
Abstract
Using viruses to our advantage has been a huge leap for humanity. Their ability to mediate horizontal gene transfer has made them useful tools for gene therapy, vaccine development, and cancer treatment. Adenoviruses, adeno-associated viruses, retroviruses, lentiviruses, alphaviruses, and herpesviruses are a few of the most common candidates for use as therapeutic agents or efficient gene delivery systems. Efforts are being made to improve and perfect viral-vector-based therapies to overcome potential or reported drawbacks. Some preclinical trials of viral vector vaccines have yielded positive results, indicating their potential as prophylactic or therapeutic vaccine candidates. Utilization of the oncolytic activity of viruses is the future of cancer therapy, as patients will then be free from the harmful effects of chemo- or radiotherapy. This review discusses in vitro and in vivo studies showing the brilliant therapeutic potential of viruses.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Viruses are the most abundant biological entities, with remarkable genetic diversity, substantial resilience to environmental changes, and significant ability to mutate and survive [68]. As effective agents of horizontal gene transfer, viruses have played an important role in influencing the immune system and genetic makeup of every cellular organism and have thus driven biological evolution [13]. The genetic and morphological diversity of viruses provides a large manipulable viral library that can be utilized for tailored applications in research and medicine [30].
Viruses are currently being employed in gene therapy, oncolysis, and vaccine development. Since viruses utilize the host’s cellular machinery for their replication, they have evolved to overcome or evade the host’s immune response and effectively deliver their nucleic acid to the infected cells. These features can be manipulated to allow them be used as gene delivery agents in gene therapy. Novel viral vectors that are human-friendly, non-immunogenic, and non-pathogenic and can transport large genes for stable and long-term expression are being developed to transfer therapeutic genes efficiently [41]. Self-amplifying RNA viral vectors have been shown to elicit strong humoral and cellular immunity in animal models as well [85].
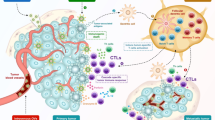
In October 2015, the FDA approved the first oncolytic virus therapy, T-VEC, for the treatment of melanoma. It is a modified herpes simplex virus that acts on cancer cells and promotes their lysis [130]. Oncolytic viruses target dendritic cells, make the tumor immunogenic, and stimulate the immune response against cancerous cells [84]. They regulate the microenvironment of cancerous cells such that T cell therapies can be used to treat solid tumors [45].
With the increasing world population and constant evolution of infectious agents, there is a dire need to develop more-efficient and novel immunization methods for which vaccines can be produced on a large scale. Viral-vector-based vaccines eliminate the need for additional adjuvants and induce a strong immune response [157].
In this review, we present an account of the major classes of viruses, both DNA viruses, including adenoviruses, adeno-associated viruses (AAVs), poxviruses, and herpesviruses, and RNA viruses, including lentiviruses, retroviruses, alphaviruses, measles virus, and Newcastle disease virus, that have shown great promise for current and future medical treatments and related applications (Table 1).
DNA viruses
Adenoviruses
Adenoviruses (Ads), are the largest non-enveloped viruses (90–100 nm) and are efficient vehicles for gene delivery. They contain a linear, non-segmented, double-stranded DNA genome of 26 to 45 kbp in their icosahedral capsid [126]. Adenoviruses can deliver transgenes of more than 8 kbp, and the viral genome resides in the nucleus episomally when injected by an adenovector, as it cannot integrate into the host genome. The genome of the adenovirus is divided into early units (E1 to E4), whose genes are responsible for the expression of non-structural proteins, and late units (L1 to L5), whose genes encode the structural components of the virion [112]. The first-generation adenovirus vector generates a strong immune response and has a capacity of around 8.2 kb for the insertion of a transgene because it lacks the E1 and E3 regions. Second-generation adenovectors are made by eliminating all of the early genes. Third-generation adenovectors, which are also known as "gutless" adenovectors, high-capacity adenoviruses (HC-Ad), or helper-dependent adenoviruses (HD-Ad), retain only the ITRs and the packaging signal (Ψ), which is essential for the final assembly of the virus [74, 83]. Adenoviral vectors have been extensively studied for their ability to carry transgenes for gene therapy. An adenovector encoding hypoxia-inducible factor 1 alpha (AdHIF-1α) has been shown to have an antiapoptotic and neuroprotective effect on ischemia and reperfusion in rats and is thought to inhibit apoptosis in nerve cells [175]. A helper-dependent HDAd5/35++ adenovector expressing CRISPR/Cas9 was generated for potential hematopoietic stem cell (HSC) gene therapy of sickle cell disease and β-thalassemia via re-activation of fetal γ-globin expression (HDAd-globin-CRISPR). This construct caused an increase in γ-globin expression in HSCs isolated from transplanted mice after genome editing [76]. An adenovirus vector Ad-E4-122aT was used in neonatal mice with hemophilia B and found to be a promising gene delivery vector for treating this disease [52].
The current emphasis in research is to utilize adenoviral vectors for development of novel vaccines. A single intranasal dose of chimpanzee adenovirus (simian Ad36)-based SARS-CoV-2 vaccine encoding the S protein (ChAd-SARS-CoV-2-S) was shown to induce the production of neutralizing antibodies and T cell responses and to limit infection in the respiratory tract after challenge with SARS-CoV-2 [44]. Another COVID-19 vaccine made using a recombinant adenovirus type 5 (Ad5) vector expressing the S protein of SARS-CoV-2 was shown to be safe and immunogenic 28 days post-vaccination in a phase 1 trial [178] A single shot of a replication-defective human Ad5 encoding the S protein of SARS-CoV-2 (Ad5-nCoV) protected mice and ferrets completely against SARS-CoV-2 infection, suggesting that mucosal vaccination may provide sufficient protection, and this can be investigated further in human clinical trials [168]. A single dose of replication-competent, highly attenuated Ad26 vector expressing mosaic HIV-1 Env (rcAd26.MOS1.HIV-Env, “rcAd26”) was found to be poorly immunogenic, suggesting that less-attenuated adenoviral vector HIV-1 vaccines should be used for oral administration [136]. The reactogenicity and immune response of a replication-defective recombinant chimpanzee Ad3-vectored Ebola virus vaccine (cAd3-EBO) was found to be dose-dependent in a phase 1 clinical trial [73]. A monovalent, recombinant, chimpanzee Ad3-vectored Zaire Ebola glycoprotein vaccine (ChAd3-EBO-Z) was found to be immunogenic and well tolerated in children aged 1 to 17 years in a phase 2 clinical trial [147]. Induction of humoral immunity by a replication-deficient human Ad5 vector expressing an empty foot-and-mouth disease virus (FMDV) capsid (AdtFMD) was studied, and the data revealed the short span of anti-FMDV- antibody-secreting cells (ASCs) and important performance characteristics of needle-free vaccination for FMDV [133]. Compared to the humoral responses induced by an inactivated influenza vaccine, the humoral responses induced by an adenovirus-vectored vaccine against the conserved stalk domain mediated cross-protection against heterosubtypic influenza viruses [63]. The efficacy of ChAdOx1, a replication-deficient simian adenovirus vaccine vector, for Middle East respiratory syndrome coronavirus (MERS-CoV), was shown to be highly immunogenic and to confer protection against lethal viral challenge [102].
Virotherapy using oncolytic adenoviruses is an effective anticancer strategy. Adenoviruses are excellent vectors in terms of manipulability and tolerance of transgenes. The potency and duration of anti-tumor activity of chimeric antigen receptor (CAR) T cells was increased using an adenovirus producing a cytokine, a checkpoint blockade, and a bispecific tumor-targeted T cell engager (BiTE) molecule, and this also ensured the dual targeting of two tumor antigens and significantly improved tumor control and survival [115]. Tumor growth was significantly inhibited by infection with a photoactivatable oncolytic adenovirus (paOAd), followed by blue light irradiation in vitro and in vivo. In addition, paOAd also showed a therapeutic effect on cancer stem cells [42]. Expression of p14 FAST from adenovirus can induce widespread syncytium formation, reduce the tumor growth rate, and improve vector efficacy for cancer treatment [28]. An oncolytic adenovirus, Delta-24-RGDOX, expressing the immune co-stimulator OX40 ligand (OX40L) showed tumor-specific activation of lymphocytes and proliferation of CD8+ T cells specific for tumor-associated antigens, resulting in immunity [57].
Adeno-associated virus
Adeno-associated virus, a member of the family Parvoviridae is a non-enveloped ssDNA virus. The small size and nonpathogenic nature of AAV make it an ideal candidate for gene transfer, with an insert capacity of ∼4.8 kilobases [129]. In primates, twelve different serotypes of AAV have been identified based on their capsid components. Most of them are being used in gene therapy [105, 129]. The use of adeno-associated viruses as vectors is not limited only to research. They are also being successfully employed as gene transfer systems for clinical purposes. To be used as a gene expression vector, the viral genome is engineered in a way that the viral DNA between the two ITRs is excised and the gene of interest is inserted in that region [129]. In one study, a novel variant of AAV called AAV-inner ear (AAV-ie) was designed and shown to be highly efficient for transduction of cochlear supporting cells and has been successfully employed for gene therapy of cochlea in a mouse model [145]. Another study showed that AAV-ANF, a variant of the AAV9 vector that contains the atrial natriuretic factor (ANF) promoter, is an efficient gene transfer vehicle for atrial-specific gene therapy [107]. Gene therapy using a recombinant AAV vector containing human rhodopsin replacement complementary DNA along with shRNA to silence mutated rhodopsin was used to treat retinitis pigmentosa in a canine model [23]. In another study, an engineered variant of AAV with better uncoating ability was shown to be effective as a carrier system for gene therapy of dendritic cells because it stimulated a stronger humoral response and induced a better adaptive response by producing anti-capsid CD8+ T cells [123]. Moreover, a study in which the AAV2/4 RPE65.RPE65 vector was used for gene therapy of Leber congenital amaurosis showed good tolerance of this vector in all nine patients tested, but the efficacy level differed among them (https://doi.org/10.1016/j.ymthe.2017.09.014). The AAV vector plays a therapeutic role in treatment of some neuromuscular disorders as well. It has been reported that a single dose of AAV9 vector containing cDNA encoding the survival motor neuron 1 (SMN1) protein provided an efficient gene replacement therapy for the treatment of spinal muscular atrophy (https://doi.org/10.1056/NEJMoa1706198). In another study, two AAV vectors were used simultaneously to deliver the CRISPR/Cas9 system for the treatment of Duchenne muscular dystrophy in a mouse model. Guide RNAs of CRISPR were inserted into scAAV, while Cas9 was inserted into ssAAV. This delivery system harnessing two AAV vectors was able to restore the expression of dystrophin and also decreased the severity of the disease (https://doi.org/10.1126/sciadv.aay6812). In hamsters that show deficiency in apolipoprotein C2, gene therapy using a recombinant AAV8 vector carrying human apolipoprotein C2 gene has been shown to significantly reduce the levels of triglycerides in the blood (https://doi.org/10.1016/j.omtm.2020.07.011).
