
Abstract
The complete genome sequence of the virulent bacteriophage PMBT3, isolated on the proteolytic Pseudomonas grimontii strain MBTL2-21, showed no significant similarity to other known phage genome sequences, making this phage the first reported to infect a strain of P. grimontii. Electron microscopy revealed PMBT3 to be a member of the family Siphoviridae, with notably long and flexible whiskers. The linear, double-stranded genome of 87,196 bp has a mol% G+C content of 60.4 and contains 116 predicted protein-encoding genes. A putative tellurite resistance (terB) gene, originally reported to occur in the genome of a bacterium, was detected in the genome of phage PMBT3.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Pseudomonas grimontii was originally isolated from French natural mineral water in 2002 [1]. The bacterium also appears to be associated with soil and has been reported to cause turnip bacterial rot disease in Japan [2]. As an environmental microorganism capable of biofilm formation, P. grimontii may become problematic in the food industry. Currently, there are more than 300 genome sequences of Pseudomonas phages available in the National Center for Biotechnology Information (NCBI) and European Nucleotide Archive (ENA) databases, with no phages so far having been isolated specifically from P. grimontii. Moreover, numerous studies have demonstrated the successful application of Pseudomonas phages or their endolysins against plant [3,4,5,6,7] and human [8,9,10,11,12,13] pathogens. However, only a few studies have investigated phages against strains of Pseudomonas species that cause food spoilage [14,15,16]. In this study, we present an analysis of the genome sequence of the novel virulent phage PMBT3, which was isolated from sewage at a municipal wastewater treatment plant located close to Kiel in Germany. The phage was detected in a double-layer agar assay using P. grimontii strain MBTL2-21 as the host bacterium. Since large volumes of drinking water are routinely used in the dairy industry for cleaning purposes, strains of this species may be introduced post-pasteurization into dairy environments [17]. In addition, phage PMBT3 showed lysis of Pseudomonas strains that are associated with milk spoilage. Therefore, phage PMBT3 could have the potential to make dairy foods safer and improve their shelf-life. To our knowledge, this is the first report of a bacteriophage infecting P. grimontii.
Materials and methods
The proteolytic activity of P. grimontii MBTL2-21 was tested with 50 µl of overnight-grown culture spotted on skim milk agar prepared as described by Kazanas [18] and incubated at room temperature (20 °C to 22 °C) overnight. Afterwards, the strain was used in a phage screening assay, and for this, a sewage sample obtained from the local municipal wastewater treatment plant located in Bülk, close to the city of Kiel, was filtered through a 0.45-µm-pore-size membrane filter (Whatman® FP30, Schleicher & Schuell, Dassel, Germany). Five milliliters of the filtrate was combined with 0.1 ml of P. grimontii MBTL2-21 (overnight culture), 1 mM CaCl2, and 10 ml of Caso broth (Carl Roth, Karlsruhe, Germany). After incubation for 18 h, the mixture was again filtered (0.45 µm), and 0.1 ml was spotted onto a lawn consisting of 0.3 ml of P. grimontii strain MBTL2-21 overnight culture and 1 mM CaCl2 in 3 ml of molten Caso soft agar (Caso broth with 0.7% [w/w] agar) poured onto Caso agar (Carl Roth, Karlsruhe, Germany). After incubation for 18 h at room temperature, the phage-produced lysis spot was scraped off the plate, and the phage was subjected to three successive rounds of single-plaque purification as described previously [19]. For propagation of phage PMBT3, 10 plates from plaque assays with confluent lysis were selected. The plates were each washed with 3 ml of modified SM buffer (0.58% NaCl, 0.25% MgSO4 × 7H2O, 0.24% Tris-HCl [pH 7.4]) [20] for 2 h at 200 rpm. After this, the lysates were pooled and filtered through 0.45-µm-pore-size membrane filters. This lysate was further concentrated and purified using cesium chloride (CsCl) density gradient ultracentrifugation as described by Sambrook and Russell [20].
