
Abstract
Red swamp crayfish is particularly prone to exposure to hypoxia-reoxygenation stress on account of the respiration and rhythmic, light-dependent photosynthetic activity of the algae and aquatic grass. Up to now, the regulation mechanisms of the adverse effects of hypoxia-reoxygenation for this species were still unknown, especially the roles of the antioxidant enzymes in reducing oxidative damage during reoxygenation. To screen for vital genes or pathways upon hypoxic-reoxygenation stress, hepatopancreas gene expression profiles were investigated by using a strategy combining second and third generation sequencing. Five groups of samples, including hypoxia for 1 and 6 h with DO of 1.0 mg/L, reoxygenation for 1 and 12 h with DO of 6.8 mg/L, and the samples under normoxia condition, were used for transcriptome sequencing. Twenty Illumina cDNA libraries were prepared to screen for the differentially expressed genes (DEGs) among the 5 groups of samples. Based on the assembled reference full-length transcriptome, 389 and 533 significantly DEGs were identified in the groups under severe hypoxia treatment for 1 and 6 h, respectively. The top three enriched pathways for these DEGs were “protein processing in endoplasmic reticulum,” “MAPK signaling pathway,” and “endocytosis.” Among these DEGs, hypoxia-inducible factor 1α (Hif-1α) and some Hif-1 downstream genes, such as Ugt-1, Egfr, Igfbp-1, Pk, and Hsp70, were significant differentially expressed when exposed to hypoxia stress. A series of antioxidant enzymes, including two types of superoxide dismutase (Cu/ZnSOD and MnSOD), catalase (CAT), and glutathione peroxidase (GPx), were identified to be differentially expressed during hypoxia-reoxygenation treatment, implying their distinct modulation roles on reoxygenation-induced oxidative stress. The full-length transcriptome and the critical genes characterized should contribute to the revelation of intrinsic molecular mechanism being associated with hypoxia/reoxygenation regulation and provide useful foundation for future genetic breeding of the red swamp crayfish.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Red swamp crayfish Procambarus clarkii is a kind of widespread freshwater crayfish species and has become the most produced commercial crustacean around the world in 2020 (Oficialdegui et al. 2020; Xu et al. 2021). In China, because of its great tasty, red swamp crayfish is the most popular casual delicacies around the year, especially in summer. In 2020, the gross cultivation area of the species in China was 1,456,420 hectares, producing approximately 2,393,700 tons of crayfish, which was ranked as the 6th most produced freshwater species in China in 2020 (Fishery Bureau of Ministry of Agriculture PRC 2021).
In aquaculture, dissolved oxygen (DO) is the most important determining factor because oxygen at efficient levels is necessary for aerobic metabolism of all creatures living in the water. During crayfish farming, to create an ideal living space and maintain excellent water quality, aquatic vegetation usually covers even more than 60% of the total areas in intensive crayfish rearing ponds. Owning to the aerobic respiration of the abundant aquatic grass plus algae and other aerobic creatures, hypoxia stress is commonly existing during night, especially at early hours of the morning, when all organisms will consume oxygen through respiration. Consequently, crayfish living in this kind of environments should clearly be more likely to expose to severe hypoxia for periods of several hours daily. Adverse effects of hypoxic stress have been widely revealed in various crustacean cultivation, leading to suppressed immune function, low feed conversion rate, slowed growth, or high-frequency of disease occurring (Hardy et al. 2012; Sun et al. 2019; Zhang et al. 2006). In red swamp crayfish rearing, hypoxia caused behavior disorders are commonly detected, severely hypoxia usually leads crayfish to climb vegetation to reach the surface where crayfish take the oxygen from the air (Reiber and McMahon 1998). In recent years, the gene regulation mechanism implicated in multiple signal pathways upon hypoxia stress has got a lot of attentions in various species (Toombs et al. 2011; Zhang et al. 2017). In vertebrates, hypoxia inducible factor 1 (HIF-1) was proved as a critical transcriptional activator of a series of genes associated with different adaption responses to decreased oxygen availability (Wang and Semenza 1995). Nevertheless, up to now, the molecular regulation mechanism of the key genes and pathways related to the hypoxia stress is still poorly understood in crustaceans, especially in red swamp crayfish.
Despite the depletion of DO concentration at night (hypoxia), however, ideal DO concentration or even super-saturation at a certain time period will generally be easy to keep during sunlit periods when oxygen is produced by photosynthesis of the aquatic grass. Therefore, red swamp crayfish are especially likely to be exposed to a kind of hypoxic-reoxygenation stress due to the aerobic respiration of aquatic grass, combined with rhythmic, light-dependent photosynthetic activity. In life activities of humans and other organisms, reoxygenation has been proved to be a special condition of high reactive oxygen species (ROS) production. Cellular damage occurring upon reoxygenation might be induced by the overproduced ROS (Ma et al. 2021; Radka and Gabriela 2018). Antioxidant enzymes possess critical functions in the ROS removal. Therefore, antioxidant enzymes play significant roles in the reduction of the oxidative damage in life activities (Zhao et al. 2014; Cui et al. 2021). The understanding of the specific functions for the antioxidant enzymes could help to reveal the mechanism responsible for the features associated with hypoxia-tolerant.
