
Abstract
Self-control is defined as the ability or capacity to obtain an objectively more valuable outcome rather than an objectively less valuable outcome though tolerating a longer delay or a greater effort requirement (or both) in obtaining that more valuable outcome. A number of tests have been devised to assess self-control in non-human animals, including exchange tasks. In this study, three chimpanzees (Pan troglodytes) participated in a delay of gratification task that required food exchange as the behavioral response that reflected self-control. The chimpanzees were offered opportunities to inhibit eating and instead exchange a currently possessed food item for a different (and sometimes better) item, often needing to exchange several food items before obtaining the highest valued reward. We manipulated reward type, reward size, reward visibility, delay to exchange, and location of the highest valued reward in the sequence of exchange events to compare performance within the same individuals. The chimpanzees successfully traded until obtaining the best item in most cases, although there were individual differences among participants in some variations of the test. These results support the idea that self-control is robust in chimpanzees even in contexts in which they perhaps anticipate future rewards and sustain delay of gratification until they can obtain the ultimately most valuable item.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Sometimes, waiting leads to something better than what one has right now. This waiting can take a form in which nothing must be done to generate a better future outcome, as when one allows money to accumulate interest or one allows food to ripen or grow rather than be eaten more immediately. Or, it can involve more direct choices, as when one chooses to give back an immediate and currently possessed reward for something better that will be delivered later, as when one refrains from taking $5 now for $20 next week (an inter-temporal choice situation). All of these situations reflect what is called self-control, a capacity that is linked with clear advantages in a number of circumstances ranging from dietary habits to financial well-being (Baumeister et al. 1994; Logue 1988; Mischel 2014). This capacity can be measured in childhood as a means of anticipating objective outcomes much later in life (e.g., Mischel et al. 1988; Moffitt et al. 2011; Shoda et al. 1990), with greater self-control exhibited at a young age related to better outcomes later in life.
A large amount of research has been conducted both developmentally and comparatively to understand the emergence of this capacity across the life span and across species. Many of these tests make use of inter-temporal choice tests with choices between dichotomous smaller-sooner versus larger-later options (e.g., Ainslie 1974; Berns et al. 2007; Logue 1988; Tobin et al. 1993, 1996; Rachlin and Green 1972; Stevens et al. 2005; Stevens and Mühlhoff 2012). Other studies use delay of gratification tests where subjects must maintain their inhibition of taking and eating more immediate rewards to receive better, later rewards (e.g., Anderson et al. 2010; Beran 2002; Beran and Evans 2006; Beran et al. 1999; Evans and Beran 2007a; Evans et al. 2012; Grosch and Neuringer 1981; Parrish et al. 2014; Stevens et al. 2011). The focus of the present article is on another delay of gratification task that makes use of item exchange as the active behavioral response that reflects self-control, and the focus also is on comparing performance across self-control tasks within the same individuals.
Exchange tasks have become increasingly popular in assessing a variety of cognitive behaviors. Some tasks involve the exchange of tokens with experimenters to obtain foods immediately, and these tasks are used to assess things such as inequity aversion and numerical cognition (e.g., Addessi et al. 2007, 2008; Brosnan and de Waal 2003, 2004). Other tests examine exchange between conspecifics, to assess whether animals can understand others’ needs in terms of token value and whether this might result in cooperative behavior (e.g., Brosnan and Beran 2009; Parrish et al. 2013; Pelé et al. 2010a, b).
The use of exchangeable tokens also allows for investigations of self-control. For example, Hackenberg and Vaidya (2003) trained pigeons (Columba livia) to make inter-temporal choices for token rewards that later could be exchanged for access to food. Choice behavior for the smaller-sooner or larger-later outcomes depended on the duration of the exchange delay and the food access delay between the two choices, and the results showed that choices were governed by reinforcer immediacy when exchange and food delays were unequal and by reinforcer amount when exchange and food delays were equal (see also Jackson and Hackenberg 1996). Judge and Essler (2013) designed a token exchange paradigm to test self-control in capuchin monkeys (Cebus apella). Different types of tokens were exchangeable for different types of food, and when monkeys were given tokens for lower-value food items that they could exchange for either that food type or a token for a higher-value food, some monkeys (two of seven) made the token-for-token exchange. This allowed them to later obtain a better food, showing self-control in not taking the more immediate food. Similarly, Bourjade et al. (2012) tested brown capuchin monkeys and Tonkean macaques (Macaca tonkeana) with a token exchange task that also introduced a temporal component that required subjects to collect tokens at one time point, but trade them for food rewards at a later point. The question was whether these species could anticipate the need to have tokens later, even when collecting them in the present offered no immediate value to the monkeys in terms of food acquisition. This was a difficult test, and most monkeys failed to accumulate any tokens, suggesting that they did not seem able to plan for a future exchange situation.
When exchange is more immediate, some species such as chimpanzees (Pan troglodytes) can learn to select symbolic tokens representing high-value foods that can be exchanged for those foods 2–3 min later over more immediately available but less preferred food items (Beran and Evans 2012). In some cases, those animals will wait to collect many tokens rather than trade the first token or few tokens immediately (e.g., Evans et al. 2012). Chimpanzees also have demonstrated the ability to exchange a token repeatedly with a conspecific to facilitate the accumulation of food rewards in a joint accumulation self-control task (Parrish et al. 2013). This task required the apes to monitor the accumulating rewards, work with a partner to accumulate those rewards, and most importantly, refrain from taking the food items so as to maximize the total reward amount that could be obtained.
The use of tokens in self-control tasks has a number of advantages, including that it allows one to assess more “symbolic” choice behavior once subjects learn the value of tokens that, in and of themselves, have no inherent or prepotent appetitive qualities or other physically appealing properties. This point was perhaps most notably expressed in the reverse-reward paradigm in which chimpanzees were required to indicate the food set that would be given to a partner animal (Boysen and Berntson 1995). In essence, the chimpanzees had to learn to select the smaller of two food sets in order to receive the larger food set for themselves. This task only was successful with the introduction of symbolic icons (much like tokens), further supporting the notion that iconic symbols and tokens do not carry the same appetitive qualities and prepotency that actual food items carry. However, if one wants to look at a more tempting and motivationally challenging self-control test that involves exchange, one can use food items as the stimuli that are given to subjects. In these tests, animals are given a particular type (or size) of a food and then offered the chance to exchange that uneaten food for a different food item immediately or after some delay. To obtain the best item, the animal must inhibit eating the food that is in its possession. Thus, this task explicitly requires delay of gratification as in the famous marshmallow test (e.g., Mischel 2014) in which food items were always present but had to be ignored or avoided by children in order for better (or more) food to be obtained later.