The AAV vector is also playing a role in combating infectious diseases. Recently, it has been employed in the development of an efficient antiviral therapy against orthopoxviruses in vitro for successful and safe delivery of the CRISPR/Cas 9 system [131]. Interestingly, a single dose of vaccine that was developed against SARS-CoV-2 using engineered AAV with the AAVrh32.33 capsid was shown to induce a strong immune response in mice and non-human primates, producing a high titer of neutralizing antibodies along with the production of memory T cells [177]. AAV-vector-based gene transfer of monoclonal antibodies has been shown to provide complete protection against Ebola virus in a mouse model [155]. This vector has also shown efficacy by inducing a strong immune response including broadly neutralizing antibodies and non-neutralizing antibodies against the envelope protein of HIV-1 [116],van den [154]. Additionally, a single dose of AAV vector encoding an anti-SIV antibody has been shown to provide strong and long-lasting protection against simian immunodeficiency virus (SIV) in monkeys [93]. With regard to cancer treatment, AAV2‑shE6E7, which encodes an shRNA targeting the E6 and E7 proteins of the human papillomavirus (HPV), has demonstrated the potential to eliminate cervical cancer both in vivo and in vitro [127]. The use of AAVP particles, which are a hybrid of AAV and phage, for target-specific delivery of transgenes into tumor cells has also been reported [119, 143].
Recently, AAV-based gene therapy of X-linked myotubular myopathy has resulted in the death of two patients [149]. The safety of AAV vectors is still being debated, and further research is required to address their efficacy and safety concerns.
Herpesviruses
Herpes simplex virus is a double-stranded DNA virus with icosahedral symmetry [78]. It is being used for gene therapy because it has a large genome that comprises almost 152 kbp and has 50 genes. By deletion in the immediate early (IE) genes, the virus can accommodate almost 30 kb of exogenous DNA and transfer it to host cells [35]. Transduction of the HSV-1 vector H24B-FXNlac carrying a reduced version of the human genomic loci FXN into fetal rat dorsal root ganglia neurons resulted in long-term expression of the human FXN transgene, allowing treatment of Friedreich's ataxia neuropathy [158]. Injection of the HSV-based vector JΔN16 into the hippocampus of a mouse resulted in robust transgene expression, and injection of JΔN16 into different parts of the brain of a mouse also resulted in transgene expression and helped in the treatment of various neurological diseases, such as Parkinson’s disease [159]. Nasal application of an HSV-based vector expressing the human proenkephalin gene (SHPE) targeted the trigeminal ganglia and gave positive results in treating post-TBI (traumatic brain injury) craniofacial neuropathic pain in a rat model [96]. A recent suicide gene therapy with HSV-TK, which activates ganciclovir (GCV) by phosphorylation and targets tumor cells, gave promising results when treating mice with non-small-cell lung cancer xenografts [56].
Intramuscular administration of a single dose of HSV1716, an oncolytic virus, showed promising results in children with relapsed/refractory non-CNS solid tumors [138]. The use of rRp450, an attenuated HSV-1 vector that is used in the treatment of medulloblastoma and atypical teratoid/rhabdoid tumors (AT/RT), was associated with improved results in orthotopic xenograft pediatric brain tumor models, and 40% and 25% of mice with the BT-12 and BT-16 cell line, respectively, of AT/RT showed an increase in survival, whereas 41.6% and 27.3% with the D283med cell line and the D425med cell line, respectively, showed a decline in tumor cells [140]. The oHSV vector KNTc-gD:GDNFΔ38 infects breast cancer cells through high expression of GFRα1 in the MCF7 flank tumor model of nude mice and showed a decrease in tumor cells [43]. Three mouse models were used to study the anti-tumor effect of G47Δ, which is a third-generation oncolytic HSV. The three models with MNK-45, MNK-74, and 44As3 tumors injected intratumorally with G47Δ showed a significant decrease in the growth of the cancer cells, and various human gastric cancers were inhibited [141].
Recombinant rhCMV with antigens from SIV induces immunity against SIV with good efficacy. This rhCMV induces a wide range of T cells that are able to recognize MHC 1 and 2 epitopes and provide protection against this particular disease [7]. The HSV-d106 recombinant vector is being used as a vaccine vector against SARS-CoV-1. The S proteins expressed by d106-SARS-CoV1S localize to the surface of infected cells and promote the fusion of 293T cells expressing ACE2, the receptor for SARS. CoV-1 cells infected with d106-SARS-CoV-1S showed production of binding and neutralizing antibodies [70]. A replication-defective HSV recombinant vector has been used for vaccine development against West Nile virus. The recombinant d106-WNV expresses the prM and E proteins, leading to the formation of extracellular VLPs. This recombinant vector also induces the production of neutralizing anti-WNV IgG antibodies in immunized mice [148]. Cells of BALB/c mice vaccinated with HSV [VP6C] amplicon vector showed the presence of the VP6 protein and specific anti-RVC antibodies against group 6 rotavirus [125].
Poxviruses
Poxviruses are large enveloped DNA viruses that replicate in the cytoplasm. The oncolytic properties of these viruses make them promising candidates for treating cancer [118]. Removing the FIL gene from the poxvirus vaccinia virus greatly enhances its oncolytic activity and safety, as has been observed in a glioblastoma cell line [113]. Its ability to spread in a systemic manner in the bloodstream helps in treating metastasis, as shown in preclinical models [71]. Myxoma virus can also kill cancer cells in humans as well as in mice [118]. A myxoma virus that expresses tumor necrosis factor, called murine LIGHT, which is preloaded in mesenchymal stem cells derived from adipose tissue, is effective for treating murine pancreatic adenocarcinoma [55]. Five antiviral and three pro-viral RNA helicases have been identified that strongly affect the efficiency of myxoma virus replication in several types of human cancer cells [117]. Poxviruses are also being used as viral vectors for melanoma-based gene therapy applications in the treatment of pancreatic, prostate, and colon cancer [49]. In cats with vaccine-associated sarcomas, poxviruses have been injected postoperatively, and a lower rate of tumor reappearance was observed [87].
Poxviruses have importance as vaccines candidates, as they encode an essential element, i.e., nuclear factor kappa light chain enhancer. By inducing certain modifications, they can be optimized to be used as vaccines [139]. Synthetically modified vaccinia vectors co-expressing the spike and nucleocapsid proteins of SARS-CoV-2 induced a strong antigen-specific immune response in mice and were therefore used as a candidate for a multi-antigenic SARS-CoV-2 vaccine [20]. In laboratory experiments, oronasal or tropical exposure of bats and mice with a recombinant raccoon poxvirus vaccine protected them against rabies [135]. In phase 2 clinical trials, intramuscular injection with a recombinant attenuated poxvirus expressing cytomegalovirus antigens is being used to prevent CMV in patients who received transplants [3]. A modified vaccinia virus Ankara expressing the viral glycoprotein and the VP40 protein to form VLPs showed immunogenicity against Ebola virus [72].
Recombinant poxviruses to be used as vectors were developed by targeted insertion inactivation of the viral thymidine kinase gene and insertion of a heterologous gene in the thymidine kinase locus of the genome [62]. Modified vaccinia viruses have been shown to be immunogenic against cancer and several infectious diseases and are thus potentially effective vector systems. In clinical trials, promising results have been obtained against Ebola virus and influenza virus infections [86]. Modified vaccinia virus Ankara has been applied to the mucosal surface of the respiratory tract to provide defensive immunity in the lungs to combat SARS-CoV-2 [37]. The safety, immunogenicity, and tolerability of a modified vaccinia virus glycoprotein expressing the MERS-CoV spike were assessed for Middle East respiratory syndrome by using it as a vaccine candidate that in healthy adults [64]. In a phase 1 clinical study, a replication-deficient modified vaccinia virus Ankara and a chimpanzee adenovirus expressing conserved antigens of influenza virus were administered to patients. The results showed these modified viruses to be safe, and in response to influenza antigens, they also boosted T cell levels [24]. In a phase 2 clinical study, a multivalent respiratory syncytial virus vaccine was developed in which a poxvirus with RSV antigens and surface proteins induced a broad immune response with no adverse effects, with antibodies remaining above baseline levels for 6 months [61].
RNA viruses
Lentiviruses
Lentiviruses (LVs) are one of the three families of retroviruses. They are associated with diseases of the human immune system which can become chronic and affect the nervous system [146]. Due to their ability to tolerate large inserts and their efficient transduction ability, lentiviruses have been employed to transfer the complete dystrophin gene in Duchenne muscular dystrophy cells, resulting in the successful restoration of dystrophin production ex vivo [25]. Moreover, at the clinical stage, the lentivirus equine infectious anemia virus has been shown to be a safe and effective vector for long-term expression of angiostatin and endostatin for gene therapy of neovascular age-related macular degeneration-based vision loss [17]. Autologous hematopoietic stem cells modified by γ-globin LV and reduced-intensity conditioning transplant results showed it to be a promising method for the treatment of sickle cell anemia [89]. Clinical trials on humans who received hematopoietic stem cells as well as lentiviral-based gene therapy against chronic granulomatous disease showed that they had no new disease-related infections, and some patients were able to discontinue antibiotic prophylaxis [65]. In vitro and in vivo lentivirus-based RNA silencing of PD-L1 signaling in a pancreatic cancer cell line and in SCID-hu mice with pancreatic cancer showed improved results after dendritic cell (DC) immunization. An in vitro investigation showed a cytotoxic T-cell-based antitumor response, while in vivo, lung metastasis and tumor growth in mice were inhibited and survival rates increased [164]. In one study, an integrase-defective lentiviral vector (IDLV) was engineered to express genes for the influenza virus nucleoprotein (NP) and hemagglutinin (HA), and this was used to inoculate CB6F1 mice. A single dose of multi-antigen IDLV efficiently induced production of antiviral antibodies after 24 weeks. An H1N1 subunit vaccine was used as a positive control [39]. Receptor-targeted LVs have higher gene transfer rates than VSV LVs, and it was reported by Jamali et al. that vectofusin-1 gene delivery was considerably enhanced by CD4 and CD8 LVs [54]. A lentiviral vector-based vaccine eliciting neutralizing antibodies against the spike glycoprotein of SARS-CoV-2 was developed, and marked prophylactic effects of LV-based vaccination against SARS-CoV-2 were seen in preclinical trials. The study also showed intranasal immunization to be effective against COVID-19 [69]. In another study, an HIV vaccine using lentiviral-vector-based dendritic cells expressing CD40 ligand (CD40L), soluble programmed cell death (PD-1) dimer, and the HIV-1 SL9 epitope induced antigen-specific T cell proliferation and memory differentiation in humanized mice, and upon challenge with HIV-1, the viral load was suppressed by 2 logs for 6 weeks [108]. Phagocytosis-shielded LVs with a large amount of CD47 on the virion surface showed enhanced transduction efficacy, increased hepatocyte gene transfer, and selective targeting to spleen and liver without symptoms of toxicity [97]. Gene delivery to human T cells via CD4- and CD8-targeted lentiviral vectors was enhanced with vectofusin-1 transduction enhancer. Vectofusin-1 improved gene delivery of CD4 and CD8-LV without affecting the killing potential of CAR T cells, and the selectivity of the target cell was also maintained [53]. Using a cancer immunotherapy vaccine based on an integrase-defective lentiviral vector with IDLVs delivering ovalbumin (OVA) as a non-self antigen, tumor growth was eliminated after a single immunisation, indicating that non-self tumor-associated antigens delivered by IDLVs have a robust potential for countering the growth of various tumors [99]. A recent study also showed a 46% ??restoration of the chloride response?? after the cystic fibrosis transmembrane conductance regulator (CFTR)-knockout rats were treated with airway delivery of a lentiviral vector with the CFTR gene [120].
