
Abstract
Animals may build refuges to avoid predation, to communicate, to mate, and to protect against extreme temperatures, among other factors. Allocosa senex is a wolf spider that constructs burrows in the coastal sand dunes of South America. The distribution of A. senex includes beaches with different granulometry, which could affect burrow characteristics and energetic costs of excavation. Males are the digging sex and their burrows are mating and oviposition sites. We aimed to test substrate preference of males of A. senex simultaneously exposed to coarse- (CS) and fine-grain sand (FS) (n = 30) and study digging behavior in both substrates. Regardless of their substrate of origin, males significantly preferred CS for digging (males from CS habitat, 23/30, P = 0.001; males from FS habitat, 20/30, P = 0.02). Males from the FS habitat showed higher weight and better body condition. Digging behavior showed similar patterns in both types of substrate but resting duration was longer when males dug in CS. Although males of A. senex can dig in both substrates, this study shows their preference for digging in CS, a substrate that is possibly less energetically demanding for that behavior.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Constructions such as shelters, burrows, or nests may allow their inhabitants to avoid predators, attract mates, and protect themselves and their progeny from extreme weather conditions (Hansell 2005). Construction processes imply considerable energetic costs that often involve a trade-off with other physical or behavioral traits (Okano et al. 2010). The quality of the materials used in these constructions may affect construction behavior and the final structure, therefore influencing the future success of the builder. Several coastal animal species transform their extreme and/or unpredictable microhabitats by constructing structures that generate more stable conditions for protection, mating, or brooding offspring. These constructions can vary according to the type of sediment. Austrohelice crassa (Dana 1851), a crab inhabitant of intertidal sediments of northern New Zealand, constructs shelters that vary in height according to the topography and geochemistry of the sediment (Morrisey et al. 1999). Another interesting example is that of the sand crab Emerita analoga Stimpson, 1857 that constructs burrows in the swash and shallow subtidal zones of Californian beaches. In this species burrowing time is dependent not only on sediment grain sizes but also on body size, parasite load and reproductive condition of the individuals (Dugan et al. 2000; Kolluru et al. 2011).
Several arachnid species construct burrows where they reside temporarily or permanently (Gwynne and Watkiss 1975; Henschel and Lubin 1992; Henschel 1995; Tso et al. 2003; Pérez-Miles et al. 2005; Suter et al. 2011; Hembree 2013). Spider burrows are frequently lined with silk, several threads of which are sometimes interlocked in a complex manner (Foelix et al. 2017). In scorpions and solpugids, burrow digging consists of extracting and tamping the substrate, but without using any connecting or cementing substances (Polis 1990; Warburg and Polis 1990; Punzo 1998). Spider burrows can be relatively simple modifications of natural fissures in the ground, or complex deep constructions with one or more openings, or with trapdoors (Foelix 2011). Furthermore, burrows can externally show turrets, aboveground extensions of the construction composed of silk, particles of soil or sand, plant debris or pebbles. The possible functions of these external constructions are detection of potential prey outside the burrow, protection from predators and prevention of flooding inside the burrow (Shook 1978; Alderweireldt and Jocque 1991; Henschel 1995; Henschel and Lubin 1997; Logunov 2011). Retreats also buffer temperature and humidity variations, providing a safe refuge in which to hide, mate, oviposit and/or brood progeny (Aisenberg et al. 2011a; Suter et al. 2011). Soil characteristics in the digging site will affect the energetic costs of the excavation activity, the dimensions of the resultant structure and, consequently, its effectiveness (Henschel and Lubin 1997; Suter et al. 2011).
The nocturnal wolf spider Allocosa senex (Mello-Leitão, 1945), recent revalidated with the junior synonym Allocosa brasiliensis (Petrunkevitch, 1910) (Simó et al. 2017), inhabits the sandy coasts of Argentina, Brazil and Uruguay (Capocasale 1990). Individuals build silk-lined burrows in which they remain during the day and in winter, becoming active during summer nights (Costa 1995). Male burrows are longer than those of females (under laboratory conditions, 10 cm and 3.5 cm length, respectively) (Aisenberg et al. 2007). This species shows a reversal in typical sex roles expected in spiders, as well as in sexual size dimorphism: males are sedentary and larger than females, remaining within their burrows for long periods, while females are the mobile sex which looks for males and initiates courtship (Aisenberg et al. 2007; Aisenberg 2014). Copulation occurs inside the male burrow and after it ends the male exits and the female seals the burrow entrance from inside. The female lays the egg sac inside the male burrow and leaves it when it is time for the progeny to disperse (Aisenberg et al. 2007). Both sexes are selective at the time of mating: females prefer to mate with males who own the longest burrows, whilst males choose virgins females in good body condition (Aisenberg et al. 2007, 2011b). According to recent studies, males of A. senex modify their building behavior and elongate their burrows according to the availability of females (Carballo et al. 2017). Summarizing, the male burrow is a refuge, mating site and breeding nest, so high selective pressures are expected to determine male digging in this species.