Drapier et al. (2005) taught capuchin monkeys to exchange food items for other food items. Within a trial, the food that could be obtained for an exchange could be either qualitatively or quantitatively better than the one the monkey already possessed, and the monkeys often would exchange items when there was a large difference between the value of what they had and what they could get, and this could include as many as three exchanges in a trial. In general, qualitative increases seemed to produce more exchange behavior, but even quantitative changes often led to exchange, although the monkeys sometimes took small bites from an item before exchanging it. Interestingly, not all capuchin monkeys make such responses, suggesting that demonstrations of self-control may be difficult for that species (e.g., Pelé et al. 2011; Ramseyer et al. 2006), a finding also seen in other delay of gratification tests with capuchin monkeys (e.g., Evans et al. 2012). Chimpanzees (Pan troglodytes) have shown greater proclivity for such food-based self-control tasks. When offered small pieces of cookie that could be exchanged for a number of larger-sized pieces, some chimpanzees waited for as long as 8 min without eating the smaller piece (e.g., Dufour et al. 2007). Other animals such as dogs (Canis familiaris; Leonardi et al. 2012), crows (Corvus corone), and ravens (Corvus corax; Dufour et al. 2012) show degrees of success in such food exchange tasks, as do human children (Steelandt et al. 2012), indicating the strong value of this paradigm for comparative assessments of self-control because of easy adaptation for use with so many species due to its intuitive and motivating nature.
The present series of experiments assessed food exchange behavior in a context in which chimpanzees perhaps would anticipate what future rewards might be available, and how long it would be until those items would become available. On each trial, a number of serially presented food items were shown to the chimpanzees, and they could exchange each item, in turn, for the next item in the series. Our manipulations of the series of food items on each trial through the sequence of experiments allowed us to assess the role of individual item value, how long until the best item was presented, the role of item visibility, and the role of delay until exchange could occur on this form of self-control behavior with anticipation of future reward delivery.
Experiment 1
This experiment assessed whether the chimpanzees would exchange food items they were handed for other food items when they could see the entire sequence of items that were available. Different trial types introduced varying degrees of difficulty from the perspective of whether the exchange of an item led to the presentation of an immediately more preferred or less preferred item than the one just traded and from the perspective of whether the exchange of a valued, but not most valuable item, led to more immediate or later opportunities to obtain the highest value item.
Methods
Participants
Three adult chimpanzees were tested: Lana, female, 44 years old; Sherman, male, 41 years old; and Mercury, male, 28 years old. These chimpanzees have long testing histories, and most relevant to this experiment are their histories in self-control tasks that require delay of gratification (Beran et al. 1999) including with accumulation of food reward (e.g., Beran 2002; Beran and Evans 2006; Evans and Beran 2007a). They also have experience in exchanging arbitrary stimuli with each other and with human experimenters (e.g., Beran et al. 2011; Brosnan and Beran 2009) including in some tests in which those exchanges were part of a self-control test (e.g., Evans et al. 2012; Parrish et al. 2013). However, they had never exchanged food items with an experimenter as part of a progression of introduced food items of varying degrees of preference such as in the procedure used in this study.
The chimpanzees lived together as a social group that had access to multiple indoor and outdoor enclosures that included climbing structures, towers, swings, and other forms of enrichment. The chimpanzees voluntarily separated from each other for these tests, but they always had visual and auditory access to the other chimpanzees. Water was available ad libitum, and the chimpanzees were otherwise maintained on their normal diet of fruits, vegetables, and protein sources independent of their performance on this task.
Apparatus
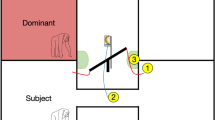
On each trial, the chimpanzees were shown a series of food items placed on a 80-cm-wide, 28-cm-deep tray mounted to a 45-cm-high bench outside of their enclosure. The bench allowed food items to be arranged so that the item that would be delivered first was closest to the subject and the item that would be delivered last was farthest away. A retractable mini blind was mounted to the bench so that, when it was lowered, the two experimenters present during the trials could not see each other although food items could be moved underneath the blind from one experimenter to the other.
Design and procedure
At the beginning of the experiment, we determined the chimpanzees’ food preferences for the following items: banana, orange, apple, sweet potato, carrot, fig bar, and chow biscuit. To do this, pieces of all seven food items were placed on the apparatus in a linear arrangement, equally distant from the chimpanzee, and the chimpanzees selected items, one a time, by reaching and pointing to them (a very common response for these chimpanzees to make in this kind of situation). Items were retrieved and given to the chimpanzee in the order selected, and three or four such trials were given to each chimpanzee. We tallied the ranked choice for each item across those trials and then generated clear preference levels that were the same across all three chimpanzees. Banana and fig bar were the two high-preference food items, so only one of those two items could be presented on a given test trial. Orange and apple were medium-preference items, with orange being more preferred between the two, and sweet potato, carrot, and chow biscuit were the low-preference items, with relative indifference between these items in the preference pretest. Then, the formal test began.
At the start of each test trial, the blind on the apparatus was lowered so that Experimenter 1 and Experimenter 2 were visually separated. Experimenter 1 entered the test area with a bowl of different food items. This experimenter arranged those items, in full view, on the apparatus so that the first presented item was closest to the chimpanzee and then each item that might be exchanged was successively further away, in a linear arrangement on the apparatus. These items were separated by approximately 5–10 cm on the apparatus. With the blind lowered, Experimenter 2, who was seated on the other side, could not see these items and could not see Experimenter 1. Experimenter 2 then raised the blind approximately 5 cm to where food items could be passed under but nothing else could be viewed on the other side. Experimenter 2 announced the start of the trial and began a digital timer. Experimenter 1 then pushed the first food item under the blind to Experimenter 2, who handed it to the chimpanzee. At the same time, this experimenter placed his or her hand under a small port attached to the enclosure that allowed the chimpanzees to drop the item through and back into the experimenter’s hand. If the chimpanzee made this response, Experimenter 2 announced “trade” and Experimenter 1 then pushed the next food item in the sequence under the blind. The trial continued in this manner until the chimpanzee ate a food item, at which point no more food items were exchanged. The total duration of the trial was 90 s, no matter how early or late in the trial the chimpanzee ended the exchanges and ate a food item. After this 90-s period, the next trial immediately began.