To elucidate the special regulation mechanism of the key genes or pathways under hypoxic-reoxygenation stress for the red swamp crayfish, we investigated the gene expression profiles of the hepatopancreas by using a method combining the second-(Illumina sequencing) and the third-generation transcriptome sequencing (PacBio full-length sequencing). Compared with Illumina sequencing, full-length transcriptome sequencing can provide a more accurate reference sequence data set, which is helpful for subsequent differential expression analysis. In addition, full-length transcriptome sequencing also facilitate us obtaining a larger number of genes with complete cDNA sequences containing both the 5′and 3′ ends (Wang et al. 2021). Based on the strategy combining second and third generation sequencing, the critical genes that are involved in the potential roles of regulating the hypoxia response and the tolerance process against oxidative stress were identified, and their features were characterized. The hypoxia-reoxygenation induced gene expressions, and the full-length transcriptome should help us not only to understand the molecular mechanism in response to hypoxia-reoxygenation, but also to reveal candidate target genes in future red swamp crayfish genetic breeding.
Materials and Methods
Animal Preparation and Hypoxia-Reoxygenation Treatment
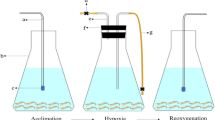
Healthy female red swamp crayfish were sampled in Nanjing, China, in April 2020. All samples were cultured in circulating water system at 20–22°C under a normal light-dark cycle and controlled DO of 6.8 ± 0.2 mg O2 L−1. After an acclimation period of 3 days, we divided the samples into experimental groups and control groups; each group contained three replicates respectively (15 crayfish per replicate). The control group was maintained under normal conditions with DO concentration of 6.8 ± 0.2 mg O2 L−1. For the experimental group, the DO concentration of 1.0 ± 0.1 mg O2 L−1 was kept for 1 and 6 h by nitrogen bubbling. Four samples from the experimental groups were randomly collected from the different replicates at each different time point respectively: at 1 h and 6 h under hypoxia and at 1 h and 12 h under reoxygenation (groups of hyp-1h, hyp-6h, reo-1h, and reo-12h). At the same time, four crayfish being held in normoxia conditions, which correspond with the time-matched samples (the groups with hypoxia-reoxygenation treatment), were pooled and used as controls (group of nor). Hepatopancreas from each sample was freshly dissected and frozen in liquid nitrogen for the transcriptome sequencing.
Total RNA Extraction, Library Preparation, and Sequencing
Total RNA was extracted from the hepatopancreas of each crayfish using TRIzol reagent (Invitrogen, USA). A quantity of 2 µg total RNAs from each crayfish was utilized to build an Illumina cDNA library. Four samples from each group were selected and used for cDNA libraries constructing. Accordingly, 20 libraries were prepared for the screening of critical DEGs related to the hypoxia-reoxygenation stress. On the same time, equal amounts of the total RNA from every crayfish in the 5 groups were pooled to get a reference full-length transcriptome by using PacBio sequencing. PacBio and Illumina cDNA library preparation and sequencing were performed by Genedenovo Biotechnology Co., Ltd (Guangzhou, China).
Reconstruction of the Full-Length Transcriptome and Identification of DEGs Correlated to Hypoxia-Reoxygenation
Full-length transcriptome reconstruction was performed using the SMRT Link v5.0.1 pipeline supplied by Pacific Biosciences (Gordon et al. 2015). The reconstructed reference full-length transcriptome was used for the subsequent gene expression quantification analysis. The gene expressions were calculated and normalized to FPKM (fragments per kilobase per million). The differential expression analysis was performed between the hypoxia-reoxygenation treated groups and control by using DESeq2 software and between two different samples by using edgeR software (Love et al. 2014). The significantly differentially expressed genes were judged with a false discovery rate (FDR) below 0.05 and absolute fold change more than 2. Kyoto Encyclopedia of Genes and Genomes (KEGG) pathways enrichment and Gene Ontology (GO) enrichment analysis were performed for the identified DEGs (Mao et al. 2005; Young et al. 2010). Trend analysis of the DEGs upon hypoxia-reoxygenation stress was carried out by using the Short Time-series Expression Miner (STEM, version 1.2.2b) software.
To confirm the accuracy of the transcriptome sequencing, 12 DEGs that potentially associated with the regulating of hypoxia-reoxygenation were chosen and measured with RT-qPCR. RNA sample used in the transcriptome sequencing was reverse transcribed with an Advantage® RT-for-PCR Kit it (Clontech, USA). Fold changes in expressions of the selected genes were evaluated with a reference gene of 18S rRNA according to the 2-ΔΔCt method (Livak and Schmittgen 2001). Primers were designed according to the transcriptome sequencing data (Table S1). All RT-qPCR were performed using SYBR Green PCR Master Mix (Takara, Dalian, China) on an ABI 7300 Real-Time PCR System (Applied Biosystems Inc., Waltham, MA, USA), and performed in triplicate.
Characterization of Vital Genes Involved in Hypoxia-Reoxygenation Response
The ORF (Open Reading Frame) of the vital candidate genes being associated with the regulations of hypoxia-reoxygenation, which included some hypoxia-induced and antioxidant-related genes in Hif-1 and antioxidant pathways, were determined with the online program of ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.html). Domains were predicted using SMART server (http://smart.embl-heidelberg.de/). Phylogenetic relationship was constructed using neighbor-joining (NJ) method with MEGA6.0 software according to the deduced amino acid sequences (Tamura et al. 2013). Bootstrap trials were replicated 1000 times to derive the confidence value for the phylogeny analysis. The amino acid sequence of the Pc-Hif-1α was further blast to the database of STRING (http://string-db.org/) to get the predicted protein-protein interaction (PPI), which were further represented with Cytoscape (Shannon et al. 2003).