Other Retroviruses
Retroviruses are enveloped viruses containing two copies of the non-segmented ssRNA genome [84]. Retrovirus-based vectors have been shown to be a reliable vector system to deliver transgenes and are suitable for long-term gene therapy applications. [82]. A codon-optimized UNC-13D expression cassette delivered by a retrovirus into a T-cell culture derived from familial hemophagocytic lymphohistiocytosis-3 (FHL-3) in a patient restored expression of the functional Munc13-4 protein and lessened disease severity [29]. The A2UCOE.EFS and CBX3.EFS vectors of RV exhibited the highest levels of transgene expression in patient-specific pluripotent stem cell gene therapy and are therefore suitable for treatment for monogenetic disorders [47]. Toca511, a tumor-selective replicating retroviral vector that encodes cytokine deaminase, has been used in prodrug gene therapy in high-grade glioma patients and has shown promising results [46]. Severe combined immunodeficiency (SCID) caused by adenosine deaminase (ADA) deficiency due to a defect in the ADA gene has been treated with the help of gamma retrovirus gene therapy by transducing CD34+ cells with a gamma retrovirus encoding the human ADA cDNA sequence [36]. Retroviruses are also highly effective for killing glioma and other cancer cells [19]. Replication of the MoMLV-VSV-G vector was shown to be cytotoxic to human rhabdomyosarcoma cells and different prostate cancer cells due to syncytium formation [58]. The retroviral vector MoMLV-10A1, which encodes an R-peptide-truncated 10A1 envelope glycoprotein, has also been shown to be effective against human rhabdomyosarcoma cells [75]. Oncolytic foamy virus (oFV), which is a non-pathogenic, complex retrovirus, has shown promising results for slowing tumor growth in intraperitoneal ovarian cancer xenografts [14]. Toca511 and Toca 5C clinical trials showed effective results in patients with recurrent high-grade glioma, especially in metastatic colorectal cancer [174]. A non-replicating baculovirus expressing a human endogenous retrovirus envelope gene has been used in the development of vaccines against MERS-CoV and SARS-CoV-2 and has been shown to provide complete protection against both viruses [21]. HIV pseudotyped ncRNA VLPs have been shown to induce strong cellular and humoral immune responses [114]. Moreover, the introduction of the host proteins into retrovirus-derived VLPs induced the production of antibodies against hepatitis C virus (HCV) in mice [134]. Intramuscular immunization of mice and pigs with Ac mPERV-C5/C6 induced strong humoral and cellular responses against porcine reproductive and respiratory syndrome virus [22].
Alphaviruses
Alphaviruses, with their positive-sense single-stranded self-amplifying RNA, a broad spectrum of hosts, and significant transgene expression levels, are attractive candidates for exploitation in gene therapy and vaccine-based applications [81, 85, 137]. The high potency of alphavirus-vector-based vaccines makes them very desirable [153]. A Venezuelan equine encephalitis virus (VEEV)-based vaccine against Zika virus (ZIKV) proved effective, and viremia was not detected in mice after challenge with ZIKV [33]. In another study, the VEE TC-83 vaccine was used as construct multivalent virus-like particle vectors (VLPVs) in which the glycoprotein genes of Lassa virus (LASV) were replaced with VEEV structural genes. The VLVP vaccine stimulated dendritic and T cells and was shown to be protective [165]. Similarly, VEEV VLVPs were used to develop vaccines against two mammalian arenaviruses, Machupo virus and Junin virus, and guinea pigs were shown to be protected against these viruses [60]. A novel vaccine, V4020, which was prepared from VEEV, made mice resistant to WT-VEEV, with a high titer of antibodies. Macaques vaccinated with V4020 also showed resistance to VEEV infection after aerosol challenge [150, 151]. VEEV TC83 was used to produce RNA replicons for a SARS-CoV-2 vaccine, and a strong antibody response was observed, with antibodies being present for at least 70 days [32]. VEEV was used as a vector in a study to make ??self-amplifying RNA?? (saRNA) as a vaccine strategy for SARS-CoV-2. Animals immunized with lipid-nanoparticle-encapsulated saRNA expressing the spike protein of SARS-CoV-2 showed high antibody titers, and on re-stimulation with the viral peptides, strong cellular responses were observed [95].
In another study, Indian HIV-1C env/gag/polRT genes expressed in virus-like replicon particles based on Semliki Forest virus showed significant immunogenic potential [2]. SFV-based DNA-launched self-replicating RNA replicon (DREP) Ebola vaccines elicited a cellular and humoral immune response that was specific for Ebola virus [110]. Sindbis virus was employed to develop a vaccine against influenza A virus, and the E2S1-M2e vaccine, when given via the intranasal route, induced glycoprotein-M2e-specific antibodies, and the mice were protected from lethal challenge with the virus [67].
In 2017, the SVF-VA7 strain was observed to destroy various human prostate cancer cell lines. The infectivity and killing ability of this oncolytic alphavirus were independent of the hormone response status of the cell lines, and a nonmalignant cell line showed resistance to the virus [92]. An SFV-based DREP, developed to target cancers induced by papillomaviruses with the HPV oncogenes E6 and E7 showed anti-tumor activity, and after 108 days, 85% of the mice tested were free of tumors [153]. The M1 alphavirus showed the selective killing of muscle-invasive bladder cancer cells by inducing apoptosis without affecting normal cells [50]. SFV-AM6-124T, which is resistant to IFN-1, along with anti-PD1, increased CD8+ cell counts and infiltration of immune cells in GL261 gliomas was also enhanced [91].
The HPV16-derived antigens E6 and E7, encoded by replicon particles of Semliki Forest virus that were replication-incompetent, were used in the Vvax001 vaccine, whose phase I clinical trials took place in 2020. It was observed to be safe and well-tolerated, eliciting a response with CD4+ and CD8+ T cells against E7 and E6. The vaccine showed a positive immune response in all 12 participants in the trial [66].
Newcastle disease virus
Newcastle disease virus (NDV), also called avian avulavirus 1, is a spherical-shaped and enveloped zoonotic virus that belongs to the family Paramyxoviridae. Below the viral envelope, the nucleocapsid encloses the helical, non-segmented, negative-sense, single-stranded RNA genome of the virus [40]. NDV is an ideal vector for therapeutic use due to its ability to replicate in the host cytoplasm, thereby eliminating the risk of foreign gene incorporation into the host genome [9]. This virus has been used as a therapeutic agent in oncolysate vaccines, oncolytic and immunogenic vectors, signaling vectors, and gene delivery agents [128, 172].
Various studies have been conducted to improve or enhance the existing oncolytic properties of the virus [128]. An engineered vesicular stomatitis virus-NDV chimera (rVSV-NDV) efficiently and selectively initiated immunogenic apoptosis in hepatocellular carcinoma (HCC) [1]. Another study showed effective suppression of HCC upon exposure to a recombinant oncolytic NDV vector containing tumor necrosis factor-related apoptosis-inducing ligand (TRIAL) [169]. Expression of NDV-vectored p53 protein (rNDV-p53) in malignant glioma cells resulted in inhibition of growth, induction of apoptosis, and stimulation of a tumor-specific cytotoxic T-cell response against the glioma [34].
When NDV infects tumor cells, it attracts both innate and adaptive immune cells to the site, thus initiating immune responses against the cancer cell [16]. Various studies have used genetically engineered NDV to express immunostimulatory agents such as cytokines to combat and treat cancer [128]. An in vivo study of a recombinant NDV tumor vaccine (rNDV-TV) revealed the ability of the vaccine, mediated by natural killer cells, to induce degradation of WEHI164 fibrosarcoma [144]. In another study, macrophage inflammatory protein 3α (MIP-3α) incorporated in rNDV (rNDV-MIP-3α) significantly suppressed B16 and CT26 tumors by oncolysis and a tumor-specific immune response [51]. Immunization with an autologous recombinant NDV vaccine expressing IL-12 (rNDV-IL12) demonstrated the strong biological anti-tumor activity of IL-12 against murine melanoma in vitro and in vivo [172]. The rNDV-IL12 vaccine also showed selective cytotoxicity against human breast cancer cells in a BALB/c mouse model [98]. When tested in HT29 colon cancer cells, rNDV-IL12 induced apoptosis as well as an immune response [104]. Likewise, IL-24 incorporated into the rNDV induced apoptosis and T-cell-mediated immunity against murine melanoma [173]. In a novel approach, a combined therapeutic and preventive anti-cancer vaccine was designed to coexpress IL-7 and IL-15. Exposure of the recombinant vaccine to B16 murine melanoma degraded the tumor and stimulated the production of memory cells for prolonged protection against cancer [171].
Recombinant-NDV-vectored antibody-mediated inhibition of immunity regulators has shown great potential for treating cancer [16]. One study showed that an rNDV-vectored anti-CTLA4 single-chain variable fragment (scFv) enhanced radiotherapy-mediated myeloma oncolysis in a mouse model [161]. In another study, six different rNDV vaccines containing checkpoint inhibitor antibodies (anti-PD1 and anti-PDL1), super-antagonists (anti-CD28), and antibody-fused cytokines (anti-CD24-murine IL-12, antiPD1-mIL-12, and antiPDL1-mIL12) caused a reduction in murine myeloma, with reduced cytotoxic effects [160].
An rNDV strain expressing the immunogenic gp160 envelope protein of simian immunodeficiency virus (SIV) induced specific humoral and mucosal responses to SIV when tested in a guinea pig model [90]. Another study demonstrated the stimulation of host immunity against poliomyelitis virus by injection with rNDV expressing the poliovirus precursor capsid protein and the associated protease [162]. Similarly, induction of immunity against Ebola virus (EBOV) was also reported when an rNDV vaccine expressing EBOV glycoprotein was administered to guinea pigs [176]. Two rNDV strains expressing Japanese encephalitis virus (JEV) envelope protein and non-structural protein 1 were found to stimulate host immunity against JEV [106]. A modified NDV vector for expression of the full-length spike (S) glycoprotein of SARS-CoV-2 conferred an immune response [122]. Another group expressed wild-type and membrane-anchored S glycoprotein using an NDV vector and reported an immune response in the mice [142].
Picornavirus
The picornaviruses are a large and important family of small, non-enveloped viruses with a single-stranded, positive-sense RNA genome. The family Picornaviridae has 47 genera and 110 species [12, 167]. Their small size, lack of a genotoxic effect, lack of oncogenes, and easy manipulation as cDNA makes them suitable as vaccines and vectors in cancer gene therapy [94].
Inactivated and oral live-attenuated vaccines (IPV and OPV) against poliovirus have been used successfully worldwide, but they carry a finite risk of causing a dangerous infection. Therefore, recombinant virus-like particles (VLPs) have been developed and have been found to be a safe and sustainable alternative [6]. Formalin-inactivated coxsackievirus A5 used as a vaccine showed good efficacy against multivalent hand, foot, and mouth disease HFMD in a mouse model [59]. In another study, the A6, A10, and A16 strains of coxsackieviruses (CA6, CA10, and CA16) were also found to be effective vaccines against HFMD after inactivation with formalin or β-propiolactone (BPL), with BPL-treated vaccines yielding better results [77]. Another novel vaccine candidate was developed recently by introducing codon-deoptimized viral protein 1 (CD-VP1) together with high-fidelity 3D polymerase (3D-HF) into enterovirus A71 (EV-A71). This vaccine induced an efficient immune response against neurological as well as HFDM-causing pathogens [152].