Transmission electron microscopic analysis of a high-titer lysate of the phage was performed using a Tecnai 10 transmission electron microscope (FEI Thermo Fisher, Eindhoven, The Netherlands) as described elsewhere [21].
The host range of phage PMBT3 was evaluated using different Pseudomonas target species (Online Resource 1) in a modified overlay assay performed in duplicate. Briefly, 0.3 ml of bacterial culture grown to an OD620 of 0.3 was combined with 0.1 ml of phage lysate, 0.25% (v/v) glycine, 10 mM CaCl2, and 10 mM MgCl2. After adsorption for 10 min at room temperature, 2.5 ml of Caso soft agar (0.35%) was added, and the mixture was poured onto Plate Count Agar (VWR, Darmstadt, Germany) supplemented with 0.25% glycine, 10 mM CaCl2, and 10 mM MgCl2. Finally, 10 µl of tenfold serially diluted phage lysates (prepared with phage dilution buffer consisting of ¼-strength Ringer's solution [Merck, Darmstadt, Germany] and 10% (v/v) Caso broth) was spotted onto the bacterial lawn, and the plates were incubated for 18 h, after which they were examined for phage-produced lysis zones or single plaques. Plates were usually incubated at room temperature (20 °C to 22 °C), except for those containing Pseudomonas koreensis (30 °C), Pseudomonas rhodesiae (30 °C), or Pseudomonas aeruginosa (37 °C), which were also used in the host range screening. The efficiency of plating (EOP) values for phage PMBT3 were calculated relative to the titer of this phage on the original host strain MBTL2-21 (3×109 plaque-forming units [pfu] per ml).
Phage DNA was isolated from 2 ml of high-titer phage lysate using a Phage DNA Isolation Kit (Norgen Biotek Corp., Thorold, Canada) according to the manufacturer's protocol. DNA was quantified and adjusted using a Qubit 3.0 fluorometer (Invitrogen, Germany). For DNA library preparation and genome sequencing, a Nextera XT Library Preparation Kit and a MiSeq Reagent Kit V2 were used according to the manufacturer's instructions on a MiSeq high-throughput sequencer (Illumina, Munich, Germany). The raw sequence data were evaluated and assembled de novo using Shovill 1.0.9 [22]. Open reading frames (ORFs) were predicted automatically using Rapid Annotation using Subsystem Technology (RAST) [23] and then analyzed manually for their putative functions using BLASTp [24] and SMART [25]. Their locations and predicted functions are shown in Online Resource 2. Putative host-dependent promoters (TTGACA[N15-19]TATAAT) in intergenic regions were identified using Kodon (Applied Maths, Austin, TX, USA), allowing a maximum of a 2-bp mismatch. Putative rho-independent terminators were identified using ARNold [26] at http://rssf.i2bc.paris-saclay.fr/toolbox/arnold/, with those occurring in intergenic regions and possessing a free energy in the stem-loop region of < -10 kcal mol-1 being retained. Their locations and sequences are shown in Online Resource 3. The amino acid sequences of the putative terminase large subunit (TerL) and tellurite resistance proteins (TerB) from PMBT3 were compared to the most closely related proteins using BLASTp. An amino acid sequence alignment of these protein sequences with selected related proteins encoded in the genomes of bacteria and other phages (GenBank accession numbers [GBANs] are listed in Supplementary Tables S5 and S6 in Online Resource 4) was performed in Geneious version 11.0.2 using the Geneious aligner [27]. A comparison of the phage PMBT3 genome to other phage genomes published in the databases (NCBI and ENA) was performed at the nucleotide level with megablast on the BLASTn platform [24]. Furthermore, the Virus Classification and Tree Building Online Resource (VICTOR) [28] (freely available at https://victor.dsmz.de) was used for whole-amino-acid-sequence-based phylogeny and classification. For this purpose, the phage PMBT3 proteome was compared to that of its closest relative, Pseudomonas phage Lana [29], and 30 Pseudomonas phages in the databases, with a focus on P. fluorescens phages, including the recently described P. fluorescens phage PMBT14 [30] (GBANs and their characteristics are listed in Supplementary Table S7 in Online Resource 4). All pairwise amino acid sequence comparisons were conducted using the Genome-BLAST Distance Phylogeny (GBDP) method with optimal settings as described by Meier-Kolthoff et al. [28]. More precisely, taxon boundaries were estimated with the clustering thresholds for species (i.e., 0.118980), genus (i.e., 0.749680), and family (i.e., 0.985225) [28]. Automatically generated phylogenetic trees were rooted at the midpoint [31] and visualized using FigTree [32].
