
Abstract
Innovative behavior is considered one of the main factors facilitating the adaptation of animals to urban life. However, the relationship between urbanization and innovativeness is equivocal, perhaps reflecting aspects of urban environments that influence differently the behavioural traits underlying the occurrence of an innovation. In this work, we analysed the variation in innovative problem-solving performance between urban and rural individuals of the Caracara Chimango (Milvago chimango), with the goal of determining which behavioural trait (or combination) most explained such variation. We found that urban raptors outperformed rural ones in their solving speed and solving level (number of solutions) with a multiaccess box. They also showed more persistence, motor flexibility and diversity, as well as higher effectiveness in their solving attempts than rural chimangos. Sex was not an important factor. Urban chimangos showed less neophobia and spent more time exploring the box than rural birds during the initial habituation period, which probably determined the amount of information about the system that each individual had at the beginning of first problem solving trial. This difference in novelty response both directly and indirectly, through its relationship with persistence, motor flexibility and proportion of effective attempts, explained variability in solving performance. All individuals showed a decrease in solving latency, and an increase in solving level with experience, indicating that learning occurred in both raptor groups. This improvement occurred in parallel with changes in the afore-mentioned traits, though the pattern of improvement differed between urban and rural chimangos. We suggest that the characteristics of urban areas modulate the novelty response of chimangos, along with other correlated non-cognitive behavioural traits, which act in combination to increase the chances that novel problems could be quickly solved, and the resulting new behaviours established in city populations of this species.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Behavioural innovation, defined as the process that results in the invention of new behaviours, or the use of pre-existing ones in novel contexts (Reader and Laland 2003), is considered an expression of behavioural flexibility and, so, is a rich source of phenotypic plasticity in animals (Sol et al. 2005; Lefebvre 2011). This process is assumed to be a fundamental attribute facilitating animals’ adjustment to novel or changing environments (Griffin and Guez 2014; Reader et al. 2016; Griffin 2016), allowing the invasion of new habitats, the exploitation of novel resources, and coping with rapid habitat modifications (Reader and Laland 2003; Ramsey et al. 2007). When innovating, animals may enter novel selection landscapes which can, in turn, generate new innovation opportunities, thus enhancing the probability of adaptive radiation (West-Eberhard 2003). The ability to innovate has also been linked to reproductive life-history variation and competitive ability (Cole and Quinn 2011; Cole et al. 2012; Cauchard et al. 2013), and thus may have important fitness implications.
Since innovations cannot be easily observed in nature, their study requires an experimental condition that allows animals to express their potential to innovate. In this sense, the novel problem-solving paradigm (or innovative problem-solving) provides an ecologically meaningful assay for measuring the natural proclivity of animals to innovate (Griffin and Guez 2014). Typically, this test involves presenting individuals with an extractive foraging task that needs to be solved to reach a food reward, though also individuals may be required to interact with objects to gain access to others rewards, such as their nest and chicks (Cauchard et al 2013). Innovative problem-solving tasks have enabled a thorough investigation of the mechanisms and factors that contribute to interindividual variation in innovation propensity (reviewed in Griffin and Guez 2014, 2016).
An important source of interindividual variability in innovative problem-solving is the environment. In this sense, the benefits of innovative behaviours may vary between habitat types, and selection may favour an innovative phenotype in relatively challenging environments more strongly. In this context, the ability to innovate, along with the behavioural traits that promote innovativeness, such as low neophobia and high exploratory behaviour, are considered key attributes associated with the successful adjustment of animals to urbanization (Sol et al. 2013, 2020; Griffin 2016). This is because urban animals are exposed to several novel or variable stimuli, such as fragmented landscapes, noise and light pollution, disturbance by domestic animals and humans, and novel food (e.g., food inside human-made packages), or novel sources of it (e.g., garbage) (Sol et al. 2013, 2020). Indeed, individuals from more urbanized habitats were found to be more successful in solving innovative problem-solving tasks in several avian species (Liker and Bokony 2009; Sol et al. 2011; Audet et al. 2016; Kozlovsky et al. 2017). Still, the relationship between urbanization and innovativeness is equivocal and can be context and species specific (Griffin et al. 2017), perhaps reflecting aspects of urban environments that could differentially influence the behavioural traits underlying innovation.
In this context, several studies have been performed with the objective to elucidate whether interindividual differences in problem-solving performance are related to cognitive or motor skills, a particular motivational state, the way animals deal with novel situations, or a combination of all these characteristics. For instance, Sol et al. (2012) showed that in common mynas the proximal factors that determine innovation included both personality traits, such as neophobia, and more state-dependent variable effects, such as motivation. Similarly, van Horik and Madden (2016) working with pheasant chicks, observed that problem solving success was mediated by differences between individuals in their latencies to interact with the test apparatus, and not by variation in cognitive abilities. Another factor with a central role in the innovation process is the diversity in the species’ motor repertoire (Greenberg 2003; Griffin et al. 2014; Tebbich et al. 2016). A more diverse motor repertoire can result in a greater diversity of ways that a physical problem could be handled, which, in turn, could increase the chance that a successful interaction, one that would lead to the solution of the problem, will occur. Indeed, several works in both mammals and birds have demonstrated that variation in motor diversity is central to explaining why some individuals are more likely to innovate than others (Benson-Amram and Holekamp 2012; Thornton and Samson 2012; Griffin et al. 2014; see other references in Guez and Griffin 2016). Recently, a study comparing several bird species in field experiments, revealed that a more balanced distribution of expression of motor actions was more important to innovative foraging than was the total number of motor actions (Diquelou et al. 2016; Guez and Griffin 2016).
Moreover, persistence, a measure of task-directed engagement, can also accelerate solving speed. That is, a more persistent interaction with a specific problem (i.e., novel food container) would increase an animal´s chances of finding a way to solve it. Persistence is a personality trait, which is considered closely related to explorative behaviour (as part of the same behavioural syndrome) in non-human animals (Massen et al. 2013). It has been measured in diverse ways, such as the time an individual spends trying to solve the problem of reaching a desired reward (work time, e.g., in hyaenas: Benson-Amram and Holekamp 2012; Benson-Amram et al. 2013; and racoons: Daniels et al. 2019); the number of times individuals contact the test apparatus (Sol et al 2011), or the rate of contacts (e.g., in chimango caracaras: Biondi et al. 2010, and grey squirrels: Chow et al. 2016). Moreover, some researchers have used the contacting time or the number of contacts made with the functional or relevant parts of the test apparatus (i.e., areas with lids) divided by the total contacting time or the total number of contacts made, respectively, during the problem-solving test (e.g., great tits: Cauchard et al. 2013, meerkats: Thornton and Samson 2012). This last way to measure persistence may also reflect the effectivity of the individuals’ solving attempts, highlighting the difference between an animal being persistent with the right behavioural actions and being just persistent, sometimes in an inaccurate way (Chow et al 2016).