This procedure ensured that the experimenter exchanging with the chimpanzee could not know what food item might be presented next and what food items still remained to (possibly) be exchanged, a critical manipulation in preventing any inadvertent cuing of the chimpanzee as to whether or not to exchange. Experimenter 1, who pushed food items to Experimenter 2, did not interact with the chimpanzee at all with regard to exchanging items and thus could not influence performance through any such cues either. The constant trial duration also ensured that the chimpanzees could not speed the presentation of the next trial through any responses of their own.
On each test trial, three, four, five, or six food items were presented. When a high-preference food item was used, only one of the two high-preference food items (banana or fig bar) was presented on that trial. All other items were possible additions to the sequence of exchangeable items on those trials. Not all trials included banana or fig bar, although orange or apple or both were presented on every trial. Thus, orange sometimes was the best item available on a trial, even though in the full food set it was a mid-preference item in comparison with banana and fig bar.
Each test trial also presented the best food item (in terms of preference) in any of the serial positions from the first to the last in however long the sequence was on that trial. There were three test conditions. In the increasing value condition, each exchange led to a more preferred (or equally preferred) food item (e.g., chow biscuit, sweet potato, apple, orange, banana). In the decreasing value condition, each exchange led to lower (or equal)-preference item until the final (fifth) exchange, which produced the more preferred item in the entire set (e.g., orange, apple, sweet potato, chow biscuit, carrot, banana). The random condition presented the food items in a random order, with some exchanges leading to a more preferred next food item (e.g., chow biscuit exchanged for orange) and some to a less preferred next food item (e.g., orange exchanged for chow biscuit). In this way, the chimpanzees could see what would come next and what would come later in the trial if exchanges continued, and then they could determine when a food item they possessed was worth eating versus exchanging, provided they could show the self-control and anticipation necessary to make profitable exchanges. Each chimpanzee completed 76 total trials: 24 trials in the increasing value condition, 24 trials in the decreasing value condition, and 28 trials in the random condition. Sherman completed these trials in seven sessions, and Lana and Mercury did so in eight sessions.
On each trial, one of three outcomes could occur. If a chimpanzee obtained the best item, this was scored as a success. Failures, however, could be of two types and were both defined as trials in which the highest preference food item was not obtained. The first type involved eating a less preferred food item and presumably reflected a failure of self-control, given that all items could be seen. The second type involved trading all items and eating nothing. This would suggest a failure of stimulus control in the sense of understanding what item, of those available, would be the best possible outcome.
Results
The performance of each chimpanzee in obtaining the highest preference item in trials in each condition is shown in Fig. 1. Sherman performed at a very high level in the experiment in this regard, and he was equally proficient in obtaining the highest preference item in all three conditions, χ 2 (2, N = 76) = 0.02, P = .98. Although Mercury did not obtain the most preferred item as often as did Sherman, he still obtained that item on over 67 % of the trials and he too was equally proficient in obtaining the highest preference item in all three conditions, χ 2 (2, N = 76) = 2.32, P = .31. Lana performed much poorer in terms of obtaining the highest preference item, exchanging for that item on only 39 % of the trials. Her performance also was equivalent across the three conditions, χ 2 (2, N = 76) = 2.40, P = .30.

Performance of each chimpanzee in Experiment 1 in obtaining the most preferred (best) item as a function of trial type
An examination of the “errors” in which an item other than the most preferred item was eaten revealed differences among the chimpanzees as well. When Sherman ate an item other than the highest preference item for a trial, it was nearly always the orange slice (10 of 12 error trials), and on the final two error trials the best item was the orange slice, and Sherman exchanged all food items and ended with nothing to eat. Mercury had 25 trials where he did not obtain the highest preference items. On 10 of those trials, he ate the orange slice. On the other 15 trials, neither of the highest preference items was presented, and Mercury traded out all items and ate nothing. Lana had 46 trials where she did not obtain one of the two highest preference items. These were distributed consistently throughout the entire experiment and did not occur more during the beginning or end of the experiment. She ate the orange slice on 19 of those trials, but she also ate low-preference items often (carrot on 16 trials, chow biscuit on 4 trials), and she sometimes traded out all food items when neither of the highest preference items was presented (7 trials).
At the conclusion of the experiment, we re-presented the food preference test to check whether food preferences had changed, especially given Lana’s large proportion of trials choosing the carrot. Each chimpanzee again exhausted all 7 food items presented at once, with a one-by-one selection from among those items. The food preferences shown before the experiment were confirmed after the experiment as remaining the same.
Discussion
Sherman performed at a high level in all conditions and, in essence, showed the self-control necessary to obtain the best item in nearly all trials. Mercury was not as successful as Sherman, but still showed the self-control necessary to exchange items on many trials and obtain the best possible item. His errors, however, suggested that perhaps he did not fully comprehend the nature of the task, especially for trials where high-preference items were not going to be available, given that he traded out all items on some of his error trials. Additional trials may have enhanced his performance (as will be seen by his subsequent performance in the following experiments). That said, there was not a systematic improvement across the trials in Experiment 1 for Mercury, with these types of errors distributed throughout the period of data collection (i.e., he did not make all of these errors early and then become more proficient at the end of the experiment).
Lana struggled with the task, failing to obtain the best item on even half of the trials. This was surprising, given that she has proved to be equally proficient as other chimpanzees in other self-control tasks (e.g., Beran and Evans 2006; Evans and Beran 2007a). The one issue for Lana was her repeated acceptance and consumption of carrot slices, despite the fact that the pretest and posttest food preference assessment showed carrot to be a low-value item when directly compared to the other items that were presented all at the same time. This suggested that her failures to obtain the better food items really were self-control failures, not her choice of carrot as a high-preference item. However, she also had a number of trials in which she received nothing, and as with Mercury, this could reflect an incomplete understanding on her part of the task, especially when high-preference food items were not available. On those trials, she should have accepted and eaten a lower-preference item. It also could be the case that Lana and Mercury simply did not want to consume those items, in the context of this task, even though they regularly eat those items in other contexts. Given that they were receiving high-preference items on many trials, and that they had to either trade all items out or eat something from an array on a given trial, these errors may instead reflect instances in which the chimpanzees did not want anything in the array and traded all items out so as to move to the next trial where a high-preference item might again be presented.
None of the chimpanzees showed an effect of condition. Compared to random trials and increasing value trials, the chimpanzees were equally proficient with decreasing value trials, where they had to exchange the item they possessed for one that was of lower value for all exchanges until the last one, at which point they received the best item possible for that trial. This suggested that the chimpanzees were viewing these trials globally and noting that a better food item eventually would be forthcoming on those trials if exchange behavior was sustained rather than looking to the next item only. Experiment 2 further assessed this anticipation by the chimpanzees by manipulating other factors in how the food items were presented in sequences.
