
Abstract
Chitin-metabolizing products are of high industrial relevance in current scenario due to their wide biological applications, relatively lower cost, greater abundance, and sustainable supply. Chitooligosaccharides have remarkably wide spectrum of applications in therapeutics such as antitumor agents, immunomodulators, drug delivery, gene therapy, wound dressings, as chitinase inhibitors to prevent malaria. Hypocholesterolemic and antimicrobial activities of chitooligosaccharides make them a molecule of choice for food industry, and their functional profile depends on the physicochemical characteristics. Recently, chitin-based nanomaterials are also gaining tremendous importance in biomedical and agricultural applications. Crystallinity and insolubility of chitin imposes a major hurdle in the way of polymer utilization. Chemical production processes are known to produce chitooligosaccharides with variable degree of polymerization and properties along with ecological concerns. Biological production routes mainly involve chitinases, chitosanases, and chitin-binding proteins. Development of bio-catalytic production routes for chitin will not only enhance the production of commercially viable chitooligosaccharides with defined molecular properties but will also provide a means to combat marine pollution with value addition.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
One of the major objectives of the present age of biotechnology is heading towards production and application of a range of bio-based specialized high-value products for emerging bioeconomy. Biopolymers are synthesized in living organisms by repeating, covalently bonded units of nucleotides, amino acids, or monosaccharides. Possession of a biocompatibility, low toxic, and environment-friendly nature leads to great interest in their production and application over current synthetic polymers. Chitin is considered as the second most abundant polymer on earth with immense biotechnological potential due to its huge prospects of structural modification which results in novel properties, functions, and applications (Pillai et al. 2009). This plentiful and renewable polymer is a major component of the cell wall of fungi, exoskeleton of crabs, lobster, shrimps, insects, squid, and octopus (Tharanathan and Kittur 2003). Chitin is a white, tough, inflexible, insoluble, nitrogenous polysaccharide contemplated to be responsible for the surface contamination of coastal regions. It arises in nature by way of well-ordered crystal-like microfibrils and occurs predominantly in three crystalline polymorphs: α, β, and γ that differ in the orientation of microfibrils. α-Chitin is predominantly present in fungal cell walls, tendons, and shells of lobster and crab, shrimp shells, and insect cuticle, formed by the antiparallel organization of microfibrils. Sheets are arranged in parallel fashion in the β-chitin, which is present in pens and vestimentiferan worms. The γ-chitin contains a mixture of antiparallel and parallel sheets and is detected in the insect cocoons (Rinaudo 2006). In spite of its abundance and being biocompatible, no major industrial application of this vastly available material has so far been developed due to its high degree of crystallinity and insolubility in aqueous as well as organic solvents. Subjecting it to inter-conversion to generate derivatives viz. chitosan, chitooligosaccharides (COSs), and glucosamine with novel and improved characteristics viz. increased solubility and less crystallinity may augment its commercial feasibility.
Chitosan, the polycationic biopolymer is the deacetylated form of chitin. It is soluble in aqueous acidic media such as dilute acetic acid and formic acid due to protonation. The solubility of chitosan depends on its degree of crystallization and on the distribution of the acetyl groups along with the main chain in addition to the molecular weight (Dutta et al. 2004). Chitosan possesses three types of reactive functional groups, an amino/acetamido group, both primary and secondary hydroxyl groups at the C6, C2, and C3 positions, respectively. The amino content is the main reason for the difference between structures and physicochemical properties of chitosan and is correlated with their chelation, flocculation, and biological functions (Pareek et al. 2011). The high nitrogen content (about 6.89%) makes it a commercially noticeable compound. Besides chitin and chitosan, there is great interest in their degradation products, i.e., COS which show a better adsorption level in the human intestine, hence contribute in various biomedical applications.
COSs are homo or heterooligomers of N-acetylglucosamine and D-glucosamine units in varying proportions. Functional properties of COSs have largely been depending on their degree of polymerization (DP) along with the number and sequence of acetylated and deacetylated units. The biological activity of COS increases with the escalation in DP but it is solubility dependent (Das et al. 2013). The exceptional biotic properties like antimicrobial activity, anticancer effect, antioxidant effect, and immune-stimulating activity make COS molecule of choice in the present era of biotechnology. Safety and ease of control of the enzymatic preparation of COS is gaining attention over the chemical preparations. The enzymatic preparations utilized for COS production includes chitinases, chitosanases, cellulases, lipases, and proteases (Xia et al. 2011). High emphasis from various groups has been given on the enzymatic production of COS for enhanced yields along with better control on the DP and degree of acetylation (DA) to get improved applicability. This review is focused on the different aspects of improved COS production along with applications in diverse industrial sectors. Moreover, this study will also put a light on the latest molecular and biochemical approaches in the field.
Production of Chitooligosaccharides
Crystallinity and insolubility of chitin present a formidable challenge towards its industrial application. Commercial feasibility of chitin may be augmented by its interconversion into COS which is one of the major degradation products of chitin and chitosan. COS may be produced by depolymerization of chitin or chitosan using physical and chemical hydrolysis as well as enzymatic degradation. Physical methods of production include hydrothermal treatment, microwave irradiation, ultra-sonication, and gamma-rays. Among the chemical methods for the preparation of COS, acid phosphoric acid, hydrofluoric acid, formic acid, lactic acid, and hydrogen peroxide for depolymerization. Chemical methods have several limitations due to the formation of toxic secondary compounds, which are difficult to remove and create higher risk to environment because of process complexity. Moreover, the acid hydrolysis mostly produces oligomers of low DP mainly monomer to tetramer not the more beneficial pentamer to heptamer (Mourya et al. 2011).
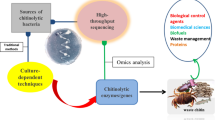
Since the last two decades, enzymatic hydrolysis of chitin and chitosan to prepare COS (MW ≤ 10 kDa) is of much interest due to the pharmaceutical, medical, agricultural, and nutritional applications of the produced oligomers. Enzymatic production of COS not only gives a better control on the DP and DA but also has no negative impact on the environment (Lodhi et al. 2014). An array of carbohydrate-degrading enzymes specifically chitinolytic and chitosanolytic enzymes viz. chitinase, chitosanase, lysozyme, cellulase, pectinase, protease, lipase, and pepsin have been involved in the bioactive oligosaccharide preparation (Jung and Park 2014). Among these enzymes, chitinase and chitosanase have specificity to chitin and chitosan degradation respectively with varying mode of action (Fig. 1). Chitinases mainly attack highly N-acetylated polymers whereas chitosanases prefer low N-acetyl content.

Production of chitin, chitosan, and chitooligosaccharides
Chitinases in COS Production
Chitinases (E.C. 3.2.1.14) are the glycosyl hydrolases having a molecular weight ranging from 20 to 90 kDa with capability to degrade chitin to low-molecular-weight chitooligomers. Among the microbial chitinases, bacterial chitinases are well classified and mostly belong to the family 18 of the glycosyl hydrolases with some exceptions present in family 19. The chitin-binding domain is located either at the amino or carboxyl terminal in bacterial chitinases. However, most of the fungal chitinases belong to the family 18 of the glycosyl hydrolases and shows high amino acid homology with class III plant chitinases. The basic structure of the fungal chitinases consists of five domains viz. catalytic domain, chitin-binding domain, serine/threonine-rich domain, N-terminal signal peptide regions, and C-terminal extension region (Hamid et al. 2013). Chitinases may be divided into two major groups, i.e., endochitinases and exochitinases, according to the mode of action. The former randomly split chitin at internal sites whereas the latter have a progressive catalytic action starting at the non-reducing end of polymer with the release of chitobiose or glucosamine units. The endochitinases forms the diacetylchitobiose and soluble low molecular mass oligomers of GlcNAc for instance chitotriose and chitotetrose. The exochitinases may further be distributed into two subgroups: chitobiosidases (E.C. 3.2.1.29), which are involved in catalyzing the progressive release of diacetyl chitobiose starting at the non-reducing end of the chitin microfibril and 1,4 β-glucosaminidases (E.C. 3.2.1.30), which cleave the oligomeric products resulting from action of endochitinases and chitobiosidases and release glucosamine monomer (Harman et al. 1993; Sahai and Manocha 1993).
