertically without rim, uppermost margin below level of dorsal margin of orbit. Gill rakers slender, moderately long. Gill filaments shorter than gill rakers. Single, flat, obtuse opercular spine. Preopercular and opercular margins smooth.
Body scales cycloid, thin, deciduous. Lateral line well developed, more or less parallel with dorsal contour of body, ending a little beyond caudal-fin base (three pored scales posterior to hypural, last scale pointed). No scales on dorsal and anal fins, but a low sheath along bases of second dorsal and anal fins; small scales basally on caudal fin; mid-ventral scales on bases of pelvic fins, paired fins otherwise lacking scales.
First dorsal-fin origin posterior to vertical through pectoral-fin base; second spine of first dorsal fin longest, its length 3.0 (2.4–3.8) in HL. Posterior end of first dorsal-fin base anterior to vertical through posterior tip of depressed pelvic fin. Second dorsal-fin origin anterior to vertical through anal-fin origin; second dorsal fin with concave dorsal margin; length of second dorsal fin spine 3.2 (2.4–3.8) in HL; first soft ray of second dorsal fin longest, its length 1.8 (1.5–2.0) in HL; all second dorsal-fin rays branched. Posterior end of second dorsal-fin base posterior to vertical through base of ninth (or eighth) anal-fin soft ray. Anal-fin origin below base of second soft ray of second dorsal fin; first anal-fin spine short, its length 16.0 (8.7–26.8) in HL; second anal-fin spine long, its length 4.8 (2.9–5.2) in HL; first soft ray of anal fin long, its length 2.2 (1.7–2.6) in HL; all anal-fin rays branched. Pelvic-fin origin anterior to vertical through origin of first dorsal fin; pelvic-fin spine damaged (length 2.7–3.7 in HL). Posterior tip of depressed fin reaching to vertical through sixth spine base of first dorsal fin. Pectoral fin long, its length 1.2 (1.1–1.4) in HL, posterior tip extending beyond vertical through base of third soft ray of second dorsal fin; pectoral-fin base narrow, its width 7.0 (4.7–8.4) in HL. Caudal fin forked, length of upper lobe 1.3 (1.2–1.6) in HL. Anus midway between origins of pelvic and anal fins.
Color when fresh. Body semi-translucent whitish with silvery abdomen, black pigmentation scattered on upper part of body from below first dorsal-fin base to anterior to caudal peduncle. A subdermal black blotch or spot on lower caudal peduncle. Snout and tip of lower jaw reddish-yellow with black pigmentation. Lower part of opercle and anterior part of pectoral-fin base usually without black pigmentation. Posteroventral margin of orbit usually without black pigmentation. First dorsal fin translucent red with black pigmentation. Second dorsal fin translucent red, anterior part yellowish, ray tips with black pigmentation. Pectoral and pelvic fins translucent. Anal fin translucent red, base with black pigmentation. Caudal fin translucent red, lobes yellowish with black pigmentation (sometimes a distinct blotch) on tips.
Color in alcohol. Head and trunk pale yellow. Snout and tip of jaws, behind eye, posteroventral margin of orbit and lower part of opercle with black pigmentation. Black pigmentation scattered on upper part of body from below first dorsal-fin base to just before caudal peduncle. A subdermal black blotch or spot on lower caudal peduncle. First dorsal and caudal fins blackish; other fins translucent.
Distribution. Currently known from the western Pacific Ocean, including Japan, the Marshall Islands, Indonesia, Papua New Guinea, the Solomon Islands, Australia and Fiji (Fig. 4). Collection data include capture depths of 3–32 m.
Etymology. The specific name “novaluna” is derived from Latin meaning “new moon”, in reference to the subdermal black blotch or spot on the lower caudal peduncle.
Remarks. Mabuchi (2001) regarded Rhabdamia gracilis, as commonly recognized, to include two distinct species, one with a black spot on the lower part of the caudal-fin base and the other lacking the spot, identifying the latter as true R. gracilis. On the other hand, Yoshida et al. (2010: fig. 40A, B) treated both species as conspecific, underwater observations including both in a single school and frequently paired with each other at Yaku-shima island, Japan. Subsequently, Yoshida and Motomura (2013) noted considerable size variation in the black spot on the lower part of the caudal-fin base (the spot sometimes being absent on underwater photographs). Reassessment of a pair of individuals with and without a black spot (Yoshida et al. 2010: fig. 40A) revealed that the upper individual figured (appearing to lack a spot) in fact probably had a very small black spot (Yoshida and Motomura 2013). Recently, Yoshida and Motomura (2018) regarded the species lacking a black spot on the lower part of the caudal-fin base as R. gracilis, the other (with a spot) being an undescribed species (described herein as Rhabdamia novaluna).