A. senex has a wide geographical distribution (Capocasale 1990; Simó et al. 2017) that includes beaches with drastic differences in substrate granulometry and composition. However, information about the potential costs of digging in different substrates and how this might affect male fitness or female choice (strong sexual selection on burrow length) in A. senex is unknown. Furthermore, plasticity of male burrowing behavior in different substrates could be crucial for the species’ survival under climate change and human disturbances or habitat degradation. The objectives of this study were to test substrate preference in males of A. senex simultaneously exposed to coarse sand (CS) and fine sand (FS) and compare burrow building behavior between the two substrates. We expected that males would prefer coarse substrate due to the lower number of silk layers needed to maintain a stable burrow in this substrate. We also expected to find variations in burrow digging according to the substrate.
Materials and methods
Study site, field captures and breeding
We performed granulometry analyses of sand samples from the beach at San José de Carrasco, Canelones, Uruguay (34°51′06.06″S, 55°58′46.71″W) and Paso del Molino, Lavalleja, Uruguay (34º16′40.10″S, 55º14′00.80″W), at the Departamento de Evolución de Cuencas, Facultad de Ciencias, Universidad de la República, Montevideo, Uruguay. The sand was sifted to measure the proportion of grains of each size in the sediment sample. We confirmed that San José de Carrasco shows a higher percentage of FS compared to Paso del Molino beach which shows a predominance of CS (Table 1).
We collected 30 adult males of A. senex at each beach between January 2013 and March 2015 (n = 60). We captured individuals overnight using a miner flashlight. We placed the specimens individually in Petri dishes of 9.5 cm in diameter and 1.5 cm high, with sand from their original location as substrate and water provided ad libitum using a moist cotton wick. We maintained spiders under natural light cycles. The temperature during maintenance was 26.25 °C (± 1.5 SD) and humidity averaged 66.25% (± 5.41 SD). We fed spiders three times a week with one larva of Tenebrio molitor Linnaeus 1758 (Coleoptera, Tenebrionidae).
Substrate choice tests
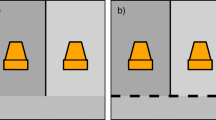
We used males in the trials 7 days after their capture for their acclimation to laboratory conditions. We placed each male (n = 30 males from each location) in a glass terrarium of 30-cm length, 16-cm width and 20-cm height, with sand as substrate and a layer of 5 cm of moistened sand at the base (Aisenberg et al. 2007, 2011b). Each male was selected only once for observation. We placed FS and CS from the original sampling localities in one half each of the terrarium, without barriers between the two. The animals were introduced into the terrarium where the two types of substrates met. Once inside, the animals inspected the substrates and walked around the terrarium until finally selecting the place where they started to build their burrows (Fig. 1). The substrates used in the tests originated from the original locations of the spiders. The position of the CS and FS in the terrarium was alternated between the tests (right and left side of the terrarium) to reduce the potential effects of the method. Forty-eight hours after placing each male in the terrarium, we checked for burrow occurrence and, if present, measured the dimensions (burrow length and diameter). A. senex males construct their burrows against terrarium walls (Aisenberg et al. 2007, 2011b), which allowed us to measure burrow dimensions through the glass wall. We measured carapace width [a characteristic considered representative of body size in spiders (Marshall and Gittleman 1994; Eberhard et al. 1998)], opisthosomal width, and weighed each individual immediately before the trials. We considered the index opisthosomal width/carapace with as representative of body condition, following Moya-Laraño et al. (2003).