The production of N-acetylglucosamine from α chitin was reported by Il’ina et al. (Il'ina et al. 2004) through the enzymatic preparation-Celloviridin G20x. A 22 kDa chitinase purified from thermophilic Bacillus licheniformis strain JS showed production of NI,NII-diacetylchitobiose from colloidal chitin as substrate (Waghmare and Ghosh 2010). Songsiriritthigul et al. (2010) illustrated the potential of a recombinant endochitinase (ChiA) from B. licheniformis strain DSM8785 to convert colloidal chitin to N-acetyl glucosamine. Variants of B. licheniformis chitinase with improved catalytic efficiency were obtained by employing directed evolution approach (Songsiriritthigul et al. 2009). The mutant chitinase was reported to have high catalytic potential for bioconversion of chitin to COS. Wang et al. (2012) reported the production of a range of COS with varying DP (chitobiose, 0.44 mg ml−1; chitotriose, 0.08 mg ml−1; chitotetraose, 0.09 mg ml−1; and chitopentaose, 0.43 mg ml−1) from chitinase I and chitinase II employing purified chitinases from the culture supernatant of B. cereus TKU027 with shrimp head powder as the substrate. Yang et al. (2016) observed the highest conversion yield of COS viz. 89.5% (w/w) from colloidal chitin by employing recombinant enzyme (70 kDa) from Paenicibacillus barengoltzii. Recently, Kumar et al. (2017) reported COS conversion potentiality of the extracellular chitinase produced from thermophilic Humicola grisea. The researchers also stated that 1.43-fold enhancement in chitinase production has been achieved through optimization of the nutritional variables by the employment of response surface methodology. Ueda et al. (2017) reported the production of N-acetyl-chitobiose by a 60 kDa chitinase derived from recombinant Eisenia fetida. Phylogenetic analyses revealed that the E. fetida chitinase is an amphioxus chitotriosidase-like protein exhibiting maximum catalytic activity at 60 °C. Bacterium Chitiniphilus LZ32 was investigated for the conversion of chitin extracted from the larvae of housefly into COS (Zhang et al. 2017). The study illustrated that the maximum production of (GlcNAc)5 (51.2 μg ml−1) and (GlcNAc)6 (36.1 μg ml−1) was achieved following hydrolysis for 72 and 84 h, respectively.
Chitosanase in COS Production
Chitosanase (EC 3.2.1.132) catalyzes the endohydrolysis of β-1,4 glycosidic linkage between d-glucosamine residues of chitosan (Lombard et al. 2014). Chitosanases are mainly grouped in family 46 of the glycoside hydrolase class on the basis of sequence. Apart from this family, 3, 5, 7, 8, 75, and 80 also includes enzymes with chitosanase activities. Chitosanase acts on the reducing end of chitosan molecule and produces COS exclusively as the end product. Based on the specificity of the cleavage positions for the partially acetylated chitosan, chitosanases can be classified into three distinct subclasses viz. subclass I, II, and III. Subclass I chitosanases can split both GlcN–GlcN and GlcNAc–GlcN linkages, while subclass II enzymes could only cleave GlcN–GlcN linkages. Subclass III chitosanases are capable of splitting both GlcN–GlcN and GlcN–GlcNAc linkages (Viens et al. 2015). Numerous researchers have reported the ability of chitosanase to hydrolyze chitosan for the production of COS. Nidheesh et al. (2015) reported 4.43 mM of chitooligomers production from the hydrolysis of colloidal chitosan by using chitosanase purified from the Purpureocillium lilacinum CFRNT12 strain. de Assis et al. (2010) studied the COS production from enzymatic hydrolysis of chitosan from the fungus Metarhizium anisopliae. The study reported the production of diverse COS species (chitobiose, 0.2 mg ml−1; chitotriose 0.19 mg ml−1; chitotetraose, 0.06 mg ml−1; chitopentaose, 0.04 mg ml−1; and chitohexaose, 0.004 mg ml−1). A 45 kDa chitosanase from Bacillus sp. strain KCTC 0377BP has been reported to produce COS ranging from chitotriose to chitooctaose using chitosan as the substrate. GH46 chitosanase from B. subtilis was expressed in Escherichia coli by fusing the gene encoding mature chitosanase to the Escherichia coli ompA signal peptide sequence and further evaluated for the production of COS (Pechsrichuang et al. 2013). The authors reported the complete conversion of chitosan hexamer to a mixture of (GlcN)2, (GlcN)3, and (GlcN)4 following incubation for an hour at 30 °C. Su et al. (2017) also reported enhancement in chitosanase production through recombinant B. subtilis PT5 and statistical optimization of fermentation medium followed by COS production. The TLC analysis revealed the presence of dimer, trimer, and tetramer chitosan oligosaccharides. Sanchez et al. (Sánchez et al. 2017) studied the effect of production process of COS on physicochemical characteristics and antibacterial activity. The authors employed two different processes for COS preparation. The first process involved hydrolysis of chitosan by commercially available chitosanase from Streptomyces griseus while in the second process, a two-stage hydrolysis strategy was adopted (chemical one followed by chitosanase). The study illustrated that the COS produced by the first method exhibited better antibacterial potential against E. coli and Listeria monocytogenes as compared to the COS generated via second process in spite of the fact that more deacetylated COSs have been produced by later procedure.
Chitin-Binding Proteins
Employment of fundamentally slow processive enzymes can be concentrated by improving the substrate accessibility due to the utilization of CBPs and substrate-disrupting accessory proteins or domains (Vaaje-Kolstad et al. 2005a). CBP plays a significant dual role in the chitin degradation mechanisms. It may either enhance the substrate availability due to the disruption of crystalline structure or aids in the microbial attachment. According to the carbohydrate active enzymes database (http://www.cazy.org/), the carbohydrate active enzyme protein domains with chitin-binding properties are classified into families 1, 2, 12, 14, 18, 19, and 33. Among these, families 14, 18, and 33 contain the non-catalytic CBPs. Family 14 and 18 CBPs are chiefly present in fungi and family 33 CBPs generally belong to bacteria and viruses (Lombard et al. 2014). Suzuki et al. (2002) reported the synergism of chitinases and CBP21 of Serratia marcescens 2170. The study reported the role of CBP21 in chitin degradation in addition to the three chitinases (chi), i.e., chiA, chiB, and chiC1. Vaaje-Kolstad et al. (2005b) also studied the crystal structure and binding properties of CBP21 of S. marcescens. By site-directed mutagenesis and subsequent binding studies, they showed that the single-point mutation lowered the affinity of CBP21 for β-chitin. The essentiality of non-catalytic CBP21 in chitin degradation from S. marcescens was also proved by Vaaje-Kolstad et al. (2005a). CBP synergistic activity with chitinases has been studied by Das et al. (2013). The authors reported the presence of family 33 CBPs, i.e., CBP21, CBP28, and CBP50, in the genome sequence of S. proteamaculans 568. Among these CBPs, CBP21 and CBP50 show synergism with chitinases. Manjeet et al. (2013) observed that B. thuringiensis serovar kurstaki chitinases and S. proteamaculans chitinases B exhibit synergism with family 33 CBPs in the hydrolysis of chitin. Khoushab and Yamabhai (2010) reviewed and explained the role of CBPs in increasing the efficiency of chitinolytic enzymes. The review also summarized a repertoire of CBPs obtained from different sources viz. from bacteria, silkworms, and plants. Khoushab et al. (2012) also reported peptide-assisted synthesis of chitin-based nanomaterials. The chitin-binding peptides used were selected through phase display technology. The peptides can induce colloidal chitin and chitohexaose to form polymeric networks at comparatively higher temperatures (up to 42 °C). Biocompatibility, relatively smaller size, and ease of modification of chitin-binding peptides will open up a new era of chitin research for development of specifically functionalized chitin nanomaterials. The substrate-disrupting protein-CBPs can be considered as the novel tool to improve the substrate accessibility by specific affinity to crystalline structure and thus contribute to more efficient enzymatic chitin degradation that leads to the enhanced bioactive COS production.