Six nominal species are included in the genus Rhabdamia (see Yoshida and Motomura 2018). Rhabdamia novaluna sp. nov. is most similar to R. gracilis (Bleeker 1856) [junior synonyms Apogonichthys nudus Regan 1905, Apogonichthys mentalis Evermann and Seale 1907 and Rhabdamia clupeiformis Weber 1909 (see below)] in sharing VI-I, 9 dorsal-fin rays, II, 12 anal-fin rays, 13 pectoral-fin rays and the absence of reddish-brown to blackish blotches on the opercle and anterior portion of the body. However, it is distinguished from R. gracilis by having a large number of developed gill rakers [26–31 (mode 28–29) vs. 22–28 (24) in R. gracilis; Table 3] and total gill rakers (including rudiments) [27–31 (28) vs. 23–28 (25); Table 4], and greater body depth at the second dorsal-fin origin [25.5–33.1% of SL (mean 30.2%) vs. 19.2–30.6% (25.7%); Tables 1–2]. The coloration of R. novaluna sp. nov. also differs from that of R. gracilis, the former having a subdermal black blotch or spot on the lower caudal peduncle (vs. absent).
The new species is easily distinguished from Rhabdamia nigrimentum (Smith 1961) by having 6 first dorsal-fin spines (vs. 7 in R. nigrimentum), 9 second dorsal-fin rays (vs. 11), 12 or 13 pectoral-fin rays (vs. 16), and large numbers of anal-fin rays (12 vs. 11) and total gill rakers [developed gill rakers 26–31 (mode 28–29) vs. 20; total gill rakers (including rudiments) 27–31 (28) vs. 22] (Smith 1961; this study). It differs from Rhabdamia spilota Allen and Kuiter 1994 in lacking blotches on the opercle and anterior portion of the body (vs. 3–6 reddish-brown to blackish blotches in the latter) and has fewer total gill rakers [developed gill rakers and total gill rakers (including rudiments) 27–33 (30–31) and 27–33 (30), respectively, in R. spilota] (Yoshida and Motomura 2018; this study).
Synonymies ofR. gracilis. Rhabdamia gracilis was originally described as Apogonichthys gracilis by Bleeker (1856), based on 39 specimens from Ternate Island, Moluccas, Indonesia. The species was recently redescribed by Yoshida and Motomura (2018) on the basis of 98 specimens, including the type series, and a lectotype designated (RMNH.PISC. 5609, 30.7 mm SL; Fig. 5a).

Primary type specimens identified as Rhabdamia gracilis. a lectotype of Rhabdamia gracilis (RMNH.PISC. 5609, 30.7 mm SL, Indonesia); b syntype of Apogonichthys nudus (BMNH 1904.5.25.104–112, 42.6 mm SL, Indonesia); c holotype of Apogonichthys mentalis (USNM 55905, 52.2 mm SL, Philippines); d syntype of Rhabdamia clupeiformis (RMNH.PISC. 10026, 36.4 mm SL, Pakistan)
Apogonichthys nudus, originally described by Regan (1905) from nine specimens (BMNH 1904.5.25.104–112; Fig. 5b) from Karachi, Pakistan, is regarded here as a junior synonym of R. gracilis (Tables 2–4). However, 10 specimens are currently assigned the above BMNH registration number, being contained in a single jar. Although four of those specimens have been badly damaged and could not be measured due to lacking the posterior portion of the body, the remaining specimens (37.0–43.4 mm SL) were within the range of the syntypes given by Regan (1905), viz. “Length, to base of caudal, 44 mm”. Regan (1905) also described A. nudus as having “A blackish line along the middle of the side from the operculum to below the end of the soft dorsal”. One specimen (one of those damaged) lacked such a stripe and is therefore unlikely to have been a syntype.
Apogonichthys mentalis, originally described from two specimens from Bacon, Sorsogon, Philippines [holotype USNM 55905, 52.2 mm SL (Fig. 5c); paratype CAS-SU 120002, 47.8 mm SL], was also found upon examination to be a junior synonym of R. gracilis (Tables 2–4).