Experimental design for substrate selectivity. Right Fine sand (FS) from San José de Carrasco, Canelones, Uruguay; left coarse sand (CS) from Paso del Molino, Lavalleja, Uruguay. The positions of FS and CS were exchanged between the trials
Burrow-digging behavior
As fine-scale data on male burrow digging of A. senex from the FS substrate at San José de Carrasco, Canelones, Uruguay, were available (Aisenberg and Peretti 2011), we decided to record male burrow digging only for CS and then compare the results for both types of substrate.
To record male burrow digging in CS, we collected another 20 adult males of A. senex between December 2014 and January 2015 at Paso del Molino (CS) (34º16′40.10″S, 55°14′00.80″W) Lavalleja, Uruguay. Feeding and maintenance conditions were similar to those described in the first section of Materials and methods. During burrow digging observations the temperature averaged 25.90 °C (± 2.80 SD) and humidity was 64.00% (± 3.10 SD). Each male was placed individually in a glass terrarium (30 cm × 16 cm × 20 cm), with CS from its original location as substrate. We placed 15 cm of dry sand and 5 cm of moistened sand on the base of the terrarium following the protocol of Aisenberg and Peretti (2011). We performed the observations between 2 and 5 p.m. to examine rapid burrow construction as a response to daylight, again in agreement with Aisenberg and Peretti (2011).
We recorded building behavior by ten males of A. senex during 1 h after the males started removing the sand with their chelicerae, forelegs and palps. If digging behavior did not begin in the first 30 min, we finished the observation. After 1 h of observation, we measured burrow dimensions (length and diameter). We recorded all the observations with a Sony DCR-SR85 video camera. We analyzed the number of occurrences and duration of digging behavioral patterns with J Watcher software (Blumstein et al. 2000).
Statistics
We analyzed the results using NCSS (copyright 2000 Jerry Hintze) and Past Paleontological Statistics, version 1.18 (Hammer et al. 2003) statistical packages. We compared the frequencies of burrow digging in each substrate with a binomial test. We compared burrow dimensions (length and width) between the burrows constructed in each substrate and body characteristics of males from each location. We also compared the number of occurrences and duration of burrowing behaviors in CS with those reported for FS by Aisenberg and Peretti (2011). We checked for normality (Shapiro–Wilk test) and homogeneity of variances (Levene test) of the data, and accordingly applied Student t-tests (parametric test) or Mann–Whitney U-tests (non-parametric test). We deposited voucher specimens from each beach at the arachnological collection of the Facultad de Ciencias, Montevideo, Uruguay.
Results
Substrate selectivity tests
We summarize the results on substrate preferences and burrow dimensions in Figs. 2 and 3, respectively. Males belonging to both the FS and CS groups showed preference for digging their burrows in CS (Fig. 2). In general, we did not find differences in burrow dimensions between males from FS or CS locations that constructed burrows in each substrate. However, males from FS that constructed in CS built longer burrows than CS males that dug burrows in the same substrate (Fig. 3).

Number of males from each location which dug in FS or CS, with the results of their corresponding statistical comparisons. For abbreviations, see Fig. 1

Box plots showing length and width of burrows constructed in FS or CS by males from each location (FS or CS beaches), with the results of their corresponding statistical comparisons. For abbreviations, see Fig. 1
Data on body characteristics of males from each location are summarized in Table 2, with their corresponding statistical comparisons. We did not find differences in body size between FS and CS males. Nevertheless, FS males weighed more and had higher body condition indexes than CS males (Table 2).
Burrow-digging behavior
The average depth of burrows by males in CS during 1 h was 3.76 ± 1.11 cm, with an average width of 1.44 ± 0.11 cm. The burrows were tubular and vertical with a single entry. We distinguished eight behaviors associated with burrow digging: extracting sand, exiting the burrow, entering the burrow, laying silk on burrow walls, releasing silk around entrance, turning, blocking entrance, and resting (Fig. 4). After 1 h of observations, burrow length values were higher in CS compared to previous observations in FS under similar conditions (Aisenberg and Peretti 2011) (FS: 2.45 ± 0.39 cm; t = −3.23, P = 0.006). Also, burrows were wider in CS compared to FS (FS: 1.01 ± 0.02 cm; t = −6.68, P = 0.0001).