Results
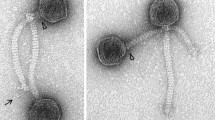
P. grimontii MBTL2-21 is capable of hydrolyzing casein on skim milk agar (proteolysis assay; data not shown). Therefore, this strain was selected as a presumable milk-spoilage strain for isolation of lytic phages. The isolated phage PMBT3 produced small (0.5 mm in diameter), turbid plaques on P. grimontii strain MBTL2-21 lawns grown on Caso soft agar at room temperature. Electron microscopy and measurement of dimensions of 22 phage particles revealed a morphotype with an isometric head of 69 ± 2.3 nm in diameter and a 268 ± 5.1-nm-long, non-contractile and flexible tail (Fig. 1). These characteristics indicate that phage PMBT3 belongs to the family Siphoviridae. A thin neck passage (nps) or collar structure (width 15.5 ± 0.9 nm) is indicated by triangles in Fig. 1a, d, and f. Notably, the phage showed up to three extraordinarily long and highly flexible whiskers (154 ± 13.3 nm) adhering at the nps structures that were found at various random positions in the vicinity of the tails. Occasionally these appendages were also detected in a bent loop formation (Fig. 1c). The distal ends of these whisker structures were also shaped with cylindrical extensions of variable length (Fig. 1a, b, c, f, h). A thin central tail fiber (length: 57 ± 8.6 nm) at the conical tail tip is indicated by asterisks in Fig. 1d and h.

Transmission electron micrographs of Siphoviridae phage PMBT3 of P. grimontii strain MBTL2-21. The arrows indicate the notably thin and highly flexible long whiskers attached to a distinct collar (neck passage) structure (see triangles). The asterisks indicate a thin tail fiber structure visible on the phage particles on the right (d and h).
In the host range assay, phage PMBT3 showed not only lytic activity against P. grimontii strain MBTL2-21 but also against several P. fluorescens strains (4 out of 7 strains tested) isolated from food samples and against P. koreensis R05-1 (1 strain tested), P. lactis strains G-8961 and G-8962 (2 out of 3 strains tested), P. protegens G-number 9102 (1 out of 3 strains tested), and P. rhodesiae B03-5 (1 strain tested) (Online Resource 1). The phage produced distinct (pinpoint) plaques on its original host strain MBTL2-21 and one tested P. fluorescens strain 17-L-08580-2-1, while it only formed cleared lysis zones on other Pseudomonas spp. strains. However, EOP values were significantly reduced for four P. fluorescens strains (10-2 to 10-6), one P. rhodesiae strain (10-4), one P. koreensis strain (10-3), two P. lactis strains (10-4), and one P. protegens strain (10-6).