The Chimango Caracara, Milvago chimango (hereafter chimango) is a small/medium (250–350 g) Neotropical raptor that lives in a wide range of habitats, usually associated with diverse anthropic activities (White et al. 1994; Ferguson-Lees and Christie 2001). It is a highly opportunistic and generalist bird, preying on arthropods, small mammals, birds, reptiles, amphibians and carrion (Biondi et al. 2005). In urban settings, it feeds mainly on urban prey and anthropogenic organic waste (White et al. 1994; Biondi et al. 2008). Previous studies performed with this species revealed a remarkable capacity for solving a novel extractive feeding problem and transmitting this ability to conspecifics via social learning. This species has also been shown to have a high tendency to explore and play with novel objects without an immediate food reward, and low neophobia compared to other generalist bird species (Biondi et al. 2008, 2010, 2013, 2015; Guido et al. 2017). All of these traits reflect the characteristic behavioural plasticity of this species. Moreover, this is the only raptor species whose abundance is positively associated with the level of anthropogenic disturbance in Argentina (Carrete et al. 2009), a trend also observed for other generalist raptors in Latin American cities (Ferrer-Sánchez and Rodríguez-Estrella 2015). Recently, the novel problem-solving capacity of chimangos has been compared between periurban and rural areas (Solaro and Sarasola 2019). As was expected for a highly generalist species, these authors observed higher problem-solving success in periurban birds. However, innovative problem-solving ability is a composite trait driven by a range of diverse behavioural characteristics, so to understand why this difference occurs we need experimental studies to unpack the behavioural traits underlying innovation and problem-solving, evaluate how the characteristics of urban habitats influence the expression of these traits, and examine the resulting effect on the innovative problem-solving capacity of this raptor species.
Here, we explore the influence of neophobia, exploratory behaviour, persistence, and motor diversity and flexibility on the interindividual and temporal variation in the innovative problem-solving performance, of chimangos from two areas with contrasting urbanization levels. In a recent study, we have already observed that chimangos from the city were less neophobic and showed higher exploratory behavior and boldness than rural chimangos (Biondi et al. 2020). These findings lead us to predict an effect of urbanization on problem-solving capacity mediated by the neophobia level and exploratory behavior of individuals. Furthermore, we expect to find higher solving persistence in urban than rural birds, as urban areas are associated with high exploratory behavior, low predatory risk and high tolerance to human disturbances, which could promote a more exploratory and persistent behavior toward a novel problem. Moreover, considering that urban chimangos in particular, may frequently contact and manipulate different non-natural objects, such as garbage bags or man-made food packages, when looking for food (Biondi et al. 2015), we could expect a high flexibility in their motor actions. That is, urban birds may use a greater diversity of different behavioural techniques during the exploration of novel situations, but also switch these techniques more frequently. Consequently, as is the case for neophobia and exploration, the predicted higher persistence and motor skills in urban birds, compared to rural conspecifics, can also underline the possible differences in the innovative problem-solving performance between chimangos of these two habitats.
Methods
The study area corresponded to urban and rural habitats in the southwestern Pampas region of Argentina. The urban habitat was represented by capture sites within Mar del Plata City, the largest coastal city of Argentina, holding > 619,000 inhabitants throughout the year, and receiving c. 3 million tourists during the summer (EMTUR, 2016). The rural habitat corresponded to two private lands with agriculture and livestock activities, located approximately 30 km north from Mar del Plata city. Twenty-four adult individuals of M. chimango were captured (12 from the city and 12 from rural areas) with baited walk-in traps (Bloom 1987) between May and July of 2017 (non-breeding period). Birds were weighed, and a blood sample was taken from the brachial vein to perform molecular sexing (Fridolfsson and Ellegren 1999), analysis that resulted in 8 females and 4 males from both urban and rural habitats. Immediately after capture, birds were housed in individual outdoor aviaries (1.5 × 1.5 × 1.2 m) following housing and care conditions described by Bloom (1987) and Aprile and Bertonatti (1996). Aviaries were visually isolated from one another by black synthetic fabric, ensuring that individuals performed on their own, without social influences (Biondi et al. 2008, 2010). Birds were given at least a 5-day period to become acclimated to captivity and to the continuous presence of the researcher in charge of the experiments. During this period, we fed the birds once a day between 9 and 10 am from a dish containing beef meat, and provided water ad libitum. Individuals were considered to have acclimated to captivity when they were comfortable enough to feed shortly after food presentation (Biondi et al. 2008, 2010), a period that ranged from 3 to 5 days for the individuals tested in this study. During all tests the subjects were video-recorded with a Sony Hdr-cx440 camcorder placed at 10 m from the aviaries. Videos were analysed by continuous recording by GF. At the end of the experiments, all individuals were identified with a plastic ring on their tarsus and then released at their capture sites at the end of the experimental tests. In this way, we prevented the use of the same individuals in subsequent experimental bouts. The captivity period was not longer than 12 days (range: 9–12).
Problem-solving test
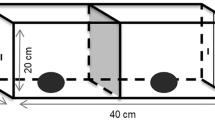
All trials were conducted between 9 to 11 am, and we altered the order of the individual tested between trials. Test subjects were presented with a Plexiglass box filled with small pieces of beef (5 g each), placed in a central spot of the aviary. The orientation of the box´s sides with respect to the location of the perches was kept consistent, during all trials. This same apparatus and experimental protocol were already published in Biondi et al. (2008, 2010). Briefly, the box has four isolated sections containing pieces of food. Each section can be opened by lifting, pushing, pulling or sliding the lids, respectively (Fig. 1). To habituate the subjects to the experimental apparatus before the test and simultaneously record their initial reactions to the novel test apparatus (i.e., neophobia, exploration), we first presented the box with all the lids open and the food accessible during a single 15-min trial (initial exploratory period or D0). During this trial we recorded the feeding latency as the time elapsed from the box presentation until the bird first retrieved a piece of food. This latency represented a measure of neophobia to the box (Biondi et al. 2010). We also recorded the number of food pieces extracted from the test apparatus and the exploration time, as the time each individual spent exploring the box. We considered that individuals were exploring the box when they contacted it or closely looked inside it (less than 5 cm from the box). This activity could last even after the bird consumed all of the available pieces of food.