Of the 14 specimens from Lamakera (Solor Island), Lirung (Salibabu Island) and Saleyer, Indonesia, which formed the basis of Rhabdamia clupeiformis, only a single specimen (RMNH.PISC. 10026, 36.4 mm SL) (Fig. 5d) was available for examination, the others (ZMA.PISC. 112204, 112205, and 112207) being apparently lost (R. de Ruiter, pers. comm.). Examination of the former with the lectotype of R. gracilis, and a large number of non-type specimens considered here as R. gracilis (Table 2) showed them to share identical characters. Accordingly, R. clupeiformis is regarded here as a junior synonym of R. gracilis.
Comparative material examined.Rhabdamia gracilis and Rhabdamia spilota, as listed in Yoshida and Motomura (2018), with the following addition: R. gracilis —36 specimens, 30.6–56.1 mm SL. SOUTH AFRICA: SAIAB 202984, 7 specimens, 38.3–44.5 mm SL, Stringer Reef, Sodwana Bay, Kwazulu Natal, 27°31.784′S, 32°40.969′E, D. King and N. Ayliffe, 11 May 2016. MOZAMBIQUE: SAIAB 18447, 2, 30.6–31.6 mm SL, Das Rolas Island, J. L. B. Smith and M. Smith, 11August 1951; SAIAB 18449, 42.6 mm SL, Pinda Island, J. L. B. Smith and M. Smith, 18 July 1950; SAIAB 18452, 38.5 mm SL, Pinda Island, J. L. B. Smith and M. Smith, 18 September 1956; SAIAB 82229, 8, 52.7–56.1 mm SL, 20°05′24″S, 36°07′54″E, 66–68 m depth, P. Heemstra and E. Heemstra, 21 October 2007; SAM 34732, 8, 42.7–52.7 mm SL, 20°26.0′S, 35°53.0′E, bottom trawl, 54 m depth, R/V Algoa, 18 June 1994. MAURITIUS: SAIAB 84066, 42.6 mm SL, Mascarene Plateau, Mascarene Islands, 17°15′19″S, 59°19′23″E, D. Tweddle and O. Alvheim, 16 October 2008. SEYCHELLES: SAIAB 18448, 34.0 mm SL, Mahé, J. L. B. Smith and M. Smith, 1 October 1954; SAIAB 18450, 37.2 mm SL, Aldabra Island, J. L. B. Smith and M. Smith, 1 November 1954. MYANMAR: SAIAB 203823, 43.8 mm SL, 4 April 2015. AUSTRALIA: WAM P. 29060-009, 36.7 mm SL, Cartier Island, Western Australia, 12°31′S, 123°23′E, 23–26 m, G. Allen, 20 September 1986. PHILIPPINES: KAUM–I. 63065, 44.6 mm SL, off Iloilo, Panay Island, 10°41′N, 122°35′E, 1 August 2014. TAIWAN: NMMB-P 26024, 50.7 mm SL, NMMB-P 25990, 2, 48.0–54.6 mm SL, off Dong-gang, Pingtung, 15 April 2017. Type specimens examined included: BMNH 1904.5.25.104–112, 10 syntypes of Apogonichthys nudus, 37.0–43.4 mm SL, Karachi, Pakistan; USNM 55905, holotype of Apogonichthys mentalis, 55.2 mm SL, Bacon, Sorsogon, Philippines; CAS-SU 120002, paratype of A. mentalis, 47.8 mm SL, Bacon, Sorsogon, Philippines, C. J. Pierson; RMNH.PISC. 10026, syntype of Rhabdamia clupeiformis, 36.4 mm SL, Lamakera, Solor, Indonesia; SAIAB (RUSI) 359, holotype of Bentuviaichthys nigrimentum, 50.6 mm SL, off Eritrea.