Experiment 2
Methods
Participants and apparatus
These were the same as in Experiment 1.
Design and procedure
The procedure for arranging food items on the apparatus and exchanging with the chimpanzees was the same as in Experiment 1 as were the controls and trial durations used in Experiment 1. In this experiment, all trials involved the presentation of six food items at the start of the trial. A high-preference food item was always included (banana or fig bar) along with orange as the most preferred mid-preference item and apple, chow biscuit, sweet potato, and cucumber (which was substituted for carrot from Experiment 1). The variables that were manipulated included the position of the best food item in the sequence (banana or fig bar; ranging from position 2 to position 6), the point in the trial when the second-best item (orange) was presented (position 1, position 3, or position 5), and the distance between the second-best item and the best item (which could range from 1 to 5 items). These variables created a number of different trial types in which the relative position of the best item, second-best item, and absolute distance between those items varied, allowing us to examine the relative importance of each of these factors on the chimpanzees’ self-control and ability to anticipate a better future reward while being faced with a present reward that they would eat or exchange. Each chimpanzee completed 64 total trials across 6 test sessions in this experiment.
Results
It is difficult in this procedure to define what constitutes statistically significant performance in terms of obtaining the best item that one could obtain. One possibility is to assume that the food that is eaten is taken at chance from the set of six items. With that criterion for establishing a chance level, the results showed that all chimpanzees were significantly above chance levels in their consumption of the best food item available on the trial. Lana consumed the best item on 53 of 64 trials, Mercury consumed the best item on 59 of 64 trials, and Sherman consumed the best item on 51 of 64 trials, all P < .001 as assessed with a binomial test.
Next, we examined whether the chimpanzees obtained the most preferred item on trials as a function of where in the sequence that item appeared. Because of the small number of trials with the most preferred food item in position 2 (N = 4) and position 3 (N = 4), these two trial types were combined to be compared to position 4 (N = 12), position 5 (N = 12), and position 6 (N = 32). None of the chimpanzees showed a difference in performance as a function of where the most preferred item was located in the sequence: Sherman χ 2 (3, N = 64) = 1.71, P = .63; Mercury χ 2 (3, N = 64) = 5.42, P = .14; Lana χ 2 (3, N = 64) = 0.40, P = .94.
Next, we examined performance depending on where in the sequence the second most preferred item was presented. Although Sherman obtained more of the high-preference items in trials in which the second-best item was presented in position 3 or position 5 relative to position 1, this was not a statistically significant difference, χ 2 (2, N = 64) = 4.18, P = .12. Mercury showed little or no difference as a function of where the second-best item appeared, χ 2 (2, N = 64) = 2.10, P = .35. This was also true for Lana, χ 2 (2, N = 64) = 1.30, P = .52.
Finally, we examined performance depending on the distance between the second-best item and the subsequent delivery of the most preferred item if exchanges continued (Fig. 2). Again, we combined the trials with distances of four or five items because these distances alone each only occurred in 4 trials. Sherman showed a significant difference, χ 2 (3, N = 64) = 11.17, P = .011. Mercury, however, did not, χ 2 (3, N = 64) = 0.36, P = .95. Lana also did not, χ 2 (3, N = 64) = 1.57, P = .66.

Performance of each chimpanzee in Experiment 2 as a function of the number of positions between the second-best item and the best item that could be obtained on trials through sustained exchanges
Discussion
Experiment 2 showed that these chimpanzees were equally proficient in obtaining the best item across a variety of manipulations of the presentation sequence: where the best item was located in the sequence, where the most tempting other item was in the sequence, and how large a distance there was between those two items. Only Sherman showed an effect of that distance, with it being more difficult for him to give up the orange slice to get the best item when the best item was a larger number of exchanges away from being presented. This might indicate that Sherman struggled more than Lana and Mercury to see that the best item eventually was forthcoming, or perhaps that temporal delay was clear to Sherman but was too aversive for him to maintain his exchange behavior to the same degree that he did when the best item was more immediately forthcoming. Lana and Mercury showed substantial improvement in this experiment compared to Experiment 1. This could reflect greater experience in the task and specifically more opportunity to learn the rule that they should trade out low-preference items until they obtained a high-preference item.
To this point, all exchanges led to qualitative differences in the food items that were obtained for an exchange. There is some evidence that qualitative increases in reward value generate better-sustained delay of gratification than quantitative increases (e.g., Evans and Beran 2007b; Hillemann et al. 2014). To assess whether this was true in the present exchange paradigm, we introduced two of the highest preference items on each trial but varied their size to see if the chimpanzees still would exchange until receiving the larger piece. In addition, Experiment 1 and Experiment 2 allowed the chimpanzees to use a rule such as “trade items until a high-value item is given.” Experiment 3 and Experiment 4 eliminated this strategy by presenting multiple high-value items of the same type on each trial, but with differences in their size. Now, the chimpanzees had to take into account the size of high-preference items as part of the process of choosing what item they most wanted to eat and, if they could, waiting to obtain that item.
Experiment 3
Methods
Participants and apparatus
These were the same as in Experiment 1 and Experiment 2.
Design and procedure
The procedure for arranging food items on the apparatus and exchanging with the chimpanzees was the same as in the first two experiments as were the controls and trial durations used in those experiments. In this experiment, all trials involved the presentation of five food items at the start of the trial, and in this experiment, two of those items were always a small banana slice (5 ± 1 g) and a large banana slice (20 ± 1 g). The remaining food items could include apple, chow biscuit, cucumber, or sweet potato. As in Experiment 2, the variables of interest were the relative positions of the small and large banana slice. The small slice could appear early (position 1), in the middle (position 3), or late (position 5), and the large banana slice could appear in positions 2 through 5 so that distances between the small and large banana slice could range from 1 to 4 positions. Each chimpanzee completed 2 sessions of 12 trials each.
Results and discussion
The results of this experiment were identical for all chimpanzees. On all 24 trials, each chimpanzee always exchanged until the large banana slice was presented and then consumed that item. This indicated the capacity for tolerating exchanges of the highest value food type when it was not the largest food item of that type, and showed that the chimpanzees were exchanging low-preference items on the basis of a rule that went beyond just “trade until a high-value food type is given.” Thus, we next presented trials in which all items were banana slices, and each was of a unique size within the trial.
Experiment 4
Methods
Participants and apparatus
These were the same as in the previous experiments.