Diagram of the clear Plexiglas box used for the innovative problem-solving test. Black dots represent the pieces of meat given to the subjects in each experimental test, and the arrows indicate the principal movement direction necessary to open each box lid
Twenty-four hours after this habituation trial, and for five consecutive days (D1–D5), each bird was presented with the closed box loaded with food, for one 15-min trial per day. The first trial with the box closed (D1) was considered to be the innovative problem-solving test per se. We included four additional trials to evaluate the repetition of the innovative behaviour, the possible improvement in the solving performance with experience (i.e., learning), and the possible change in the influence of variables considered as potential mechanisms of innovation. During each of these five trials, we recorded the contact and solving latency, as well as the number of box-lids opened (hereafter named solving level). We measured contact latency as the time elapsed from the box presentation until the first contact of the box by the individual, and solving latency as the time from this first contact to access to the first piece of food. For those individuals that did not contact the box, or they contacted it but did not successfully open and feed from the box within an experimental session, we recorded a latency of 15 min (scored in seconds). We used two measurements of persistence. The first was the rate of attempts in which an “attempt” was defined as the action of contacting one section of the box with the beak, talons or both. When subjects stopped making contact with the box for at least 10 s or switch to another section of the box, the attempt was considered finished. Thus, an attempt could involve a single motor technique (e.g., peck the box once and then stop for more than 10 s, or switch to another box area), the continuous repetition of the same motor technique (e.g., peck an area of the box in a persistent manner), or a sum of two or more techniques (e.g., pecking and grasping or kicking). Then, to calculate the rate, we divided the number of attempts by the time elapsed between the first box contact and the first lid opening. This measurement was used as an explanatory variable of the first lid solving latency. In addition, we recorded a second persistence measurement, the work time, which was used as an explanatory variable of the solving level reached during the whole trial. This variable was represented by the total time devoted to physically exploring the box, excluding the periods when birds ate the meat after a successful opening, or when they stopped contacting the box and flew up to the perch or walked away from the test apparatus.
Moreover, we calculated the proportion of effective attempts, as the number of attempts made on the lids, divided by the total number of attempts (towards any part of the box) in each trial. This calculation was made for the whole trial (total), and before the first lid opening (initial). We also registered all clearly identified motor techniques used during the opening attempts (Table 6). From these techniques we used the number of different techniques exhibited during each trial as a measurement of motor diversity, and the changes in techniques before individuals opened a lid (or until the session was over) as motor flexibility. As with persistence, motor flexibility was calculated before the first-lid opening, in which case we divided it by the time until first solving (or the individual gives up the task) and, also, for the whole trial (total) in which case we use the total number of technique changes recorded from the beginning of a trial. All time-related variables were scored in seconds, and all values are presented as mean ± SE,
Data analysis
Univariate models for D0 and D1
Since all latency variables were capped at 900 s when an individual did not contact or solve the problem, these values were treated as censored data; that is, data coming from an individual for which a desired event (in our case the contact or solving of the box) was not recorded within the trial period (e.g., Sol et al. 2011). Consequently, we used Cox Proportional Hazards Models to evaluate the effect of habitat and sex, as well as their interaction, on feeding latency during D0 and contact and solving latencies during D1 (coxph function of survival R package, Therneau 2000). We evaluated the validity of these models’ assumptions, using the function cox.zph in the survival package, and ggcoxdiagnostics in survminer package. In the case of solving level, we considered the number of lids opened as an ordinal variable (0, 1, 2, or 3), and to analyse these data from D1 we applied a Cumulative Link Model (CLM) (clm function of ordinal R package, Christensen 2018) with habitat type and sex, as well as its interaction, as explanatory variables. For the rest of the variables (initial attempts rate, motor flexibility and diversity, proportion of effective attempts and working time) we used the Shapiro-test to evaluate normality of the data, and Levene's test of equality of variances. Then, to analyse the effect of habitat type and sex on these parameters, we performed General or Generalized Linear Models (LM or GLM), according to its normality or lack of it, respectively.
Univariate models for variation across trials
To evaluate the effect of test repetition on the behavioural variables studied here, we run mixed models with trial as numeric variable, individual identity as random factor and habitat type nested with trial to obtain a separate analysis for rural and urban chimangos. For contact and solving latency, we used Cox proportional hazards mixed models (coxme function of coxme R package, Therneau et al. 2003); in the case of solving level, we used a Cumulative Mixed Link Model (CLMM) (clmm function of ordinal R package, Christensen 2018); and Generalized Mixed Linear Models (GLMM), for the rest of the variables.
Multivariate models
To infer causal relationships between neophobia, exploration, persistence, motor flexibility and effective solving attempts with the innovative problem-solving performance variables during D1, we fitted a structural equation model for both the solving latency and solving level as the response variables using the R package “piecewiseSEM” (Lefcheck 2016). With this procedure, SEMs are built using a list of structured equations (or causal paths), which can be specified using most common linear modelling functions in R, and thus can accommodate non-normal distributions, hierarchical structures and different estimation procedures (Lefcheck 2016). We tested the overall fit of the piecewise SEMs using the Shipley test of d separation (Shipley 2009), which validates important missing paths that are not included. The combined probabilities of all the paths are compared with a Chi-square distribution with 2 k degrees of freedom (i.e., number of independent claims). If the resulting Fisher’s C value is unlikely to have occurred by chance (P < 0.05), then the SEM model is rejected, because it does not have a good fit to the data (Shipley 2009; Lefcheck 2016). In each SEM, we included neophobia and exploration time as exogenous variables, motor flexibility, persistence, and proportion of effective attempts as intermediate variables, and the solving latency or solving level as each end point (endogenous variable) (Fig. 4). We did not include motor diversity due to its significantly high correlation with motor flexibility in both rural (r = 0.98, p < 0.001) and urban chimangos (r = 0.91, p < 0.001). As the piecewiseSEM package does not support CLM or Survival models, in the case of the path including solving latency, we transformed this variable with base 10 logarithm and applied a LM with Gaussian distribution. In addition, in the case of the path with solving level as response variable we analysed this path using a GLM with Poisson distribution family. We report the marginal R2 for the fixed effects included in each piecewiseSEM analysis. It was not possible to calculate standardized path coefficients for generalized linear models with Poisson distributed response variables (causal path including solving level as response). For the rest we showed both the unstandardized and standardized estimates of each individual path. In addition, we tested whether habitat type had contrasting effects by comparing a multigroup SEM with an overall model (Shipley 2002). With this approach, we could test for the habitat effects both on the entire model and on each causal path independently (Shipley 2002).