Genome sequencing of phage PMBT3 produced 526,502 reads. A total of 518,833 paired-end reads were assembled de novo using Shovill 1.0.9 [22] into a single contig with a length of 87,196 bp. The DNA sequence had a mol% G+C content of 60.4. Automated annotation of the contig was performed using RAST [23], and this was followed by manual curation, resulting in 116 predicted open reading frames (ORF1-116) that contained both a start and stop codon, as well as a ribosome binding site (Online Resource 2). The ORFs were arranged in functional modules for DNA packaging, tail morphogenesis, DNA replication and transcription, and host lysis (Fig. 2). Amongst the total predicted open reading frames, only 31 ORFs (i.e., 27%) encoded proteins with a putative function, with only eight of these being assigned to phage-related proteins. The remaining 85 ORFs (i.e., 73%) were unclassified with no assigned category (i.e., encoding hypothetical proteins). The absolute numbers of start codons included 99 ATG, 11 GTG, and 6 TTG codons, respectively. Genomic analysis with PHACTS [33] revealed no lysogeny-related genes (e.g., genes encoding integrase, repressor, or antirepressor proteins), thus confirming the lytic nature of phage PMBT3. Using tRNAscan-SE version 2.0 [34], three tRNA genes for proline, glutamine, and methionine, respectively, were identified, and these were located between ORFs coding for DNA transcription and packaging proteins. Directly upstream of the thymidylate synthase gene (thyX) in the postulated replication module of PMBT3, a gene encoding a putative tellurite resistance protein (terB) was detected by BLASTp (Fig. 2) and verified using HHpred [35]. The encoded 150-amino-acid-long TerB protein showed the closest similarity to TerB proteins of bacteria, i.e., Pseudomonas sp. HMWF031 (GBAN PTU03211; coverage, 99%; identity, 60.4%) and Pseudomonas sp. B1(2018) (GBAN WP_116583222; coverage, 99%; identity, 61.7%), and less similarity to TerB of the first described member of the genus Lanavirus, i.e., the Pseudomonas phage Lana [29] (GBAN YP_009820378.1; coverage, 99%; identity, 50.3%). An amino acid sequence alignment with selected TerB proteins deduced from bacterial and phage genomes (i.e., e-values from 1e-53 to 3e-43) showed a distribution of homologous amino acids over the entire length of the protein, as demonstrated by the consensus sequence and identical amino acids in the individual sequences shown in Supplementary Fig. S1 in Online Resource 5. Structural analysis of the terminase large subunit protein using BLASTp [24] revealed a conserved domain belonging to the terminase GpA superfamily (pfam05876), which has also been detected in cos-site Escherichia coli phage lambda and Salmonella prophage Gifsy-2 [36, 37].

Genome map of PMBT3 (87,196 bp) with structural and functional annotations and GC content displayed in the inner circle. The genome starts with the ORF coding for the putative terminase large subunit (ORF1) shown at the top of the map. The putative tellurite resistance gene (terB) between the predicted replication and transcription modules is indicated by a pink arrow. The three tRNA genes in the postulated transcription and replication module are also shown. An automated annotation was obtained from RAST, and all ORFs were also annotated manually using BLASTp and SMART. The genome was subdivided into functional modules as indicated by different colors (for details, see legend). The map was generated using Geneious 11.0.2.
A search of the databases using megablast [24] indicated that the phage PMBT3 genome showed only low similarity at the nucleic acid level (i.e., coverage, 35%; identity, 81.7%) to Pseudomonas phage Lana [29] (GBAN NC_048166 [38]), which has a similar genome size of 88,342 bp. The resulting amino-acid-based phylogenetic GBDP tree generated with VICTOR [28] for the phages PMBT3, Lana, PMBT14, and 30 other Pseudomonas phages is shown in Fig. 3. The tree was reconstructed with the formula D6 and yielded an average support of 71%. It shows a branch distance of about 0.23 between the closest relative phages PMBT3 and Lana. In addition, a comparison of the predicted terminase large subunit proteins from the most closely related bacteria (or prophages) and phages in the NCBI database with that of PMBT3 showed moderate sequence similarity (i.e., 81% identity) to the corresponding protein of phage Lana, but only low similarity (i.e., a maximum of 58% identity) to the corresponding proteins of bacteria and of other phages (Supplementary Fig. S2 in Online Resource 5).

