
Abstract
Animals commonly face choices requiring them to wait and postpone action. The ability to delay gratification is a prerequisite for making future-oriented decisions. We investigated the ability of brown capuchins (Cebus apella) and Tonkean macaques (Macaca tonkeana) to delay benefits in several experiments. In exchange tasks, subjects had to return a piece of cookie after a given time lag to obtain a larger one from an experimenter. Capuchins could wait 10–40 s and macaques 20–80 s depending on subjects and the size of rewards. Both groups were able to anticipate delay durations, but unlike macaques, capuchins discounted all sizes of reward at the same speed, meaning that their delay-maintenance was not affected by the reward size. When the subjects could give the initial piece of cookie back immediately and then wait for the return, performances increased to 10–21 min for capuchins and 21–42 min for macaques, demonstrating the role of consumption inhibition in postponing gratification. In a further task, we presented subjects with an accumulation of food pieces added at short intervals until they seized them. On average, brown capuchins could wait 33–42 s and macaques 38–72 s before seizing the rewards. Our results confirmed that brown capuchins were more impulsive than Tonkean macaques in several tasks. We did not find significant differences between the waiting performances of the Tonkean macaques and those previously reported in long-tailed macaques. The contrasting performances of macaques and capuchins might be related to their different skills in the physical and social domains.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Individuals commonly face decisions requiring them to delay action and wait. The ability to delay gratification is crucial for complex goal-directed action and planning (Kacelnik 2003; Mischel 1974). In humans, Mischel (1974) measured the ability of children to forgo an immediate, less preferred reward for a future, more preferred one. He offered them a choice between 1 marshmallow available immediately and 2 marshmallows available only after some waiting. To obtain the larger reward, subjects first needed to choose waiting (delay-choice) and then to bridge the delay interval by maintaining their decision (delay-maintenance). Depending on their age and the type of instruction received, children either demonstrated patience for >15 min, or were impulsive and ate the marshmallow without waiting (Mischel 1974).
Researchers have studied delay of gratification in animals via similar procedures. In delay-choice tasks, subjects are given a choice between an immediate reward and a delayed but more valued one. When subjects opt for the delayed option, they have no further opportunity to modify their responses during the entire duration of the trial. In such tasks, domestic fowl (Gallus gallus), pigeons (Columbia livia), and rats (Rattus norvegicus) opted for the small immediate reward when the delay exceeded a few seconds (Abeyesinghe et al. 2005; Mazur 1987; Richards et al. 1997). Marmosets (Callithrix jacchus) and tamarins (Saguinus oedipus) postponed gratification for 5–20 s (Stevens et al. 2005a), whereas long-tailed macaques (Macaca fascicularis) and brown capuchins (Cebus apella) waited ca. 30 s (Amici et al. 2008), and spider monkeys (Ateles geoffroyi), bonobos (Pan paniscus), and chimpanzees (Pan troglodytes) between 1 and 2 min (Amici et al. 2008; Rosati et al. 2007).
In delay-maintenance tasks, subjects have the possibility to alter their choice at any time during the waiting period (Toner and Smith 1977; Toner et al. 1979). The ability to postpone gratification is then measured as the duration of time a subject refrains from making an impulsive response during a trial. In such tasks, pigeons (Grosch and Neuringer 1981) and rats (Killeen et al. 1981) waited for several seconds. In nonhuman primates, rhesus macaques (Macaca mulatta) waited for 2 min to obtain preferred food (Evans and Beran 2007a) and chimpanzees up to 18 min (Evans and Beran 2007b).
A further task consists of presenting subjects with food items accumulating at regular time intervals—usually seconds—within reachable distance of the subject. Once the subject reaches for the accumulated pile, no further items are presented. As in previous tasks, the subject must restrain food taking for as long as possible to maximize its gain. In this food-accumulation task, on average macaques did not delay gratification beyond 15–60 s (Evans and Beran 2007a; Pelé et al. 2010) and chimpanzees and orangutans beyond 2–3 min (Beran 2002).
Nonhuman primates can readily engage in exchanges of valuable goods with humans, which creates situations in which we can examine their skills in comparing costs and benefits (Addessi et al. 2007, 2008; Brosnan and de Waal 2004, 2005; Chalmeau and Peignot 1998; Drapier et al. 2005; Hyatt and Hopkins 1998; Lefebvre 1982; Westergaard et al. 2004). The ability to delay benefits appears to be a basic capacity needed for economic transactions, as there is often a delay to the benefits in comparison to costs in reciprocal interactions. Individuals must be able to bridge the time interval between the gift and the return. We previously investigated this ability directly in an exchange task wherein subjects were required to return a piece of cookie after a given time lag to obtain a larger one from an experimenter. When offered food rewards 40 times the size of the initial piece, brown capuchins (Cebus apella) did not wait >20–40 s before returning food (Ramseyer et al. 2006), whereas chimpanzees postponed returns for up to 4–8 min (Dufour et al. 2007). Tested in the same conditions, most long-tailed macaques tolerated time lags of ca. 2–5 min, but a few of them were able to postpone returns up to 10–21 min in several trials (Pelé et al. 2010).