Results
Habituation trial (D0)
During D0, all but three rural individuals (2 females, 1 male) obtained at least one piece of meat from the plexiglass box during the 15-min trial. Individuals that did not feed from the box did not even approach and make contact with the test apparatus during the habituation trial. On average, urban birds showed a lower feeding latency (153.5 ± 33.0 s) than rural chimangos (557.5 ± 91.7 s) (Table 1). Neither sex nor its interaction with habitat type resulted statistically significant (Table 1). All urban subjects obtained the four pieces of meat from the box. Rural birds, however, obtained an average of 2.1 ± 0.5 pieces during D0. The time devoted to exploring the box was higher in urban (258.6 ± 19.5 s) compared to rural birds (107.1 ± 28.9 s) (Table1). We didn’t find an effect of sex or its interaction with habitat on this variable during D0 (Table 1).
Innovative problem-solving trial (D1)
During the first day with the lids closed, all but three rural birds contacted the box at least once. The contact latency was higher in rural (316.5 ± 113.7 s) than in urban raptors (88.8 ± 27.4 s) (Table 2). After this first contact, all urban and five rural chimangos opened the test apparatus and obtained at least one piece of food. The solving latency was higher in rural (641.36 ± 99.3 s) than in urban birds (122.1 ± 30.9 s) (Table 2). The rate of attempts was higher in urban (0.16 ± 0.05) compared to rural chimangos (0.04 ± 0.01) (Table 3). The first-lid motor flexibility did not differ between urban (0.05 ± 0.03) and rural birds (0.02 ± 0.01) (Table 3). However, we found a significant difference between habitats regarding the proportion of effective attempts made by the chimangos before the first lid opening. This value was lower in rural birds (0.4 ± 0.1) compared to urban raptors (0.7 ± 0.1) (Table 3).
When analysing variables measured over the full duration of D1, we found a significant effect of habitat type, but not sex or their interaction (Table 3). Urban raptors opened more lids, reaching a higher solving level during D1, than rural birds (3.1 ± 0.3 and 1.1 ± 0.4, respectively). The work time was also higher in urban (553.7 ± 62.5 s) compared to rural birds (165.1 ± 43.9 s). Moreover, rural birds showed a lower value of the proportion of effective attempts (0.3 ± 0.1) compared to urban birds (0.8 ± 0.03). Finally, we observed that total diversity and motor flexibility were higher in urban chimangos (3.5 ± 0.3 and 16 ± 1.8, respectively) compared with rural birds (1.5 ± 0.4 and 5.4 ± 2.1, respectively).
As sex was not an important factor explaining the variation in any of the behavioural variables recorded during D0 or D1, and taking into account the low representation of males in the sample data from the two habitats, we opted not to include this factor in the remaining analysis to avoid an unnecessary decrease in the robustness of our models.
Direct and indirect effects on problem-solving performance
Since neophobia and exploration time during D0 were highly correlated (r = 0.91, p < 0.001), we performed a principal component analysis from which we extracted the first component (which explained 92.7% of the total variance) and used it as a combined variable (henceforth termed the neo-exp variable) in further analyses. Negative (and lower) values of this variable reflect individuals with lower neophobia and higher exploration times than those with more positive values. Global piecewise SEMs performed in relation to the two solving performance variables showed a good fit (psem1 [solving latency]: Fisher's C = 0.233, p = 0.89, AIC: 30.2; psem2 [solving level]: Fisher's C = 0.38, p= 0.827, AIC: 32.3). As the Fisher’s C values cannot be computed in a saturated model, we omitted the path connecting persistence (rate of attempts and working time) with the two solving performance variables. These missing paths were non-significant in both cases (solving latency ~ rate of attempts: p= 0.890; solving level ~ working time: p = 0.699). The multigroup analysis of these two models showed differences in some of the paths between rural and urban chimangos (Table 4). Thus, according to the psem1, in urban birds the solving latency was directly and positively influenced by the neo-exp variable and negatively influenced by the first-lid proportion of effective attempts (Table 4a, Fig. 2a). The first-lid proportion of effective attempts was, in turn, related negatively to the neo-exp variable (Table 4a, Fig. 2a.). Both first-lid rate of attempts and motor flexibility had not effect (direct or indirect) on solving latency during D1 in chimangos from the city (Fig. 2a, Table 4a). Rate of attempts was negatively influenced by neo-exp and showed a positive effect on motor flexibility. In rural birds, motor flexibility and the neo-exp variable had a direct effect on solving latency (Table 4a, Fig. 2a.), with the rest of the paths in the model being similar to those of urban chimangos.

Diagram of the Piecewise structural equation model showing direct and indirect of behavioural traits on a) solving latency and b) solving level, observed in chimangos during D1. Arrows represent significant (p < 0.05) interactions (paths) between variables; black arrows show significant paths in both rural and urban chimangos, whereas white and grey arrows represent significant interactions only for rural or urban birds, respectively. Next to the arrows, we include the significant path standardized coefficients for the two habitats (urban: BU and rural: BR), resulted from applying the multigroup function of the “piecewiseSEM” R package on the global model (Fig. 4). We also include R2 values for component models in the boxes of their response variables
According to the psem2, solving level was directly and positively affected by the total proportion of effective attempts in both urban and rural individuals (Table 4b, Fig. 2b). This proportion of effective attempts had a positive relationship with total motor flexibility and a negative one with the neo-exp variable. In turn, motor flexibility was affected positively by work and work time was negatively related to neo-exp. In rural birds, the only difference with urban chimangos was found in the relationship between the total proportion of effective attempts and working time, which was positive and significant for this group, whereas in urban birds this relation was clearly non-significant. Otherwise, the significant paths in the model remained the same for urban and rural chimangos (Table 4b, Fig. 2b).
Problem-solving repetition phase
All individuals decreased their latency to contact the box and open the first lid across trials (Table 5, Fig. 3). Moreover, the rate of attempts recorded before the first lid opening increased in both urban and rural raptors, though it only showed significant variation in urban chimangos (Table 5). The initial proportion of effective attempts increased across days in raptors from the two habitats. In addition, the initial motor flexibility increased with time in rural individuals, though for urban chimangos this variable decreased across successive presentations of the box (Table 5, Fig. 3).