Results
Full-Length Transcriptome Sequencing and Functional Annotations
Through full-length transcriptome sequencing, 59.85 Gb clean data that included 907,959 circular consensuses (CSS), with a mean length of 1,836 bp, were acquired after applying the filters. By clustering and correcting the full-length non-chimeric reads (FLNC), 24,311 high-quality transcripts and 292 low-quality transcripts were obtained. After eliminating the redundancy, 21,970 unique isoforms were remained. Functional annotations were carried out for the isoforms with 4 protein databases. Among the 18,553 annotated transcripts, 18,512 sequences showed significant matches within the Nr database, and 16,044 sequences yielded significant matches in the Swissport database. The full-length transcriptome reconstruction is shown in Table 1. The data sets have been submitted to NCBI Sequence Read Archive (SRA) under Accession PRJNA746726.
Identification of DEGs Being Relevant to the Hypoxia-Reoxygenation
High-quality clean reads ranging from 39,044,058 to 51,167,500 were acquired from the samples of the groups of nor, hyp-1h, hyp-6h, reo-1h, and reo-12h with Illumina sequencing (Table S2). On the reference of the reconstructed full-length transcriptome, differentially expressed genes being related to hypoxia-reoxygenation stress were screened and functionally annotated. During the hypoxia period, 389 and 533 significantly different expressed genes were detected in the groups of hyp-1h and hyp-6h respectively. Among the screened genes, 280 and 362 DEGs were up-regulated, while 109 and 171 DEGs were down-regulated. When exposed to reoxygenation, 533 and 783 significantly DEGs were identified at 1 h and 12 h, respectively (Fig. 1A, B) (Table S3). Further functional categorization assigned 351, 466, 457, and 600 DEGs to 148, 118, 165, and 150 KEGG pathways for the four groups of samples with different treatment respectively. When exposed to hypoxia (hyp-6h), the top 3 enriched pathways were “MAPK signaling pathway,” “protein processing in endoplasmic reticulum,” and “endocytosis.” When exposed to reoxygenation (reo-12h), the top three abundant pathways were “protein processing in endoplasmic reticulum,” “spliceosome,” and “endocytosis” (Fig. 1C, D). Hierarchical cluster and trend analysis were further performed to investigate the gene expression patterns associated with hypoxia-reoxygenation stress (Fig. 2A). A total of 1,605 DEGs were categorized in 20 profiles. The composition and function annotation of the 20 profiles are shown in Table S4. The significantly enriched profiles included two up-regulated patterns throughout the process of hypoxia-reoxygenation (profiles 18 and 19) and two temporary down-regulated patterns during the early reoxygenation stage (profile 11, 17) (Fig. 2B). Profile 18 (368 DEGs) and profile 19 (246 DEGs) comprise the largest amount of DEGs whose expressions were up-regulated constantly, indicating an increasing expression of these genes occurred at the whole hypoxia-reoxygenation stage. In profile 18, the DEGs with the greatest fold changes when exposed to hypoxia were heat shock 70 kDa protein (Hsp70), sortilin-related receptor, and UDP-glucuronosyltransferase (UGT). In profile 17, the DEGs with the greatest fold changes when exposed to reoxygenation were endo-beta-1, 4-glucanase (Egla), amylase (Amy), and Arg kinase (Ak), implying the potential influences of hypoxia-reoxygenation on nutrient absorption and energy metabolism of the red swamp crayfish.

Distribution and functional enrichment of the differentially expressed genes (DEGs) upon hypoxia-reoxygenation. A The number of DEGs upon the hypoxia-reoxygenation treatment. B The number of DEGs identified in different groups. C KEGG pathway enrichment of the DEGs upon hypoxia (hyp-6h). D KEGG pathway enrichment of the DEGs upon reoxygenation (reo-12h)

Clustering and trend analysis of differentially expressed genes (DEGs) upon hypoxia-reoxygenation. A Hierarchical clustering of the DEGs based on the averaged log2 (FPKM + 1) values. B Trend analysis of the DEGs during hypoxia-reoxygenation treatment. The 1,605 DEGs were enriched in 20 profiles. Profiles with grey backgrounds indicate the significantly enriched patterns with a Q-value less than 0.05
Characterization of Critical Genes Being Related to Hypoxia-Reoxygenation
Hypoxia-Induced Genes
On reference of the obtained full-length transcriptome, we identified two transcripts with complete ORFs that encode the subunit of hypoxia-inducible factor 1 (Hif-1, Pc-Hif-1α/β), which is a transcriptional activator of a variety of genes closely related to the hypoxia adaptions (Fig. 3). Pc-Hif-1α cDNAs contains a 3,279 bp full-length ORF which encodes a protein with 1,092 amino acids (aa) (NCBI Accession number MZ476857). Pc-Hif-1α has a conserved HLH domain (residue 25-82 aa), a PAS domain (residue 98-157 aa), and a PAS-B domain of NCOA1 (residue 232-333 aa) (Fig. 3A). Pc-Hif-1β cDNAs contains a 1,668 bp ORF that encodes a protein with 555 aa (NCBI Accession number MZ476858). Pc-Hif-1β has a highly conserved PAS (residues 32–94 aa) and a PAS-B domain of NCOA1 (nuclear receptor coactivator 1) (223–324 aa) (Fig. 3B). When exposed to hypoxic-reoxygenation, the expression of Pc-Hif-1α was significantly up-regulated in the hyp-1h, hyp-6h, and reo-1h group; however, no significant difference was detected in the reo-12h group.