The most commonly used immunotherapeutic and oncolytic virus of the family Picornaviridae is coxsackievirus. The use of strain CAVATAK (CVA21) together with ipilimumab has shown promising results against advanced melanoma in patients receiving anti-PD1 blockade therapy in phase I clinical trials [27]. In phase II, the combination of CVA21 and ipilimumab was given to melanoma patients without any previous therapy, and again it provided successful treatment without any serious side effects [26]. CAVATAK with a subtherapeutic dose of mitomycin C has been shown to be effective against non-muscle-invasive bladder cancer (NMIBC), and after treatment with pembrolizumab, CAVATAK showed effective oncolytic activity against advanced melanoma [4, 132]. Recently, an infectious synthetic RNA (iRNA) based on coxsackievirus A21 (R-CVA21) exhibited an efficient oncolytic effect in an animal model [31]. A novel picornavirus plasmid was designed in which the P2A site was modified to express interleukin-12 (IL-12p70) to improve its therapeutic efficacy against tumor cells [15].
A chimera of poliovirus and rhinovirus (PVSRIPO) has been developed that was found to mediate the anti-tumor effect of CD8T cells and activated dendritic cells and thereby increase the survival rate in rodent tumor models [100]. Another picornavirus, Seneca Valley virus (SVV) 001, has the ability to target tumor cells in various cancers with minimal toxicity. In phase I clinical trials, SVV-001 in an attenuated form was given intravenously to 36 patients with small-cell lung carcinoma (SCLC), neuroendocrine tumors, or carcinoid tumors, and the results were positive at a dose of 107 to 1011 virus particles per kilogram (VP/kg). In phase II, 40 patients with ED-SCLC (extensive disease) were given 1011 VP/kg, and the results were the same, i.e. antiviral antibodies were produced after 14 days without any significant symptoms [88].
Measles virus
Measles virus is an enveloped, non-segmented single-stranded negative-sense RNA virus [8]. A vaccine based on a modified measles virus strain, MeV-Stealth, has been used to treat ovarian cancer in mice by causing lysis of cancer cells expressing CD46 [101]. Measles vaccines encoding interleukin 12 and interleukin 15 agonists were tested in tumor models in vitro. A construct expressing IL12 showed robust viral gene expression and activation of immune cells, resulting in stronger anti-tumor activity [5]. For treatment of colon cancer in a rat model, MeVac FmIL-12, a measles virus encoding IL12 fusion proteins, was found to enhance anti-tumor immunity both in vitro and in vivo [163]. PEI-GOS-PEG-FA was used for coating of measles virus to form a viral GOS complex, and in mice, this resulted in increased viral replication in the tumor mass and an enhanced antitumor response [170]. An engineered measles virus Edmonston strain expressing human sodium iodide symporter or carcinoembryonic antigen was used as oncolytic viral therapy, as it provoked oncolytic effects in human hepatocellular carcinoma cell lines [10]. A recombinant measles virus, i.e., SLAM-blind measles virus, showed antitumor activity, and it is also a promising candidate for the treatment of metastatic and nectin-4-positive triple negative breast cancer, as was demonstrated in xenografted mouse model [38]. In patients with multiple myeloma, an enhanced tumor-antigen-specific T-cell response was observed after oncolytic measles virus therapy [111].
A measles-virus-based recombinant vaccine expressing the spike protein of SARS-CoV-2 was developed and shown to induce high levels of neutralizing antibodies in animals, and it was found to be protective against SARS-CoV-2 in hamsters [79]. Mice were immunized with a measles virus vaccine containing the spike glycoprotein gene of SARS-CoV-2 at two different positions in the MeV genome. It was observed that the variant with lower protein expression induced an increased Th1-biased antibody and T-cell response [48]. A measles virus encoding a soluble form of the E protein and the prM protein of Zika virus (ZIKV) conferred protection against ZIKV infection in an allogenic mouse pregnancy model [109]. A recombinant measles virus expressing the HPV capsid protein was used as a vaccine candidate for prevention of cervical cancer, and transgenic mice immunized with this virus exhibited a strong humoral immune response [18]. In a mouse model, a robust and multifunctional T-cell response was generated by immunization with a live attenuated bivalent vaccine derived from measles virus targeting Middle East respiratory syndrome coronavirus [11].
A one-cycle measles virus vector was developed by substituting the gene for the viral attachment protein with genes encoding four reprogramming factors (OCT4, SOX2, KLF4 and c-Myc), and this induced pluripotent stem cell reprogramming and was used as a platform for delivery of multiple genes [166]. A measles-virus-vectored vaccine has been used to prevent chikungunya fever in cynomolgus macaques. This vaccine was shown to induce neutralizing antibodies and to be well tolerated [124].
Conclusion
All of the aforementioned viruses have the potential to be applied in clinical settings for vaccine, gene therapy, and oncolytic strategies after appropriate safety testing. Despite their potential benefits, some of the viral vectors have drawbacks that have to be overcome. A large amount of research has been done to make adenoviral vectors more efficient for therapeutic use, although concerns such as a lack of specificity, the immunodominance of adenoviral antigens over the vaccine transgene antigen, and immune modulation by viral antigens still exist. Nevertheless, some approaches based on adenoviral vectors have been approved for gene therapy and oncotherapy. Similarly AAV, due to its very low immunogenicity and low risk of proliferation, is a promising viral vector that has been shown to be useful for long-term expression of genes. However, recent studies have raised safety concerns regarding AAV that need to be addressed. Herpes simplex virus, with its latent infection cycle, ability to be attenuated and capacity to carry large genes, has considerable potential as a vector, but it is a human pathogen, and when its genes are “turned off”, shutdown of the transgene can also occur. Clinical and pre-clinical investigations on oncolytic poxviruses have indicated that they are extremely safe and can potentially be used to treat a variety of cancers that are currently incurable. In clinical trials, vaccines against Ebola virus and influenza virus have yielded promising results. When using poxviruses as a vector, difficulty in making recombinant constructs, the transient nature of transgene expression, and possible cytotoxicity are some of the concerns that arise. Compared to other mRNAs, the immune response produced by the self-amplifying RNA of alphaviruses is more robust. saRNA vaccines based on alphaviruses have the potential to protect against diseases such as SARS-CoV-2, and are being tested in clinical trials. The main concern regarding picornavirus-based vectors, i.e., genetic instability, has been resolved by developing genetically stable expression vectors. The use of a combination of two or more picornaviral vectors such as PVSRIPO is being considered for building engineered vectors with increased stability and anti-tumor and immunogenic properties. Measles virus is a potential oncolytic platform because of its selectivity towards tumor cells and its capacity to retain its oncolytic characteristics even when it is modified. Furthermore, because it is naturally lymphotropic, it has potential for use in the development of HIV vaccines. Similarly, Newcastle disease virus is potentially useful for cancer therapy.
Although studies have suggested that the benefits of using these viruses for vaccine development, gene therapy, and oncolysis outweigh the associated risks, more research is needed to rule out potential threats before human trials can be initiated.
References
Abdullahi S, Jäkel M, Behrend SJ, Steiger K, Topping G, Krabbe T, Colombo A, Sandig V, Schiergens TS, Thasler WE, Werner J, Lichtenthaler SF, Schmid RM, Ebert O, Altomonte J (2018) A novel chimeric oncolytic virus vector for improved safety and efficacy as a platform for the treatment of hepatocellular carcinoma. J Virol 92:e01386-18
Ajbani SP, Velhal SM, Kadam RB, Patel VV, Lundstrom K, Bandivdekar AH (2017) Immunogenicity of virus-like Semliki Forest virus replicon particles expressing Indian HIV-1C gag, env and polRT genes. Immunol Lett 190:221–232
Aldoss I, La Rosa C, Baden LR, Longmate J, Ariza-Heredia EJ, Rida WN, Lingaraju CR, Zhou Q, Martinez J, Kaltcheva T, Dagis A, Hardwick N, Issa NC, Farol L, Nademanee A, Al Malki MM, Forman S, Nakamura R, Diamond DJ (2020) Poxvirus vectored cytomegalovirus vaccine to prevent cytomegalovirus viremia in transplant recipients. Ann Intern Med 172:306–316
Annels NE, Mansfield D, Arif M, Ballesteros-Merino C, Simpson GR, Denyer M, Sandhu SS, Melcher AA, Harrington KJ, Davies B (2019) Phase I trial of an ICAM-1-targeted immunotherapeutic-coxsackievirus A21 (CVA21) as an oncolytic agent against non muscle-invasive bladder cancer. Clin Cancer Res 25:5818–5831
Backhaus PS, Veinalde R, Hartmann L, Dunder JE, Jeworowski LM, Albert J, Hoyler B, Poth T, Jäger D, Ungerechts G, Engeland CE (2019) Immunological effects and viral gene expression determine the efficacy of oncolytic measles vaccines encoding IL-12 or IL-15 agonists. Viruses 11:914
Bahar MW, Porta C, Fox H, Macadam AJ, Fry EE, Stuart DI (2021) Mammalian expression of virus-like particles as a proof of principle for next generation polio vaccines. npj Vaccines 6:5
Bailer SM, Funk C, Riedl A, Ruzsics Z (2017) Herpesviral vectors and their application in oncolytic therapy, vaccination, and gene transfer. Virus Genes 53:741–748