Phylogenomic analysis of phage PMBT3 with Pseudomonas phages from the NCBI database (GBANs are listed in Online Resource 4) at the amino acid level using VICTOR. The tree is based on the recommended formula D6 and shows GBDP pseudo-bootstrap support values from 100 replications above branches. The branch lengths of the resulting VICTOR tree are scaled in terms of the respective distance formula used.
Discussion
P. grimontii strain MBTL2-21 might be a proteolytic strain, as it hydrolyzed casein when grown on skim milk agar. This characteristic is of relevance to the dairy industry, as it may cause milk spoilage. Other Pseudomonas spp., such as P. fluorescens, P. gessardii, P. fragi, P. proteolytica, P. lundensis, and P. lactis are known to be major milk spoilage microorganisms at low temperatures in raw milk bulk tanks in dairies (or in the refrigerator at home), based on their ability to produce proteolytic and/or lipolytic enzymes [39, 40]. P. grimontii may be introduced into the dairy environment via the washing water used in large volumes in dairies. Therefore, (heat-treated) milk may not play a significant role in the dissemination of these strains. The use of phages, which can eliminate such “problematic” Pseudomonas spp. strains during milk storage may thus prevent spoilage or product defects that might occur during subsequent milk processing. Phage PMBT3 is a potential candidate for this purpose, as the results of the host range assay showed that, in addition to the P. grimontii strain MBTL2-21, it also lysed a strain of P. fluorescens. Furthermore, it showed lysis zones when tested on strains of P. koreensis, P. lactis, and P. rhodesiae, the last of which belongs to the P. fluorescens group [41]. The reason why no single plaques were visible could be that phage PMBT3 has not been adapted to other strains. Alternatively, it was not able to infect the strains and the lysis zones were caused by lysis from without or by other toxic effects. Even though phage titers could not be determined with the various Pseudomonas strains (except for one P. fluorescens strain) due to the lack of single plaque formation, different efficiencies of plating when compared to the host strain could be identified.
Electron microscopy showed a unique feature of this phage, as it contained extraordinary long and highly flexible whiskers with so far unknown functions. Regarding whisker-encoding ORFs, we failed to identify any ORF coding for the two long whiskers in the phage PMBT3 genome. Furthermore, we could not assign any of the six hypothetical structural ORFs (i.e., ORFs 8-13) between the predicted DNA packaging and tail morphogenesis module to the function of a capsid protein. The head of PMBT3 thus seems to be constructed from completely different capsid proteins to those described previously for other phages. However, for phage tail morphogenesis, we found six ORFs located between ORF13 and ORF20, which encoded a tail tape measure protein (ORF14), a minor tail protein (ORF15 and ORF16), a tail assembly protein (ORF17 and ORF18), and a tail fiber protein (ORF19) (Fig. 2 and Online Resource 2), respectively. It has been suggested that tRNAs are randomly gained from their hosts and then lost either neutrally or according to a set of different selection mechanisms [42]. The fact that phage PMBT3 and its closest relative phage, Lana, harbor different numbers of tRNAs could be an explanation for their different target hosts (i.e., P. grimontii for PMBT3 and Pseudomonas sp., which, however, could also include P. grimontii for Lana). The putative tellurite resistance protein TerB (ORF50)-encoding gene in the genome of PMBT3 might have originated from a bacterial genome, which is supported by the fact that ORFs for similar gene products have been found in Pseudomonas spp. genomes (Supplementary Fig. S1 in Online Resource 5). Potassium tellurite (K2TeO3) was used as antimicrobial agent before modern antibiotics were introduced [43]. Two other phages that infect Salmonella, i.e., FSL SP-076 and FSL SP-058, have previously also been described to harbor a single TerB gene [44]. Furthermore, a TerB-like gene was also found in the genome of Pseudomonas phage Lana [29] (GBAN YP_009820378.1). All of these proteins, however, showed relatively low similarity to TerB of phage PMBT3, which shows more similarity to bacterial TerB gene products (Supplementary Fig. S1 in Online Resource 5). Since the terB gene detected in PMBT3 is not accompanied by other TelR genes, which are necessary for activity [45], and since neither a promoter (i.e., -35 and -10 region) nor a rho-independent terminator could be identified upstream and downstream of the gene, respectively (Online Resource 3), this putative TerB gene product is suspected to have no tellurite-resistance-mediating activity.