Design and procedure
The procedure for arranging food items on the apparatus and exchanging with the chimpanzees was the same as in the first three experiments as were the controls and trial durations used in those experiments. In this experiment, all trials involved the presentation of only banana slices in one of four sizes (5 ± 1 g; 10 ± 1 g; 20 ± 1 g; 40 ± 1 g). As in Experiment 1, we presented increasing value trials, decreasing value trials, and random trials with regard to banana slice size as the factor related to preference and value. Each chimpanzee completed 2 sessions of 8 trials each.
Results and discussion
Sherman exchanged until he obtained the largest banana slice on all 16 trials. Lana exchanged until she received the largest slice on 12 of 16 trials. If one assumes that eating a banana slice from the array occurred by chance (25 %), both of these performances are significantly better than chance, P < .01, binomial test. For trials in which Lana failed to consume the largest slice, she selected the next largest slice on the other four trials, once in an increasing value trial, and three times in random trials. Three of her four choices of the second-largest item occurred when it was the first item presented. Mercury performed far worse, eating the first banana slice he received on 15 of 16 trials. We do know why this manipulation was so detrimental to his performance, especially given that in Experiment 3 he showed that he could exchange banana slices when such exchanges led to even better rewards. Experiment 4 was conducted across more than 1 day, so it was not simply a “bad session” but appeared to be a real failure. Perhaps that many banana options was simply too tempting, or too difficult for him to discriminate in this particular context, and so he ate the first item he received. His failures highlighted the large differences among the chimpanzees for this manipulation.
Our next experiment in the study moved in a new direction. Now, we required the chimpanzees to make use of memory of the possible food items that could be obtained during a trial. Here, the question was whether the chimpanzees could remember what food item was the best possible reward for a trial, and then exchange other items until that item was received.
Experiment 5
Methods
Participants and apparatus
These were the same as in previous experiments.
Design and procedure
The procedure for arranging food items on the apparatus and exchanging with the chimpanzees as well as the controls for cuing were the same as in previous experiments. Now, however, the trial duration was set to 60 s. We also substituted graham cracker as an additional food item in this experiment given the large number of recently completed trials with banana slices. Graham crackers are very high-preference food items like the fig bars that also were used in this experiment.
All trials involved the initial presentation of five food items at the start of the trial, but then those items all were placed into an opaque box on the apparatus so that the chimpanzees could not see what was going to be exchanged next, and they could not see what remained to be gained potentially through exchange. Instead, they had to attend to the items at the start of the trial and then remember what food item was most preferred and whether it had yet to be presented in an exchange. The variables of interest were the position of the best food item in the sequence (graham cracker, fig bar, banana, or orange; ranging from position 3 to position 5) and the distance between the second-best item and the best item (which could range from 1 to 4 items, although on trials where orange was the best item there was no such distance). Each chimpanzee completed 35 total trials across three test sessions in this experiment. In eight of those trials, orange was the best food item. For the remaining trials, a higher-preference banana, fig bar, or graham cracker also was present.
Results
On four trials, Lana exchanged all items she received and thus did not consume anything. Mercury also did this on three trials. In all of those cases, the highest preference item was not shown as part of the set of items, and thus, orange was the best item that could have been obtained. These trials were removed from all subsequent analyses.
First, we examined whether the chimpanzees obtained the most preferred item on trials as a function of where in the sequence that item appeared (Fig. 3a). None of the chimpanzees showed a difference in performance as a function of where the most preferred item was located in the sequence: Fisher’s exact tests—Sherman P = .83; Mercury P = 1.0; Lana P = .68.

a Performance of each chimpanzee in Experiment 5 as a function of where in the sequence the best item was presented. b Performance of each chimpanzee as a function of the number of positions between the second-best item and the best item that could be obtained on trials through sustained exchanges
Next, we examined performance depending on the distance between the second-best item and the subsequent delivery of the most preferred item if exchanges continued (Fig. 3b). Here, we excluded trials that did not contain one of the highest preference items. We combined the trials with distances of three or four items because these distances alone each only occurred in 6 and 3 trials, respectively. None of the chimpanzees showed a significant difference as a function of distance: Fisher’s exact test—Sherman P = .56; Mercury P = .99; Lana P = .99.
Discussion
As with previous experiments, there was variability among the chimpanzees. In this experiment, Lana and Mercury excelled, obtaining the best item on nearly all trials, even though they only saw that item (and the other items) at the outset of the trial before it went into the box. Although this interval between seeing the item and exchanging until they could receive it was fairly short (less than a minute), it does show that these two chimpanzees can make exchanges for a specifically remembered food item. Sherman, however, performed very poorly in this experiment. He showed a strong preference to eat the orange slice once he received it (29 of 35 trials), suggesting that he did not remember or chose not to wait to receive the better item that he saw at trial outset. An informal test at the end of the experiment confirmed that fig bar and graham cracker both were repeatedly selected over orange slices when both were presented to Sherman simultaneously. Thus, his preferences when given choices of items to immediately consume did not appear to have changed at this point in the study.
Experiment 6
The purpose of Experiment 6 was to introduce a delay between the time at which the chimpanzees saw the potential food items that could be obtained through sustained exchange and when that exchange opportunity was presented. The trials were similar to Experiment 5 except that the Delay condition introduced a 5-min interval between when the chimpanzees saw the foods items (which subsequently were placed in the opaque container) and when exchange began. We assessed whether performance suffered with this memory load for the potential items that could be obtained through exchange.
Methods
Participants and apparatus
These were the same as in previous experiments.
Design and procedure
The procedure for presenting the food items and then placing them in the opaque container and the controls for cuing were the same as in Experiment 5 except that now, on some trials, that container remained in the chimpanzee’s area for 5 min while the experimenters moved away from the chimpanzee. The best item again could appear in the third, fourth, or fifth position in the sequence and could be a cracker, a fig bar, a banana slice, or an orange slice. Critically, the orange slice trials allowed us to determine whether the chimpanzees were using a strategy of trading out all items until they received one of the highest preference rewards, which would not require memory of the specific foods, or exchanged for the best possible reward on that trial, a response that would indicate a larger reliance on their memory.
In the no delay condition, Experimenter 1 and Experimenter 2 entered the test area at the same time. Experimenter 2 moved to the exchange location and prepared to exchange items with the chimpanzee. Experimenter 1 presented a bowl with five food items clearly visible to the chimpanzee for 5 s. Those items then were placed into the opaque container, and the first item was removed and given to Experimenter 2. He gave that item to the chimpanzee and then allowed an exchange if the chimpanzee chose to do that. If an item was exchanged, he announced the exchange, and Experimenter 1 handed him the next scheduled item in the food sequence. This continued until the chimpanzee ate an item or until there were no more items to present. Experimenter 1 then showed the remaining contents of the container. These trials typically required about 30–45 s to complete. At the end of 60 s, the two experimenters then moved to a second chimpanzee, repeated that process for the next 60 s, and then moved to the third chimpanzee for the same kind of trial. Critically, the food items and their order of presentation differed across chimpanzees within these blocks of trials, so that Experimenter 2 (and the other chimpanzees that were watching) could not know exactly what food items remained during a given trial. At the end of the three trials, one with each chimpanzee, a 60-s inter-trial interval occurred before presentation of the next block of trials.