Catalytic Mechanism
The binding mechanism of the carbohydrate substrate to their enzymes is difficult to analyze due to the speedy hydrolysis of the substrates. There are many carbohydrate-hydrolyzing enzymes that share similar domains. Most of the microbial chitinases belong to family 18 glycosidases among which chitinases from S. marcescence have been extensively studied. The enzymes contain four domains, i.e., N-terminal signal sequence, a catalytic domain, a serine/threonine-rich region, and a C-terminal chitin-binding domain. Family 18 chitinases possess a typical (α/β) eight-barrel structure composed of eight α-helices and β-sheets. It also contains an N-terminal β-strand-rich domain and a small (α + β) domain. These chitinases have the ability to degrade chitin with retention of stereochemistry at the anomeric carbon. The overall mechanism of chitonolysis involves firstly cleavage of the sugar chain followed by the formation of an oxazolinium ion and lastly hydrolysis of the ion (Fukamizo 2000; Van Aalten et al. 2001). Based on the X-ray crystal structures of chitinases and their complexes with substrate analogues, Fukamizo (2000) has discussed the catalytic mechanism of family 18 and family 46 chitosanases and showed that family 46 chitosanases has two carboxyl groups at the catalytic center. In this family of chitosanases, catalysis takes place through a single displacement mechanism. On the other hand, family 18 chitinases have one catalytic carboxylate as a proton donor with a substrate-assisted catalytic reaction.
Purification and Characterization of COS
COS purification is mainly done by using a non-volatile mixture separation technique, i.e., thin-layer chromatography (TLC), followed by one of the most reliant analytical technique of separation, identification, and quantification viz. high performance liquid chromatography (HPLC). Songsiriritthigul et al. (2010) illustrated the utilization of TLC for the analysis of the COS. The authors analyzed the conversion of colloidal chitin to N-acetyl glucosamine at pH 4.0, 6.0, and 9.0 (50 °C). Nidheesh et al. (2015) purified chitosan hydrolysates by using HPLC and reported the presence of chitosan-trimer and chitosan-tetramer. Yang et al. (2016) reported N,N-diacetyl chitobiose about 89.5% through HPLC-ELSD equipped with a Cosmosil Sugar-D column (4.61*250 mm). Wang et al. (2012) also used HPLC equipped with Nucleosil 5 NH2 column (4.6 *250 mm) and reported the concentrations of chitobiose, chitotriose, chitotetraose, and chitopentose as 0.44, 0.08, 0.09, and 0.43 mg/ml, respectively. The most reliable technique for COS characterization is mass spectrometry. It measures the characteristics of individual molecules on the basis of their mass to charge ratio (m/z). Wang et al. (2012) showed the MALDI-TOF MS spectra of the COS in which ions with m/z ratio 608 contains GlcN-(GlcNAc)2, m/z ratio 650 contains (GlcNAc)3, m/z ratio 811 contains GlcN-(GlcNAc)3, and m/z ratio 1014 contain GlcN-(GlcNAc)4.
Chitosan and COS-Based Nanobiotechnology
In the contemporary era of nanobiotechnology, chitin and its derivative-based nanomaterials are gaining significant consideration for biomedical applications (Fig. 2) (Manivasagan and Oh 2016; Elieh-Ali-Komi and Hamblin 2016). Biodegradable polymeric nanoparticles as well as metal nanoparticles decorated with biopolymers have been commonly used as drug delivery carriers as they own outstanding bioavailability, better encapsulation, controlled release, and less toxic properties (Kumari et al. 2010; Rai et al. 2015). The chitosan and COS-based nanomaterials exhibited improved physical and chemical properties viz. high surface area, porosity, tensile strength, conductivity, and mechanical properties that make them highly beneficial in drug delivery, gene delivery, and wound healing (Shukla et al. 2013; Elgadir et al. 2015). The composite network of chitosan nanofibers loaded with gold nanoparticles can improve the performance of cholesterol biosensors, fabricated by immobilizing cholesterol oxidase onto the composite network (Gomathi et al. 2011). Kono and Teshirogi (2015) have reported cyclodextrin-grafted chitosan hydrogel for drug delivery. The study reported utilization of carboxymethyl chitosan and carboxymethyl β-chitosan for the preparation of hydrogels using carbodiimide as crosslinker. The surface modification of nanoparticles through polymeric protective agents can play a key role in drug delivery and targeting as the surface-protected nanoparticles cannot be recognized by the reticuloendothelial system (Mogoşanu et al. 2016). Manivasagan et al. (2016) synthesized biocompatible paclitaxel-loaded gold nanoparticles using chitosan oligosaccharides as reducing and stabilizing agents. The prepared biocompatible nanoparticles loaded with paclitaxel were used in drug delivery and photoacoustic imaging of cancer cells. Antidiabetic ability of chitosan nanoparticles loaded with Stevia rebaudiana leaf extract has been studied in streptozotocin-induced diabetes mellitus rat models (Perumal et al. 2016). Chitosan nanoparticles fabricated using sodium tripolyphosphate as crosslinking agent were used to load letrozole for sustained release of the drug during cancer chemotherapy (Gomathi et al. 2017). The prepared formulation was biocompatible as well as hemocompatible. Li et al. (2017) synthesized chitosan oligosaccharide silver nanofibers by using polyvinyl alcohol through an electrospinning technique. The study presented that the prepared nanofibers promote fibroblast proliferation, collagen synthesis, and cell adherence. Ramezanzade et al. (2017) reported employment of chitosan-coated nanoliposomes as a carrier for antioxidant peptides derived from Oncorhynchus mykiss. Skin gelatin hydrolysates were encapsulated in chitosan-coated nanoliposomes with mean size in the range of 163.4–234 nm. The study showed that the antioxidant activity of peptide fractions (molecular mass ˂ 30 kDa) was at maximum in case of nanoliposomes coated with 0.4% (w/v) chitosan. Wang et al. (2017) synthesized and utilized hyaluronic acid-coated chitosan nanoparticles to enhance delivery of chemotherapeutic drug 5-fluorouracil into tumor cells, overexpressing CD44. In another study, Subhapradha and Shanmugam et al. (Subhapradha and Shanmugam 2017) fabricated β-chitosan nanoparticles and studied their anticancer potential against human hepatoma cells. The study reported that β-chitosan nanoparticles were showed optimum inhibitory action when employed at concentration of 30 μg ml−1.

Production and application of COS-based nanomaterials
Applications of Chitooligosaccharides
Chitin-derived oligosaccharides are observed to have applicability in diverse sectors viz. medicine, food, biotechnology, agriculture, and wastewater treatment (Table 1). Among these, COSs find widespread applications in the biomedical field viz. antimicrobial, anticancer, antioxidant, an anticoagulant agent, wound healing, immunomodulation, osteoconductivity, antihyperlipidermic functioning, apoptosis induction, hypoglycemic, chemoprevention, enzyme inhibition, drug delivery (Hamed et al. 2016; Muanprasat and Chatsudthipong 2017; Olicón-Hernández et al. 2017). These biological applications of COS are mainly attributed to their low molecular weight that enables them to be easily absorbed through the intestine and quickly get into the blood flow.
Medicine
COSs have been molecules of interest for the last two decades due to their reported abilities in the field of medicine that include drugs against asthma; as antibacterial and antifungal agent, anti-metastasis, and anticancer; wound-dressing materials; for increasing bone-strength; as anti-malaria; and as a vector in gene therapy and diabetics. However, the molecular mechanism behind these biological activities is largely unknown.