Temporal variation of solving performance variables (solving latency and level) and the explicative behavioural traits analysed for urban (full lines) and rural chimangos (dotted lines). Markers represent the mean values and vertical bars denote ± SE
Solving level significantly increased across trials in rural raptors (Table 5, Fig. 3). In chimangos from the city there was only a slight increase in the quantity of lids opened with time (Fig. 3). Regarding working time, there was significant variation in this value across days for urban and rural birds, but they differed in the direction of this trend. That is, while rural birds showed an increase in work time across trials, the opposite occurred in urban chimangos (Fig. 3). Similarly, the total motor flexibility and diversity increased across session days in rural birds but decreased in urban raptors (Fig. 3). Finally, the total proportion of effective attempts increased with time in rural birds, whereas for urban chimangos this value was always high and it did not vary significantly (Table 5).
Discussion
The first phases of the innovation process correspond to the encountering by an individual of a novel opportunity, and the behavioural interaction with it (Tebbich et al. 2016). In the innovative problem-solving paradigm these phases can be represented by the first interaction of the individuals with the Plexiglass box filled with food; that is, D0 with the open box, and the first day with the closed box, or D1 (the innovative problem-solving test per se). The main reason for the inclusion of a D0 in our experimental protocol was measuring the initial reaction of chimangos to the box itself. In this sense, novelty responses (i.e., neophobia–neophilia, exploration) are among the most well-known mechanisms underlying the occurrence of an innovation (Greenberg 2003), and are considered to have an important role in the adjustment of animals to urban life (Griffin et al. 2017; Sol et al. 2020). Indeed, in this study we found that urban birds reached the food from the open box faster and spent more time exploring it, than rural birds. This result is in line with recent work showing a lower level of object neophobia and higher boldness and object explorative behaviour in chimangos coming from the city compared to rural ones (Biondi et al 2020). We also found that the birds’ time exploring the open box during D0 was negatively related to their neophobia level, a result that was already shown by Biondi and collaborators (2020) with object neophobia and exploration in chimangos from these two habitats. This gives additional support to the idea that neophobia can influence the level of the exploratory behaviour exhibited (i.e., O’Hara et al. 2017).
As we predicted, chimangos from the city outperformed rural individuals in both solving speed and solving level during the first confrontation with the closed box (D1). To reach a deeper understanding of why this difference may occur, we went beyond the habitat comparison and investigated several of the behavioural traits considered key mechanisms of the innovation process (Tebbich et al. 2016). In addition to neophobia and exploration during D0, we analysed the role of persistence, diversity and flexibility in the use of behavioural techniques, as well as the proportion of effective attempts. First, we found a clear difference in these variables between habitats, with urban birds showing higher values of persistence, motor diversity and flexibility, as well as proportion of effective attempts compared to rural chimangos. Second, we found that all these behavioural variables contributed directly and indirectly to the variation in innovative problem-solving performance, though the importance of the causal paths connecting these variables differed between rural and urban chimangos.
There are only a few studies comparing persistence, in a problem-solving context, between individuals from habitats with different urbanization levels (Sol et al. 2011; Papp et al. 2015; Prasher et al. 2019). For example, Sol and collaborators (2011), working with common mynas, found that individuals from more urbanized areas had a higher contact frequency with a test apparatus and a higher propensity to innovate compared with birds from less urbanized habitats. In line with our initial expectations, and like this previous study, our results showed that the two measurements of persistence—rate of attempts made by the individuals before the first-lid solving, and the work time registered during the whole trial—were significantly higher in urban than rural raptors during D1. Moreover, similar to Sol and collaborators (2012), both the rate of attempts and work time were related to the novelty response to the box—individuals with lower neophobia and higher exploratory behaviour during D0 were those who showed higher values of persistence. This result can be expected considering that solving persistence and box initial explorative behaviour probably represent two aspects of the same explorative personality (i.e., Massen et al. 2013).
Persistence did not directly explain the variability in problem-solving performance, but did so indirectly especially through its positive relationship with motor flexibility. Motor flexibility and motor diversity have been found to lead to faster solutions or a higher probability of innovative problem-solving in numerous species (i.e., Griffin and Diquelou 2015; Diquelou et al. 2016, Daniel et al. 2019). However, we need to point out that our definition motor of flexibility (“number of switches between behavioural techniques”) differs slightly from how motor flexibility has been defined in other studies (“a combination between total number of techniques used and their relative distribution”, in Griffin and Diquelou 2015). Flexibility of foraging motor actions represents a characteristic of dietary generalists, such as chimangos. These generalist raptors have a notable dexterity for handling all kind of prey items, including artificial objects, such as plastic containers or other types of man-made food packaging (Biondi et al. 2013, 2015). This last fact is particularly true for chimangos living in anthropized areas, and may be one of the possible explanations for the higher motor diversity and flexibility observed in urban raptors. The importance of motor flexibility varied between the two measures of problem-solving performance. For solving latency, an individual's initial motor flexibility had a significant and direct positive effect only in rural raptors. Contrarily, in chimangos from the city we did not find a significant connection between motor flexibility and solving latency, but it was directly explained by the proportion of effective attempts and the initial novelty response toward the box. It seems that, when the effective solving attempts are relatively low, as occurred in rural birds, motor flexibility becomes an important explanatory factor for the variation in first-lid solving speed. In contrast, when effective solving attempt are relatively high, as in urban birds, motor flexibility loses explanatory power. As for solving level, motor flexibility indirectly influenced the number of lids opened during D1, through its positive relationship with the proportion of effective attempts; this pattern was observed in both rural and urban chimangos alike.
From these results it follows that the most important mediating factor between these behavioural factors (initial novelty response, motor flexibility and persistence) and the two problem-solving performance variables was the proportion of effective attempts. Individuals that contacted the box with higher frequency on lid areas were those who solved the first lid quicker, and obtained more pieces of meat from the test apparatus during D1. It has been suggested that contacts aimed at mobile elements of a test apparatus (i.e., lids) may provide an individual with secondary cues leading to the access of food, so increasing persistence on these particular areas would increase problem-solving success and speed (Overington et al. 2011; Guez and Griffin 2016). This idea, might parallel with what we observed in this study with chimangos. In addition, the novelty reaction to the box during D0 was the common factor influencing the two measurements of the proportion of effective attempts (before the first-lid opening and during the whole trial). This means that individuals who showed lower neophobia and more initial explorative behaviour toward the box, were those showing higher proportions of effective attempts. It seems plausible that chimangos, while exploring the box during D0, acquired knowledge about its characteristics (i.e., the presence of mobile lids, the areas, where the food was accessible). This information could have been used by the individuals during D1 to focus their attention on the functional parts of the test apparatus, thus influencing the time needed to access the reward, as well as the number of lids opened. In addition, the total proportion of effective attempts was positively related to motor flexibility and persistence registered during the whole D1 trial. A higher persistence for contacting the box in areas with mobile lids may have motivated the individuals to switch between different contact techniques more frequently, compared to locations without lids, which would explain the observed link connecting persistence and motor flexibility with attempt effectiveness.