Protein sequence characteristics and phylogenetic analysis of the key genes in hypoxia-inducible factor 1 (Hif-1) pathway of the red swamp crayfish. A Hif-1α; B Hif-1β; C prolyl-4-hydroxylase, PHD; D factor-inhibiting Hif-1, FIH-1. The diagrams showed the structures of the encoded proteins, and the boxes in the diagrams indicated the conserved domains. Phylogenetic analysis was performed with the deduced protein sequences of the related species
Two Hif-1 negative regulators, prolyl-4-hydroxylase (PHD) and factor-inhibiting Hif-1 (FIH-1), which both function as sensors of oxygen concentrations and possess the functions of down-regulating the HIF-1α expression and transactivation activity with their hydroxylation activities, have also been identified. Pc-PHD cDNA comprises a 1,260 bp full-length ORF that encodes a putative protein with 419 aa (NCBI accession number MZ476859). Pc-PHD has a conserved zf-MYND domain (residue 34-72 aa) and a P4Hc domain (Prolyl 4-hydroxylase alpha subunit homologues) (residue 254-399 aa) (Fig. 3C). Pc-FIH-1 cDNA contains a 1,029 bp ORF that encodes a putative protein with 342 aa (NCBI Accession number MZ476860). Pc-FIH-1 has a highly conserved Cupin_8 domain (residues 38–280 aa) (Fig. 3D). During hypoxic-reoxygenation, the expression of Pc-PHD was significantly up-regulated in the groups of hyp-6h, reo-1h, and reo-12h, and no significant difference in FIH-1 expression was observed in different groups.
Based on the PPI network associated with the hypoxia regulation of Pc-Hif-1, 115 proteins that include 43 DEGs were predicted to be interacted with Pc-Hif-1α (Fig. 4). The network of the PPI between the Pc-Hif-1α and its partners contained 115 nodes and 2205 edges (Table S5). Among the potential interacting proteins, Ugt-1, Egfr, CASP2, EGLN1, Hsp90, and Hsp70 have been experimentally examined and proved to be the key genes within the Hif-1 pathway in other organisms (Luo et al. 2010; Zeng et al. 2021).

Protein-protein interaction network between the pc-Hif-1α and the transcripts identified in the full-length transcriptome sequencing. The nodes represent proteins, and the edges represent interactions
Antioxidant-Related Genes
Reoxygenation-induced oxidative damage can be significantly relieved through the regulation roles of antioxidant enzymes. Based on the reconstructed full-length transcriptome, a series of antioxidant enzymes including two types of superoxide dismutase (Cu/ZnSOD and MnSOD), catalase (CAT), and glutathione peroxidase (GPx) were identified, which might be responded for the cellular damage protection during reoxygenation (Fig. 5). Pc-Cu/ZnSOD cDNAs contains a 1,491 bp full-length ORF that encodes a putative protein with 496 aa (NCBI Accession number MZ476863). Pc-Cu/ZnSOD contains a Cu-Zn_Superoxide_Dismutase domain and 8 conserved amino acids responsible for binding Cu2+ and Zn2+ (Fig. 5A). Both mitochondrial (mMnSOD) and cytosolic MnSODs (cMnSOD) were identified in the red swamp crayfish, and the full-length ORF of the Pc-mMnSOD (Nucleotide Accession number MZ476862) and Pc-cMnSOD (NCBI Accession number MZ476861) encode polypeptide with 286 aa and 218 aa respectively. A conserved SodA domain (residue 83-282 aa) and a conserved PLN02471 domain (residue 2-208 aa) were identified in the Pc-mMnSOD and Pc-cMnSOD, respectively (Fig. 5B). Pc-CAT cDNA comprises a 1,548 bp ORF that encodes a putative protein with 515 aa (NCBI Accession number MZ476864). Pc-CAT has a highly conserved catalase_clade_3 (residues 64–495 aa) domain and 15 conserved amino acids responsible for NADPH binding (Fig. 5C). The 408bp full-length ORF of the Pc-GPx encodes a 135 aa protein with a conserved GSH_Peroxidase domain (Fig. 5D) (NCBI Accession number MZ476865). During hypoxic-reoxygenation, the expression of Cu/ZnSOD was significantly up-regulated in the hyp-6h group and was significantly down-regulated in the reoxygenation stages. However, no significant different expressions were observed for both mMnSOD and cMnSOD in the whole hypoxic-reoxygenation treatment. Hypoxia caused a significant up-regulation of CAT in the groups of hyp-1h and reo-1h and a significant down-regulation in the group of reo-12h. The expressions of GPx were severely influenced in the whole hypoxia/reoxygenation process, implying its vital roles in the antioxidant defense of the red swamp crayfish.