In the food-exchange task, subjects must assess the value of rewards in relation to incurred delays. Analysis of the timing of failure to return food in chimpanzees and long-tailed macaques showed that individuals anticipated the delay times and decided accordingly whether to wait or give up (Dufour et al. 2007; Pelé et al. 2010). Moreover, temporal discounting—whereby the subjective value of benefits declines with time—could also affect choices when current and future rewards are in conflict (Mazur 1987; Stevens et al. 2005b). This decrease in value may occur because of the increased risk involved in waiting for rewards, or because of the decreased rate of reward associated with increased waiting time (Green and Myerson 1996). Another feature of the food-exchange task is that subjects must restrain their impulse to consume an available food. When the food-keeping condition was relaxed by asking the subjects to return the initial item before applying the time lag, long-tailed macaques doubled their waiting performances up to 40 min (Pelé et al. 2010). It is noteworthy that the waiting duration sustained by animals depends on the task and the experimental procedure. In delay-maintenance tasks, for instance, animals show better performances than in tasks in which they cannot modify their response. Having some degree of control over the end time of the trial in delay-maintenance tasks might make the situation more predictable and safer for the individuals than delay-choice tasks. This makes it necessary to test subjects in different experimental situations to appreciate their abilities better.
According to Amici et al. (2008), action is motivated more by immediate goals and interests in species living in cohesive groups, e.g., macaques and capuchins, than in species undergoing frequent splitting and merging of subgroups, e.g., chimpanzees and orangutans. By favoring behavioral flexibility, social dynamics based on fission and fusion would enhance inhibitory skills in the latter species. However, we found comparable waiting performances in chimpanzees and long-tailed macaques in the food-exchange task, whereas capuchins sustained significantly lower time lags than the previous 2 species (Dufour et al. 2007; Pelé et al. 2010; Ramseyer et al. 2006).
We submitted brown capuchins and Tonkean macaques (Macaca tonkeana) to different conditions of delay maintenance in the food-exchange task and the food-accumulation task to test the robustness of our previous findings and extend them to another macaque species. Tonkean macaques display higher levels of social tolerance than long-tailed macaques and they are characterized by more relaxed dominance relationships (Thierry 2007). We investigated whether their ability to delay gratification differed significantly from that measured in long-tailed macaques.
General Methods
Subjects
We tested 10 Tonkean macaques and 9 brown capuchins maintained at the Primatology Centre of the University of Strasbourg, France. The Tonkean macaques belonged to 3 different social groups. Four subjects (Rim, She, Sim, Syb) were housed together in an indoor-outdoor enclosure of 15 m2. Two subjects (Jan, Mil) were housed together in an indoor-outdoor enclosure of 15 m2. Four subjects (Gae, Lad, Olg, Sha) belonged to a social group of 12 individuals living in a 0.5-ha park. The capuchins belonged to a social group of 18 individuals and were housed in an indoor-outdoor enclosure of 70 m2. Commercial monkey diet pellets and water were provided ad libitum. Subjects were never deprived of food. For testing, subjects were temporarily separated from their conspecifics. The age and sex of subjects are provided in Table I.
Some data for capuchins are from Ramseyer et al. (2006) for Experiments 1 and 2 (subjects Acc, Arn, Aso, Bib, Clo, Pis). One capuchin (Bib) died before Experiment 2 and was tested in Experiment 1 only. Another (Clo) left the Primatology Centre before Experiment 3 and was tested in Experiments 1 and 2 only. We tested 3 additional capuchins (Pet, Pop, Rav) to analyze the time of giving up in Experiments 1 and 2.
Exchange Procedure
We conducted Experiments 1–3 using food-exchange tasks. Before these experiments, subjects had been trained to exchange food items with humans (Electronic Supplementary Material [ESM 1]). During testing, the experimenter stood in front of the subject’s compartment, showing the subject 2 different sized pieces of cookie in each hand for 3 s. The experimenter then gave the smaller piece to the subject. The larger piece, i.e., the reward, remained visible in the experimenter’s second hand for the duration of the waiting period. After the waiting period had elapsed, the experimenter held out her empty hand while still presenting the reward in her other hand. If the subject gave back the small piece of cookie, he received the larger one. If the subject did not give back the initial piece, the experimenter did not reward the subject and stepped back, thereby ending the trial. Whenever the individual had dropped or eaten the cookie before the end of the waiting period, the experimenter ended the trial by stepping back. The experimenter waited for 30 s after the end of food consumption before beginning another trial.
Statistics
We used the Friedman test to compare the percentages of return for the different sizes of cookies at each time lag in Experiment 1, the Wilcoxon matched-pairs test to compare percentages of return in Experiments 2 and 3, and the Mann-Whitney test to compare performances of capuchins and macaques in the different experiments (Siegel and Castellan 1988). Given the small number of individuals, we applied the exact test procedure of SPSS 16©.
We compared the distribution of observed giving-up times—the time at which capuchins and macaques gave up waiting before the end of the trial—with the distribution of times to be expected under the null hypothesis of a constant giving-up chance during the trial. If capuchins and macaques anticipate the duration of the delay that remained, they should decide early on whether to wait or not. To test this hypothesis, we calculated via Kaplan Meier survival analysis the estimated (observed) probability of continuing to wait at each point of time that the subject gave up waiting. This analysis included both the failure times and the times of successful return at the end of the trial (as censored data). The survival probability was expressed as percentage of chance to wait longer than the time already elapsed in the trial. Against this observed survival function, we calculated the expected (exponential) distribution of giving up times under the null hypothesis of a constant chance of giving up. We compared the expected and the observed distributions using an adjusted Kolmogorov–Smirnov test (Haccou and Meelis 1992). A statistically significant difference between both distributions leads to the rejection of the null hypothesis that a subject has a constant chance to give up independent of the time elapsed in the trial. We performed the analysis on Experiment 1 by combining all cookie sizes: 2, 4, and 8 times the original size (N = 48). In Experiment 2, we tested only 1 reward size; therefore, the analysis was performed on a smaller number of events (N = 12).