Tumor Growth Inhibition
The antitumor potential of COS has been explored since 1970 by various groups of researchers. Cationic and structural properties viz. molecular weight and DDA pose effects on the tumor cell growth. COS with a molecular weight ranging from 1.5 to 5.5 kDa could effectively inhibit the growth of Sarcoma 180 solid (S180) or Uterine cervix carcinoma No. 14 (U14) tumor in BALB/c mice reference. S180 tumor cell growth in mice may be inhibited by the water-soluble COS mixture of tetramer and pentamer after oral and intraperitoneal administration. The antitumor properties of COS have been observed to be related to their inhibitory effects on angiogenesis, i.e., the formation of new capillary blood vessels from already existing ones (Jeon and Kim 2002; Prashanth and Tharanathan 2005; Wang et al. 2007; Xiong et al. 2009). According to Xiong et al. (2009), chitohexaose was observed as the most effective inhibitor, whereas Wang et al. (2007) reported that 81% N-acetylated COS are the most effective agents in preventing angiogenesis both in vitro and in vivo. Recently, COSs have been observed to inhibit cell proliferation in human gastric cancer cell lines (Luo et al. 2016). Recently, chitosan- and COS-based nanomedicines are gaining much more attention in cancer research. Wu et al. (2016) fabricated a nanodrug delivery system using biotinylated carboxymethyl chitosan and calcium carbonate for co-delivery of an antitumor drug (doxorubicin hydrochloride) and a drug resistance inhibitor (tariquidar) to overcome multidrug resistance in cancer chemotherapy. Manivasagan et al. (2017) reported chitosan-polypyrrole nanocomposites as a novel means for photoacoustic imaging-guided photothermal ablation of breast cancer.
Wound Healing
COS can accelerate the wound healing process by enhancing functions of inflammatory and repairing cells. Chitohexaose both N-acetylated and deacetylated have been reported to be effective wound healing agents. The healing potential of COS can be attributed to their ability to form biocompatible ordered tissue-like structures and their hemostatic effects. COS had been observed to accelerate wound healing alone as well as with polyvinyl alcohol sponges (You et al. 2004). Biocompatibility of COS with nanofibrous membranes adds to their potentiality as wound-dressing agents (Aam et al. 2010; Wang et al. 2011; You et al. 2004). Sathiyaseelan et al. (2017) has developed nanocomposite sponges by employing fungal chitosan (from Cunninghamella elegans), aloe vera extract, and Cuscuta reflexa-mediated silver nanoparticles. The aloe vera extract reduced the toxicity of silver nanoparticles used in the nanocomposite. The synthesized nanocomposite had potentiality as an effective wound-dressing material and exhibited remarkable antibacterial activity against E. coli, B. subtilis, S. aureus, and Klebsiella pneumoniae. Sandri et al. (2017) synthesized a nanocomposite from halloysite and chitosan oligosaccharides for the treatment of chronic wounds. The researchers investigated the healing capacity of the nanocomposite in murine model and found that the prepared nanocomposite was able to promote skin reepithelization and reorganization.
Immunostimulation
Animal response against the microbial invasion is initiated by the non-specific immune system due to immunostimulants by increasing the defending activity of macrophages and neutrophils. Immunostimulants may bind specifically with the cell surface receptor proteins of phagocytes or lymphocytes that evoke stimulation for the production of immune response compounds, i.e., interferons, interleukin, and complement proteins (Kim and Rajapakse 2005; Kim et al. 2006). COSs have been well documented as immunostimulating agents due to their potential to enhance the migratory activity of macrophages. Oral administration may exert effects on specific cell-mediated immunity. N-acetylated chitohexaose has shown to possess cell proliferating activity on mouse lymphocytes (Wu and Tsai 2004). The study also concluded that the immune-enhancing activity of COS is due to their ability to increase serum levels of IgG and IgM. COS could also activate the macrophages to trigger immunity by enhancing the activity of inducible nitric oxide synthase that results in the accumulation of nitrous oxide and TGF-α (Han et al. 2005). Dou et al. (2007) reported that COS can also regulate the activation of neutrophils, the active phagocytic cells that enhance the production of oxygen and nitrogen intermediates. In the study, researchers showed that COS at concentrations ranging from 50 to 150 μg/ml is capable of reducing phorbol myristate acetate-induced neutrophil activation.
Antioxidant Activity
Oxidation of biomolecules by the reactive oxygen species (ROS), i.e., hydrogen peroxide, hydroxyl radicals, and superoxide anions produced during the normal metabolism results in oxidative stress. Combating oxidative stress is performed by the cell following synthesis of antioxidant agents viz. catalase, superoxide dismutase, glutathione peroxidase, and thioredoxins. Excessive generation of ROS due to the inability of cellular defense mechanisms results in aging, cancer, wrinkle formation, rheumatoid arthritis, and inflammation. In the recent years, ample findings in medicinal research showed usage of N-acetyl COS with MW up to 3 kDa for balancing the oxidative and reductive state inside the body. According to a research conducted by Fernandes et al. (2010), the mixture of COS (MW 3 and 5 kDa) with 80–85% degree of deacetylation is capable of reducing hemolysis and DNA damage by inhibiting hydrogen peroxide and 2,2′-azobis (2-amidinopropane) hydrochloride radicals. Je et al. (2004) reported that COS (90 kDa) prepared from 90% deacetylated chitosan showed notable radical scavenging activity. The scavenging activity of COS is also dependent on the degree of substitution. Increase in the degree of substitution of N-carboxymethyl COS led to decrease in scavenging activity (Sun et al. 2008). Ngo et al. (2011) observed the antioxidant activity of gallete COS showed decreased reactive oxygen species-induced activation of the nuclear transcription factor. Recently, the antioxidant activity of COS has been exploited to enhance flavor stability in beer (Yang et al. 2017). Addition of COS (0.001–0.01%) in stale beer averted the formation of staling compounds and scavenge radicals. Decreased production of 5-hydroxymethylfurfural, trans-2-nonenal, and phenylacetaldehyde by 105, 360, and 27% respectively was also observed upon addition of COS.
Antimicrobial Activity
The antimicrobial activity of COS against different groups of microorganisms, such as bacteria, yeast, and fungi, has received significant attention in the scientific community in recent years. The antimicrobial activity of COS is the consequence of factors like DP, level of deacetylation, type of microorganisms, and physiochemical properties. COS can alter permeability characteristics of the microbial cell membrane and further check the admission of materials or cause leakage of cell constituents that finally leads to the death of bacteria (Chung et al. 2004; Park et al. 2004). Electron microscopic studies of bacterial cells by Choi et al. (2001) also proved that COSs act on the bacterial envelop and kill the organism as the consequence of membrane disruption. Positively charged COSs also inhibit bacterial cell growth by binding to their cell wall. The mechanism that lies behind the growth inhibiting activity of COS depends on the interaction between positively charged amino group (C-2 position) of the glucosamine monomer with negatively charged carboxylic acid group present at bacterial cell surface. This interaction led to formation of polyelectrolyte complexes that act as an impermeable layer around the cell and suppress the metabolic activity of bacteria by blocking nutrient penetration through the cell wall. The charge distribution on the bacterial cell wall also plays a crucial role in the antibacterial activity of COS (Chung et al. 2004). Hydrophilicity and negative charge distribution on the bacterial cell surface have been observed to be closely related. The adsorption of COS to the cell surface is in the order of higher to lower negatively charged Gram-negative bacteria (Gerasimenko et al. 2004; Kim et al. 2003). Chitosan nanoparticles loaded with leaf extract of Ocimum basilicum exhibited antibacterial properties against Gram-positive B. valllsmortis and Gram-negative E. coli. The spherical nanoparticle was in the size range of 135–729 nm (Rasaee et al. 2016). Recently, Luo et al. (2017) investigated the antibacterial potential and cytocompatibility of COS-modified polyurethane membrane. Antibacterial activity of the COS-polyurethane membrane was significantly enhanced against E. coli and S. aureus. The study also concluded that the COS-functionalized polyurethane membrane was more beneficial to the adhesion and proliferation of NH-3T3 cells as compared to the normal polyurethane membrane.