Bellini WJ, Rota JS, Rota PA (1994) Virology of measles virus. J Infect Dis 170:S15–S23
Bello MB, Yusoff K, Ideris A, Hair-Bejo M, Jibril AH, Peeters BPH, Omar AR (2020) Exploring the prospects of engineered newcastle disease virus in modern vaccinology. Viruses 12:451
Blechacz B, Splinter PL, Greiner S, Myers R, Peng K-W, Federspiel MJ, Russell SJ, LaRusso NF (2006) Engineered measles virus as a novel oncolytic viral therapy system for hepatocellular carcinoma. Hepatology 44:1465–1477
Bodmer BS, Fiedler AH, Hanauer JRH, Prüfer S, Mühlebach MD (2018) Live-attenuated bivalent measles virus-derived vaccines targeting Middle East respiratory syndrome coronavirus induce robust and multifunctional T cell responses against both viruses in an appropriate mouse model. Virology 521:99–107
Bouin A, Semler BL (2020) Picornavirus cellular remodeling: doubling down in response to viral-induced inflammation. Current Clinical Microbiology Reports. 7:31–37
Broecker F, Moelling K (2019) What viruses tell us about evolution and immunity: beyond Darwin? Ann N Y Acad Sci 1447:53
Budzik KM, Nace RA, Ikeda Y, Russell SJ (2021) Oncolytic foamy virus—generation and properties of a nonpathogenic replicating retroviral vector system that targets chronically proliferating cancer cells. J Virol 95:e00015-21
Burkart C, Mukhopadhyay A, Shirley SA, Connolly RJ, Wright JH, Bahrami A, Campbell JS, Pierce RH, Canton DA (2018) Improving therapeutic efficacy of IL-12 intratumoral gene electrotransfer through novel plasmid design and modified parameters. Gene Ther 25:93–103
Burman B, Pesci G, Zamarin D (2020) Newcastle disease virus at the forefront of cancer immunotherapy. Cancers (Basel) 12:3552
Campochiaro PA, Lauer AK, Sohn EH, Mir TA, Naylor S, Anderton MC, Kelleher M, Harrop R, Ellis S, Mitrophanous KA (2017) Lentiviral vector gene transfer of endostatin/angiostatin for macular degeneration (GEM) study. Hum Gene Ther 28:99–111
Cantarella G, Liniger M, Zuniga A, Schiller JT, Billeter M, Naim HY, Glueck R (2009) Recombinant measles virus-HPV vaccine candidates for prevention of cervical carcinoma. Vaccine 27:3385–3390
Chen S-H, Sun J-M, Chen B-M, Lin S-C, Chang H-F, Collins S, Chang D, Wu S-F, Lu Y-C, Wang W, Chen TC, Kasahara N, Wang H-E, Tai C-K (2020) Efficient prodrug activator gene therapy by retroviral replicating vectors prolongs survival in an immune-competent intracerebral glioma model. Int J Mol Sci 21:1433
Chiuppesi F, Salazar MA, Contreras H, Nguyen VH, Martinez J, Park Y, Nguyen J, Kha M, Iniguez A, Zhou Q, Kaltcheva T, Levytskyy R, Ebelt ND, Kang TH, Wu X, Rogers TF, Manuel ER, Shostak Y, Diamond DJ, Wussow F (2020) Development of a multi-antigenic SARS-CoV-2 vaccine candidate using a synthetic poxvirus platform. Nat Commun 11:6121
Cho H, Jang Y, Park K-H, Choi H, Nowakowska A, Lee H-J, Kim M, Kang M-H, Kim J-H, Shin HY, Oh Y-K, Kim YB (2021) Human endogenous retrovirus-enveloped baculoviral DNA vaccines against MERS-CoV and SARS-CoV2. npj Vaccines 6:37
Cho Y, Heo Y, Choi H, Park KH, Kim S, Jang Y, Lee H-J, Kim M, Kim YB (2020) Porcine endogenous retrovirus envelope coated baculoviral DNA vaccine against porcine reproductive and respiratory syndrome virus. Anim Biotechnol 31:32–41
Cideciyan AV, Sudharsan R, Dufour VL, Massengill MT, Iwabe S, Swider M, Lisi B, Sumaroka A, Marinho LF, Appelbaum T, Rossmiller B, Hauswirth WW, Jacobson SG, Lewin AS, Aguirre GD, Beltran WA (2018) Mutation-independent rhodopsin gene therapy by knockdown and replacement with a single AAV vector. Proc Natl Acad Sci 115:E8547–E8556
Coughlan L, Sridhar S, Payne R, Edmans M, Milicic A, Venkatraman N, Lugonja B, Clifton L, Qi C, Folegatti PM, Lawrie AM, Roberts R, de Graaf H, Sukhtankar P, Faust SN, Lewis DJM, Lambe T, Hill AVS, Gilbert SC (2018) Heterologous Two-dose vaccination with simian adenovirus and poxvirus vectors elicits long-lasting cellular immunity to influenza virus a in healthy adults. EBioMedicine 29:146–154
Counsell JR, Asgarian Z, Meng J, Ferrer V, Vink CA, Howe SJ, Waddington SN, Thrasher AJ, Muntoni F, Morgan JE, Danos O (2017) Lentiviral vectors can be used for full-length dystrophin gene therapy. Sci Rep 7:44775
Curti B, Richards J, Hallmeyer S, Faries M, Andtbacka R, Daniels G, Grose M, Shafren DR (2017) Abstract CT114: The MITCI (Phase 1b) study: a novel immunotherapy combination of intralesional Coxsackievirus A21 and systemic ipilimumab in advanced melanoma patients with or without previous immune checkpoint therapy treatment. 77:CT114
Curti BD, Richards JM, Hallmeyer S, Faries MB, Andtbacka RHI, Daniels GA, Grose M, Shafren D (2017) Activity of a novel immunotherapy combination of intralesional Coxsackievirus A21 and systemic ipilimumab in advanced melanoma patients previously treated with anti-PD1 blockade therapy. 35:3014–3014
Del Papa J, Petryk J, Bell JC, Parks RJ (2019) An oncolytic adenovirus vector expressing p14 FAST protein induces widespread syncytium formation and reduces tumor growth rate in vivo. Mol Ther Oncolytics 14:107–120
Dettmer V, Bloom K, Gross M, Weissert K, Aichele P, Ehl S, Cathomen T (2019) Retroviral unc13d gene transfer restores cytotoxic activity of T cells derived from familial hemophagocytic lymphohistiocytosis type 3 patients in vitro. Hum Gene Ther 30:975–984
Douglas T, Young M (2006) Viruses: making friends with old foes. Science 312:873–875
Elsedawy NB, Nace RA, Russell SJ, Schulze AJ (2020) Oncolytic activity of targeted picornaviruses formulated as synthetic infectious RNA. Mol Ther Oncolytics 17:484–495
Erasmus Jesse H, Khandhar Amit P, O’Connor Megan A, Walls Alexandra C, Hemann Emily A, Murapa P, Archer J, Leventhal S, Fuller James T, Lewis Thomas B, Draves Kevin E, Randall S, Guerriero Kathryn A, Duthie Malcolm S, Carter D, Reed Steven G, Hawman David W, Feldmann H, Gale M, Veesler D, Berglund P, Fuller Deborah H (2020) An Alphavirus-derived replicon RNA vaccine induces SARS-CoV-2 neutralizing antibody and T cell responses in mice and nonhuman primates. Sci Transl Med. 12:eabc9396
Erasmus JH, Khandhar AP, Guderian J, Granger B, Archer J, Archer M, Gage E, Fuerte-Stone J, Larson E, Lin S, Kramer R, Coler RN, Fox CB, Stinchcomb DT, Reed SG, Van Hoeven N (2018) A nanostructured lipid carrier for delivery of a replicating viral RNA provides single, low-dose protection against zika. Mol Ther 26:2507–2522
Fan X, Lu H, Cui Y, Hou X, Huang C, Liu G (2018) Overexpression of p53 delivered using recombinant NDV induces apoptosis in glioma cells by regulating the apoptotic signaling pathway. Exp Ther Med 15:4522–4530
Fernández-Frías I, Pérez-Luz S, Díaz-Nido J (2020) Enhanced production of herpes simplex virus 1 amplicon vectors by gene modification and optimization of packaging cell growth medium. Mol Ther Methods Clin Dev 17:491–496
Ferrua F, Aiuti A (2017) Twenty-five years of gene therapy for ADA-SCID: from bubble babies to an approved drug. Hum Gene Ther 28:972–981
Förster R, Fleige H, Sutter G (2020) Combating COVID-19: MVA vector vaccines applied to the respiratory tract as promising approach toward protective immunity in the lung. Front Immunol 11:1959
Fujiyuki T, Amagai Y, Shoji K, Kuraishi T, Sugai A, Awano M, Sato H, Hattori S, Yoneda M, Kai C (2020) Recombinant SLAMblind measles virus is a promising candidate for nectin-4-positive triple negative breast cancer therapy. Mol Ther Oncolytics 19:127–135
Gallinaro A, Borghi M, Bona R, Grasso F, Calzoletti L, Palladino L, Cecchetti S, Vescio MF, Macchia D, Morante V, Canitano A, Temperton N, Castrucci MR, Salvatore M, Michelini Z, Cara A, Negri D (2018) Integrase defective lentiviral vector as a vaccine platform for delivering influenza antigens. Front Immunol 9:171
Ganar K, Das M, Sinha S, Kumar S (2014) Newcastle disease virus: current status and our understanding. Virus Res 184:71–81
Goswami R, Subramanian G, Silayeva L, Newkirk I, Doctor D, Chawla K, Chattopadhyay S, Chandra D, Chilukuri N, Betapudi V (2019) Gene therapy leaves a vicious cycle. Front Oncol 9:297
Hagihara Y, Sakamoto A, Tokuda T, Yamashita T, Ikemoto S, Kimura A, Haruta M, Sasagawa K, Ohta J, Takayama K (2020) Photoactivatable oncolytic adenovirus for optogenetic cancer therapy. Cell Death Dis 11:1–9
Hall BL, Leronni D, Miyagawa Y, Goins WF, Glorioso JC, Cohen JB (2020) Generation of an oncolytic herpes simplex viral vector completely retargeted to the GDNF receptor GFRα1 for specific infection of breast cancer cells. Int J Mol Sci 21:8815
Hassan AO, Feldmann F, Zhao H, Curiel DT, Okumura A, Tang-Huau T-L, Case JB, Meade-White K, Callison J, Chen RE (2021) A single intranasal dose of chimpanzee adenovirus-vectored vaccine protects against SARS-CoV-2 infection in rhesus macaques. Cell Rep Med 2:100230
Hemminki O, Dos Santos JM, Hemminki A (2020) Oncolytic viruses for cancer immunotherapy. J Hematol Oncol 13:1–15
Hiraoka K, Inagaki A, Kato Y, Huang TT, Mitchell LA, Kamijima S, Takahashi M, Matsumoto H, Hacke K, Kruse CA, Ostertag D, Robbins JM, Gruber HE, Jolly DJ, Kasahara N (2017) Retroviral replicating vector–mediated gene therapy achieves long-term control of tumor recurrence and leads to durable anticancer immunity. Neuro Oncol 19:918–929
Hoffmann D, Schott JW, Geis FK, Lange L, Müller FJ, Lenz D, Zychlinski D, Steinemann D, Morgan M, Moritz T, Schambach A (2017) Detailed comparison of retroviral vectors and promoter configurations for stable and high transgene expression in human induced pluripotent stem cells. Gene Ther 24:298–307
Hörner C, Schürmann C, Auste A, Ebenig A, Muraleedharan S, Dinnon KH, Scholz T, Herrmann M, Schnierle BS, Baric RS, Mühlebach MD (2020) A highly immunogenic and effective measles virus-based Th1-biased COVID-19 vaccine. Proc Natl Acad Sci 117:32657