Box plots showing the number of occurrences (a) and duration (b) of burrow-digging patterns in FS and CS, with the results of the statistical comparisons. FS values correspond to those described by Aisenberg and Peretti (2011). White Data for males originating from the FS location, gray data for males originating from the CS location. For abbreviations, see Fig. 1
The number of occurrences and duration of burrow-digging behavioral patterns in CS (present study) and FS (Aisenberg and Peretti 2011) are summarized in Fig. 4. The construction of burrows began with the removal of sand to make a hole (hollowing); once inside the hole, the spider began to extract sand using its chelicerae, forelegs and pedipalps. Males repeatedly left the burrow to deposit the sand outside, then returned inside the burrow. They combined sand extraction with silk deposition on the inner walls of the burrow and resting. Three of ten individuals did not close their burrows. When males blocked their burrows, they closed them by depositing silk and sand until a complete plug had been created. When we compared burrow digging in CS with data on FS, general digging patterns were similar, but occurrences and duration of resting behavior were higher in CS compared to FS (Fig. 4).
Discussion
This study shows that males of A. senex, when given the choice, prefer to dig in CS compared to FS, independently of their substrate of origin. Possibly, in this species, the choice of substrate is influenced by characteristics of the sand such as grain size and hardness, among other factors, which could play an important role when burrows are built. A. senex individuals use their palps, forelegs and chelicerae during digging (Aisenberg and Peretti 2011), and possibly males test substrate characteristics with these appendages during the first digging event, when making the initial hole.
Males of A. senex may prefer CS because digging is faster or less energetically demanding in this substrate than in FS; this is in agreement with the creation of longer burrows in CS compared to FS after 1 h of observation. Silk production and deposition have been reported as energetically demanding activities in spiders (Prestwich 1977; Henschel and Lubin 1992). Burrow construction in the sand implies the deposition of several layers of silk to maintain a firm structure, particularly in FS. The deposition of multiple layers of silk during burrow construction has been described for other wolf spiders inhabiting sandy habitats (Gwynne and Watkiss 1975; Henschel 1990; Foelix et al. 2017). In A. senex, burrows of males are their refuges but also the place where mating, oviposition and brooding of the spiderlings occur (Aisenberg et al. 2007; Aisenberg 2014). Furthermore, there is strong female choice for male burrow length. However, in this study we did not find higher frequencies of duration of silk deposition in FS, so hypothesize that mechanical properties of CS, such as grain composition and weight, but not differences in silk deposition behavior, are crucial for determining the observed preference by males of A. senex. Soil hardness, particle size, and sediment density have been reported as important factors determining habitat selection and burrow architecture in other spiders (Main 1982; Punzo and Henderson 1999), scorpions (Polis 1990), and harvestmen (Lamoral 1978). On the other hand, though we did not find differences in the number of occurrences or duration of silk deposition units between the two substrates, other unregistered characteristics such as the type of silk mesh or the thickness of the silk fibers could vary in FS and CS burrows. According to recent studies on A. senex and other burrowing spiders, silk mesh complexity seems to be an essential factor for providing stability to the burrow when digging on sand (Foelix et al. 2017).
The burrows constructed by males of A. senex in CS were tubular with a single entry, similar to those described for this species in FS (Aisenberg et al. 2007; Albín et al. 2015). Though general digging patterns in FS or CS agreed, resting behavior occurred more frequently and for a longer duration in CS. This result, in addition to the longer burrows dug in coarse substrate in a similar period, indicate that digging is faster in CS and that males reach a minimum optimum of burrow length in a shorter time period in this substrate. Moreover, it is remarkable that males from FS constructed longer burrows in CS compared to males that originated from the CS location.
Males from the CS locality showed lower weight and body condition values compared to FS males. Intraspecific differences in body characteristics between localities have been reported for other animal groups (Bansemer et al. 2002; Schäuble 2004), and arachnids such as scorpions (Olivero et al. 2012), spiders (González et al. 2015; Puzin et al. 2014), and particularly for A. senex (Bollatti et al. 2017). Variations in the microhabitats and prey availability in each location could shape body differences, affecting not only morphology, but also physiology and behavior of the individuals (Berger et al. 2012; Hausch et al. 2013). Prey abundance could in turn determine the availability of fat reserves and affect energy expenditure during digging, and consequently resting needs, of males of A. senex. Males from the locality with FS may require better body condition for the more demanding burrowing effort in this substrate.