The antifungal potential of COS has been comparatively less investigated. There has been a growing interest in the antifungal activity of COS due to its application in the agriculture sector against phytopathogenic fungi (Rahman et al. 2015). The antifungal activity of COS is mainly owing to their interaction with lipids in the plasma membrane that leads to morphological changes and the cell surface disruption which occurs due to their polycationic nature that allows them to react with negatively charged groups on the fungal surface. The formation of polyelectrolyte complexes between COS and negatively charged groups on the surface of the cell interferes with the normal growth and physiological functions of fungi (Hirano and Nagao 1989; Palma-Guerrero et al. 2009; Park et al. 2008).
Agriculture
The potential of COS in the field of agriculture has been studied tremendously in the last two decades for plant growth and protection. These bioactive oligosaccharides show a wide range of biological activities in plants, and so their use in agriculture may be very promising for the problems such as disease resistance to bactericides, fungicides, and high cost of developing new products (Falcón-Rodríguez et al. 2012). COS at low concentration acts as the plant growth regulator whereas at higher concentration shows the protective activity. COS had also been investigated for inducing innate immunity in plants (Das et al. 2013), enhancing plant physiological responses (Katiyar et al. 2015), mineral accumulation for plant growth (Chatelain et al. 2014), and as a potential disease vaccine (Yin et al. 2010). In the recent years, chitosan-based nanomaterials have been extensively researched for the development of controlled delivery systems for slow and sustainable release of agrochemicals or genetic materials viz. pesticides, fertilizers, herbicides, and micronutrients (Kashyap et al. 2015). Phu et al. (2017) fabricated chitosan nanosilica by the dispersion of nanosilica in oligochitosan solution and investigated its foliar effect on soybean seed yield. Seventeen percent enhancement in the yield was observed with respect to control.
Plant Immunity
Different from animals, plants are sessile and therefore they have developed sophisticated mechanisms to adapt to a range of biotic (fungi, bacteria, and insects) and abiotic (wounding, salinity, drought, salt, and cold) stresses. Plants initiate various defense responses viz. hypersensitive responses, production of phytoalexins, and reinforcement of cell walls to combat stress conditions (Yin et al. 2010). A plant’s immune system defends it from infection by other organisms and abiotic stress in a non-specific manner. Antimicrobial properties and plant innate immunity elicitation activities attribute to the ability of COS in enhancing the plant immunity. Potential of COS as immune-activator depends on several factors viz. molecular weight, DDA, solubility, concentration, positive charge density, chelating capacity, chemical modification, and type of microorganisms (Katiyar et al. 2015). Low-molecular-weight chitosan has been reported to control postharvest diseases of fruits as pear, strawberry, tomato, etc. (Badawy and Rabea 2009). Kulikov et al. (2006) observed the antiviral activity of COS against tobacco mosaic virus. Vander et al. (1998) reported that N-acetyl oligomer (DoP7) stimulated peroxidase activity for lignin biosynthesis. COSs have been known to stimulate other systems, i.e., transduction, cascades, and the elicitor-responsive genes that are involved in resistance of plants to various infections. COSs also play a role as plant defense booster by triggering various morphological and physiological responses. COSs have also been proved to be effective on several plant-disease interactions and can be used in cooperation with other pesticides and biological agents. Yin et al. (2013) showed that innate immunity was triggered by providing resistance to rapeseed against Sclerotinia sclerotiorum. Kheiri et al. (2017) reported the fusarium head blight disease controlling efficiency of the chitosan and chitosan nanoparticles in wheat crop. The chitosan nanoparticles were able to reduce the growth of the disease causative agent Fusarium graminearum up to 77.5% at 5000 ppm concentration. Recently, Dzung et al. (2017) have reported the induction of anthracnose disease resistance (caused by Colletrichum gloeosporioides) on chili fruit through the application of oligochitosan-nanosilica hybrid material. The study suggested that the foliar application of oligochitosan-nanosilca at concentration of 60–60 mg l−1 was most effective to reduce the severity of the disease up to 22.2%.
Plant Growth and Development
COSs also have great potential for improving plant growth and development in the context of germination capacity, root length, plant height, increase in root activity, seedling size, shoot growth, and hastened flowering time (Falcón-Rodríguez et al. 2012). Chatelain et al. (2014) reported the alteration power of COS in root length, mineral accumulation, and shoot biomass. The study showed that the plants have high mineral accumulation capacity when treated with COS at high concentration. Schultze and Kondorosi (1996) suggested the role of lipochitooligosaccharides (LCOS), i.e., a nod signal excreted by Rhizobia to promote the plant growth by inducing the development of symbiotic root nodules in leguminous plants. The plant growth and development properties of LCOS had also been proved by Schwinghamer et al. (2014). The authors showed that chitopentaose containing LCOS may increase the germination by 76%. Vasconcelos (2014) also supported the effect of COS in crop development as well as in phytoremediation. Recently, nanochitin has been reported to improve tobacco stem length, stem girth, leaf number, and leaf area (Zhou et al. 2017). The study reported that nanochitin (0.001%) showed synergistic effects on inhibition of tobacco root rot when mixed with metalaxyl mancozeb and thiophanate methyl fungicides. In another study, Zong et al. (2017) reported the enhancement in cadmium tolerance of Brassica rapa by the exogenous utilization of COS. The study illustrated that the spraying of COS onto leaves resulted in the increase of plant growth and leaf chlorophyll contents whereas decrease in the malondialdehyde and Cd++ levels.
Conclusion
Degradation of chitin and chitosan to produce COS has been studied for years, and a number of reports are available for biological production as well. However, still, the success is rather limited due to the high crystalline index of the parent polymer and variable properties (affinity, stability, and processivity) of the enzyme system employed. Direct fermentation of chitinous waste has also been exploited to get high yields of COS production, yet there is still plenty of room for improvement. Crystallinity of chitin needs to be overcome to improve its bioconversion and increase its commercial value. Employment of mono-enzyme system has not been proved to be a successful one. Synergistic action of dual enzyme systems may be explored to increase the catalytic efficiency of conversion. Concomitant employment of both CBPs and chitinases may provide novel direction towards production of these bioactive oligosaccharides. Scaling up of the production process may generate high yields of versatile polymer.