Hromic-Jahjefendic A, Lundstrom K (2020) Viral vector-based melanoma gene therapy. Biomedicines 8:60
Hu C, Liu Y, Lin Y, Liang J-K, Zhong W-W, Li K, Huang W-T, Wang D-J, Yan G-M, Zhu W-B, Qiu J-G, Gao X (2018) Intravenous injections of the oncolytic virus M1 as a novel therapy for muscle-invasive bladder cancer. Cell Death Dis 9:274
Huang F-Y, Wang J-Y, Dai S-Z, Lin Y-Y, Sun Y, Zhang L, Lu Z, Cao R, Tan G-H (2020) A recombinant oncolytic Newcastle virus expressing MIP-3α promotes systemic antitumor immunity. J Immunother Cancer 8:e000330
Iizuka S, Sakurai F, Tachibana M, Ohashi K, Mizuguchi H (2017) Neonatal gene therapy for hemophilia B by a novel adenovirus vector showing reduced leaky expression of viral genes. Mol Ther Methods Clin Dev 6:183–193
Jamali A, Kapitza L, Schaser T, Johnston IC, Buchholz CJ, Hartmann J (2019) Highly efficient and selective CAR-gene transfer using CD4-and CD8-targeted lentiviral vectors. Mol Ther Methods Clin Dev 13:371–379
Jamali A, Kapitza L, Schaser T, Johnston ICD, Buchholz CJ, Hartmann J (2019) Highly efficient and selective CAR-gene transfer using CD4- and CD8-targeted lentiviral vectors. Mol Ther Methods Clin Dev 13:371–379
Jazowiecka-Rakus J, Hadrys A, Rahman MM, McFadden G, Fidyk W, Chmielik E, Pazdzior M, Grajek M, Kozik V, Sochanik A (2021) Myxoma virus expressing LIGHT (TNFSF14) pre-loaded into adipose-derived mesenchymal stem cells is effective treatment for murine pancreatic adenocarcinoma. Cancers 13:1394
Ji J, Weng Q, Zhang F, Xiong F, Jin Y, Hui J, Song J, Gao J, Chen M, Li Q (2018) Non-small-cell lung cancer: feasibility of intratumoral radiofrequency hyperthermia–enhanced herpes simplex virus thymidine kinase gene therapy. Radiology 288:612–620
Jiang H, Rivera-Molina Y, Gomez-Manzano C, Clise-Dwyer K, Bover L, Vence LM, Yuan Y, Lang FF, Toniatti C, Hossain MB (2017) Oncolytic adenovirus and tumor-targeting immune modulatory therapy improve autologous cancer vaccination. Cancer Res 77:3894–3907
Jin SY, Jung Y-T (2020) Construction of a replication-competent retroviral vector for expression of the VSV-G envelope glycoprotein for cancer gene therapy. Adv Virol 165:1089–1097
Jin W-P, Lu J, Zhang X-Y, Wu J, Wei Z-N, Mai J-Y, Qian S-S, Yu Y-T, Meng S-L, Wang Z-J, Shen S (2021) Efficacy of coxsackievirus A5 vaccine candidates in an actively immunized mouse model. J Virol 95:e01743-20
Johnson DM, Jokinen JD, Wang M, Pfeffer T, Tretyakova I, Carrion R, Griffiths A, Pushko P, Lukashevich IS (2020) Bivalent Junin & Machupo experimental vaccine based on alphavirus RNA replicon vector. Vaccine 38:2949–2959
Jordan E, Lawrence SJ, Meyer TPH, Schmidt D, Schultz S, Mueller J, Stroukova D, Koenen B, Gruenert R, Silbernagl G, Vidojkovic S, Chen LM, Weidenthaler H, Samy N, Chaplin P (2020) Broad antibody and cellular immune response from a phase 2 clinical trial with a novel multivalent poxvirus-based respiratory syncytial virus vaccine. J Infect Dis 223:1062–1072
Joshi LR, Diel DG (2021) Poxvirus vectors. In: Vanniasinkam T, Tikoo SK, Samal SK (eds) Viral vectors in veterinary vaccine development: a textbook. Springer International Publishing, Cham, pp 71–94
Kim EH, Han G-Y, Nguyen H (2017) An adenovirus-vectored influenza vaccine induces durable cross-protective hemagglutinin stalk antibody responses in mice. Viruses 9:234
Koch T, Dahlke C, Fathi A, Kupke A, Krähling V, Okba NMA, Halwe S, Rohde C, Eickmann M, Volz A, Hesterkamp T, Jambrecina A, Borregaard S, Ly ML, Zinser ME, Bartels E, Poetsch JSH, Neumann R, Fux R, Schmiedel S, Lohse AW, Haagmans BL, Sutter G, Becker S, Addo MM (2020) Safety and immunogenicity of a modified vaccinia virus Ankara vector vaccine candidate for Middle East respiratory syndrome: an open-label, phase 1 trial. Lancet Infect Dis 20:827–838
Kohn DB, Booth C, Kang EM, Pai S-Y, Shaw KL, Santilli G, Armant M, Buckland KF, Choi U, De Ravin SS, Dorsey MJ, Kuo CY, Leon-Rico D, Rivat C, Izotova N, Gilmour K, Snell K, Dip JX-B, Darwish J, Morris EC, Terrazas D, Wang LD, Bauser CA, Paprotka T, Kuhns DB, Gregg J, Raymond HE, Everett JK, Honnet G, Biasco L, Newburger PE, Bushman FD, Grez M, Gaspar HB, Williams DA, Malech HL, Galy A, Thrasher AJ, Buckland KF, Bauser CA, Honnet G, Grez M, Gaspar HB, Galy A, Thrasher AJ, C.G.D.c. the Net (2020) Lentiviral gene therapy for X-linked chronic granulomatous disease. Nat Med 26:200–206
Komdeur FL, Singh A, van de Wall S, Meulenberg JJM, Boerma A, Hoogeboom BN, Paijens ST, Oyarce C, de Bruyn M, Schuuring E, Regts J, Marra R, Werner N, Sluis J, van der Zee AGJ, Wilschut JC, Allersma DP, van Zanten CJ, Kosterink JGW, Jorritsma-Smit A, Yigit R, Nijman HW, Daemen T (2021) First-in-human phase I clinical trial of an SFV-based RNA replicon cancer vaccine against HPV-induced cancers. Mol Ther 29:611–625
Krishnavajhala HR, Williams J, Heidner H (2018) An influenza A virus vaccine based on an M2e-modified alphavirus. Adv Virol 163:483–488
Kristensen DM, Mushegian AR, Dolja VV, Koonin EV (2010) New dimensions of the virus world discovered through metagenomics. Trends Microbiol 18:11–19
Ku M-W, Bourgine M, Authié P, Lopez J, Nemirov K, Moncoq F, Noirat A, Vesin B, Nevo F, Blanc C, Souque P, Tabbal H, Simon E, Hardy D, Le Dudal M, Guinet F, Fiette L, Mouquet H, Anna F, Martin A, Escriou N, Majlessi L, Charneau P (2021) Intranasal vaccination with a lentiviral vector protects against SARS-CoV-2 in preclinical animal models. Cell Host Microbe 29:236-249.e236
Kurt-Jones EA, Dudek TE, Watanabe D, Mandell L, Che J, Zhou S, Cao L, Greenough T, Babcock GJ, Diaz F, Oh HS, Zhou C, Finberg RW, Knipe DM (2021) Expression of SARS coronavirus 1 spike protein from a herpesviral vector induces innate immune signaling and neutralizing antibody responses. Virology 559:165–172
Lawler SE, Speranza M-C, Cho C-F, Chiocca EA (2017) Oncolytic viruses in cancer treatment: a review. JAMA Oncol 3:841–849
Lázaro-Frías A, Gómez-Medina S, Sánchez-Sampedro L, Ljungberg K, Ustav M, Liljeström P, Muñoz-Fontela C, Esteban M, García-Arriaza J (2018) Distinct immunogenicity and efficacy of poxvirus-based vaccine candidates against Ebola virus expressing GP and VP40 proteins. J Virol 92:e00363-e1318
Ledgerwood JE, DeZure AD, Stanley DA, Coates EE, Novik L, Enama ME, Berkowitz NM, Hu Z, Joshi G, Ploquin A (2017) Chimpanzee adenovirus vector Ebola vaccine. N Engl J Med 376:928–938
Lee CS, Bishop ES, Zhang R, Yu X, Farina EM, Yan S, Zhao C, Zeng Z, Shu Y, Wu X (2017) Adenovirus-mediated gene delivery: potential applications for gene and cell-based therapies in the new era of personalized medicine. Genes Dis 4:43–63
Lee ES, Jin SY, Kang BK, Jung Y-T (2019) Construction of replication-competent oncolytic retroviral vectors expressing R peptide-truncated 10A1 envelope glycoprotein. J Virol Methods 268:32–36
Li C, Psatha N, Gil S, Wang H, Papayannopoulou T, Lieber A (2018) HDAd5/35++ adenovirus vector expressing anti-CRISPR peptides decreases CRISPR/Cas9 toxicity in human hematopoietic stem cells. Mol Ther Methods Clin Dev 9:390–401
Lim H, In HJ, Lee J-A, Sik Yoo J, Lee S-W, Chung GT, Choi YK, Chung JK, Cho SJ, Lee J-W (2018) The immunogenicity and protection effect of an inactivated coxsackievirus A6, A10, and A16 vaccine against hand, foot, and mouth disease. Vaccine 36:3445–3452
Liu Y-T, Jih J, Dai X, Bi G-Q, Zhou ZH (2019) Cryo-EM structures of herpes simplex virus type 1 portal vertex and packaged genome. Nature 570:257–261
Lu M, Dravid P, Zhang Y, Trivedi S, Li A, Harder O, Kc M, Chaiwatpongsakorn S, Zani A, Kenney A, Zeng C, Cai C, Ye C, Liang X, Shimamura M, Liu S-L, Mejias A, Ramilo O, Boyaka PN, Qiu J, Martinez-Sobrido L, Yount JS, Peeples ME, Kapoor A, Niewiesk S, Li J (2021) A safe and highly efficacious measles virus-based vaccine expressing SARS-CoV-2 stabilized prefusion spike. Proc Natl Acad Sci 118:e2026153118
Lundstrom K (2004) Gene therapy applications of viral vectors. Technol Cancer Res Treat 3:467–477
Lundstrom K (2005) Biology and application of alphaviruses in gene therapy. Gene Ther 12(Suppl 1):S92-97
Lundstrom K (2018) Viral vectors in gene. Therapy 6:42
Lundstrom K (2018) Viral vectors in gene therapy. Diseases 6:42
Lundstrom K (2019) RNA viruses as tools in gene therapy and vaccine development. Genes (Basel). 10:189
Lundstrom K (2020) Self-amplifying RNA viruses as RNA vaccines. Int J Mol Sci 21:5130
Lundstrom K (2021) Viral vectors for COVID-19 vaccine development. Viruses 13:317
MacNeill AL (2020) The potential of the combined use of targeted type I interferon pathway inhibitors and oncolytic viruses to treat sarcomas. Vet Comp Oncol 18:36–42
Maemondo M (2020) Perspective of gene therapy with replication competent viruses. Transl Lung Cancer Res. 9:2511–2513
Malik P, Grimley M, Quinn CT, Shova A, Courtney L, Lutzko C, Kalfa TA, Niss O, Mehta PA, Chandra S, Grassman E, Van der Loo JCM, Witting S, Nordling D, Shreshta A, Felker S, Terrell C, Reeves L, Pillis D, Anastacia L, Bushman FD, Knight-Madden J, Kalinyak K, Davies SM, Asnani M (2018) Gene therapy for sickle cell anemia using a modified gamma globin lentivirus vector and reduced intensity conditioning transplant shows promising correction of the disease phenotype. Blood 132:1021
Manoharan VK, Khattar SK, LaBranche CC, Montefiori DC, Samal SK (2018) Modified Newcastle Disease virus as an improved vaccine vector against Simian Immunodeficiency virus. Sci Rep 8:8952
Martikainen M, Ramachandran M, Lugano R, Ma J, Martikainen M-M, Dimberg A, Yu D, Merits A, Essand M (2021) IFN-I-tolerant oncolytic Semliki Forest virus in combination with anti-PD1 enhances T cell response against mouse glioma. Mol Ther Oncolytics 21:37–46
Martikainen M, Ruotsalainen J, Tuomela J, Härkönen P, Essand M, Heikkilä J, Hinkkanen A (2017) Oncolytic alphavirus SFV-VA7 efficiently eradicates subcutaneous and orthotopic human prostate tumours in mice. Br J Cancer 117:51–55