In the delay condition, Experimenter 2 remained outside of the test area, and Experimenter 1 entered the area where the first chimpanzee was located to present the food items and then put them in the opaque container with a lid on top to prevent the chimpanzees from looking inside the container during the delay interval. At the end of 60 s, she moved to the second chimpanzee and did the same thing, and at the end of 120 s, she presented foods to the third chimpanzee and then put those foods into that chimpanzee’s opaque container. Then, she left the test area, and three minutes elapsed before she and Experimenter 2 went to the location where the first chimpanzee was located and began the exchange procedure. After 60 s (which was more time than needed for a chimpanzee to exchange all items if it chose to do that), both experimenters moved to the second chimpanzee and offered the exchange opportunity, and after another 60 s, they moved to the third chimpanzee to do the same. Then, the 60-s inter-trial interval began.
Thus, in each block of three trials, each chimpanzee had one exchange opportunity, and all three either could exchange with no delay or with a 5-min delay between when they saw the food items and when they could begin to exchange. So, in those blocks of three trials, the delay was the same for all three chimpanzees, but across blocks of trials the use of a delay or no delay was randomized so that each chimpanzee completed a total of 40 trials with the delay and 40 trials without any delay. Daily sessions consisted of 8 trials per chimpanzee, and 10 sessions were completed by each chimpanzee.
Results
First, we assessed whether there was a difference in performance as a function of delay time (0 delay or 5-min delay) in obtaining the best item when that item was one of the highest preferred rewards (fig bar, banana, or cracker). These results are shown in Fig. 4a. We assessed whether such a difference was evident using a one-tailed Fisher’s exact test, given that our prediction was that longer delays would disrupt performance relative to the 0 delay condition. None of the chimpanzees showed a statistically significant difference—Sherman P = .08; Mercury P = .50; Lana P = .24. It was clear, however, that Sherman performed worse than Lana or Mercury in obtaining the best item on these trials, and in all cases in which he failed, he chose to eat the orange slice that was delivered before the best item.

a Performance of each chimpanzee in Experiment 6 as a function of the delay interval for trials in which the best food item was a high-preference item (fig bar, banana, or cracker). b Performance of each chimpanzee as a function of the delay interval for trials in which orange slice was the best food item that could be obtained
We analyzed Sherman’s data in greater detail. On each trial, our designated “highest preference” item was banana, cracker, or fig bar. When banana was the item included in the test trials, Sherman obtained it in 16 of 20 trials. When cracker was the item included, Sherman obtained it in 7 of 20 trials. When fig bar was the item included, Sherman obtained it in 5 of 20 trials. For all of the trials in which those items were not obtained, Sherman took the orange slice. After the experiment concluded, we re-presented Sherman with the food preference test because of concerns that perhaps his preferences had changed, and the results indicated that they had. We presented nine preference trials like those of Experiment 1 in which apple, sweet potato, cucumber, and orange were presented. In three of each of those trials, banana, cracker, or fig bar were included as the fifth item. Sherman always selected banana first when it was available, but he selected orange over cracker one time, and he selected orange before fig bar on all three trials. Thus, the results of the experiment for Sherman must be qualified by his changed preference for at least one of the three items (fig bar) that we originally considered of highest preference. For the one clear preference that was available in the test trials (banana), he obtained that item on 80 % of the trials, a level more comparable to that of Lana and Mercury for all food items.
Next, we assessed whether there was a difference in performance as a function of delay time (0 delay or 5-min delay) in obtaining the orange slice when it was the best item. These trials were critical because a failure of chimpanzees in this condition would suggest they were not remembering what specific item was the best they could get from the occluded container of items but instead were anticipating only the delivery of the highest preference items, even on these trials when those items were never seen at the trial outset. The results for these trials are shown in Fig. 4b. Sherman almost always ate the orange slice in the 0 delay and 5-min delay conditions, P = .50. Although Lana and Mercury ate the orange slice more often in the 0 delay trials on these critical trials (9/10 trials for Lana and 8/10 trials for Mercury) than the 5-min delay trials (5/10 trials for each chimpanzee), these differences did not exceed statistically significant levels—Mercury P = .17; Lana P = .08. On all of these trials at both delay intervals where Lana or Mercury failed to eat the orange slice, they exchanged all items with the experimenter and thus consumed nothing.
Discussion
There were clear individual differences among the chimpanzees. Sherman ate orange slices far more often than did Mercury and Lana. Thus, when those were the best items in the sequence, he performed at ceiling. However, when more preferred items were available, he was less likely to obtain those items than Lana and Mercury. However, there was no apparent effect of delay interval for Sherman in either case. Lana and Mercury were very good at obtaining the highest preference items no matter whether a delay was inserted between the hiding of items and the exchange period. However, when the best item they could get was the orange slice, they more often failed to obtain that item, and this was especially true for the longer delay, even though this effect failed to reach conventional level of statistical significance. Thus, for these two chimpanzees, there seems to be an effect of memory demands for lower-preference items in this self-control test, and likely one that would have reached statistical significance had we conducted a large number trials. At the same time, when high-preference rewards were available during trials, the general outcome of this experiment was that time delays of this duration had little overall effect on the exchange behavior and the self-control of these chimpanzees, and especially for Lana and Mercury who consistently obtained those best items. It is possible that the chimpanzees by this point in the study (or earlier) were using a strategy of giving back low-preference things until some high-preference item was presented without needing to anticipate the exact item that they could obtain on that trial. Future manipulations will be needed to more fully outline the role of memory in exchange-based self-control tasks.
General discussion
In these experiments, chimpanzees “traded up” for a better but later reward by exchanging a food item currently in their possession to receive a higher-valued reward. Often, this higher-value item was not delivered immediately, but only could be obtained from additional exchanges beyond the present one. In some cases, the task required using memory for the rewards that remained available for exchange in the occluded food set. The current paradigm offers several advantages to a comparative assessment of self-control. As demonstrated nicely with several species in previous research (e.g., Drapier et al. 2005; Dufour et al. 2007, 2012; Pelé et al. 2011; Ramseyer et al. 2006), exchanging food items as a means to obtain a better, later reward is an intuitive task in which the animals control the pacing of the exchange and the termination of the trial by simply consuming the desired reward. In our task, the items available for exchange sometimes were displayed in a linear ascending order in relation to the animal so that the closest reward was the next item for exchange. This layout allowed the animals to easily monitor the available rewards in the order of their delivery.