The genome sequence of PMBT3 exhibited low similarity (i.e., 35% coverage and 83% identity with the megablast algorithm or 57% coverage and 78% identity with the discontiguous megablast algorithm [29]) to only one phage in the NCBI database, i.e. Pseudomonas sp. phage Lana, and clustered with this nearest neighbor in an amino-acid-based phylogenetic tree. The branching value exceeded the distance of 0.12 that has been recommended as the cutoff value for phage species [28], indicating that phage PMBT3 represents a novel taxon, separate from the Pseudomonas sp. phage Lana taxon. In fact, new taxons have recently been proposed and approved by the International Committee on Taxonomy of Viruses (ICTV) for both phages, with phage PMBT3 as the sole member of the new genus Maxrubnervirus [46] and phage Lana as the type member of the new genus Lanavirus [29, 38] in the family Siphoviridae. Finally, a comparative analysis of the predicted terminase large subunit gene also confirmed the hypothesis that phage PMBT3 represents a new species (Supplementary Fig. S2 in Online Resource 5). The catalytically active ATPase in the terminase large subunit is a housekeeping gene product that is well suited for phylogenetic comparison between phages, since it appears to be relatively conserved in dsDNA phages [36]. It is the main component of the terminase holoenzyme and is required for the highly specific process of phage DNA packaging. In conclusion, phage PMBT3 is (so far) unique regarding its genome sequence as well as its morphology. To our knowledge, this is the first reported phage infecting a P. grimontii strain.
Nucleotide sequence accession number
The complete genome sequence of phage PMBT3 generated in this project was deposited in the GenBank database (National Center for Biotechnology Information [NCBI]) under the accession number MG596799.1.
References
Baïda N, Yazourh A, Singer E et al (2002) Pseudomonas grimontii sp. nov. Int J Syst Evol Microbiol 52:1497–1503. https://doi.org/10.1099/00207713-52-5-1497
Sawada H, Horita H, Misawa T et al (2019) Pseudomonas grimontii, causal agent of turnip bacterial rot disease in Japan. J Gen Plant Pathol 85:413–423. https://doi.org/10.1007/s10327-019-00869-3
Pinheiro LAM, Pereira C, Frazão C et al (2019) Efficiency of phage φ6 for biocontrol of Pseudomonas syringae pv. syringae: an in vitro preliminary study. Microorganisms. https://doi.org/10.3390/microorganisms7090286
Frampton RA, Taylor C, Holguín Moreno AV et al (2014) Identification of bacteriophages for biocontrol of the kiwifruit canker phytopathogen Pseudomonas syringae pv. actinidiae. Appl Environ Microbiol 80:2216–2228. https://doi.org/10.1128/AEM.00062-14
Rombouts S, Volckaert A, Venneman S et al (2016) Characterization of novel bacteriophages for biocontrol of bacterial blight in leek caused by Pseudomonas syringae pv. porri. Front Microbiol 7:279. https://doi.org/10.3389/fmicb.2016.00279
Buttimer C, McAuliffe O, Ross RP et al (2017) Bacteriophages and bacterial plant diseases. Front Microbiol 8:34. https://doi.org/10.3389/fmicb.2017.00034
Tanaka H, Negishi H, Maeda H (1990) Control of tobacco bacterial wilt by an avirulent strain of Pseudomonas solanacearum M4S and its bacteriophage. Jpn J Phytopathol (Japanese Journal of Phytopathology) 56:243–246. https://doi.org/10.3186/jjphytopath.56.243