Though males of A. senex preferred digging in CS, they showed the ability to dig similarly long burrows in both substrates. Phenotypic plasticity allows an organism with a given genotype to change its phenotype in response to changes in the environment (Réale et al. 2003; Nussey et al. 2007), i.e. the ability of individuals to change behavior in situations that may have important ecological and evolutionary implications for them (West-Eberhard 1989). Due to a strong anthropogenic impact, the Uruguayan coast has been modified adversely during the last decades (Costa et al. 2006) affecting the populations of A. senex. Sand dunes there have been drastically reduced, fragmented and modified due to the introduction of exotic vegetation, which has gradually changed the composition, hardness and compaction of the substrate where A. senex constructs its burrows (Jorge et al. 2015). Thus, adaptation to different substrates is of vital importance for the existence of this species in sandy environments in Uruguay.
It will be interesting to determine whether burrow-digging duration and burrow architecture of males of A. senex vary in different substrates under natural conditions. Finally, in future studies we will test the costs associated with burrowing behavior of males of A. senex through immunocompetence tests performed before and after burrow digging, testing the costs associated with oxygen consumption during the process in order to determine if the high male reproductive investment in this trait, and burrow delivery after mating, shape male mate choice in this species.
References
Aisenberg A (2014) Adventurous females and the demanding males: sex role reversal in a Neotropical spider. In: Macedo RH, Machado G (eds) Sexual selection: perspectives and models from the neotropics. Academic Press, California, pp 163–182
Aisenberg A, Peretti AV (2011) Male burrow digging in a sex-role reversed spider inhabiting water-margin environments. Bull Br Arach Soc 15:201–204
Aisenberg A, Viera C, Costa FG (2007) Daring females, devoted males, and reversed sexual size dimorphism in the sand dwelling spider Allocosa brasiliensis (Araneae, Lycosidae). Behav Ecol Sociobiol 62:29–35
Aisenberg A, González M, Laborda A, Postiglioni R, Simó M (2011a) Spatial distribution, burrow depth and temperature: implications for the sexual strategies in two Allocosa wolf spiders. Stud Neotrop Fauna Environ 46:147–152
Aisenberg A, Costa FG, González M (2011b) Male sexual cannibalism in a sand-dwelling wolf spider with sex role reversal. Biol J Linn Soc 103:68–75
Albín A, Simó M, Aisenberg A (2015) Characterization of burrow architecture under natural conditions in the sand-dwelling wolf spider Allocosa brasiliensis. J Nat Hist 14:22–29
Alderweireldt M, Jocque R (1991) A remarkable new genus of wolf spiders from southwestern Spain (Araneae, Lycosidae). Bull Inst R Sci Nat Belg Entomol 61:103–111
Bansemer C, Grutter AS, Poulin R (2002) Geographic variation in the behaviour of the cleaner fish. Ethology 108:353–366
Berger D, Olofsson M, Friberg M, Karlsson B, Wiklund C, Gotthard K (2012) Intraspecific variation in body size and the rate of reproduction in female insects–adaptive allometry or biophysical constraint? J Anim Ecol 81:1244–1258
Blumstein DT, Evans CS, Daniel JC (2000) JWatcher. http://galliform.psy.mq.edu.au/jwatcher/. Accessed 2015 May 16
Bollatti F, Garcia Diaz V, Peretti AV, Aisenberg A (2017) Geographical variation in sexual behavior and body traits in a sex role reversed wolf spider. Sci Nat 104:40
Capocasale RM (1990) Las especies de la subfamilia Hipassinae de América del Sur (Araneae, Lycosidae). J Arachnol 18:131–141
Carballo M, Baldenegro F, Bollatti F, Peretti AV, Aisenberg A (2017) No pain, no gain: male plasticity in burrow digging according to female rejection in a sand-dwelling wolf spider. Behav Proc 140:171–180