We used linear regression analyses to assess relationships between the percentage of return and time. To study the influence of temporal discounting on the subjects' abilities to wait, we contrasted the decreasing rates of returns as a function of time for each reward size in Experiments 1 and 2 using comparison tests of regression slopes.
Data were processed via SPSS 16.0 (SPSS Inc., Chicago, IL, USA) and R (The R Foundation for Statistical Computing, Vienna, Austria). The significance level was set at 0.05. Average values are given in the text and figures as means and SEM.
Experiment 1: Testing the Effect of Quantity on Waiting Periods
We tested whether the duration of the waiting period sustained by Tonkean macaques and brown capuchins would vary according to the relative value of the items being exchanged.
Methods
Subjects were 10 Tonkean macaques and 9 brown capuchins. For each trial, the experimenter first offered a small piece of cookie (1 × 2 × 0.5 cm), then presented the subject with a larger piece of cookie: 2 × 2 × 0.5 cm (2-fold the size of the initial item), 2 × 4 × 0.5 cm (4-fold), or 4 × 4 × 0.5 cm (8-fold). The experimenter tested subjects in a succession of stages comprising 4 sessions in total. Each stage differed by the time lag tested: 2, 5, 10, 20, 40 s, etc. Each session comprised 12 trials in which each of the 3 possible cookie sizes was presented 4 times in random order. The experimenter tested successive time lags until the individual performance dropped to 0% of return. Subjects received 1 session per half-day. The experimenter recorded failure to return the initial cookie as well as its timing.
Results
Percentage of Return
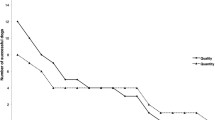
In Experiment 1, most of the macaques succeeded in exchanging until 40 s of delay. Only 4 subjects (Gae, Olg, Sha, She) stopped exchanging at lesser time lags. Two macaques (Lad, Sim) stopped returning the initial item at 80 s and 2 others (Mil, Syb) at 160 s of delay. The last 2 subjects (Jan, Rim) still returned the small item at 2 min 40 s and stopped at 5 min 20 s (ESM 1). All capuchins succeeded until 10 s of delay. Five subjects (Arn, Aso, Clo, Pis, Pop) stopped exchanging at 20 s. Two others (Acc, Bib) stopped returning the initial item at 40 s and 1 (Rav) at 80 s. A last subject (Pet) still returned the small item at a time lag equal to 80 s and stopped at 160 s (ESM 1). In comparing the longest time lag sustained at least once by each subject in both groups, we found that macaques waited longer than capuchins did (Mann-Whitney test, z = 2.14, p = 0.034, N 1 = 10, N 2 = 9, \( {m_{{1}}} = {63}\pm {\hbox{18 s}} \), \( {m_{{2}}} = {23}\pm {\hbox{8 s}} \)). The waiting periods of macaques significantly increased with the size of the reward at time lags of 2 s (Friedman test, df = 2, p = 0.016), 5 s (p = 0.0002), 10 s (p = 0.0004), 20 s (p = 0.0003), 40 s (p = 0.005), and 80 s (p = 0.022) (Fig. 1). The waiting periods of capuchins significantly increased with size at 5 s (Friedman test, df = 2, p = 0.0006), 10 s (p = 0.0086), and 20 s (p = 0.038) (Fig. 1).

Percentages of return for each size of cookie (2, 4, and 8 times) at each time lag in Experiment 1 for macaques (N = 10) and capuchins (N = 9). Friedman test, *p < 0.05, **p < 0.01, ***p < 0.001.
Time of Giving Up
The size of the reward affected the waiting time of subjects. To test whether subjects decided early on whether to wait or not, we compared the distribution of observed giving-up times of macaques with the distribution of times to be expected under the null hypothesis of a constant giving-up chance during the trial. All but 1 subject (She) gave up earlier than predicted by the model; from a time lag of 5 s for Rim, Sha; 10 s for Gae, Jan; 20 s for Lad, Mil, Olg, Sim; and 40 s for Syb (Table II). In capuchins —those tested only in this study—the 3 subjects gave up significantly earlier than predicted, from a time lag of 5 s for Pop, 10 s for Pet, and 40 s for Rav (Table II).
Experiment 2: Assessing Maximal Waiting Periods
In this experiment, we considerably increased the final pay-off to assess whether macaques and capuchins could sustain longer waiting periods than those measured in the previous experiment.
Methods
Subjects were 10 Tonkean macaques and 8 brown capuchins. The experimenter offered subjects a reward 40-fold the size of the initial food item: 5 pieces of cookie, each 4 × 4 × 0.5 cm. The experimenter tested each subject in 12 trials for each time lag: 10, 20, 40, 80 s, etc. Considering the larger size of the reward, we reduced sessions to 2 trials per day, separated by a minimum 20-min interval. If a subject succeeded at least once in a session, we tested it at the next time lag. As in Experiment 1, the experimenter recorded the timing of failures.