Protein sequence characteristics and phylogenetic analysis of the key genes in antioxidant pathway in the red swamp crayfish. A Cu/Zn superoxide dismutase, Cu/ZnSOD; B MnSOD, mitochondrial MnSOD (mMnSOD) and cytosolic MnSODs (cMnSOD); C catalase, CAT; D glutathione peroxidase, GPx. The diagrams showed the structures of the encoded proteins, and the boxes in the diagrams indicated the conserved domains. Phylogenetic analysis was performed with the deduced protein sequences of the related species
Based on the expression profiles of the upstream and downstream genes in Hif-1 and antioxidant signaling pathways, we suggested a speculated outline illustration for the molecular pathways of hypoxia/reoxygenation regulation of the red swamp crayfish (Fig. 6). During hypoxia/reoxygenation, Hif-1α and some Hif-1 downstream genes, such as Ugt-1, Egfr, Igfbp-1, Pk, Hsp70, and Hc-1, were significant differentially expressed when exposed to hypoxia stress. A series of antioxidant enzymes, including two types of superoxide dismutase (Cu/ZnSOD and MnSOD), catalase (CAT), and glutathione peroxidase (GPx), were identified to be differentially expressed during hypoxia-reoxygenation treatment, implying their distinct modulation roles on reoxygenation-induced oxidative stress in red swamp crayfish.

The speculated outline illustration reveals the relationship among the key candidate genes in Hif-1 and antioxidant pathways in response to hypoxia-reoxygenation in the hepatopancreas of the red swamp crayfish. The FPKM levels of the significant differentially expressed genes in Hif-1 and antioxidant pathways have been provided as line charts beside the gene symbol. The numbers of 0, 1, 2, 3, and 4 on X-axis represented the different experimental groups of nor, hyp-1h, hyp-6h, reo-1h, and reo-12, respectively
Confirming the Accuracy of Transcriptome Sequencing with RT-qPCR
Twelve candidate genes potentially involved in the regulation of hypoxic-reoxygenation were randomly selected for RT-qPCR assays to confirm the accuracy and reliability of the Illumina sequencing. The selected genes included Hif-1α, UDP-glucose transferase 1 (Ugt-1), epidermal growth factor receptor (Egfr), hemocyanin 1 (Hc-1), chitinase 1 (Cht1), insulin-like growth factor binding protein (Igfbp-1), peritrophin-44 (Pm), hemocyanin subunit 2 (Hc-2), heat shock protein 70 (Hsp70), pyruvate kinase (Pk), hexokinase (Hk), and Cu/ZnSOD. Among the selected genes, Ugt-1, Egfr, Igfbp-1, Pk, Hsp70, and Hc-1 have been experimentally examined and proved to be the Hif-1 downstream genes in other organisms (Luo et al. 2010; Zeng et al. 2021). The results showed that the fold changes in expression generated through RNA-seq sequencing did not exactly match the expressions detected by RT-qPCR assays, but the expression profiles were essentially consistent for all the selected genes. The results confirmed the reliability and accuracy of the gene expressions generated from the RNA-seq sequencing (Fig. 7).

Confirmation of the accuracy of the gene expressions generated from the RNA-seq sequencing with RT-qPCR analysis. Candidate genes potentially related to the regulation of hypoxic-reoxygenation responses were selected for the RT-qPCR assay with the RNAs derived from the transcriptome sequencing. Fold change was determined with a reference gene of 18S rRNA according to the 2-ΔΔCt method. The Y-axis on the right showed the relative gene expression analyzed by RT-qPCR (blue lines), while the Y-axis on the left showed the corresponding expression data (FPKM) of the RNA-Seq (red lines). Each value represented the mean ± standard error for three to four replicates. The X-axis represented the different experimental groups
Discussion
Photosynthesis and aerobic respiration of the abundant aquatic grass and algae usually caused diurnal or daily fluctuations of dissolved oxygen in crayfish rearing. In addition, if the water quality of the pond was not exactly managed, overproduced organic pollutant, including residual fodder and decayed creatures, would deposit in the pond bottom, the sediment will become the hotbeds of various aerobic and many harmful anaerobic bacteria. Therefore, severe hypoxia during night often occurs in the red swamp crayfish rearing pond. For a long time, people have thought that red swamp crayfish possess the great ability to tolerate hypoxia. According to the record reports, in times of drought, red swamp crayfish can even survive periods in excess of 28 days without any access to free water (David 2002). Therefore, it is generally believed that red swamp crayfish can survive and grow vigorously under low DO concentrations. However, in red swamp crayfish rearing, it has been found that mild or moderate hypoxia may cause physiology disorder, leading to a complex physiological response to hypoxia stress. In severely hypoxia rearing conditions, red swamp crayfish will have to consume the oxygen from the air by climbing to the vegetation to reach the water surface. Reiber and McMahon (1998) observed a declined heart rate and an increased hemolymph flow in red swamp crayfish when exposed to hypoxia, implying a mechanism to maintain oxygen supply in response to hypoxia. Changes in gene expression regulations in many energy producing pathways have been implicated in compensation to hypoxia exposure in several other aquaculture species (Bernd and Margit 2018); however, up to now, little information is available for the red swamp crayfish.