References
Ahlstrom EH, Butler JL, Sumida BY (1976) Pelagic stromateoid fishes (Pisces, Perciformes) of the eastern Pacific: kinds, distributions, and early life histories and observations on five of these from the northwest Atlantic. Bull Mar Sci 26:285–402
Allen GR, Erdmann MV (2012) Reef fishes of the East Indies. Vols 1–3. Tropical Reef Research, Perth
Allen GR, Kuiter RH (1994) Description of two new species of cardinalfishes (Apogonidae) from Malaysia and Indonesia. Rev Fr Aquariol Herpétol 21:19–23
Bleeker P (1856) Zevende bijdrage tot de kennis der ichthyologische fauna van Ternate. Natuurkundig Tijdschrift voor Nederlandsch Indië, 10:357–386
Evermann BW, Seale A (1907) Fishes of the Philippine Islands. Bull Bur Fish 26:49–110
Fraser TH (1972) Comparative osteology of the shallow water cardinal fishes [Perciformes: Apogonidae] with reference to the systematics and evolution of the family. Ichthyol Bull JLB Smith Inst Ichthyol 34:i–v + 1–105
Hayashi M (1993) Apogonidae. In: Nakabo T (ed) Fishes of Japan with pictorial keys to the species. Tokai University Press, Tokyo, pp 655–681, 1316–1317
Hayashi M (1997) Apogonidae. In: Okamura O, Amaoka K (eds.) Sea fishes of Japan. Yama-kei, Tokyo, pp 288–307
Hayashi M, Kishimoto H (1983) Fish fauna of Iriomote-Island, Ryukyu Island III. Apogonidae (Apogoninae). Sci Rep Yokosuka City Mus 31:15–46
Ida H, Moyer JT (1974) Apogonid fishes of Miyake-jima and Ishigaki-jima, Japan, with description of a new species. Jpn J Ichthyol 21:113–128
Mabuchi K (2001) Rhabdamia gracilis. In: Nakabo T, Machida Y, Yamaoka K, Nishida K (eds) Fishes of the Kuroshio Current, Japan. Osaka Aquarium Kaiyukan, Osaka, p 185
Mabuchi K, Fraser TH, Song H, Azuma Y, Nishida M (2014) Revision of the systematics of the cardinalfishes (Percomorpha: Apogonidae) based on molecular analyses and comparative reevaluation of morphological characters. Zootaxa 3846:151–203
Masuda H, Kobayashi Y (1996) Grand atlas of fish life modes. Color variation in Japanese fish. Tokai University Press, Tokyo
Motomura H, Ishikawa S (eds) (2013) Fish collection building and procedures manual. English edition. The Kagoshima University Museum, Kagoshima and the Research Institute for Humanity and Nature, Kyoto (Available at http://www.museum.kagoshima-u.ac.jp/staff/motomura/dl_en.html)
Randall JE (2005) Reef and shore fishes of the South Pacific. New Caledonia to Tahiti and Pitcairn Islands. University of Hawaiʻi Press, Honolulu
Regan CT (1905) On fishes from the Persian Gulf, the Sea of Oman, and Karachi, collected by Mr. F. W. Townsend. J Bombay Nat. Hist. Soc. 16:318–333, pls A–C
Sabaj MH (2016) Standard symbolic codes for institutional resource collections in herpetology and ichthyology: an online reference. Ver 6.5. American Society of Ichthyologists and Herpetologists, Washington, DC. http://www.asih.org/. Accessed 16 February 2018
Senou H, Kodato H, Nomura T, Yunokawa, K (2006) Coastal fishes of Ie-jima Island, the Ryukyu Islands, Okinawa, Japan. Bull Kanagawa Prefect Mus (Nat Sci) 35:67–92
Senou H, Kobayashi Y, Kobayashi N (2007) Coastal fishes of the Miyako Group, the Ryukyu Island, Japan. Bull Kanagawa Prefect Mus (Nat Sci) 36:47–74
Smith JLB (1961) Fishes of the family Apogonidae of the western Indian Ocean and the Red Sea. Ichthyol Bull JLB Smith Inst Ichthyol (22):373–418
Watai M, Miyazaki Y, Murase A, Senou H (2009) Fish fauna of Tokashiku Bay, Tokashiki Island, the Kerama Islands, Okinawa Prefecture. Bull Kanagawa Prefect Mus (Nat Sci) 38:119–132
Weber M (1909) Diagnosen neuer fische der Siboga-Expedition. Notes Leyden Mus 31:143–169
Yoshida T, Motomura H (2013) Rhabdamia gracilis. In: Motomura H, Dewa S, Furuta K, Matsuura K (eds) Fishes of Iou-jima and Take-shima islands, Mishima, Kagoshima, Japan. The Kagoshima University Museum, Kagoshima, and the National Museum of Nature and Science, Tsukuba, pp 136–138
Yoshida T, Motomura H (2016) A new cardinalfish, Verulux solmaculata (Perciformes: Apogonidae), from Papua New Guinea and Australia. Ichthyol Res https://doi.org/10.1007/s10228-016-0539-2 (also appeared in Ichthyol Res 64:64–70)