Results
Percentage of Return
Compared with Experiment 1, subjects of both species increased their performances. All macaques still returned the initial item until a time lag of 40 s. The percentage of return of 4 individuals (Gae, Olg, Sha, She) dropped to 0% for a time lag of 80 s. Five other individuals (Jan, Mil, Rim, Sim, Syb) stopped exchanging at 160 s of delay. A last individual (Lad) continued to return the initial item until 160 s and stopped at 320 s (ESM 1). In capuchins, 2 subjects (Pis, Pop) stopped exchanging the initial item at 20 s of delay. Three subjects (Arn, Aso, Clo) stopped returning the small piece of cookie at 40 s and 2 others (Acc, Rav) at 80 s. A last individual (Pet) continued to exchange the initial item until 160 s and stopped for a time lag of 320 s (ESM 1). The comparison of subjects' performances in both groups showed that macaques sustained longer time lags than capuchins did (Mann-Whitney test, z = 2.43, p = 0.040, N 1 = 10, N 2 = 8, \( {m_{{1}}} = {72}\pm {\hbox{12 s}} \), \( {m_{{2}}} = {4}0\pm {\hbox{18 s}} \)).
Time of Giving Up
Four macaques (Gae, She, Sim, Syb) never gave up earlier than predicted by the model (Table III). The other 6 macaques gave up significantly earlier than predicted by the model; Olg and Sha at the 80-s time lag; Jan, Mil and Rim at 160 s, and Lad at 5 min 20 s (Table III). All capuchins (Pet, Pop, Rav) gave up significantly earlier than predicted from a time lag of 20 s for Pop, 40 s for Rav and 80 s for Pet (Table III).
We calculated the extinction of exchanges for each individual by examining the waiting time per trial for the 3 longest time lags it sustained (ESM 1). Ten subjects (macaques: Jan, Lad, Olg, Rim, Sha, She, Sim, Syb; capuchins: Pop, Rav) gradually stopped exchanging as delays were extended. The other 3 subjects (macaques: Gae, Mil; capuchin: Pet) displayed different patterns; they resumed exchanges after some failures in several consecutive trials. In this experiment, both macaques and capuchins sustained especially long periods when offered a reward 40 times the size of the initial item.
Temporal Discounting
Engaging with a partner in a delayed exchange incurs the risk that the partner will not return the gift. As the delay to the outcome increases, the subjects' return expectations may decrease. Temporal discounting appears as a possible response to the risk associated with waiting for delayed rewards (Kagel et al. 1986). To check for the possible influence of temporal discounting on the subjects' abilities to wait, we contrasted the decreasing rates of return as a function of time for each reward size in Experiments 1 and 2 using comparison tests of regression slopes (Fig. 2). Macaques neither devalued rewards 2-fold the size of the initial item more rapidly than 4-fold rewards (t = –1.88, df = 5, p = 0.118), nor did they devalue 8-fold rewards more rapidly than 40-fold rewards (t = 0.15, df = 8, p = 0.880). However, they devalued rapidly 4-fold rewards significantly more than 8-fold (t = –2.91, df = 8, p = 0.019) and 40-fold rewards (t = –3.11, df = 6, p = 0.020). Capuchins devalued 2-fold rewards at the same speed as 4-fold (t = 0.25, df = 4, p = 0.810) and 8-fold rewards (t = –0.04, df = 4, p = 0.968). They also devalued 4-fold rewards at the same speed as 8-fold rewards (t = –0.42, df = 4, p = 0.692). Capuchins’ devaluation of 40-fold rewards was slower compared to 2-fold (t = –5.17, df = 3, p = 0.014), 4-fold (t = –8.21, df = 3, p = 0.003), and 8-fold rewards (t = –5.13, df = 3, p = 0.014).

Linear regressions of the percentage of return with time for each reward size in Experiments 1 and 2 for each species.
Experiment 3: Testing the Effect of Action Inhibition on Waiting
In Experiments 1 and 2, macaques and capuchins needed to decide whether to exchange or not. They also had to keep the small piece of cookie intact throughout the waiting period, thus inhibiting the consumption of the initial item. To test how refraining from action may have affected their performances, we relaxed the food-keeping condition and asked that subjects give back the initial food item 3 s after having received it. Only then was the waiting period applied. Once it ended, the experimenter gave the larger reward.
Methods
Subjects were 10 Tonkean macaques and 7 brown capuchins. We applied the same procedure as in Experiment 2. The experimenter tested subjects in 2 daily trials separated by a 20-min interval. Once subjects had returned the initial piece of cookie, the experimenter placed it beside the larger reward (40-fold the initial piece), in front but out of reach of the subjects. Then, the experimenter started the waiting period: 10, 20, 40, 80 s, etc. If subjects failed to return the initial item, the trial ended. If they succeeded, they obtained the larger reward at the end of the time lag.
Results
Percentage of Return
In Experiment 3, all macaques but 1 (She) waited until a time lag of 640 s. Four of them (Jan, Sha, Sim, Syb) stopped waiting at 1280 s. Lad stopped exchanging at 2560 s, whereas the last 4 subjects (Gae, Mil, Olg, Rim) continued to wait at this time lag (ESM 1). All capuchins were able to wait for ≥40 s. Two of them (Aso, Pop) stopped exchanging at 80 s and 320 s, respectively. The other 5 subjects waited until 640 s; 2 (Pet, Rav) stopped waiting at a time lag of 1280 s, and the other 3 (Acc, Arn, Pis) at 2560 s (ESM 1). Comparison of subjects' performances in Experiments 2 and 3 showed that the suppression of the food-keeping condition in the latter experiment significantly increased waiting durations for both species (Fig. 3). The time lags sustained by subjects did not differ significantly between macaques and capuchins (Mann-Whitney test, z = 1.21, p = 0.220, N 1 = 10, N 2 = 7, \( {m_{{1}}} = {1424}\pm {32}0\,{\hbox{s}} \), \( {m_{{2}}} = {76}0\pm {2}0{\hbox{2 s}} \)).