The abnormality of physiological and metabolic in creatures induced by hypoxic stress seemed to share a common model of molecular regulation, which is usually regulated by the hypoxia-inducible factors. In invertebrates and vertebrates, hypoxia-inducible factor 1 (Hif-1) serves as a transcriptional activator of numerous genes with various functions in life activities (Koyasu et al. 2018). In the past few decades, much attention has been focused on the elucidating of the molecular mechanism regarding the Hif-1 activation in many biological processes. The regulation of Hif-1α expression is a critical determinant of HIF-1 activity. The major mechanisms underlying the regulation of Hif-1α protein stability has been detailed investigated in model organisms. HIF-1α stability was controlled by a kind of O2/PHD/VHL-dependent and independent mechanisms (Gassmann et al. 2002). Besides the proteolytic mechanism triggered by PHD, another dioxygenase, FIH-1, was proven to possess critical roles for the HIF-1 inactivation in normoxic conditions. In our present study, Hif-1α/β and its negative regulator, PHD and FIH-1, were both recognized and characterized from the red swamp crayfish hepatopancreas full-length transcriptome data. Be similar to the findings in various model organisms, when exposed to hypoxia, Hif-1α expression was significantly up-regulated, and Hif-1β expression remained stable in hepatopancreas of the red swamp crayfish. The activated HIF-1α/β heterodimeric complex can bind to the hypoxia-responsive elements (HREs) of the target genes and regulate the expressions of these target genes to restore tissue homeostasis under hypoxia stress (Poellinger and Johnson 2004). In this research, some experimentally proven Hif-1 downstream genes, such as Hsp70, Egfr, and Igfbp-1, were significant differentially expressed in hepatopancreas when exposed to hypoxia stress (Luo et al. 2010; Zeng et al. 2021). Among the different heat shock proteins, Hsp70 family is the most conserved and widely distributed. In recent years, Hsp70 has drawn much attention because it is fundamental to cellular and developmental processes and functions during stress when the organism is subjected to various external stresses (Fang et al. 2012). Among the differentially expressed genes, a molting related gene encoding the chitinase was identified. In crustaceans, Chitinases possess critical functions both in digesting chitinous food and degrading exoskeletons during molting cycles (Li et al. 2015). In the present study, the chitinase expression was significantly down-regulated when exposed to hypoxia stress, probably implying adverse effects of hypoxia on growth or development in the red swamp crayfish. KEGG pathway enrichment of the DEGs identified in the hypoxia group revealed that the top 3 enriched pathways were “protein processing in endoplasmic reticulum,” “MAPK signaling pathway,” and “endocytosis.” Mitogen-activated protein kinase (MAPK) signaling pathway was pivotal and intensively studied signaling pathways in hypoxic conditions in a wide variety of creatures (Minet et al. 2001). The activity of HIF-1 is mediated by phosphorylation and subsequent activation, which is regulated by various kinase pathways. The MAPK signal transduction pathways have been proposed to act as the downstream oxygen sensors and play a key role in the activation of these kinases, which is essential for the modulation of cell growth, differentiation, and cell death in various life activities (Mylonis et al. 2006). In red swamp crayfish, when exposed to hypoxia, some MAPK pathway genes including Hsp70, Egfr, and Elk3, showed significantly differential expression in response to the treatment, implying the potential critical regulation roles of the MAPK/Hif-1α pathways upon hypoxia stress.
There have been extensive studies on the metabolic disorders occurring during hypoxia; however, experimental evidences have showed that the subsequent reoxygenation was another major event causing cell and tissue dysfunction (Biddlestone et al. 2015). Cellular damage occurring upon reoxygenation may be caused by the ROS overproduction. Antioxidant enzymes possess critical roles in the removal of ROS and, therefore, have significant effects in reducing the oxidative damage during reoxygenation. SOD, GPx, and CAT are the main antioxidants that protect cell compartments against oxidative damage (Milkovic et al. 2019). Among the antioxidant enzymes, SODs make up the first defense system that catalyzes the dismutation of superoxide radicals to oxygen and hydrogen peroxide, which is further reduced by a reaction catalyzed by CAT or GPx (McCord and Fridovich 1969). In Litopenaeus vannamei, CAT, SOD, and GPx enzyme activities were significantly affected by hypoxia and reoxygenation (Parrilla-Taylor and Zenteno-Savín 2011; Trasviña-Arenas et al. 2013). Recently, Estrada-Cárdenas et al. (2021) explored the influences of hypoxia/reoxygenation and high temperature on the antioxidant response of L. vannamei and observed specific gene expression and enzymatic activity changes of MnSOD, GPx, and CAT in hepatopancreas. In crayfish rearing, due to the photosynthesis and aerobic respiration of abundant aquatic grass, daily fluctuations of the concentration of dissolved oxygen are commonly observed in the pond; therefore, red swamp crayfish are particularly likely to expose to the hypoxia-reoxygenation stress. In our study, three types of SODs including Cu/ZnSOD, cytosolic MnSODs (cMnSOD), and mitochondrial matrix (mMnSOD) were identified in the full-length transcriptome data. Only Cu/ZnSOD showed significant differentially expression during the whole hypoxia-reoxygenation treatment, suggesting the vital roles of Cu/ZnSOD in antioxidant regulation of the red swamp crayfish. During the whole reoxygenation stages, CAT and GPx were significantly and continuously up-regulated, indicating the modulation roles of the two antioxidant enzymes for the reoxygenation-induced oxidative stress. In a word, our data indicated that the significant up-regulated Cu/ZnSOD, CAT, and GPx should act as critical antioxidant when ROS is supposed to reach high levels during reoxygenation. In future studies, in-depth functional analyses are required to reveal the cellular and molecular basis of the antioxidant responses in red swamp crayfish under hypoxia/reoxygenation, including the evaluation of these critical antioxidant enzymes both in transcription and translation levels in various tissues.