Within the current set of studies, we assessed the impact on performance in obtaining the best item when reward type, reward size, reward visibility, delay to the reward, and the location of the highest valued reward in the exchange set were varied. The chimpanzees performed well across nearly all variations of the task regardless of the position of the best reward, exchanging less preferred rewards even if the high-valued reward was late in the exchange set. Further, the chimpanzees performed well when the rewards that still could be obtained were non-visible during exchanges. High performance in this exchange task required sustained self-control in which the chimpanzees actively refrained from eating a number of items until they received a preferred food reward. This point also highlights a key feature and valuable aspect of exchange tasks—they require an increase in activity (performing the exchange) to obtain a better but later reward in addition to behavioral inhibition from consuming the reward in their possession.
Despite the overall high performance across experiments, there were a few notable task variations that proved to be difficult for some of the chimpanzees. These individual differences, even in a small sample of highly task-sophisticated chimpanzees, highlight the variability across individuals that can result from a variety of contextual dimensions. For example, when the exchange set contained different sizes of the same high-valued reward (Experiment 4), Lana and Sherman always maximized, exchanging until they obtained the largest piece of banana on every trial. Despite Mercury’s general high performance across other experiments, he completely failed to maximize in this case, consuming the first piece of banana regardless of its size on every trial. The sole presence of the high-valued reward in the exchange set may have increased the banana’s salience, reducing his behavioral inhibition in this trial type. Note that this outcome was likely not the result of Mercury failing to tell apart the difference in those items’ size, as he is proficient in making quantity judgments across a variety of presentation formats (e.g., Beran 2010, 2012; Beran and Beran 2004; Parrish and Beran 2014a, b). In addition, Sherman failed to maximize in the delay experiment (Experiment 6) in which we instantiated a 5-min delay between presentation of the rewards and the exchange. Part of this failure likely was a change in Sherman’s preference for fig bars versus orange slices, which highlights the need to continually assess the relative values of food items when repeated testing with the same animals occurs. But, even for an item that remained high value (cracker), Sherman struggled to obtain that item especially with the 5-min delay. Thus, memory demands appeared to differentially affect the chimpanzees. But, even memory demands may be highly task specific in their effects, as Sherman has proven to be proficient in other studies of food memory and retrieval over much longer time delays (e.g., Sayers and Menzel 2012).
Taken together with previous research (e.g., Beran 2002; Beran et al. 1999; Dufour et al. 2007, 2012; Evans and Beran 2007a; Evans et al. 2012; Parrish et al. 2013), we conclude that chimpanzees displayed self-control in the current study, maximizing food intake in terms of quality and quantity through sustained food exchange. But work remains to be done to describe and define the task conditions that best promote or disrupt self-control in this species, as well as to understand whether generalized rules such as “exchange until something good is presented” might be at work versus more trial-specific memories of what will become available later in a trial. Additional comparative assessments, too, will benefit the continued investigation of the emergence of self-control within and beyond primate species. And, future tests that assess the degree of effort that a chimpanzee would sustain through exchange or similar procedures would be of interest to the current literature, and would likely highlight another area in which individual differences emerge. Ultimately, the emerging conception of chimpanzees (and some other animals) as self-controlled individuals, at least in intuitive kinds of tasks such as exchange tasks, highlights a commonality across species that has the potential to illustrate the evolutionary emergence of some of the basic inhibitory processes at work in human behavior and cognition.
References
Addessi E, Crescimbene L, Visalberghi E (2007) Do capuchin monkeys (Cebus apella) use tokens as symbols? Proc R Soc Lond 274:2579–2585
Addessi E, Crescimbene L, Visalberghi E (2008) Food and token quantity discrimination in capuchin monkeys (Cebus apella). Anim Cogn 11:275–282
Ainslie GW (1974) Impulse control in pigeons. J Exp Anal Behav 21:485–489
Anderson JR, Kuroshima H, Fujita K (2010) Delay of gratification in capuchin monkeys (Cebus apella) and squirrel monkeys (Saimiri sciureus). J Comp Psychol 124:205–210
Baumeister RF, Heatherton TF, Tice DM (1994) Losing control: how and why people fail at self-control. Academic Press, San Diego, CA
Beran MJ (2002) Maintenance of self-imposed delay of gratification by four chimpanzees (Pan troglodytes) and an orangutan (Pongo pygmaeus). J Gen Psychol 129:49–66
Beran MJ (2010) Chimpanzees (Pan troglodytes) accurately compare poured liquid quantities. Anim Cogn 13:641–649
Beran MJ (2012) Quantity judgments of auditory and visual stimuli by chimpanzees (Pan troglodytes). J Exp Psychol Anim Behav Proc 38:23–29
Beran MJ, Beran MM (2004) Chimpanzees remember the results of one-by-one addition of food items to sets over extended time periods. Psychol Sci 15:94–99
Beran MJ, Evans TA (2006) Maintenance of delay of gratification by four chimpanzees (Pan troglodytes): the effects of delayed reward visibility, experimenter presence, and extended delay intervals. Behav Proc 73:315–324
Beran MJ, Evans TA (2012) Language-trained chimpanzees (Pan troglodytes) delay gratification by choosing token exchange over immediate reward consumption. Am J Primatol 74:864–870
Beran MJ, Savage-Rumbaugh ES, Pate JL, Rumbaugh DM (1999) Delay of gratification in chimpanzees (Pan troglodytes). Dev Psychobiol 34:119–127
Beran MJ, Evans TA, Hoyle D (2011) Numerical judgments by chimpanzees (Pan troglodytes) in a token economy. J Exp Psychol Anim Behav Proc 37:165–174