Percentage of return at each time lag in Experiments 2 and 3 for macaques (N = 10) and capuchins (N = 7). Wilcoxon test, *p < 0.05, **p < 0.01.
Temporal Discounting
Figure 4 presents the decreasing rates of return as a function of time in Experiments 2 and 3. In macaques, comparing the slope of the curve obtained for Experiment 3 to a null slope did not yield a statistically significant difference (t = –1.6, p = 0.215), which indicates that once the constraint of consumption inhibition is removed, macaques still valued the reward regardless of the delay period. We found no effect of temporal discounting on waiting performances of macaques. By contrast, comparison of the slope of the curve obtained for Experiment 3 with a null slope in capuchins produced a significant difference (t = –16.3, p = 0.038), which indicates that the subjects declined to exchange depending on the waiting duration despite the fact that consumption inhibition was unnecessary. To test whether capuchins differently devalued the rewards expected in Experiments 2 and 3, we compared the slopes of 2 linear regressions. A statistically significant difference appeared in devaluation rates between both experiments (t = 6.2, df = 2, p = 0.024).

Linear regressions of the return percentage with time in Experiments 2 and 3 for each species.
Experiment 4: Testing the Ability to Wait in an Accumulation Task
We found different waiting periods in macaques and capuchins in several conditions of the food-exchange task. To assess their performances in a different delay-maintenance test, we studied subjects in an accumulation task wherein food pieces were added at fractioned time intervals (Beran 2002). Brunner and Gibbon (1995) suggested that subjects could treat such sequences of food items not as a whole but as a collection of independent items. If subjects consider 1 interval at a time they should wait for longer periods than for nonfractioned periods.
Methods
Training
Subjects were 10 Tonkean macaques and 7 brown capuchins. The apparatus consisted of 1 PVC panel comprised of 2 tablets, 1 and 2, separated by 5 cm, and small (1 × 2 × 0.5 cm) pieces of cookie. The first training stage consisted of a demonstration of the transfer of 5 pieces of cookie, 1 at a time, at a rate of 1 every 3 s, from tablet 1 to tablet 2; both tablets were out of reach for subjects. Once the 5 pieces of cookie were on tablet 2, the experimenter moved the panel closer to subjects and allowed them to take the food items. A session was composed of 6 demonstrations separated by 2 min after the end of food consumption by the subject. We conducted 4 sessions at this stage, with ≤2 daily sessions separated by an interval ≥20 min long. In the second training stage, the experimenter alternated 6 demonstration trials with 6 training trials; in the latter, subjects had access to tablet 2, i.e., they could take items at any time, thus ending the trial. As previously, the experimenter conducted 4 sessions with ≤2 daily sessions separated by an interval of ≥20 min. In the third stage, demonstrations ceased and sessions consisted of 6 training tests, with items transferred from one tablet to another every 3 s. The experimenter waited for 30 s after the end of the subject’s food consumption before starting another trial. A minimum of 3 pieces of cookie per trial and 20 pieces of cookie per session during 2 consecutive sessions was required to proceed to the next training stage. In the final stage, the experimenter trained subjects as in stage 3 and using the same criterion of success, except that items were transferred every 6 s. If individuals had not reached success criteria after 20 training sessions, we stopped training them.
Three capuchins (Pet, Pis, Rav) and 6 Tonkean macaques (Jan, Lad, Mil, Rim, Sha, Syb) successfully passed the training criteria. Four Tonkean macaques (Gae, Olg, She, Sim) did not reach the 3-s success criterion. At such a time lag, they waited for a maximum of 5, 4, 4, and 4 pieces of cookie, respectively. Among brown capuchins, 3 (Acc, Aso, Pop) did not reach the 3-s success criterion at 3 s, and 1 (Arn) the 6 s criterion; for 3 s, Acc waited for a maximum of 2 pieces of cookie; Arn, 10, Aso, 1, and Pop, 1, respectively.
Testing
Subjects were 6 Tonkean macaques and 3 brown capuchins. During the test, the experimenter placed the panel in front of the subject, with tablet 2 remaining within its reach, and tablet 1 loaded with 81 pieces of cookie out of reach. Every 9 s, the experimenter transferred a piece of cookie from tablet 1 to tablet 2. Subjects were allowed to take the items on tablet 2 at any time. If they did, the trial ended. We conducted daily sessions of 2 trials/d separated by a 20-min interval. We submitted each subject to a total of 60 trials.