Altogether, the full-length hepatopancreas transcriptome and the critical genes characterized should help us to understand the intrinsic mechanism related to hypoxia/reoxygenation regulation and should supply us useful basis for future genetic selection of the red swamp crayfish.
References
Bernd P, Margit E (2018) Hypoxia-inducible transcription factors in fish: expression, function and interconnection with the circadian clock. J Exp Biol 221(13):jeb163709
Biddlestone J, Bandarra D, Rocha S (2015) The role of hypoxia in inflammatory disease (review). Int J Mol Med 35:859–69
Cui Z, Liu Y, Yuan J, Yuan JB, Zhang XJ, Ventura T, Ma KY, Sun S, Song CW, Zhan DL, Yang YA, Liu HR, Fan GY, Cai QL, Du J, Qin J, Shi CC, Hao SJ, Fitzgiibbon QP, Smith GG, Xiang JH, Chan TY, Hui M, Bao CC, Li FH, Chu HK (2021) The Chinese mitten crab genome provides insights into adaptive plasticity and developmental regulation. Nat Commun 12:2395
David MH (2002) Biology of freshwater crayfish [M]. Blackwell Science Ltd, Oxford, pp 541–574
Estrada-Cárdenas P, Cruz-Moreno DG, González-Ruiz R, Peregrino-Uriarte AB, Leyva-Carrillo L, Camacho-Jiménez L, Quintero-Reyes I, Yepiz-Plascencia G (2021) Combined hypoxia and high temperature affect differentially the response of antioxidant enzymes, glutathione and hydrogen peroxide in the white shrimp Litopenaeus vannamei. Comp Biochem Physiol A Mol Integr Physiol 254:110909
Fang DA, Wang Q, He L, Wang J, Wang Y (2012) Characterization of heat shock protein 70 in the red claw crayfish (Cherax quadricarinatus): evidence for its role in regulating spermatogenesis. Gene 15;492(1):138–147
Fishery Bureau of Ministry of Agriculture PRC (2021) China Fishery Statistical Yearbook. China Agriculture Press, Beijing (In Chinese)
Gassmann M, Chilov D, Wenger RH (2002) Regulation of the hypoxia-inducible factor-1α. In: Lahiri S., Prabhakar N.R., Forster R.E. (eds) Oxygen Sensing. Adv Exp Med Biol vol 475. Springer, Boston, MA
Gordon SP, Tseng E, Salamov A, Zhang JW, Meng XD, Zhao ZY, Kang DW, Underwood J, Grigoriev IV, Figueroa M, Schilling JS, Chen F, Wang Z (2015) Widespread polycistronic transcripts in fungi revealed by single-molecule mRNA sequencing. Plos One 10(7):e0132628
Hardy KM, Follett CR, Burnett LE, Lema SC (2012) Gene transcripts encoding hypoxia-inducible factor (HIF) exhibit tissue- and muscle fiber type-dependent responses to hypoxia and hypercapnic hypoxia in the Atlantic blue crab. Callinectes sapidus. Comp Biochem Physiol A 163:137–146
Koyasu S, Kobayashi M, Goto Y, Hiraoka M, Harada H (2018) Regulatory mechanisms of hypoxia-inducible factor 1 activity: Two decades of knowledge. Cancer Sci 109:60–571
Li XG, Xu ZQ, Zhou G, Lin H, Zhou J, Zeng QF, Mao ZG, Gu XH (2015) Molecular characterization and expression analysis of five chitinases associated with molting in the Chinese mitten crab, Eriocheir sinensis. Comp Biochem Physiol B: Biochem Mol Biol 187:110–120
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods 25:402–408
Love MI, Huber W, Anders S (2014) Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome biology 15:550
Luo W, Zhong J, Chang R, Hu H, Pandey A, Semenza GL (2010) Hsp70 and chip selectively mediate ubiquitination and degradation of hypoxia-inducible factor (hif)-1α but not hif-2α. J Biol Chem 285:3651–3663
Ma H, Li Y, Hou T, Li J, Gong Z (2021) Sevoflurane postconditioning attenuates hypoxia/reoxygenation injury of cardiomyocytes under high glucose by regulating hif-1α/mif/ampk pathway. Front Pharmacol 11:624809
Mao XZ, Cai T, Olyarchuk JG, Wei LP (2005) Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 21:3787–3793
McCord JM, Fridovich I (1969) Superoxide dismutase. An enzymic function for erythrocuprein (hemocuprein). J Biol Chem 244:6049–6055
Milkovic L, Cipak Gasparovic A, Cindric M, Mouthuy P-A, Zarkovic N (2019) Short overview of ROS as cell function regulators and their implications in therapy concepts. Cells 8:793
Minet E, Michel G, Mottet D, Raes M, Michiels C (2001) Transduction pathways involved in Hypoxia-Inducible Factor-1 phosphorylation and activation. Free Radic Biol Med 31(7):847–55
Mylonis I, Chachami G, Samiotaki M, Panayotou G, Paraskeva E, Kalousi A, Georgatsou E, Bonanou S, Simos G (2006) Identification of MAPK phosphorylation sites and their role in the localization and activity of hypoxia-inducible factor-1alpha. J Biol Chem 281:33095–106