Berns GS, Laibson D, Loewenstein G (2007) Intertemporal choice-toward an integrative framework. Trends Cogn Sci 11:482–488
Bourjade M, Thierry B, Call J, Dufour V (2012) Are monkeys able to plan for future exchange? Anim Cogn 15:783–795
Boysen ST, Berntson GG (1995) Responses to quantity: perceptual versus cognitive mechanisms in chimpanzees (Pan troglodytes). J Exp Psychol Anim Behav Proc 21:82–86
Brosnan SF, Beran MJ (2009) Trade between conspecifics in chimpanzees (Pan troglodytes). J Comp Psychol 123:181–194
Brosnan SF, de Waal FBM (2003) Monkeys reject unequal pay. Nature 425:297–299
Brosnan SF, de Waal FBM (2004) Socially learned preferences for differentially rewarded tokens in the brown capuchin monkey (Cebus apella). J Comp Psychol 118:133–139
Drapier M, Chauvin C, Dufour V, Uhlrich P, Thierry B (2005) Food-exchange with humans in brown capuchin monkeys. Primates 46:241–248
Dufour V, Pelé M, Sterck EHM, Thierry B (2007) Chimpanzee (Pan troglodytes) anticipation of food return: coping with waiting time in an exchange task. J Comp Psychol 121:145–155
Dufour V, Wascher CAF, Braun A, Miller R, Bugnyar T (2012) Corvids can decide if a future exchange is worth waiting for. Biol Lett 8:201–204
Evans TA, Beran MJ (2007a) Chimpanzees use self-distraction to cope with impulsivity. Biol Lett 3:599–602
Evans TA, Beran MJ (2007b) Delay of gratification and delay maintenance by rhesus macaques (Macaca mulatta). J Gen Psychol 134:199–216
Evans TA, Beran MJ, Paglieri F, Addessi E (2012) Delaying gratification for food and tokens in capuchin monkeys (Cebus apella) and chimpanzees (Pan troglodytes): when quantity is salient, symbolic stimuli do not improve performance. Anim Cogn 15:539–548
Grosch J, Neuringer A (1981) Self-control in pigeons under the Mischel paradigm. J Exp Anal Behav 35:3–21
Hackenberg TD, Vaidya M (2003) Determinants of pigeons’ choices in token-based self-control procedures. J Exp Anal Behav 79:207–218
Hillemann F, Bugnyar T, Kotrschal K, Wascher CAF (2014) Waiting for better, not for more: corvids respond to quality in two delay maintenance tasks. Anim Behav 90:1–10
Jackson K, Hackenberg TD (1996) Token reinforcement, choice, and self-control in pigeons. J Exp Anal Behav 66:29–49
Judge PG, Essler J (2013) Capuchin monkeys exercise self-control by choosing token exchange over an immediate reward. Int J Comp Psychol 26:256–266
Leonardi RJ, Vick SJ, Dufour V (2012) Waiting for more: the performance of domestic dogs (Canis familiaris) on exchange tasks. Anim Cogn 15:107–120
Logue AW (1988) Research on self-control: an integrating framework. Behav Brain Sci 11:665–709
Mischel W (2014) The marshmallow test: mastering self-control. Little, Brown and Company, New York
Mischel W, Shoda Y, Peake PK (1988) The nature of adolescent competencies predicted by preschool delay of gratification. J Pers Soc Psychol 54:687–696
Moffitt TE, Arseneault L, Belsky D, Dickson N, Hancox RJ, Harrington H, … Caspi A (2011) A gradient of childhood self-control predicts health, wealth, and public safety. Proc Nat Acad Sci 108:2693–2698
Parrish AE, Beran MJ (2014a) When less is more: like humans, chimpanzees (Pan troglodytes) misperceive food amounts based on plate size. Anim Cogn 17:427–434
Parrish AE, Beran MJ (2014b) Chimpanzees sometimes see fuller as better: judgments of food quantities based on container size and fullness. Behav Proc 103:184–191
Parrish AE, Evans TE, Perdue BM, Beran MJ (2013) Chimpanzees (Pan troglodytes) transfer tokens repeatedly with a partner to accumulate rewards in a self-control task. Anim Cogn 16:627–636
Parrish AE, Perdue BM, Stromberg EE, Bania AE, Evans TA, Beran MJ (2014) Delay of gratification by orangutans (Pongo pygmaeus) in the accumulation task. J Comp Psychol 128:209–214
Pelé M, Dufour V, Micheletta J, Thierry B (2010a) Long-tailed macaques display unexpected waiting abilities in exchange tasks. Anim Cogn 13:263–271
Pelé M, Thierry B, Call J, Dufour V (2010b) Monkeys fail to reciprocate in an exchange task. Anim Cogn 13:745–751
Pelé M, Micheletta J, Uhlrich P, Thierry B, Dufour V (2011) Delay maintenance in tonkean macaques (Macaca tonkeana) and brown capuchin monkeys (Cebus apella). Int J Primatol 32:149–166
Rachlin H, Green L (1972) Commitment, choice and self-control. J Exp Anal Behav 17:15–22
Ramseyer A, Pelé M, Dufour V, Chauvin C, Thierry B (2006) Accepting loss: the temporal limits of reciprocity in brown capuchin monkeys. Proc Royal Soc London 273:179–184
Sayers K, Menzel CR (2012) Memory and foraging theory: chimpanzee utilization of optimality heuristics in the rank-order recovery of hidden foods. Anim Behav 84:795–803
Shoda Y, Mischel W, Peake PK (1990) Predicting adolescent cognitive and self-regulatory competencies from preschool delay of gratification: identifying diagnostic conditions. Dev Psychol 26:978–986
Steelandt S, Thierry B, Broihanne M, Dufour V (2012) The ability of children to delay gratification in an exchange task. Cogn 122:416–425
Stevens JR, Mühlhoff N (2012) Intertemporal choice in lemurs. Behav Proc 89:121–127
Stevens JR, Hallinan EV, Hauser MD (2005) The ecology and evolution of patience in two new world monkeys. Biol Letters 1:223–226
Stevens JR, Rosati AG, Heilbronner SR, Mühlhoff N (2011) Waiting for grapes: expectancy and delayed gratification in bonobos. Int J Comp Psychol 24:99–111
Tobin H, Chelonis JJ, Logue AW (1993) Choice in self-control paradigms using rats. Psychol Rec 43:441–454
Tobin H, Logue AW, Chelonis JJ, Ackerman KT, May JG (1996) Self-control in the monkey Macaca fascicularis. Anim Learn Behav 24:168–174
Acknowledgments
This research was supported by National Institutes of Health Grant HD060563, a 2CI Primate Social Cognition, Evolution & Behavior Fellowship and the Rumbaugh Fellowship from Georgia State University.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Ethical standards
All aspects of this research conformed to APA standards for the ethical treatment of animals and followed the Institute of Science guidelines for ethical research with chimpanzees.
Conflict of interest
The authors declare that they have no conflicts of interest.
Rights and permissions
About this article

Cite this article
Beran, M.J., Rossettie, M.S. & Parrish, A.E. Trading up: chimpanzees (Pan troglodytes) show self-control through their exchange behavior. Anim Cogn 19, 109–121 (2016). https://doi.org/10.1007/s10071-015-0916-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-015-0916-7