Results
On average, Tonkean macaques could wait between 38 and 72 s and brown capuchins between 33 and 42 s before seizing the rewards. The maximal waiting periods recorded were 351 s for macaques and 126 s for capuchins (Fig. 5). They did not significantly differ between macaques and capuchins (Mann-Whitney test, z = 0.38, p = 0.760, N 1 = 6, N 2 = 3, \( {m_{{1}}} = {14}0\pm {\hbox{45 s}} \), \( {m_{{2}}} = {93}\pm {\hbox{16 s}} \)). In Experiment 4, the value in time of 1 unit of cookie, i.e. per cm2, was equal to 4.5 s. In Experiment 1, macaques waited on average 3.93 ± 0.39 s/cm2 and capuchins 2.25 ± 0.48 s/cm2. In macaques fractioning time periods led to performances that were comparable to those measured in Experiment 1 using similar reward amounts. In capuchins the waiting periods of tested subjects were 2 or 4 times longer than those measured in Experiment 1 for similar reward amounts.

Mean number of food items consumed by subjects of both species by series of 10 trials in Experiment 4. Maxima are indicated by black crosses.
Discussion
When exchanging with an experimenter, capuchins and Tonkean macaques sustained longer waiting periods for larger food quantities, consistent with the results of previous studies in long-tailed macaques, capuchins, and chimpanzees (Dufour et al. 2007; Pelé et al. 2010; Ramseyer et al. 2006). Such a magnitude effect was not found in rats and pigeons, nor in marmosets and tamarins, when tested in a delay-choice task (Green et al. 2004; Richards et al. 1997; Stevens et al. 2005a).
In Experiment 1 most Tonkean macaques waited up to 20–80 s for rewards 8 times the size of the initial food item, which compares well with performances of long-tailed macaques under the same conditions (Pelé et al. 2010). In brown capuchins, although 1 subject waited sometimes for 40 s, the addition of 3 subjects to our initial sample did not alter our conclusion that the waiting duration of most subjects did not exceed 10–20 s for the same reward size (Ramseyer et al. 2006). In Experiment 2, an increase in reward quantity to 40 times the size of the initial food item led both capuchins and Tonkean macaques to double their waiting periods to 20–40 s and 40–80 s, respectively. These performances were again consistent with previous measurements in brown capuchins (Ramseyer et al. 2006). In contrast, most long-tailed macaques were able to postpone returns for 2–5 min (Pelé et al. 2010).
A majority of Tonkean macaques and brown capuchins gave up earlier than predicted under a model of constant rate of giving up in Experiments 1 and 2. It is likely that they anticipated waiting durations in relation to the size of final rewards; they reversed their choice, preferring to consume the smaller reward early in trials. An alternative explanation would involve only associative learning; longer delays would heighten the difficulty of associating the initial exchange acceptance and the final reward, resulting in an extinction of returns (Dufour et al. 2007). However, in Experiment 2, 2 macaques and 1 capuchin succeeded in waiting for long delays after consecutive failures, which does not support the hypothesis of associative learning. It thus appears that Tonkean macaques and brown capuchins can anticipate the duration they had to wait for, and decide accordingly whether to wait or give up, as do long-tailed macaques (Pelé et al. 2010) and chimpanzees (Dufour et al. 2007).
In Experiment 3, we tested the hypothesis that consumption inhibition constrained the subjects' abilities to postpone gratification by asking subjects to return the initial item before waiting. As in a delayed-reward contingency task, individuals faced a single decision: to accept the exchange or not. They could not modify their choice within a given trial, but they could reevaluate it in the next one. Both Tonkean macaques and capuchins dramatically improved their maximum waiting time compared to Experiment 2 for identical reward amounts, demonstrating that the inhibition of food consumption is a limiting factor for the subjects' abilities to postpone gratification (Pelé et al. 2010). Several brown capuchins accepted a time lag of 10 min 40 s, and some tolerated a time lag of 21 min 20 s. Most Tonkean macaques could sustain the latter durations, and half of them were even able regularly to sustain a waiting period of 42 min 40 s. By contrast, most long-tailed macaques did not accept time lags longer than 21 min 20 s (Pelé et al. 2010).
The study of temporal discounting highlighted differences between Tonkean macaques and brown capuchins in the maintenance of their decision to wait. The former discounted small rewards faster than bigger ones, indicating that their subjective devaluation of future rewards was proportionally affected by the size of rewards. By contrast, capuchins discounted all rewards at the same speed in Experiment 1, which showed that the maintenance of the decision to wait was not affected by the size of rewards. Moreover, there was no more temporal discounting in Tonkean macaques when relaxing the need to inhibit food consumption in Experiment 3. By contrast, capuchins still devalued the reward even though they did not have to inhibit eating. They consistently behaved with more immediacy than macaques did.
In the food-accumulation task of Experiment 4, Tonkean macaques tended to wait longer than brown capuchins did, with the caveat that several individuals from both species did not reach the criterion needed to be tested. The performances reported in rhesus and long-tailed macaques were also lower than those measured in Tonkean macaques (Evans and Beran 2007a; Pelé et al. 2010). We assumed that fractioning the entire waiting period into a series of short intervals would minimize temporal discounting. When comparing the behavior of subjects for similar reward quantities, it appeared that in Experiment 4 subjects were satisfied with a number of food items, which was equivalent in Experiment 1 to the 8-fold reward in macaques and the 4-fold reward in capuchins. It was as if the subjects assessed the entire waiting duration together with the final reward instead of each time lag separately, indicating that their decision was affected by temporal discounting.