Costa FG (1995) Ecología y actividad diaria de las arañas de la arena Allocosa spp. (Araneae, Lycosidae) en Marindia, localidad costera del sur del Uruguay. Rev Bras Biol 55:457–466
Costa FG, Simo M, Aisenberg A (2006) Composición y ecología de la fauna epigea de Marindia (Canelones, Uruguay) con especial énfasis en las arañas: un estudio de dos años con trampas de intercepción. In: Rodríguez-Gallego L, Scarabino F, Conde D, Menafra R (eds) Bases para la conservación y el manejo de la costa uruguaya. Vida Silvestre Uruguay, Montevideo, pp 427–436
Dugan JE, Hubbard DM, Lastra M (2000) Burrowing abilities and swash behavior of three crabs, Emerita analoga Stimpson, Blepharipoda occidentalis Randall, and Lepidopa californica Efford (Anomura, Hippoidea), of exposed sandy beaches. J Exp Mar Biol Ecol 255:229–245
Eberhard WG, Huber BA, Rodrıguez SRL, Briceno RD, Salas L, Rodrıguez V (1998) One size fits all? Relationships between the size and degree of variation in genitalia and other body parts in twenty species of insects and spiders. Evolution 52:415–431
Foelix RF (2011) Biology of spiders, 3rd edn. Oxford University Press, New York
Foelix R, Rechenberg I, Erb B, Albín A, Aisenberg A (2017) Sand transport and burrow construction in sparassid and lycosid spiders. J Arachnol 45:255–264
González M, Peretti AV, Costa FG (2015) Reproductive isolation between two populations of Aglaoctenus lagotis, a funnel-web wolf spider. Biol J Linn Soc 114:646–658
Gwynne DT, Watkiss J (1975) Burrow blocking behaviour in Geolycosa wrightii (Araneae: lycosidae). Anim Behav 23:953–956
Hammer O, Harper DAT, Ryan PD (2003) Past Palaeontological, version 1.18. Copyright Hammer and Harper. http://folk.uio.no/ohammer/past. Accessed 15 Mar 2015
Hansell MH (2005) Animal architecture. Oxford University Press, New York, p 336
Hausch S, Shurin JB, Matthews B (2013) Variation in body shape across species and populations in a radiation of Diaptomid Copepods. PLoS One 8:e68272
Hembree DI (2013) Neoichnology of the whip scorpion Mastigoproctus giganteus: complex burrows of predatory terrestrial arthropods. Palaios 28:141–162
Henschel JR (1990) The biology of Leucorchestris arenicola (Araneae: Heteropodidae), a burrowing spider of the Namib dunes. In: Seely MK (eds) Namib ecology: 25 years of Namib research. Transvaal museum monograph no. 7. Transvaal Museum, Pretoria, p 115–127
Henschel JR (1995) Tool use by spiders: stone selection and placement by corolla spiders Ariadna (Segestriidae) of the Namib Desert. Ethology 101:87–199
Henschel JR, Lubin YD (1992) Environmental factors affecting the web and activity of a psammophilous spider in the Namib Desert. J Arid Environ 22:173–189
Henschel JR, Lubin YD (1997) A test of habitat selection at two spatial scales in a sit-and-wait predator: a web spider in the Namib Desert dunes. J Anim Ecol 66:401–413
Jorge C, Laborda A, Alves Días M, Aisenberg A, Simó M (2015) Habitat preference and effects of coastal fragmentation in the sand-dwelling spider Allocosa brasiliensis (Lycosidae; Allocosinae) Open. J Anim Sci 5:309–324
Kolluru GR, Green ZS, Vredovoe LK, Kuzma MR, Ramadan SN, Zosky MR (2011) Parasite infection and sand coarseness increase sand crab (Emerita analoga) burrowing time. Behav Processes 88:184–191
Lamoral BH (1978) Soil hardness, an important and limiting factor in burrowing scorpions of the genus Opisthophthalmus C. L. Koch, 1837 (Scorpionidae, scorpionida). Symp Zool Soc Lond 42:171–181
Logunov DV (2011) Sexual size dimorphism in burrowing wolf spiders (Araneae: lycosidae). Proc Zool Inst 315:274–288
Main BY (1982) Adaptations to arid habitats by mygalomorph spiders. In: Barker WR, Greenslade PJM (eds) Evolution of the flora and fauna of Australia). Peacock, Anzaas S, Australian Division, pp 273–283
Marshall SD, Gittleman JL (1994) Clutch size in spiders: is more better? Funct Ecol 8:118–124
Morrisey DJ, DeWitt TH, Roper DS, Williamson RB (1999) Variation in the depth and morphology of burrows of the mud crab Helice crassa among different types of intertidal sediment in New Zealand. Mar Ecol Prog Ser 182:231–242