Ciepluch K, Maciejewska B, Gałczyńska K et al (2019) The influence of cationic dendrimers on antibacterial activity of phage endolysin against P. aeruginosa cells. Bioorg Chem 91:103121. https://doi.org/10.1016/j.bioorg.2019.103121
Law N, Logan C, Yung G et al (2019) Successful adjunctive use of bacteriophage therapy for treatment of multidrug-resistant Pseudomonas aeruginosa infection in a cystic fibrosis patient. Infection 47:665–668. https://doi.org/10.1007/s15010-019-01319-0
Raz A, Serrano A, Hernandez A et al (2019) Isolation of phage lysins that effectively kill Pseudomonas aeruginosa in mouse models of lung and skin infection. Antimicrob Agents Chemother. https://doi.org/10.1128/AAC.00024-19
Cafora M, Deflorian G, Forti F et al (2019) Phage therapy against Pseudomonas aeruginosa infections in a cystic fibrosis zebrafish model. Sci Rep 9:1527. https://doi.org/10.1038/s41598-018-37636-x
Waters EM, Neill DR, Kaman B et al (2017) Phage therapy is highly effective against chronic lung infections with Pseudomonas aeruginosa. Thorax 72:666–667. https://doi.org/10.1136/thoraxjnl-2016-209265
Wang H, Chan HH, Ni MY et al (2020) Bacteriophage of the skin microbiome in patients with psoriasis and healthy family controls. J Invest Dermatol 140:182-190.e5. https://doi.org/10.1016/j.jid.2019.05.023
Eller MR, Salgado RL, Vidigal PMP et al (2013) Complete genome sequence of the Pseudomonas fluorescens bacteriophage UFV-P2. Genome Announc. https://doi.org/10.1128/genomeA.00006-12
Sillankorva S, Kluskens LD, Lingohr EJ et al (2011) Complete genome sequence of the lytic Pseudomonas fluorescens phage ϕIBB-PF7A. Virol J 8:142. https://doi.org/10.1186/1743-422X-8-142
Tanaka C, Yamada K, Takeuchi H et al (2018) A lytic bacteriophage for controlling Pseudomonas lactis in raw cow’s milk. Appl Environ Microbiol. https://doi.org/10.1128/AEM.00111-18
Chiesa F, Lomonaco S, Nucera D et al (2014) Distribution of Pseudomonas species in a dairy plant affected by occasional blue discoloration. Ital J Food Saf 3:1722. https://doi.org/10.4081/ijfs.2014.1722
Kazanas N (1968) Proteolytic activity of microorganisms isolated from freshwater fish. Appl Microbiol 16:128–132
Bebeacua C, Lorenzo Fajardo JC, Blangy S et al (2013) X-ray structure of a superinfection exclusion lipoprotein from phage TP-J34 and identification of the tape measure protein as its target. Mol Microbiol 89:152–165. https://doi.org/10.1111/mmi.12267
Sambrook J, Russell DW (2001) Molecular cloning: a laboratory manual, 3rd edn. Cold Spring Harbor Laboratory Press, Cold Spring Harbor
Philippe C, Levesque S, Dion MB et al (2020) Novel genus of phages infecting Streptococcus thermophilus: genomic and morphological characterization. Appl Environ Microbiol. https://doi.org/10.1128/aem.00227-20
Seemann T. (2018) Shovill: faster SPAdes assembly of Illumina reads. https://github.com/tseemann/shovill
Aziz RK, Bartels D, Best AA et al (2008) The RAST Server: rapid annotations using subsystems technology. BMC Genomics 9:75. https://doi.org/10.1186/1471-2164-9-75
Altschul SF, Gish W, Miller W et al (1990) Basic local alignment search tool. J Mol Biol 215:403–410. https://doi.org/10.1016/S0022-2836(05)80360-2
Letunic I, Bork P (2018) 20 years of the SMART protein domain annotation resource. Nucleic Acids Res 46:D493–D496. https://doi.org/10.1093/nar/gkx922