Capuchins appeared more impulsive than macaques in several tasks. We know of no obvious socioecological variations suitable as explanations for such interspecific differences. However, the propensity of capuchins to manipulate objects and use them as tools is noteworthy whereas their rates of social interactions appear relatively low compared to those of macaques (Anderson 1996; Fragaszy et al. 2004; Schino et al. 2009; Westergaard and Fragaszy 1987). If inhibitory skills are necessary to face uncertainties in social interactions (Amici et al. 2009; Pelé et al. 2010), it is understandable that the more socially oriented macaques can perform better in delay-maintenance tasks. Future research should investigate possible links between delaying gratification and the cognitive abilities respectively used in the physical and social domains.
Whereas intraspecific variation appeared weaker than interspecific variation when contrasting capuchins and macaques, we found the converse when comparing the performances of Tonkean macaques with those previously measured in long-tailed macaques. The individuals studied in each species differed by several factors, e.g., age, sex, and breeding conditions, and no consistent differences emerged between groups; long-tailed macaques outperformed Tonkean macaques in Experiment 2 but the converse was true in Experiments 3 and 4, and the results from both groups were similar in Experiment 1. Further investigation in a larger number of individuals and species should deepen our knowledge of interspecific variations in temperament and cognitive abilities among macaques. However, at present, we may conclude that although long-tailed and Tonkean macaques widely differ in their social relationships and levels of social tolerance (Aureli et al. 1997; Thierry 1986, 2007; Thierry et al. 2008), we did not find significant differences in their willingness to delay gratification. This is what we would expect under the hypothesis that all macaques possess similar abilities to cope with complex social situations.
References
Abeyesinghe, S. M., Nicol, C. J., Hartnell, S. J., & Wathes, C. M. (2005). Can domestic fowl, Gallus gallus domesticus, show self-control? Animal Behaviour, 70, 1–11.
Addessi, E., Crescimbene, L., & Visalberghi, E. (2007). Do capuchin monkeys (Cebus apella) use tokens as symbols? Proceedings of the Royal Society of London B: Biological Series, 274, 2579–2585.
Addessi, E., Crescimbene, L., & Visalberghi, E. (2008). Food and token quantity discrimination in capuchin monkeys (Cebus apella). Animal Cognition, 11, 275–282.
Amici, F., Aureli, F., & Call, J. (2008). Fission-fusion dynamics, behavioral flexibility and inhibitory control in primates. Current Biology, 18, 1415–1419.
Amici, F., Call, J., & Aureli, F. (2009). Variation in withholding of information in three monkey species. Proceedings of the Royal Society of London B: Biological Series, 276, 3311–3318.
Anderson, J. R. (1996). Chimpanzees and capuchin monkeys: comparative cognition. In A. E. Russon, K. A. Bard, & S. T. Parker (Eds.), Reaching into thought: The minds of the great apes (pp. 23–56). Cambridge: Cambridge University Press.
Aureli, F., Das, M., & Veenema, H. C. (1997). Differential kinship effect on reconciliation in three species of macaques (Macaca fascicularis, M. fuscata, and M. sylvanus). Journal of Comparative Psychology, 111, 91–99.
Beran, M. J. (2002). Maintenance of self-imposed delay of gratification by four chimpanzees (Pan troglodytes) and an orangutan (Pongo pygmaeus). The Journal of General Psychology, 129, 49–66.
Brosnan, S. F., & de Waal, F. B. M. (2004). A concept of value during experimental exchange in brown capuchin monkeys, Cebus apella. Folia Primatologica, 75, 317–330.
Brosnan, S. F., & de Waal, F. B. M. (2005). Responses to a simple barter task in chimpanzees, Pan troglodytes. Primates, 46, 173–182.
Brunner, D., & Gibbon, J. (1995). Value of food aggregates: parallel versus serial discounting. Animal Behaviour, 50, 1627–1634.
Chalmeau, R., & Peignot, P. (1998). Exchange of objects between human and captive western lowland gorillas. Primates, 39, 389–398.
Drapier, M., Chauvin, C., Dufour, V., Uhlrich, P., & Thierry, B. (2005). Food-exchange with humans in brown capuchin monkeys. Primates, 46, 241–248.
Dufour, V., Pelé, M., Sterck, E. H. M., & Thierry, B. (2007). Chimpanzee (Pan troglodytes) anticipation of food return: coping with waiting time in an exchange task. Journal of Comparative Psychology, 121, 145–155.
Evans, T. A., & Beran, M. J. (2007a). Delay of gratification and delay maintenance by rhesus macaques (Macaca mulatta). The Journal of General Psychology, 134, 199–216.
Evans, T. A., & Beran, M. J. (2007b). Chimpanzees use self-distraction to cope with impulsivity. Biology Letters, 3, 599–602.
Fragaszy, D. M., Visalberghi, E., & Fedigan, L. M. (2004). The complete capuchin: the biology of the genus Cebus. Cambridge: Cambridge University Press.
Green, L., & Myerson, J. (1996). Exponential versus hyperbolic discounting of delayed outcomes: Risk and waiting time. American Zoologist, 36, 496–505.
Green, L., Myerson, J., Holt, D. D., Slevin, J. R., & Estle, S. J. (2004). Discounting of delayed food rewards in pigeons and rats: is there a magnitude effect? Journal of the Experimental Analysis of Behavior, 81, 39–50.
Grosch, J., & Neuringer, A. (1981). Self-control in pigeons under the Mischel paradigm. Journal of the Experimental Analysis of Behavior, 35, 3–21.
Haccou, P., & Meelis, E. (1992). Statistical analysis of behavioural data. Oxford: Oxford University Press.