Martinez-Navio JM, Fuchs SP, Mendes DE, Rakasz EG, Gao G, Lifson JD, Desrosiers RC (2020) Long-term delivery of an anti-SIV monoclonal antibody with AAV. Front Immunol 11:449–449
McCarthy C, Jayawardena N, Burga LN, Bostina M (2019) Developing picornaviruses for cancer therapy. Cancers 11:685
McKay PF, Hu K, Blakney AK, Samnuan K, Brown JC, Penn R, Zhou J, Bouton CR, Rogers P, Polra K, Lin PJC, Barbosa C, Tam YK, Barclay WS, Shattock RJ (2020) Self-amplifying RNA SARS-CoV-2 lipid nanoparticle vaccine candidate induces high neutralizing antibody titers in mice. Nat Commun 11:3523
Meidahl A, Klukinov M, Tzabazis A, Sorensen J, Yeomans D (2017) Nasal application of HSV encoding human preproenkephalin blocks craniofacial pain in a rat model of traumatic brain injury. Gene Ther 24:482–486
Milani M, Annoni A, Moalli F, Liu T, Cesana D, Calabria A, Bartolaccini S, Biffi M, Russo F, Visigalli I (2019) Phagocytosis-shielded lentiviral vectors improve liver gene therapy in nonhuman primates. Sci Transl Med 11:eaav7325
Mohamed Amin Z, Che Ani MA, Tan SW, Yeap SK, Alitheen NB, Syed Najmuddin SUF, Kalyanasundram J, Chan SC, Veerakumarasivam A, Chia SL, Yusoff K (2019) Evaluation of a recombinant newcastle disease virus expressing human IL12 against human breast cancer. Sci Rep 9:13999
Morante V, Borghi M, Farina I, Michelini Z, Grasso F, Gallinaro A, Cecchetti S, Di Virgilio A, Canitano A, Pirillo MF, Bona R, Cara A, Negri D (2021) Integrase-defective lentiviral vector is an efficient vaccine platform for cancer immunotherapy. Viruses 13:355
Mosaheb MM, Dobrikova EY, Brown MC, Yang Y, Cable J, Okada H, Nair SK, Bigner DD, Ashley DM, Gromeier M (2020) Genetically stable poliovirus vectors activate dendritic cells and prime antitumor CD8 T cell immunity. Nat Commun 11:524
Muñoz-Alía MÁ, Nace RA, Tischer A, Zhang L, Bah ES, Auton M, Russell SJ (2021) MeV-Stealth: A CD46-specific oncolytic measles virus resistant to neutralization by measles-immune human serum. PLoS Pathog 17:e1009283
Munster VJ, Wells D, Lambe T, Wright D, Fischer RJ, Bushmaker T, Saturday G, Van Doremalen N, Gilbert SC, De Wit E (2017) Protective efficacy of a novel simian adenovirus vaccine against lethal MERS-CoV challenge in a transgenic human DPP4 mouse model. npj Vaccines 2:1–4
Naim HY (2015) Measles virus. Hum Vaccines Immunother 11:21–26
Najmuddin SUFS, Amin ZM, Tan SW, Yeap SK, Kalyanasundram J, Veerakumarasivam A, Chan SC, Chia SL, Yusoff K, Alitheen NBJP (2020) Oncolytic effects of the recombinant Newcastle disease virus, rAF-IL12, against colon cancer cells in vitro and in tumor-challenged NCr-Foxn1nu nude mice. PeerJ 8:e9761
Naso MF, Tomkowicz B, Perry WL, Strohl WR (2017) Adeno-associated virus (AAV) as a vector for gene therapy. BioDrugs 31:317–334
Nath B, Vandna HM, Saini MP, Kumar S (2020) Evaluation of Japanese encephalitis virus E and NS1 proteins immunogenicity using a recombinant Newcastle disease virus in mice. Vaccine 38:1860–1868
Ni L, Scott L, Campbell HM, Pan X, Alsina KM, Reynolds J, Philippen LE, Hulsurkar M, Lagor WR, Li N, Wehrens XHT (2019) Atrial-specific gene delivery using an adeno-associated viral vector. Circ Res 124:256–262
Norton TD, Zhen A, Tada T, Kim J, Kitchen S, Landau NR (2019) Lentiviral vector-based dendritic cell vaccine suppresses HIV replication in humanized mice. Mol Ther 27:960–973
Nürnberger C, Bodmer Bianca S, Fiedler Anna H, Gabriel G, Mühlebach Michael D, Heise Mark T (2019) A measles virus-based vaccine candidate mediates protection against zika virus in an allogeneic mouse pregnancy model. J Virol 93:e01485
Öhlund P, García-Arriaza J, Zusinaite E, Szurgot I, Männik A, Kraus A, Ustav M, Merits A, Esteban M, Liljeström P, Ljungberg K (2018) DNA-launched RNA replicon vaccines induce potent anti-Ebolavirus immune responses that can be further improved by a recombinant MVA boost. Sci Rep 8:12459
Packiriswamy N, Upreti D, Zhou Y, Khan R, Miller A, Diaz RM, Rooney CM, Dispenzieri A, Peng K-W, Russell SJ (2020) Oncolytic measles virus therapy enhances tumor antigen-specific T-cell responses in patients with multiple myeloma. Leukemia 34:3310–3322
Patel DH, Misra A (2011) 5—gene delivery using viral vectors. In: Misra A (ed) Challenges in delivery of therapeutic genomics and proteomics. Elsevier, London, pp 207–270
Pelin A, Foloppe J, Petryk J, Singaravelu R, Hussein M, Gossart F, Jennings VA, Stubbert LJ, Foster M, Storbeck C, Postigo A, Scut E, Laight B, Way M, Erbs P, Le Boeuf F, Bell JC (2019) Deletion of apoptosis inhibitor F1L in vaccinia virus increases safety and oncolysis for cancer therapy. Mol Ther Oncolytics 14:246–252
Pitoiset F, Vazquez T, Levacher B, Nehar-Belaid D, Dérian N, Vigneron J, Klatzmann D, Bellier B, Simon V (2017) Retrovirus-based virus-like particle immunogenicity and its modulation by toll-like receptor activation. J Virol 91:e01230-e11217
Porter CE, Shaw AR, Jung Y, Yip T, Castro PD, Sandulache VC, Sikora A, Gottschalk S, Ittman MM, Brenner MK (2020) Oncolytic adenovirus armed with BiTE, cytokine, and checkpoint inhibitor enables CAR T cells to control the growth of heterogeneous tumors. Mol Ther 28:1251–1262
Priddy FH, Lewis DJ, Gelderblom HC, Hassanin H, Streatfield C, LaBranche C, Hare J, Cox JH, Dally L, Bendel D (2019) Adeno-associated virus vectored immunoprophylaxis to prevent HIV in healthy adults: a phase 1 randomised controlled trial. Lancet HIV 6:e230–e239
Rahman MM, Bagdassarian E, Ali MAM, McFadden G (2017) Identification of host DEAD-box RNA helicases that regulate cellular tropism of oncolytic Myxoma virus in human cancer cells. Sci Rep 7:15710
Rahman MM, McFadden G (2020) Oncolytic virotherapy with myxoma. Virus 9:171
Reul J, Frisch J, Engeland CE, Thalheimer FB, Hartmann J, Ungerechts G, Buchholz CJ (2019) Tumor-specific delivery of immune checkpoint inhibitors by engineered AAV vectors. Front Oncol 9:52
Reyne N, Cmielewski P, McCarron A, Delhove J, Parsons D, Donnelley M (2021) Single-dose lentiviral mediated gene therapy recovers CFTR function in cystic fibrosis knockout rats. Front Pharmacol 12:1204
Robert-Guroff M (2007) Replicating and non-replicating viral vectors for vaccine development. Curr Opin Biotechnol 18:546–556
Rohaim MA, Munir M (2020) A scalable topical vectored vaccine candidate against SARS-CoV-2. Vaccines 8:472
Rossi A, Dupaty L, Aillot L, Zhang L, Gallien C, Hallek M, Odenthal M, Adriouch S, Salvetti A, Büning H (2019) Vector uncoating limits adeno-associated viral vector-mediated transduction of human dendritic cells and vector immunogenicity. Sci Rep 9:3631
Rossi SL, Comer JE, Wang E, Azar SR, Lawrence WS, Plante JA, Ramsauer K, Schrauf S, Weaver SC (2019) Immunogenicity and efficacy of a measles virus-vectored chikungunya vaccine in nonhuman primates. J Infect Dis 220:735–742
Rota RP, Palacios CA, Temprana CF, Argüelles MH, Mandile MG, Mattion N, Laimbacher AS, Fraefel C, Castello AA, Glikmann G (2018) Evaluation of the immunogenicity of a recombinant HSV-1 vector expressing human group C rotavirus VP6 protein. J Virol Methods 256:24–31
Saha B, Wong CM, Parks RJ (2014) The adenovirus genome contributes to the structural stability of the virion. Viruses 6:3563–3583
Sato N, Saga Y, Uchibori R, Tsukahara T, Urabe M, Kume A, Fujiwara H, Suzuki M, Ozawa K, Mizukami H (2018) Eradication of cervical cancer in vivo by an AAV vector that encodes shRNA targeting human papillomavirus type 16 E6/E7. Int J Oncol 52:687–696
Schirrmacher V, van Gool S, Stuecker W (2019) Breaking therapy resistance: an update on oncolytic newcastle disease virus for improvements of cancer therapy. Biomedicines 7:66
Schnödt M, Büning H (2017) Improving the quality of adeno-associated viral vector preparations: the challenge of product-related impurities. Hum Gene Ther Methods 28:101–108
Sheridan C (2015) First oncolytic virus edges towards approval in surprise vote. Nature Publishing Group
Siegrist CM, Kinahan SM, Settecerri T, Greene AC, Santarpia JL (2020) CRISPR/Cas9 as an antiviral against Orthopoxviruses using an AAV vector. Sci Rep 10:19307
Silk AW, Kaufman H, Gabrail N, Mehnert J, Bryan J, Norrell J, Medina D, Bommareddy P, Shafren D, Grose M, Zloza A (2017) Phase 1b study of intratumoral Coxsackievirus A21 (CVA21) and systemic pembrolizumab in advanced melanoma patients: Interim results of the CAPRA clinical trial. Cancer Res 77:CT026
Sitt T, Kenney M, Barrera J, Pandya M, Eckstrom K, Warner M, Pacheco JM, LaRocco M, Palarea-Albaladejo J, Brake D (2019) Duration of protection and humoral immunity induced by an adenovirus-vectored subunit vaccine for foot-and-mouth disease (FMD) in Holstein steers. Vaccine 37:6221–6231
Soares HR, Castro R, Tomás HA, Carrondo MJT, Alves PM, Coroadinha AS (2019) Pseudotyping retrovirus like particles vaccine candidates with Hepatitis C virus envelope protein E2 requires the cellular expression of CD81. AMB Express 9:22
Stading B, Ellison JA, Carson WC, Satheshkumar PS, Rocke TE, Osorio JE (2017) Protection of bats (Eptesicus fuscus) against rabies following topical or oronasal exposure to a recombinant raccoon poxvirus vaccine. PLOS Negl Trop Dis 11:e0005958
Stephenson KE, Keefer MC, Bunce CA, Frances D, Abbink P, Maxfield LF, Neubauer GH, Nkolola J, Peter L, Lane C (2018) First-in-human randomized controlled trial of an oral, replicating adenovirus 26 vector vaccine for HIV-1. PLoS ONE 13:e0205139
Strauss JH, Strauss EG (1994) The alphaviruses: gene expression, replication, and evolution. Microbiol Rev 58:491–562
Streby KA, Geller JI, Currier MA, Warren PS, Racadio JM, Towbin AJ, Vaughan MR, Triplet M, Ott-Napier K, Dishman DJ (2017) Intratumoral injection of HSV1716, an oncolytic herpes virus, is safe and shows evidence of immune response and viral replication in young cancer patients. Clin Cancer Res 23:3566–3574