Moya-Laraño J, Pascual J, Wise DH (2003) Mating patterns in late-maturing female Mediterranean tarantulas may reflect the costs and benefits of sexual cannibalism. Anim Behav 66:469–476
Nussey DH, Wilson AJ, Brommer JE (2007) The evolutionary ecology of individual phenotypic plasticity in wild populations. J Evol Biol 20:831–844
Okano J, Kikuchi E, Sasaki O (2010) The role of particle surface texture on case material selection and silk lining in caddis flies. Behav Ecol 21:826–835
Olivero PA, Mattoni CI, Peretti AV (2012) Morphometry and geographical variation of Bothriurus bonariensis (Scorpiones: bothriuridae). J Arachnol 40:113–122
Pérez-Miles F, Costa FG, Toscano-Gadea C, Mignone A (2005) Ecology and behaviour of the ‘road tarantulas’ Eupalaestrus weijenberghi and Acanthoscurria suina (Araneae, Theraphosidae) from Uruguay. J Nat Hist 39:483–498
Polis GA (1990) The biology of scorpions. Stanford University Press, California, p 587
Prestwich KN (1977) The energetics of web-building in spiders. Comparative Biochem Physiol 57:321–326
Punzo F (1998) The biology of camel-spiders: Arachnida, Solifugae. Kuwer, Dordrecht
Punzo F, Henderson L (1999) Aspects of the natural history and behavioural ecology of the tarantula spider Aphonopelma hentzi (Orthognatha, Theraphosidae). Bull Br Arachnol Soc 11:121–128
Puzin C, Leroy B, Pétillon J (2014) Intra-and inter-specific variation in size and habitus of two sibling spider species (Araneae: lycosidae): taxonomic and biogeographic insights from sampling across Europe. Biol J Linn Soc 113:85–96
Réale D, McAdam AG, Boutin S, Berteaux D (2003) Genetic and plastic responses of a northern mammal to climate change. Proc R Soc Lond Biol 270:591–596
Schäuble CS (2004) Variation in body size and sexual dimorphism across geographical and environmental space in the frogs Limnodynastes tasmaniensis and L. peronii. Biol J Linn Soc 82:39–56
Shook RS (1978) Ecology of the wolf spider, Lycosa carolinensis Walckenaer (Araneae, Lycosidae) in a desert community. J Arachnol 6:53–64
Simó M, Lise AA, Pompozzi G, Laborda A (2017) On the taxonomy of southern South American species of the wolf spider genus Allocosa (Araneae: Lycosidae: Allocosinae). Zootaxa 4216:61–278
Suter RB, Stratton GE, Miller PR (2011) Mechanics and energetics of excavation by burrowing wolf spiders. Geolycosa spp. J Insect Sci 11:22
Tso IM, Haupt J, Zhu MS (2003) The trapdoor spider family Ctenizidae (Arachnida: Araneae) from Taiwan. Raffles Bull Zool 51:25–33
Warburg MR, Polis GA (1990) Behavioral responses, rhythms, and activity patterns. In: Polis GA (ed) The biology of scorpions. Stanford University Press, California, pp 224–246
West-Eberhard MJ (1989) Phenotypic plasticity and the origins of diversity. Annu Rev Ecol Syst 20:249–278
Acknowledgements
We thank L. Bidegaray, F. Bollatti, M. Carballo, F. Cargnelutti, T. Casacuberta, M. Casacuberta, G. De Simone, Á. Laborda, M. González, L. Montes de Oca, and M. Trillo for their help in the fieldwork and breeding of spiders. We are grateful to P. Pintos and R. Postiglioni for their help with the box plots and figures. We thank Petr Dolejš, one anonymous reviewer, and the editor for their suggestions that improved the final version of the manuscript. This research was funded by the National Agency for Research and Innovation under the code POS_NAC_2013_1_555 awarded to A. Albín. A. Aisenberg and M. Simó acknowledge financial support from the Programa Desarrollo de Ciencias Básicas and the Sistema Nacional de Investigadores.
Author information
Authors and Affiliations
Corresponding author
About this article

Cite this article
Albín, A., Bardier, G., Peretti, A.V. et al. A matter of choice: substrate preference by burrow-digging males of a sand-dwelling spider. J Ethol 37, 13–20 (2019). https://doi.org/10.1007/s10164-018-0561-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10164-018-0561-0