Overall, these results highlight the central role of the novelty reaction to the test apparatus in explaining the variability of problem-solving performance in chimangos. In part it influenced the interactions of individuals with the test apparatus, and the amount of information about the system that individuals were able to use at the beginning of the innovative problem-solving test. This novelty response showed a direct and indirect effect on birds’ persistence, motor flexibility and effectiveness of solving attempts, all of which represented key behavioural factors that contribute to the habitat differences in innovative problem-solving found in this study. Moreover, the fact that neophobia and initial exploration of the box differed between rural and urban chimangos not only paved the way to a clear habitat difference in solving performance during the first exposure to the feeding problem, but also set a discrepancy in the way that solving performance and its associated behavioural traits changed across time in these two raptors groups. In this sense, during problem-solving test repetition, we observed a decrease of solving latency through successive trials, which was much more abrupt in rural compared to urban birds. This habitat difference in the solving improvement can be best understood by considering that city chimangos already showed a relatively short first-lid solving latency during D1, so there was less room for a decrease in solving latency, and therefore, a less pronounced improvement compared to rural raptors, which started with high latency values and thus they had much more room to improve with time. This improvement occurred alongside an increase in persistence, in the proportion of effective attempts and, for rural raptors, an increase in motor flexibility and diversity. In chimangos from the city motor diversity and flexibility decreased with time, indicating that urban birds used the additional trials to narrow in on the most efficient techniques to solve the box. This pattern observed primarily in urban chimangos was in accordance with the results found in a previous study on Grey squirrels (Chow et al. 2016), in which an improvement in solving time across trials was associated with an increase in persistence and behavioural selectivity, and a decrease in flexibility. Urban and rural chimangos also differed in how their solving level changed across time. Rural birds started with a low number of lids opened and then rapidly increased this value across days, whereas city birds already showed a solving level close to the maximum during D1, so increased only slightly with additional presentations with the problem. This dissimilar pattern was reflected by a difference in the temporal variation observed in the associated behavioural traits. In particular, the total proportion of effective attempts increased sharply with time in rural birds, whereas this variable did not change significantly in urban raptors. Moreover, persistence and motor flexibility and diversity increased across trials in rural birds, whereas in city chimangos these variables decreased with time. It seems that rural birds increased their engagement with the box over successive trials, behaving in a more diverse and flexible way, to open more lids. Urban raptors, on the other hand, having solved the majority of the box during the beginning of the problem-solving trials, improved their performance by focusing on the effective areas of the box and by reducing the number of times they switched between opening techniques (probably to focus on the most effective actions needed to solve the lids).
This fine-tuning of the opening behaviour in urban chimangos, along with the increase in the attempts focused on the lid areas of the box in rural raptors, can be taken as evidence for the involvement of operant learning in the improvement of solving performance across trials, as was observed in previous studies (e.g., Benson-Amram and Holekamp 2012; Chow et al. 2016). However, from this work, we cannot say with certainty that urban and rural raptors differed in their learning speed, mainly because differences in neophobia and initial exploration influenced the degree to which they interacted with the box and, therefore, led to a difference in the learning opportunities these individuals had from the beginning of the innovative problem-solving test. Only if all individuals experienced the same amount of learning opportunities with the box, could their learning rate be extracted and correctly compared (see Griffin and Guez 2016). Hence, to circumvent these difficulties, future studies in this raptor species should measure the relationship between innovative problem-solving and learning ability evaluating this last attribute through the use of separate associative learning test batteries (e.g., Griffin et al. 2013; Ducatez et al. 2014).
In conclusion, this work provides new evidence for the role of novelty responses in the ability of animals to cope with urbanization. In addition, our results are in line with the increasing number of studies showing that innovativeness can be predicted by an array of non-cognitive behavioural traits, which act in consort to increase the chances of quickly solving novel problems in urban areas.
References
Aprile G, Bertonatti, C (1996) Manual sobre rehabilitación de fauna. Boletín Técnico FVSA (Fundación Vida Silvestre Argentina), Buenos Aires, Argentina.
Audet JN, Ducatez S, Lefebvre L (2016) The town bird and the country bird: problem solving and immunocompetence vary with urbanization. Behav Ecol 27(2):637–644. https://doi.org/10.1093/beheco/arv201
Benson-Amram S, Holekamp KE (2012) Innovative problem solving by wild spotted hyenas. Proc R Soc B 279(1744):4087–4095. https://doi.org/10.1098/rspb.2012.1450
Benson-Amram S, Weldele ML, Holekamp KE (2013) A comparison of innovative problem-solving abilities between wild and captive spotted hyaenas Crocuta Crocuta. Anim Behav 85(2):349–356. https://doi.org/10.1016/j.anbehav.2012.11.003
Biondi LM, Bó MS, Favero M (2005) Dieta del chimango (Milvago chimango) durante el periodo reproductivo en el sudeste de la provincia de Buenos Aires. Argentina Ornitol Neotrop 16:31–42
Biondi LM, Bó MS, Vassallo AI (2008) Experimental assessment of problem solving by Milvago chimango (Aves: Falconiformes). J Ethol 26:113–118. https://doi.org/10.1007/s10164-007-0035-2
Biondi LM, Bó MS, Vassallo AI (2010) Inter-individual and age differences in exploration, neophobia and problem-solving ability in a Neotropical raptor (Milvago chimango). Anim Cogn. https://doi.org/10.1007/s10071-010-0319-8
Biondi LM, Guido J, Madrid E, Bó MS, Vassallo AI (2013) The effect of age and sex on object exploration and manipulative behavior in a neotropical raptor, the chimango caracara, Milvago chimango. Ethology 119:221–232. https://doi.org/10.1111/eth.12056
Biondi LM, Guido J, Bó MS, Muzio RN, Vassallo AI (2015) The role of stimulus complexity, age and experience in the expression of exploratory behaviour in the Chimango Caracara, Milvago chimango. Anim Cogn 18:139–150. https://doi.org/10.1007/s10071-014-0785-5
Biondi LM, Fuentes GM, Córdoba RS, Bó MS, Cavalli M, Paterlini CA, Castano MV, García GO (2020) Variation in boldness and novelty response between rural and urban predatory birds: The Chimango Caracara, Milvago chimango as study case. Behav Process 173:104064. https://doi.org/10.1016/j.beproc.2020.104064
Cabezas VM, Schlatter RP (1987) Hábitos y comportamiento alimentario de Milvago chimango Vieillot, 1816 (Aves, Falconidae). An Mus Hist Nat Valp 18:131–141
Carrete M, Tella JL, Blanco G, Bertellotti M (2009) Effects of habitat degradation on the abundance, richness and diversity of raptors across Neotropical biomes. Biol Conserv 142(10):2002–2011. https://doi.org/10.1016/j.biocon.2009.02.012
Cauchard L, Boogert NJ, Lefebvre L, Dubois F, Doligez B (2013) Problem solving performance is correlated with reproductive success in a wild bird population. Anim Behav 85:19–26. https://doi.org/10.1016/j.anbehav.2012.10.005
Chow PKY, Lea SEG, Leaver LA (2016) How practice makes perfect: the role of learning, flexibility, and persistence in problem solving efficiency. Anim Behav 112:273–283. https://doi.org/10.1016/j.anbehav.2015.11.014
Christensen RHB (2018) “Ordinal—Regression Models for Ordinal Data.” R package version 2018.8–25. http://www.cran.r-project.org/package=ordinal/.