Struzik J, Szulc-Dąbrowska L (2020) NF-κB as an important factor in optimizing poxvirus-based vaccines against viral infections. Pathogens 9:1001
Studebaker AW, Hutzen BJ, Pierson CR, Haworth KB, Cripe TP, Jackson EM, Leonard JR (2017) Oncolytic herpes virus rRp450 shows efficacy in orthotopic xenograft group 3/4 medulloblastomas and atypical teratoid/rhabdoid tumors. Mol Ther Oncolytics 6:22–30
Sugawara K, Iwai M, Yajima S, Tanaka M, Yanagihara K, Seto Y, Todo T (2020) Efficacy of a third-generation oncolytic herpes virus g47δ in advanced stage models of human gastric cancer. Mol Ther Oncolytics 17:205–215
Sun W, Leist SR, McCroskery S, Liu Y, Slamanig S, Oliva J, Amanat F, Schäfer A, Dinnon KH III, García-Sastre A, Krammer F, Baric RS, Palese P (2020) Newcastle disease virus (NDV) expressing the spike protein of SARS-CoV-2 as a live virus vaccine candidate. EBioMedicine 62:103132
Suwan K, Yata T, Waramit S, Przystal JM, Stoneham CA, Bentayebi K, Asavarut P, Chongchai A, Pothachareon P, Lee K-Y, Topanurak S, Smith TL, Gelovani JG, Sidman RL, Pasqualini R, Arap W, Hajitou A (2019) Next-generation of targeted AAVP vectors for systemic transgene delivery against cancer. Proc Natl Acad Sci 116:18571–18577
Takamura-Ishii M, Miura T, Nakaya T, Hagiwara K (2017) Induction of antitumor response to fibrosarcoma by Newcastle disease virus-infected tumor vaccine. Med Oncol 34:171
Tan F, Chu C, Qi J, Li W, You D, Li K, Chen X, Zhao W, Cheng C, Liu X, Qiao Y, Su B, He S, Zhong C, Li H, Chai R, Zhong G (2019) AAV-ie enables safe and efficient gene transfer to inner ear cells. Nat Commun 10:3733
Tang H, Kuhen KL, Wong-Staal F (1999) Lentivirus replication and regulation. Annu Rev Genet 33:133–170
Tapia MD, Sow SO, Mbaye KD, Thiongane A, Ndiaye BP, Ndour CT, Mboup S, Keshinro B, Kinge TN, Vernet G, Bigna JJ, Oguche S, Koram KA, Asante KP, Gobert P, Hogrefe WR, De Ryck I, Debois M, Bourguignon P, Jongert E, Ballou WR, Koutsoukos M, Roman F, Amusu S, Ayuk L, Bilong C, Boahen O, Camara M, Cheick Haidara F, Coly D, Dièye S, Dosoo D, Ekedi M, I. Eneida Almeida Dos Santos, S. Kaali, A. Kokogho, M. Levine, N. Opoku, S. Owusu-Agyei, S. Pitmang, F. Sall, M. Seydi, M. Sztein, M. Tejiokem, A. Traore, M.-A. Vernet, and A.K. Yawson. (2020) Safety, reactogenicity, and immunogenicity of a chimpanzee adenovirus vectored Ebola vaccine in children in Africa: a randomised, observer-blind, placebo-controlled, phase 2 trial. Lancet Infect Dis 20:719–730
Taylor TJ, Diaz F, Colgrove RC, Bernard KA, DeLuca NA, Whelan SP, Knipe DM (2016) Production of immunogenic West Nile virus-like particles using a herpes simplex virus 1 recombinant vector. Virology 496:186–193
Terry M (2020) Two patient deaths halt Audentes’ gene therapy trial
Tretyakova I, Plante KS, Rossi SL, Lawrence WS, Peel JE, Gudjohnsen S, Wang E, Mirchandani D, Tibbens A, Lamichhane TN, Lukashevich IS, Comer JE, Weaver SC, Pushko P (2020) Venezuelan equine encephalitis vaccine with rearranged genome resists reversion and protects non-human primates from viremia after aerosol challenge. Vaccine 38:3378–3386
Tretyakova I, Tibbens A, Jokinen JD, Johnson DM, Lukashevich IS, Pushko P (2019) Novel DNA-launched Venezuelan equine encephalitis virus vaccine with rearranged genome. Vaccine 37:3317–3325
Tsai Y-H, Huang S-W, Hsieh W-S, Cheng C-K, Chang C-F, Wang Y-F, Wang J-R (2019) Enterovirus A71 containing codon-deoptimized VP1 and high-fidelity polymerase as next-generation vaccine candidate. J Virol 93:e02308-02318
van de Wall S, Ljungberg K, Ip PP, Boerma A, Knudsen ML, Nijman HW, Liljeström P, Daemen T (2018) Potent therapeutic efficacy of an alphavirus replicon DNA vaccine expressing human papilloma virus E6 and E7 antigens. OncoImmunology. 7:e1487913
van den Berg FT, Makoah NA, Ali SA, Scott TA, Mapengo RE, Mutsvunguma LZ, Mkhize NN, Lambson BE, Kgagudi PD, Crowther C, Abdool Karim SS, Balazs AB, Weinberg MS, Ely A, Arbuthnot PB, Morris L (2019) AAV-mediated expression of broadly neutralizing and vaccine-like antibodies targeting the HIV-1 envelope V2 region. Mol Ther Methods Clin Dev 14:100–112
van Lieshout LP, Soule G, Sorensen D, Frost KL, He S, Tierney K, Safronetz D, Booth SA, Kobinger GP, Qiu X, Wootton SK (2018) Intramuscular adeno-associated virus-mediated expression of monoclonal antibodies provides 100% protection against ebola virus infection in mice. J Infect Dis 217:916–925
Vannucci L, Lai M, Chiuppesi F, Ceccherini-Nelli L, Pistello M (2013) Viral vectors: a look back and ahead on gene transfer technology. New Microbiol 36(1):1–22
Venkatraman N, Anagnostou N, Bliss C, Bowyer G, Wright D, Lövgren-Bengtsson K, Roberts R, Poulton I, Lawrie A, Ewer K (2017) Safety and immunogenicity of heterologous prime-boost immunization with viral-vectored malaria vaccines adjuvanted with Matrix-M™. Vaccine 35:6208–6217
Ventosa M, Wu Z, Lim F (2017) Sustained FXN expression in dorsal root ganglia from a nonreplicative genomic HSV-1 vector. J Gene Med 19:376–386
Verlengia G, Miyagawa Y, Ingusci S, Cohen JB, Simonato M, Glorioso JC (2017) Engineered HSV vector achieves safe long-term transgene expression in the central nervous system. Sci Rep 7:1–11
Vijayakumar G, McCroskery S, Palese P (2020) Engineering newcastle disease virus as an oncolytic vector for intratumoral delivery of immune checkpoint inhibitors and immunocytokines. J Virol 94:e01677-e11619
Vijayakumar G, Palese P, Goff PH (2019) Oncolytic Newcastle disease virus expressing a checkpoint inhibitor as a radioenhancing agent for murine melanoma. EBioMedicine 49:96–105
Viktorova EG, Khattar SK, Kouiavskaia D, Laassri M, Zagorodnyaya T, Dragunsky E, Samal S, Chumakov K, Belov GA (2018) Newcastle disease virus-based vectored vaccine against poliomyelitis. J Virol 92:e00976-e1918
Wang J, Liu T, Chen J (2021) Oncolytic measles virus encoding interleukin-12 mediated antitumor activity and immunologic control of colon cancer in vivo and ex vivo. Cancer Biother Radiopharm 36:774–782
Wang J, Sun M, Zhu X, Zhao H, Mao D, Zhang Z, Zhao X (2019) Lentivirus-mediated RNA interference targeting programmed death receptor ligand 1 increases the immunologic anti-tumor effect of dendritic cell vaccination against pancreatic cancer in SCID-hu mice. Oncol Lett 18:1539–1547
Wang M, Jokinen J, Tretyakova I, Pushko P, Lukashevich IS (2018) Alphavirus vector-based replicon particles expressing multivalent cross-protective Lassa virus glycoproteins. Vaccine 36:683–690
Wang Q, Vossen A, Ikeda Y, Devaux P (2019) Measles vector as a multigene delivery platform facilitating iPSC reprogramming. Gene Ther 26:151–164
Wang Y, Ma L, Stipkovits L, Szathmary S, Li X, Liu Y (2018) The strategy of picornavirus evading host antiviral responses: non-structural proteins suppress the production of IFNs. Front Microbiol 9:2943
Wu S, Zhong G, Zhang J, Shuai L, Zhang Z, Wen Z, Wang B, Zhao Z, Song X, Chen Y (2020) A single dose of an adenovirus-vectored vaccine provides protection against SARS-CoV-2 challenge. Nat Commun 11:1–7
Wu Y, He J, Geng J, An Y, Ye X, Yan S, Yu Q, Yin J, Zhang Z, Li D (2017) Recombinant Newcastle disease virus expressing human TRAIL as a potential candidate for hepatoma therapy. Eur J Pharmacol 802:85–92
Xia M, Luo D, Dong J, Zheng M, Meng G, Wu J, Wei J (2019) Graphene oxide arms oncolytic measles virus for improved effectiveness of cancer therapy. J Exp Clin Cancer Res 38:408
Xu X, Sun Q, Mei Y, Liu Y, Zhao L (2018) Newcastle disease virus co-expressing interleukin 7 and interleukin 15 modified tumor cells as a vaccine for cancer immunotherapy. Cancer Sci 109:279–288
Xu X, Sun Q, Yu X, Zhao L (2017) Rescue of nonlytic newcastle disease virus (NDV) expressing IL-15 for cancer immunotherapy. Virus Res 233:35–41
Xu X, Yi C, Yang X, Xu J, Sun Q, Liu Y, Zhao L (2019) Tumor cells modified with newcastle disease virus expressing IL-24 as a cancer vaccine. Mol Ther Oncolytics 14:213–221
Yagiz K, Rodriguez-Aguirre ME, Lopez Espinoza F, Montellano TT, Mendoza D, Mitchell LA, Ibanez CE, Kasahara N, Gruber HE, Jolly DJ, Robbins JM (2018) A retroviral replicating vector encoding cytosine deaminase and 5-FC induces immune memory in metastatic colorectal cancer models. Mol Ther Oncolytics 8:14–26
Yang M-L, Tao T, Xu J, Liu Z, Xu D (2017) Antiapoptotic effect of gene therapy with recombinant adenovirus vector containing hypoxia-inducible factor-1α after cerebral ischemia and reperfusion in rats. Chin Med J (Engl) 130:1700–1706
Yoshida A, Kim S-H, Manoharan VK, Varghese BP, Paldurai A, Samal SK (2019) Novel avian paramyxovirus-based vaccine vectors expressing the Ebola virus glycoprotein elicit mucosal and humoral immune responses in guinea pigs. Sci Rep 9:5520
Zabaleta N, Dai W, Bhatt U, Chichester JA, Sanmiguel J, Estelien R, Michalson KT, Diop C, Maciorowski D, Qi W, Hudspeth E, Cucalon A, Dyer CD, Pampena MB, Knox JJ, LaRocque RC, Charles RC, Li D, Kim M, Sheridan A, Storm N, Johnson RI, Feldman J, Hauser BM, Ryan A, Zinn E, Kobayashi DT, Chauhan R, McGlynn M, Ryan ET, Schmidt AG, Price B, Honko A, Griffiths A, Yaghmour S, Hodge R, Betts MR, Freeman MW, Wilson JM, Vandenberghe LH (2021) An AAV-based, room-temperature stable, single dose COVID-19 vaccine. bioRxiv:2021.2001.2005.422952
Zhu F-C, Li Y-H, Guan X-H, Hou L-H, Wang W-J, Li J-X, Wu S-P, Wang B-S, Wang Z, Wang L (2020) Safety, tolerability, and immunogenicity of a recombinant adenovirus type-5 vectored COVID-19 vaccine: a dose-escalation, open-label, non-randomised, first-in-human trial. Lancet 395:1845–1854
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Sheela Ramamoorthy.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
bin Umair, M., Akusa, F.N., Kashif, H. et al. Viruses as tools in gene therapy, vaccine development, and cancer treatment. Arch Virol 167, 1387–1404 (2022). https://doi.org/10.1007/s00705-022-05432-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-022-05432-8