Oficialdegui FJ, Sánchez MI, Clavero M (2020) One century away from home: how the red swamp crayfish took over the world. Rev Fish Biol Fisheries 30:121–135
Parrilla-Taylor DP, Zenteno-Savin T (2011) Antioxidant enzyme activities in Pacific white shrimp (Litopenaeus vannamei) in response to environmental hypoxia and reoxygenation. Aquculture 318:379–383
Poellinger L, Johnson RS (2004) HIF-1 and hypoxic response: the plot thickens. Curr Opin Genet Dev 14:81–85
Radka C, Gabriela P (2018) Hif-1, metabolism, and diabetes in the embryonic and adult heart. Frontiers in Endocrinology. Front Pharmacol 9:460
Reiber CL, McMahon BR (1998) The effects of progressive hypoxia on the crustacean cardiovascular system: a comparison of the freshwater crayfish, (Procambarus clarkii) and the lobster (Homarus americanus). J Comp Physiol 168B:168–176
Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T (2003) Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res 13:2498–2504
Sun S, Wu Y, Jakovli I, Fu H, Jin S (2019) Identification of neuropeptides from eyestalk transcriptome profiling analysis of female oriental river prawn (macrobrachium nipponense) under hypoxia and reoxygenation conditions. Comp Biochem Physiol B Biochem Mol Biol 241:110392
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol 30(12):2725–2729
Toombs C, Jost J, Frederich M (2011) Differential hypoxia tolerance and AMPK activity in two color morphs of the green crab. Carcinus maenas. Integr Comp Biol 51:E258
Trasviña-Arenas CH, Garcia-Triana A, Peregrino-Uriarte AB, Yepiz-Plascencia G (2013) White shrimp Litopenaeus vannamei catalase: gene structure, expression and activity under hypoxia and reoxygenation. Comp Biochem Physiol B Biochem Mol Biol 164:44–52
Wang GL, Semenza GL (1995) Purification and characterization of hypoxiainducible factor 1. J Biol Chem 270:1230–1237
Wang L, Zhu P, Mo Q, Luo W, Du Z, Jiang J, Yang S, Zhao L, Gong Q, Wang Y (2021) Comprehensive analysis of full-length transcriptomes of Schizothorax prenanti by single-molecule long-read sequencing. Genomics 28:S0888-7543(21)35-50
Xu ZQ, Gao TH, Xu Y, Li XG, Li JJ, Lin H, Yan WH, Pan JL, Tang JQ (2021) A chromosome-level reference genome of red swamp crayfish Procambarus clarkii provides insights into the gene families regarding growth or development in crustaceans. Genomics 113:3274–3284
Young MD, Wakefield MJ, Smyth GK, Oshlack A (2010) Gene ontology analysis for RNA-seq: accounting for selection bias. Genome Biol 11:R14
Zeng B, Liu L, Liao XT, Zhang CX (2021) Cardiomyocyte protective effects of thyroid hormone during hypoxia/reoxygenation injury through activating of IGF-1-mediated PI3K/Akt signalling. J Cell Mol Med 25:3205–3215
Zhang CS, Hawley SA, Zong Y, Li MQ, Wang ZC, Gray A, Ma T, Cui JW, Feng JW, Zhu MJ, Wu YQ, Li TY, Ye ZY, Lin SY, Yin HY, Piao HL, Hardie DGR, Lin SC (2017) Fructose-1, 6-bisphosphate and aldolase mediate glucose sensing by AMPK. Nature 548:112–116
Zhang PD, Zhang XM, Li J, Huang G (2006) The effects of body weight, temperature, salinity, pH, light intensity and feeding condition on lethal DO levels of whiteleg shrimp, Litopenaeus annamei (Boone, 1931). Aquaculture 256:579–587
Zhao DX, Chen LQ, Qin JG, Qin CJ, Li EC (2014) Molecular characterization of a cytosolic manganese superoxide dismutase from the Chinese mitten crab, Eriocheir sinensis. Genetics & Molecular Research Gmr 13:9429
Funding
This work was supported in part by the National Key R&D Program of China (no. 2020YFD0900303), the Agricultural Project from Jiangsu Province Science and Technology Agency (no. BE2020348), the Jiangsu Agricultural Science and Technology Innovation Fund (no. CX(20)3005), the earmarked fund for Jiangsu Agricultural Industry Technology System (no. JATS [2021] 409), the China Agriculture Research System (no. CARS-48), and the Agriculture Breeding Project supported by Jiangsu Provincial Department of Agriculture and Rural Affairs (no. PZCZ201746).
Author information
Authors and Affiliations
Corresponding authors
Ethics declarations
Conflict of Interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Xu, Y., Lin, H., Yan, W. et al. Full-Length Transcriptome of Red Swamp Crayfish Hepatopancreas Reveals Candidate Genes in Hif-1 and Antioxidant Pathways in Response to Hypoxia-Reoxygenation. Mar Biotechnol 24, 55–67 (2022). https://doi.org/10.1007/s10126-021-10086-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10126-021-10086-9