Yoshida T, Motomura H (2018) Redescription of the Indo-West Pacific cardinalfishes (Perciformes: Apogonidae) Rhabdamia spilota Allen & Kuiter 1994 and R. gracilis (Bleeker 1856). Zootaxa 4377:178–190
Yoshida T, Harazaki S, Motomura H (2010) Apogonid fishes (Teleostei: Perciformes) of Yaku-shima Island, Kagoshima Prefecture, southern Japan. In: Motomura H, Matsuura K (eds) Fishes of Yaku-shima Island – A World Heritage island in the Osumi Group, Kagoshima Prefecture, southern Japan. National Museum of Nature and Science, Tokyo, pp 27–64
Yoshigou H, Nakamura S (2002) Catalogue of fish specimens preserved in Hiwa Museum for Natural History (II). Hiwa Mus Nat His Mtl Rep (3):85–136, pl 1
Yoshigou H, Ichikawa M, Nakamura S (2005) Catalogue of fish specimens preserved in Hiwa Museum for Natural History (IV). Hiwa Mus Nat His Mtl Rep (5):1–51, pl 1
Acknowledgements
We are deeply grateful to M. McGrouther, A. Hay and S. Reader (AMS), M. Sabaj (ANSP), O. Crimmen and J. Maclaine (BMNH), A. Suzumoto (BPBM), T. Iwamoto, D. Catania and M. Hong (CAS), H. Senou (KPM), K. Koeda (NMMB), R. de Ruiter (RMNH), T. Yoshino (Okinawa Churashima Foundation), R. Bills and O. Gon (SAIAB), A. Bosman (SAM), J. Williams (USNM), and S. Morrison, G. Moore and M. Allen (WAM) for opportunities to examine specimens of Rhabdamia. We also thank Y. Haraguchi and other volunteers and students of KAUM for curatorial assistance, G. Moore (WAM) for the donation of WAM specimens to KAUM, and G. Hardy (Ngunguru, New Zealand) for reading the manuscript and providing help with English. This study was supported in part by the Sasakawa Scientific Research Grant from the Japan Science Society (29-747), Grants-in-Aid for Scientific, JSPS KAKENHI Grant Numbers JP26241027, JP24370041, JP23580259 and JP26450265; the JSPS Core-to-Core Program: B Asia–Africa Science Platforms; the “Biological Properties of Biodiversity Hotspots in Japan” project of the National Museum of Nature and Science, Tsukuba, Japan; “Establishment of Research and Education Network on Biodiversity and Its Conservation in the Satsunan Islands” project of Kagoshima University adopted by the Ministry of Education, Culture, Sports, Science and Technology, Japan; and the “Island Research” project of Kagoshima University. This study was conducted under a Memorandum of Agreement for joint research made by and among the Department of Agriculture of the Republic of the Philippines (DA), the University of the Philippines Visayas (UPV), the Kagoshima University Museum, the Research Institute for Humanity and Nature, and Tokai University, facilitated by S. L. Sanchez [Bureau of Fisheries and Aquatic Resources (BFAR), DA]. P. J. Alcala (DA) provided a Prior Informed Consent Certificate, and I. P. Cabacaba and S. M. S. Nolasco (BFAR, DA) provided a fish specimen Export Certificate (No. 2016-39812). We thank the staff of the Office of the Vice-Chancellor for Research and Extension, UPV, and UPV Museum of Natural Sciences, College of Fisheries, UPV, including R. P. Babaran, S. S. Garibay, U. B. Alama, V. G. Urbina, L. H. Mooc, C. J. N. Rubido, E. P. Abunal, A. M. T. Guzman, R. S. Cruz, A. C. Gaje and R. F. M. Traifalgar, and graduate students of the College of Fisheries, UPV, for their support of this research collaboration.
Author information
Authors and Affiliations
Corresponding author
Additional information
This article was registered in the Official Register of Zoological Nomenclature (ZooBank) as A2613F51-4512-4EC1-A4DA-EB778F892063.
This article was published as an Online First article on the online publication date shown on this page. The article should be cited by using the doi number.
About this article

Cite this article
Yoshida, T., Mabuchi, K. & Motomura, H. Rhabdamia novaluna, a new species of cardinalfish (Perciformes: Apogonidae) from the western Pacific Ocean, with comments on the synonymy of Rhabdamia gracilis. Ichthyol Res 66, 129–139 (2019). https://doi.org/10.1007/s10228-018-0664-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10228-018-0664-1