References
Aam BB, Heggset EB, Norberg AL, Sørlie M, Vårum KM, Eijsink VG (2010) Production of chitooligosaccharides and their potential applications in medicine. Mar Drugs 8:1482–1517
Babashpour S, Aminzadeh S, Farrokhi N, Karkhane A, Haghbeen K (2012) Characterization of a chitinase (Chit62) from Serratia marcescens B4A and its efficacy as a bioshield against plant fungal pathogens. Biochem Genet 50:722–735
Badawy ME, Rabea EI (2009) Potential of the biopolymer chitosan with different molecular weights to control postharvest gray mold of tomato fruit. Postharvest Biol Technol 51:110–117
Chatelain PG, Pintado ME, Vasconcelos MW (2014) Evaluation of chitooligosaccharide application on mineral accumulation and plant growth in Phaseolus vulgaris. Plant Sci 215:134–140
Choi BK, Kim KY, Yoo YJ, Oh SJ, Choi JH, Kim CY (2001) In vitro antimicrobial activity of a chitooligosaccharide mixture against Actinobacillu sactinomycetemcomitans and Streptococcus mutans. Int J Antimicrob Agents 18:553–557
Chung YC, Su YP, Chen CC, Jia G, Wang H, Wu JG, Lin JG (2004) Relationship between antibacterial activity of chitosan and surface characteristics of cell wall. Acta Pharmacol Sin 25:932–936
Das SN, Madhuprakash J, Sarma P, Purushotham P, Suma K, Manjeet K et al (2013) Biotechnological approaches for field applications of chitooligosaccharides (COS) to induce innate immunity in plants. Crit Rev Biotechnol 35:29–43
de Assis CF, Araújo NK, Pagnoncelli MGB, da Silva Pedrini MR, de Macedo GR, dos Santos ES (2010) Chitooligosaccharides enzymatic production by Metarhizium anisopliae. Bioprocess Biosyst Eng 33:893–899
Dou J, Tan C, Du Y, Bai X, Wang K, Ma X (2007) Effects of chitooligosaccharides on rabbit neutrophils in vitro. Carbohydr Polym 69:209–213
Dutta PK, Dutta J, Tripathi V (2004) Chitin and chitosan: chemistry, properties and applications. J Sci Ind Res 63:20–31
Dzung PD, Hung LT, Ngoc LS, Hiet HD, Van Le B, Thang NT, Van Phu D, Duy NN, Hien NQ (2017) Induction of anthracnose disease resistance on chili fruit by treatment of oligochitosan—nanosilica hybrid material. Agric Sci 8(10):1105–1113
Elgadir MA, Uddin MS, Ferdosh S, Adam A, Chowdhury AJK, Sarker MZI (2015) Impact of chitosan composites and chitosan nanoparticle composites on various drug delivery systems: a review. J Food Drug Anal 23(4):619–629
Elieh-Ali-Komi D, Hamblin MR (2016) Chitin and chitosan: production and application of versatile biomedical nanomaterials. Int J Adv Res 4(3):411–427
Falcón-Rodríguez A, Costales D, Rogers H, Diosdado E, González S, Cabrera G, et al (2012) Practical use of oligosaccharins in agriculture. In: Saa Silva S, et al. (eds) Developing application for oligosaccharins in agriculture. The 1st World Congress on the use of Biostimulants in Agriculture, vol 1009. Strasbourg Congress Centre, France, pp 195–212
Fernandes JC, Eaton P, Nascimento H, Gião MS, Ramos ÓS, Belo L, Santos-Silva A, Pintado ME, Malcata FX (2010) Antioxidant activity of chitooligosaccharides upon two biological systems: erythrocytes and bacteriophages. Carbohydr Polym 79:1101–1106
Fukamizo T (2000) Chitinolytic enzymes catalysis, substrate binding, and their application. Curr Protein Pept Sci 1:105–124
Gao XA, Ju WT, Jung WJ, Park RD (2008) Purification and characterization of chitosanase from Bacillus cereus D-11. Carbohydr Polym 72:513–520
Gerasimenko D, Avdienko I, Bannikova G, Zueva OY, Varlamov V (2004) Antibacterial effects of water-soluble low-molecular-weight chitosans on different microorganisms. Appl Biochem Microbiol 40:253–257
Gomathi P, Ragupathy D, Choi JH, Yeum JH, Lee SC, Kim JC, Do Ghim H (2011) Fabrication of novel chitosan nanofiber/gold nanoparticles composite towards improved performance for a cholesterol sensor. Sensors Actuators B Chem 153(1):44–49
Gomathi T, Sudha PN, Florence JAK, Venkatesan J, Anil S (2017) Fabrication of letrozole formulation using chitosan nanoparticles through ionic gelation method. Int J Biol Macromol 104:1820–1832
Hamed I, Özogul F, Regenstein JM (2016) Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): a review. Trends Food Sci Technol 48:40–50
Hamid R, Khan MA, Ahmad M, Ahmad MM, Abdin MZ, Musarrat J et al (2013) Chitinases: an update. J Pharm Bioallied Sci 5:21–29
Han Y, Zhao L, Yu Z, Feng J, Yu Q (2005) Role of mannose receptor in oligochitosan-mediated stimulation of macrophage function. Int Immunopharmacol 5:1533–1542
Harman G, Hayes C, Lorito M, Broadway R, Di Pietro A, Peterbauer C et al (1993) Chitinolytic enzymes of Trichoderma harzianum: purification of chitobiosidase and endochitinase. Phytopathology 83:313–318
Hirano S, Nagao N (1989) Effects of chitosan, pectic acid, lysozyme, and chitinase on the growth of several phytopathogens. Agric Biol Chem 53:3065–3066
Il'ina A, Zueva OY, Lopatin S, Varlamov V (2004) Enzymatic hydrolysis of α-chitin. Appl Biochem Microbiol 40:35–38
Je JY, Park PJ, Kim SK (2004) Free radical scavenging properties of hetero-chitooligosaccharides using an ESR spectroscopy. Food Chem Toxicol 42:381–387
Jeon YJ, Kim SK (2002) Antitumor activity of chitosan oligosaccharides produced in ultrafiltration membrane reactor system. J Microbiol Biotechnol 12:503–507
Jung WJ, Park RD (2014) Bioproduction of chitooligosaccharides: present and perspectives. Mar Drugs 12:5328–5356
Kashyap PL, Xiang X, Heiden P (2015) Chitosan nanoparticle based delivery systems for sustainable agriculture. Int J Biol Macromol 77:36–51
Katiyar D, Hemantaranjan A, Singh B (2015) Chitosan as a promising natural compound to enhance potential physiological responses in plant: a review. Indian J Plant Phys 20:1–9
Kheiri A, Moosawi Jorf SA, Malihipour A, Saremi H, Nikkhah M (2017) Synthesis and characterization of chitosan nanoparticles and chitosan effect on Fusarium head blight and oxidative activity in wheat. Int J Biol Macromol 102:526–538
Khoushab F, Yamabhai M (2010) Chitin research revisited. Mar Drugs 8(7):1988–2012
Khoushab F, Jaruseranee N, Tanthanuch W, Yamabhai M (2012) Formation of chitin-based nanomaterials using a chitin-binding peptide selected by phage-display. Int J Biol Macromol 50(5):1267–1274
Kim SK, Rajapakse N (2005) Enzymatic production and biological activities of chitosan oligosaccharides (COS): a review. Carbohydr Polym 62:357–368
Kim JY, Lee JK, Lee TS, Park WH (2003) Synthesis of chitooligosaccharide derivative with quaternary ammonium group and its antimicrobial activity against Streptococcus mutans. Int J Biol Macromol 32:23–27
Kim HM, Hong SH, Yoo SJ, Baek KS, Jeon YJ, Choung SY (2006) Differential effects of chitooligosaccharides on serum cytokine levels in aged subjects. J Med Food 9:427–430
Kono H, Teshirogi T (2015) Cyclodextrin-grafted chitosan hydrogels for controlled drug delivery. Int J Biol Macromol 72:299–308
Kulikov S, Chirkov S, Il’ina A, Lopatin S, Varlamov V (2006) Effect of the molecular weight of chitosan on its antiviral activity in plants. Appl Biochem Microbiol 42:200–203
Kumar M, Brar A, Vivekanand V, Pareek N (2017) Production of chitinase from thermophilic Humicola grisea and its application in production of bioactive chitooligosaccharides. Int J Biol Macromol. https://doi.org/10.1016/j.ijbiomac.2017.04.100
Kumari A, Yadav SK, Yadav SC (2010) Biodegradable polymeric nanoparticles based drug delivery systems. Colloids Surf B Biointerfaces 75(1):1–18
Liang T-W, Hsieh J-L, Wang S-L (2012) Production and purification of a protease, a chitosanase, and chitin oligosaccharides by Bacillus cereus TKU022 fermentation. Carbohydr Res 362:38–46.