Hyatt, C. W., & Hopkins, W. D. (1998). Interspecies object exchange: bartering in apes? Behavioural Processes, 42, 177–187.
Kacelnik, A. (2003). The evolution of patience. In G. Loewenstein, D. Read, & R. Baumeister (Eds.), Time and decision: Economic and psychological perspectives on intertemporal choice (pp. 115–138). New York: Russell Sage Foundation.
Kagel, J. H., Green, L., & Caraco, T. (1986). When foragers discount the future: constraint or adaptation? Animal Behaviour, 34, 271–283.
Killeen, P. R., Smith, J. P., & Hanson, S. J. (1981). Central place foraging in Rattus norvegicus. Animal Behaviour, 29, 64–70.
Lefebvre, L. (1982). Food exchange strategies in an infant chimpanzee. Journal of Human Evolution, 11, 195–204.
Mazur, J. E. (1987). An adjusting procedure for studying delayed reinforcement: The effects of delay and intervening events on reinforcement value. In M. L. Commons, J. E. Mazur, J. A. Nevin, & H. Rachlin (Eds.), Quantitative analyses of behavior (Vol. 5, pp. 55–73). Hillsdale, NJ: Lawrence Erlbaum.
Mischel, W. (1974). Processes in delay of gratification. In L. Berkowitz (Ed.), Advances in experimental social psychology (pp. 249–292). New York: Academic Press.
Pelé, M., Dufour, V., Micheletta, J., & Thierry, B. (2010). Long-tailed macaques display unexpected waiting abilities in exchange tasks. Animal Cognition, 13, 263–271.
Ramseyer, A., Pelé, M., Dufour, V., Chauvin, C., & Thierry, B. (2006). Accepting loss: the temporal limits of reciprocity in brown capuchin monkeys. Proceedings of the Royal Society of London B: Biological Series, 273, 179–184.
Richards, J. B., Mitchell, S. H., de Wit, H., & Seiden, L. S. (1997). Determination of discount functions in rats with an adjusting-amount procedure. Journal of the Experimental Analysis of Behavior, 67, 353–366.
Rosati, A. G., Stevens, J. R., Hare, B., & Hauser, M. D. (2007). The evolutionary origins of human patience: temporal preferences in chimpanzees, bonobos, and human adults. Current Biology, 17, 1663–1668.
Schino, G., di Giuseppe, F., & Visalberghi, E. (2009). Grooming, rank, and agonistic support in tufted capuchin monkeys. American Journal of Primatology, 71, 101–105.
Siegel, S., & Castellan, N. J. (1988). Nonparametric statistics for the behavioral sciences. Singapore: McGraw-Hill.
Stevens, J. R., Hallinan, E. V., & Hauser, M. D. (2005). The ecology and evolution of patience in two New World monkeys. Biology Letters, 1, 223–226.
Stevens, J. R., Cushman, F. A., & Hauser, M. D. (2005). Evolving the psychological mechanisms for cooperation. Annual Review of Ecology and Systematics, 36, 499–518.
Thierry, B. (1986). A comparative study of aggression and response to aggression in three species of macaque. In J. G. Else & P. C. Lee (Eds.), Primate ontogeny, cognition, and social behaviour (pp. 307–313). Cambridge: Cambridge University Press.
Thierry, B. (2007). Unity in diversity: lessons from macaque societies. Evolutionary Anthropology, 16, 224–238.
Thierry, B., Aureli, F., Nunn, C., Petit, O., Abegg, C., & de Waal, F. B. M. (2008). A comparative study of conflict resolution in macaques: insights into the nature of trait covariation. Animal Behaviour, 75, 847–860.
Toner, I. J., & Smith, R. A. (1977). Age and overt verbalization in delay-maintenance behavior in children. Journal of Experimental Child Psychology, 24, 123–128.
Toner, I. J., Lewis, B. C., & Gribble, C. M. (1979). Evaluative verbalization and delay maintenance behavior in children. Journal of Experimental Child Psychology, 28, 205–210.
Westergaard, G. C., & Fragaszy, D. M. (1987). The manufacture and use of tools by capuchin monkeys (Cebus apella). Journal of Comparative Psychology, 101, 159–168.
Westergaard, G. C., Liv, C., Rocca, A. M., Cleveland, A., & Suomi, S. J. (2004). Tufted capuchins (Cebus apella) attribute value to foods and tools during voluntary exchanges with humans. Animal Cognition, 7, 19–24.
Acknowledgments
We thank N. Poulin and C. Sueur for statistical advice; A. Delabouglise, C. Dilger, P. Gayet, A. Jacquemin, C. Morin, A. Navarre, F. Vogelweith, M. Plasse, C. Pichon, F.Colas, C.Rosière, and F. Wehrle for their valuable assistance in experiments, and 2 anonymous reviewers whose thoughtful comments greatly improved a previous version of the manuscript. The research was supported by a grant from the Agence Nationale de la Recherche (ANR-08-BLAN-0042-01).
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
Below is the link to the electronic supplementary material.
ESM 1
(DOC 715 kb)
Rights and permissions
About this article
Cite this article
Pelé, M., Micheletta, J., Uhlrich, P. et al. Delay Maintenance in Tonkean Macaques (Macaca tonkeana) and Brown Capuchin Monkeys (Cebus apella). Int J Primatol 32, 149–166 (2011). https://doi.org/10.1007/s10764-010-9446-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10764-010-9446-y