Cole EF, Morand-Ferron J, Hinks AE, Quinn JL (2012) Cognitive ability influences reproductive life history variation in the wild. Curr Biol 22:808–1812. https://doi.org/10.1016/j.cub.2012.07.051
Cole EF, Quinn JL (2011) Personality and problem-solving performance explain competitive ability in the wild. Proc. R. Soc. B.2791168–1175. http://doi.org/https://doi.org/10.1098/rspb.2011.1539
Daniels SE, Fanelli RE, Gilbert A, Benson-Amram S (2019) Behavioral flexibility of a generalist carnivore. Anim Cogn 22:387–396. https://doi.org/10.1007/s10071-019-01252-7
Diquelou MC, Griffin AS, Sol D (2016) The role of motor diversity in foraging innovations: A cross-species comparison in urban birds. Behav Ecol 27(2):584–591. https://doi.org/10.1093/beheco/arv190
Ducatez S, Audet JN, Lefebvre L (2014) Problem-solving and learning in Carib grackles: individuals show a consistent speed-accuracy trade-off. Anim Cogn 18:485–496. https://doi.org/10.1007/s10071-014-0817-1
Ferguson-Lees J, Christie DA (2001) Raptors of the World. Houghton Mifflin, Harcourt, Boston
Ferrer-Sánchez Y, Rodríguez-Estrella R (2015) Man-made environments relationships with island raptors: endemics do not cope with habitat changes, the case of the island of Cuba. J Biodivers Conserv Bioresour Manag 24:407–425. https://doi.org/10.1007/s10531-014-0819-y
Greenberg R (2003) The role of neophobia and neophilia in the development of innovative behaviour of birds. In: Reader SM, Laland KN (eds) Animal Innovation. Oxford University Press, Oxford, pp 175–196
Griffin AS (2016) Innovativeness as an emergent property: a new alignment of comparative and experimental research on animal innovation. Phil Trans R Soc. https://doi.org/10.1098/rstb.2015.0544
Griffin AS, Diquelou MC (2015) Innovative problem solving in birds: a cross-species comparison of two highly successful passerines. Anim Behav 100:84–94. https://doi.org/10.1016/j.anbehav.2014.11.012
Griffin AS, Guez D (2014) Innovation and problem solving: a review of common mechanisms. Behav Process 109(Pt B):121–134. https://doi.org/10.1016/j.beproc.2014.08.027
Griffin AS, Guez D (2016) Bridging the gap between cross-taxon and within species analyses of behavioral innovations in birds: Making sense of discrepant cognition e innovation relationships and the role of motor diversity. Adv Study Behav 48:1–40. https://doi.org/10.1016/bs.asb.2016.02.001
Griffin AS, Guez D, Lermite F, Patience M (2013) Tracking changing environments: innovators are fast, but not flexible learners. PLoS ONE 8:e84907. https://doi.org/10.1371/journal.pone.0084907
Griffin AS, Diquelou MC, Perea M (2014) Innovative problem solving in birds: a key role of motor diversity. Anim Behav 92:221–227. https://doi.org/10.1016/j.anbehav.2014.04.009
Griffin AS, Netto K, Peneaux C (2017) Neophilia, innovation and learning in an urbanized world: A critical evaluation of mixed findings. Curr Opin Behav Sci 16:15–22. https://doi.org/10.1016/j.cobeha.2017.01.004
Guez D, Griffin AS (2016) Unraveling the key to innovative problem solving: A test of learning versus persistence. Behav Ecol 27(5):1449–1460. https://doi.org/10.1093/beheco/arw055
Guido JM, Biondi LM, Vassallo AI, Muzio RN (2017) Neophobia is negatively related to reversal learning ability in females of a generalist bird of prey, the Chimango Caracara, Milvago chimango. Anim Cogn 20:591–602. https://doi.org/10.1007/s10071-017-1083-9
Kozlovsky DY, Weissgerber EA, Pravosudov VV (2017) What makes specialized food-caching mountain chickadees successful city slickers? Proc r Soc b Biol Sci 284:25–27. https://doi.org/10.1098/rspb.2016.2613
Lefcheck JS (2016) piecewiseSEM: Piecewise structural equation modelling in r for ecology, evolution, and systematics. Methods Ecol Evol 7:573–579. https://doi.org/10.1111/2041-210X.12512
Lefebvre L (2011) Taxonomic counts of cognition in the wild. Biol Lett. https://doi.org/10.1098/rsbl.2010.0556
Liker A, Bokony V (2009) Larger groups are more successful in innovative problem-solving in house sparrows. Proc Natl Acad Sci U S A 106(19):7893–7898. https://doi.org/10.1073/pnas.0900042106
Massen JJM, Antonides A, Arnold A-MK, Bionda T, Koski SE (2013) A behavioral view on chimpanzee personality: exploration tendency, persistence, boldness, and tool-orientation measured with group experiments. Am J Primatol 75:947–958. https://doi.org/10.1002/ajp.22159
O’Hara M, Mioduszewska B, von Bayern A, Auesperg A, Bungyar T, Wilkinson A, Huber L, Gajdon GK (2017) The temporal dependence of exploration on neotic style in birds. Sci Rep 7:4742. https://doi.org/10.1038/s41598-017-04751-0
Overington SE, Cauchard L, Cote KA, Lefebvre L (2011) Innovative foraging behaviour in birds: what characterizes an innovator? Behav Process 87(3):274–285. https://doi.org/10.1016/j.beproc.2011.06.002
Papp S, Vincze E, Preiszner B, Liker A, Bókony V (2015) A comparison of problem-solving success between urban and rural house sparrows. Behav Ecol Sociobiol 69:471–480. https://doi.org/10.1007/s00265-014-1859-8
Prasher S, Evans JC, Thompson MJ, Morand-Ferron J (2019) Characterizing innovators: Ecological and individual predictors of problem-solving performance. PLoS ONE 14(6):e0217464. https://doi.org/10.1371/journal.pone.0217464