Li ZW, Li CW, Wang Q, Shi SJ, Hu M, Zhang Q, Dang JZ (2017) The cellular and molecular mechanisms underlying silver nanoparticle/chitosan oligosaccharide/poly (vinyl alcohol) nanofiber-mediated wound healing. J Biomed Nanotechnol 13(1):17–34
Lodhi G, Kim YS, Hwang JW, Kim SK, Jeon YJ, Je J et al (2014) Chitooligosaccharide and its derivatives: preparation and biological applications. Biomed Res Int:1:1–1:113
Lombard V, Ramulu HG, Drula E, Coutinho PM, Henrissat B (2014) The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res 42:D490–D495
Luo Y, Deng L, Deng Q-J, Wen L (2016) Comparative study of the chitooligosaccharides effect on the proliferation inhibition and radio-sensitization of three types of human gastric cell line. Asian Pac J Trop Med 9:601–605
Luo C, Liu W, Luo B, Tian J, Wen W, Liu M, Zhou C (2017) Antibacterial activity and cytocompatibility of chitooligosaccharide-modified polyurethane membrane via polydopamine adhesive layer. Carbohydr Polym 156:235–243
Ma L, Li Y, Yu C, Wang Y, Li X, Li N, Chen Q, Bu N (2012) Alleviation of exogenous oligochitosan on wheat seedlings growth under salt stress. Protoplasma 249:393–399
Mahata M, Shinya S, Masaki E, Yamamoto T, Ohnuma T, Brzezinski R, Mazumder TK, Yamashita K, Narihiro K, Fukamizo T (2014) Production of chitooligosaccharides from Rhizopus oligosporus NRRL2710 cells by chitosanase digestion. Carbohydr Res 383:27–33
Manivasagan P, Oh J (2016) Marine polysaccharide-based nanomaterials as a novel source of nanobiotechnological applications. Int J Biol Macromol 82:315–327
Manivasagan P, Bharathiraja S, Bui NQ, Lim IG, Oh J (2016) Paclitaxel-loaded chitosan oligosaccharide-stabilized gold nanoparticles as novel agents for drug delivery and photoacoustic imaging of cancer cells. Int J Pharm 511(1):367–379
Manivasagan P, Bui NQ, Bharathiraja S, Moorthy MS, Oh YO, Song K, Seo H, Yoon M, Oh J (2017) Multifunctional biocompatible chitosan-polypyrrole nanocomposites as novel agents for photoacoustic imaging-guided photothermal ablation of cancer. Sci Rep 7:43593
Manjeet K, Purushotham P, Neeraja C, Podile AR (2013) Bacterial chitin binding proteins show differential substrate binding and synergy with chitinases. Microbiol Res 168:461–468
Mogoşanu GD, Grumezescu AM, Bejenaru C, Bejenaru LE (2016) Polymeric protective agents for nanoparticles in drug delivery and targeting. Int J Pharm 510(2):419–429
Mourya VK, Inamdar NN, Choudhari YM (2011) Chitooligosaccharides: synthesis, characterization and applications. Polym Sci Ser A 53:583–612
Muanprasat C, Chatsudthipong V (2017) Chitosan oligosaccharide: biological activities and potential therapeutic applications. Pharmacol Ther 170:80–97
Ngo DH, Qian ZJ, Vo TS, Ryu B, Ngo DN, Kim SK (2011) Antioxidant activity of gallate-chitooligosaccharides in mouse macrophage RAW264.7 cells. Carbhydr Polym 84:1282–1288
Nguyen AD, Huang CC, Liang TW, Pan PS, Wang SL (2014) Production and purification of a fungal chitosanase and chitooligomers from Penicillium janthinellum D4 and discovery of the enzyme activators. Carbohydr Polym 108:331–337
Nidheesh T, Pal GK, Suresh P (2015) Chitooligomers preparation by chitosanase produced under solid state fermentation using shrimp by-products as substrate. Carbohydr Polym 121:1–9
Olicón-Hernández DR, Zepeda Giraud LF, Guerra-Sánchez G (2017) Current applications of chitosan and chito-oligosaccharides. A review. J Drug Des Res 4(2):1039
Palma-Guerrero J, Huang IC, Jansson HB, Salinas J, Lopez-Llorca L, Read N (2009) Chitosan permeabilizes the plasma membrane and kills cells of Neurospora crassa in an energy dependent manner. Fungal Genet Biol 46:585–594
Pareek N, Vivekanand V, Dwivedi P, Singh RP (2011) Penicillium oxalicum SAE M-51: a mutagenised strain for enhanced production of chitin deacetylase for bioconversion to chitosan. New Biotechnol 28:118–124
Park PJ, Je JY, Byun HG, Moon SH, Kim SK (2004) Antimicrobial activity of hetero-chitosans and their oligosaccharides with different molecular weights. J Microbiol Biotechnol 14:317–323
Park Y, Kim MH, Park SC, Cheong H, Jang MK, Nah JW, Hahm KS (2008) Investigation of the antifungal activity and mechanism of action of LMWS-chitosan. J Microbiol Biotechnol 18:1729–1734
Pechsrichuang P, Yoohat K, Yamabhai M (2013) Production of recombinant Bacillus subtilis chitosanase, suitable for biosynthesis of chitosan-oligosaccharides. Bioresour Technol 127:407–414
Perumal V, Manickam T, Bang KS, Velmurugan P, Oh BT (2016) Antidiabetic potential of bioactive molecules coated chitosan nanoparticles in experimental rats. Int J Biol Macromol 92:63–69
Phu DV, Du BD, Tuan LNA, Tam HV, Hien NQ (2017) Preparation and foliar application of oligochitosan-nanosilica on the enhancement of soybean seed yield. Int J Environ Agric Biotechnol 2(1):421–428
Pillai C, Paul W, Sharma CP (2009) Chitin and chitosan polymers: chemistry, solubility and fiber formation. Prog Polym Sci 34:641–678
Prashanth KH, Tharanathan R (2005) Depolymerized products of chitosan as potent inhibitors of tumor-induced angiogenesis. Biochim Biophys Acta Gen Subj 1722:22–29
Rahman MH, Hjeljord LG, Aam BB, Sørlie M, Tronsmo A (2015) Antifungal effect of chito-oligosaccharides with different degrees of polymerization. Eur J Plant Pathol 141:147–158
Rai M, Ingle AP, Gupta I, Brandelli A (2015) Bioactivity of noble metal nanoparticles decorated with biopolymers and their application in drug delivery. Int J Pharm 496(2):159–172
Ramezanzade L, Hosseini SF, Nikkhah M (2017) Biopolymer-coated nanoliposomes as carriers of rainbow trout skin-derived antioxidant peptides. Food Chem 234:220–229
Rasaee I, Ghannadnia M, Honari H (2016) Antibacterial properties of biologically formed chitosan nanoparticles using aqueous leaf extract of Ocimum basilicum. Nanomed J 3(4):240–247
Rinaudo M (2006) Chitin and chitosan: properties and applications. Prog Polym Sci 31:603–632
Sahai A, Manocha M (1993) Chitinases of fungi and plants: their in morphogenesis and host-parasite. FEMS Microbiol Rev 11:317–338
Sánchez Á, Mengíbar M, Rivera-Rodríguez G, Moerchbacher B, Acosta N, Heras A (2017) The effect of preparation processes on the physicochemical characteristics and antibacterial activity of chitooligosaccharides. Carbohydr Polym 157:251–257
Sandri G, Aguzzi C, Rossi S, Bonferoni MC, Bruni G, Boselli C, Cornaglia AI, Riva F, Viseras C, Caramella C, Ferrari F (2017) Halloysite and chitosan oligosaccharide nanocomposite for wound healing. Acta Biomater 57:216–224
Sathiyaseelan A, Shajahan A, Kalaichelvan PT, Kaviyarasan V (2017) Fungal chitosan based nanocomposites sponges—an alternative medicine for wound dressing. Int J Biol Macromol 104:1905–1915
Schultze M, Kondorosi Á (1996) The role of lipochitooligosaccharides in root nodule organogenesis and plant cell growth. Curr Opin Genet Dev 6:631–638
Schwinghamer T, Souleimanov A, Dutilleul P, Smith D (2014) The plant growth regulator lipo-chitooligosaccharide (LCO) enhances the germination of canola (Brassica napus [L.]). J Plant Growth Regul 34:183–195
Shen KT, Chen MH, Chan HY, Jeng JH, Wang YJ (2009) Inhibitory effects of chitooligosaccharides on tumor growth and metastasis. Food Chem Toxicol 47:1864–1871
Shukla SK, Mishra AK, Arotiba OA, Mamba BB (2013) Chitosan-based nanomaterials: a state-of-the-art review. Int J Biol Macromol 59:46–58
Songsiriritthigul C, Pesatcha P, Eijsink VG, Yamabhai M (2009) Directed evolution of a Bacillus chitinase. Biotechnol J 4:501–509
Songsiriritthigul C, Lapboonrueng S, Pechsrichuang P, Pesatcha P, Yamabhai M (2010) Expression and characterization of Bacillus licheniformis chitinase (ChiA), suitable for bioconversion of chitin waste. Bioresour Technol 101(11):4096–4103
Su PC, Hsueh WC, Chang WS, Chen PT (2017) Enhancement of chitosanase secretion by Bacillus subtilis for production of chitosan oligosaccharides. J Taiwan Inst Chem Eng 79:49–54
Subhapradha N, Shanmugam A (2017) Fabrication of β-chitosan nanoparticles and its anticancer potential against human hepatoma cells. Int J Biol Macromol 94:194–201
Sun T, Yao Q, Zhou D, Mao F (2008) Antioxidant activity of N-carboxymethyl chitosan oligosaccharides. Bioorg Med Chem Lett 18:5774–5776
Suzuki K, Sugawara N, Suzuki M, Uchiyama T, Katouno F, Nikaidou N et al (2002) Chitinases A, B, and C1 of Serratia marcescens 2170 produced by recombinant Escherichia coli: enzymatic properties and synergism on chitin degradation. Biosci Biotechnol Biochem 66:1075–1083
Tharanathan RN, Kittur FS (2003) Chitin—the undisputed biomolecule of great potential. Crit Rev Food Sci Nutr 43:61–87
Ueda M, Shioyama T, Nakadoi K, Nakazawa M, Sakamoto T, Iwamoto T, Sakaguchi M (2017) Cloning and expression of a chitinase gene from Eisenia fetida. Int J Biol Macromol 104:1648–1655
Vaaje-Kolstad G, Horn SJ, van Aalten DM, Synstad B, Eijsink VG (2005a) The non-catalytic chitin-binding protein CBP21 from Serratia marcescens is essential for chitin degradation. J Biol Chem 280:28492–28497
Vaaje-Kolstad G, Houston DR, Riemen AH, Eijsink VG, van Aalten DM (2005b) Crystal structure and binding properties of the Serratia marcescens chitin-binding protein CBP21. J Biol Chem 280:11313–11319
Van Aalten D, Komander D, Synstad B, Gåseidnes S, Peter M, Eijsink V (2001) Structural insights into the catalytic mechanism of a family 18 exo-chitinase. Proc Natl Acad Sci 98:8979–8984
Vander P, Vårum KM, Domard A, El Gueddari NE, Moerschbacher BM (1998) Comparison of the ability of partially N-acetylated chitosans and chitooligosaccharides to elicit resistance reactions in wheat leaves. Plant Physiol 118:1353–1359
Vasconcelos MW (2014) Chitosan and chitooligosaccharide utilization in phytoremediation and biofortification programs: current knowledge and future perspectives. Front Plant Sci 5:616–620
Viens P, Lacombe-Harvey MÈ, Brzezinski R (2015) Chitosanases from family 46 of glycoside hydrolases: from proteins to phenotypes. Mar Drugs 13:6566–6587
Waghmare SR, Ghosh JS (2010) Chitobiose production by using a novel thermostable chitinase from Bacillus licheniformis strain JS isolated from a mushroom bed. Carbohydr Res 345:2630–2635
Wang Z, Zheng L, Yang S, Niu R, Chu E, Lin X (2007) N-acetylchitooligosaccharide is a potent angiogenic inhibitor both in vivo and in vitro. Biochem Biophys Res Commun 357:26–31
Wang Y, Zhang C, Zhang Q, Li P (2011) Composite electrospun nanomembranes of fish scale collagen peptides/chito-oligosaccharides: antibacterial properties and potential for wound dressing. Intl J Nanomed 6:667–676
Wang SL, Liu CP, Liang TW (2012) Fermented and enzymatic production of chitin/chitosan oligosaccharides by extracellular chitinases from Bacillus cereus TKU027. Carbohydr Polym 90:1305–1313
Wang T, Hou J, Su C, Zhao L, Shi Y (2017) Hyaluronic acid-coated chitosan nanoparticles induce ROS-mediated tumor cell apoptosis and enhance antitumor efficiency by targeted drug delivery via CD44. J Nanobiotechnology 15(1):7
Woo CJ, Park HD (2003) An extracellular Bacillus sp. chitinase for the production of chitotriose as a major chitinolytic product. Biotechnol Lett 25:409–412
Wu GJ, Tsai GJ (2004) Cellulase degradation of shrimp chitosan for the preparation of a water-soluble hydrolysate with immunoactivity. Fish Sci 70:1113–1120
Wu JL, He XY, Jiang PY, Gong MQ, Zhuo RX, Cheng SX (2016) Biotinylated carboxymethyl chitosan/CaCO 3 hybrid nanoparticles for targeted drug delivery to overcome tumor drug resistance. RSC Adv 6(73):69083–69093
Xia W, Liu P, Zhang J, Chen J (2011) Biological activities of chitosan and chitooligosaccharides. Food Hydrocoll 25:170–179
Xiong C, Wu H, Wei P, Pan M, Tuo Y, Kusakabe I et al (2009) Potent angiogenic inhibition effects of deacetylated chitohexaose separated from chitooligosaccharides and its mechanism of action in vitro. Carbohydr Res 344:1975–1983
Xu Q, Dou J, Wei P, Tan C, Yun X, Wu Y et al (2008) Chitooligosaccharides induce apoptosis of human hepatocellular carcinoma cells via up-regulation of Bax. Carbohydr Polym 71:509–514
Yang S, Fu X, Yan Q, Guo Y, Liu Z, Jiang Z (2016) Cloning, expression, purification and application of a novel chitinase from a thermophilic marine bacterium Paenibacillus barengoltzii. Food Chem 192:1041–1048
Yang F, Luan B, Sun Z, Yang C, Yu Z, Li X (2017) Application of chitooligosaccharides as antioxidants in beer to improve the flavour stability by protecting against beer staling during storage. Biotechnol Lett 39(2):305–310
Yano S, Rattanakit N, Honda A, Noda Y, Wakayama M, Plikomol A et al (2008) Purification and characterization of chitinase A of Streptomyces cyaneus SP-27: an enzyme participates in protoplast formation from Schizophyllum commune mycelia. Biosci Biotechnol Biochem 72:54–61
Yin H, Zhao X, Du Y (2010) Oligochitosan: a plant diseases vaccine: a review. Carbohydr Polym 82:1–8
Yin H, Li Y, Zhang HY, Wang WX, Lu H, Grevsen K et al (2013) Chitosan oligosaccharides triggered innate immunity contributes to oilseed rape resistance against Sclerotinia Sclerotiorum. Int J Plant Sci 174:722–732
You Y, Park WH, Ko BM, Min BM (2004) Effects of PVA sponge containing chitooligosaccharide in the early stage of wound healing. J Mater Sci Mater Med 15:297–301
Youssef AS, El-Sherif MF, Hassan MA, Hassan HM, El-Aassar SA (2013) Purification and properties of chitinase enzyme produced by Bacillus licheniformis. J Pure Appl Microbiol 7:179–188
Zhang Y, Zhou X, Ji L, Du X, Sang Q, Chen F (2017) Enzymatic single-step preparation and antioxidant activity of hetero-chitooligosaccharides using non-pretreated housefly larvae powder. Carbohydr Polym 172:113–119
Zhou Y, Jiang S, Jiao Y, Wang H (2017) Synergistic effects of nanochitin on inhibition of tobacco root rot disease. Int J Biol Macromol 99:205–212
Zong H, Li K, Liu S, Song L, Xing R, Chen X, Li P (2017) Improvement in cadmium tolerance of edible rape (Brassica rapa L.) with exogenous application of chitooligosaccharide. Chemosphere 181:92–100
Funding
The authors are thankful to Department of Biotechnology and Department of Science and Technology, Government of India, for financial support.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of Interest
The authors declare that they have no conflict of interest.
Rights and permissions
About this article

Cite this article
Kumar, M., Brar, A., Vivekanand, V. et al. Bioconversion of Chitin to Bioactive Chitooligosaccharides: Amelioration and Coastal Pollution Reduction by Microbial Resources. Mar Biotechnol 20, 269–281 (2018). https://doi.org/10.1007/s10126-018-9812-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10126-018-9812-x