Ramsey G, Bastian ML, van Schaik C (2007) Animal innovation defined and operationalized. Behav Brain Sci 30:393–407. https://doi.org/10.1017/S0140525X07002373
Reader SM, Laland KN (2003) Animal innovation: an introduction. In: Reader SM, Laland KN (eds) Animal Innovation. Oxford University Press, Oxford, pp 4–35
Reader SM, Morand-Ferron J, Flynn E (2016) Animal and human innovation: novel problems and novel solutions. Phil Trans R Soc. https://doi.org/10.1098/rstb.2015.0182
Shipley B (2002) Cause and correlation in biology: a user’s guide to path analysis, structural equations and causal inference. Cambridge University Press, Cambridge, UK
Shipley B (2009) Confirmatory path analysis in a generalized multilevel context. Ecology 90:363–368. https://doi.org/10.1890/08-1034.1
Sol D, Duncan RP, Blackburn TM, Cassey P, Lefebvre L (2005) Big brains, enhanced cognition, and response of birds to novel environments. Proc Natl Acad Sci U S A 102:5460–5465. https://doi.org/10.1073/pnas.0408145102
Sol D, Griffin AS, Bartomeus I, Boyce H (2011) Exploring or avoiding novel food resources? The novelty conflict in an invasive bird. PLoS ONE 6:e19535. https://doi.org/10.1371/journal.pone.0019535
Sol D, Griffin AS, Bartomeus I (2012) Consumer and motor innovation in the common myna: The role of motivation and emotional responses. Anim Behav 83(1):179–188. https://doi.org/10.1016/j.anbehav.2011.10.024
Sol D, Lapiedra O, González-Lagos C (2013) Behavioural adjustments for a life in the city. Anim Behav 85:1101–1112. https://doi.org/10.1016/j.anbehav.2013.01.023
Sol D, Lapiedra O, Ducatez S (2020) Cognition and Adaptation to Urban Environments. In: Szulkin M, Munshi-South J, Charmantier A (eds) Urban Evolutionary Biology. Oxford University Press, Oxford, pp 253–267
Solaro C, Sarasola JH (2019) Urban living predicts behavioural response in a neotropical raptor. Behav. Process 169:103995. https://doi.org/10.1016/j.beproc.2019.103995
Tebbich S, Griffin AS, Peschl MF, Sterelny K (2016) From mechanisms to function: an integrated framework of animal innovation. Phil Trans R Soc. https://doi.org/10.1098/rstb.2015.0195
Therneau T, Grambsch P, Pankratz VS (2003) Penalized survival models and frailty. J Computational and Graphical Statistics 12:156–175
Therneau T, Grambsch P (2000) Modeling Survival Data: Extending the Cox Model. Springer-Verlag.
Thornton A, Samson J (2012) Innovative problem solving in wild meerkats. Anim Behav 83:1459–1468. https://doi.org/10.1016/j.anbehav.2012.03.018
van Horik JO, Madden JR (2016) A problem with problem solving: Motivational traits, but not cognition, predict success on novel operant foraging tasks. Anim Behav 114:189–198. https://doi.org/10.1016/j.anbehav.2016.02.006
West-Eberhard MJ (2003) Developmental Plasticity and Evolution. Oxford University Press, New York
White CM, Olsen PD, Cliff LF (1994) New world vultures to Guineafowl. In: Del Hoyo J, Sargalat EA (eds) Handbook of the Birds of the World 2. Lynx Editions, Barcelona, pp 216–247
Acknowledgements
We thank Analia Medina and Rodrigo Santiago Córdoba for their assistance during the capturing and managing of raptors. We appreciate the improvements in English usage made by Sanjay Prasher through the Association of Field Ornithologists' program of editorial assistance. This work was conducted with funds provided by the Universidad Nacional de Mar del Plata (EXA 842/17, 15/E795).
Author information
Authors and Affiliations
Contributions
LMB conceived of the project, LMB, GMF and MSB caught birds, LMB and GMF performed the research and the analysis, LMB wrote the first draft of the manuscript and MSB edited it. MSB and LMB provided funding for the research project.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
The capture of raptors adhered to guidelines for the use of animals in research and to the legal requirements of Argentina: Disposition N 45, Exp. N 22500–24126/13, Dirección Contralor y Uso de Recursos Naturales y pesqueros, Ministerio de Asuntos Agrarios de la Provincia de Buenos Aires. The experimental protocol design and the housing and care conditions had the approval of the National University of Mar del Plata Animal Ethics Committee (RD 179/2015), and meets the latest ASAB/ABS ethical guidelines (https://doi.org/10.1016/j.anbehav.2019.11.002). All individuals were released with identifier-coloured bands at their original habitats after complete the experimental tests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Appendix
Appendix

General design of piece-wise structural equation models (psem) proposed to test the direct and indirect effect of several behavioural traits on problem-solving performance. The variable ‘persistence’ was represented by rate of attempts in the model including solving latency as end point, and working time in the case of solving level as model’s endogenous variable
Rights and permissions
About this article

Cite this article
Biondi, L.M., Fuentes, G. & Susana, M. Behavioural factors underlying innovative problem-solving differences in an avian predator from two contrasting habitats. Anim Cogn 25, 529–543 (2022). https://doi.org/10.1007/s10071-021-01569-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-021-01569-2