Naville M, Ghuillot-Gaudeffroy A, Marchais A et al (2011) ARNold: a web tool for the prediction of Rho-independent transcription terminators. RNA Biol 8:11–13. https://doi.org/10.4161/rna.8.1.13346
Kearse M, Moir R, Wilson A et al (2012) Geneious Basic: an integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 28:1647–1649. https://doi.org/10.1093/bioinformatics/bts199
Meier-Kolthoff JP, Göker M (2017) VICTOR: genome-based phylogeny and classification of prokaryotic viruses. Bioinformatics 33:3396–3404. https://doi.org/10.1093/bioinformatics/btx440
Hylling O, Carstens AB, Kot W et al (2020) Two novel bacteriophage genera from a groundwater reservoir highlight subsurface environments as underexplored biotopes in bacteriophage ecology. Sci Rep 10:11879. https://doi.org/10.1038/s41598-020-68389-1
Koberg S, Gieschler S, Brinks E et al (2018) Genome sequence of the novel virulent bacteriophage PMBT14 with lytic activity against Pseudomonas fluorescens DSM 50090R. Arch Virol 163:2575–2577. https://doi.org/10.1007/s00705-018-3882-y
Farris JS (1972) Estimating phylogenetic trees from distance matrices. Am Nat 106:645–668
Rambaut A (2006) FigTree 1.4.3: A graphical viewer of phylogenetic trees and a program for producing publication-ready figures. http://tree.bio.ed.ac.uk/software/figtree/
McNair K, Bailey BA, Edwards RA (2012) PHACTS, a computational approach to classifying the lifestyle of phages. Bioinformatics 28:614–618. https://doi.org/10.1093/bioinformatics/bts014
Lowe TM, Eddy SR (1997) tRNAscan-SE: a program for improved detection of transfer RNA genes in genomic sequence. Nucleic Acids Res 25:955–964. https://doi.org/10.1093/nar/25.5.955
Söding J, Biegert A, Lupas AN (2005) The HHpred interactive server for protein homology detection and structure prediction. Nucleic Acids Res 33:W244–W248. https://doi.org/10.1093/nar/gki408
Ortega ME, Gaussier H, Catalano CE (2007) The DNA maturation domain of gpA, the DNA packaging motor protein of bacteriophage lambda, contains an ATPase site associated with endonuclease activity. J Mol Biol 373:851–865. https://doi.org/10.1016/j.jmb.2007.07.067
McClelland M, Sanderson KE, Spieth J et al (2001) Complete genome sequence of Salmonella enterica serovar Typhimurium LT2. Nature 413:852–856. https://doi.org/10.1038/35101614
Kropinski AM, Adriaenssens EM (2019) Create one new genus (Lanavirus) including one new species in the family Siphoviridae. ICTV Taxonomic Proposal 2019.062B. https://talk.ictvonline.org/taxonomy/p/taxonomy-history?taxnode_id=201907877
von Neubeck M, Baur C, Krewinkel M et al (2015) Biodiversity of refrigerated raw milk microbiota and their enzymatic spoilage potential. Int J Food Microbiol 211:57–65. https://doi.org/10.1016/j.ijfoodmicro.2015.07.001
Meng L, Zhang Y, Liu H et al (2017) Characterization of Pseudomonas spp. and associated proteolytic properties in raw milk stored at low temperatures. Front Microbiol 8:2158. https://doi.org/10.3389/fmicb.2017.02158
Anzai Y, Kim H, Park JY et al (2000) Phylogenetic affiliation of the pseudomonads based on 16S rRNA sequence. Int J Syst Evol Microbiol 50(Pt 4):1563–1589. https://doi.org/10.1099/00207713-50-4-1563
Bailly-Bechet M, Vergassola M, Rocha E (2007) Causes for the intriguing presence of tRNAs in phages. Genome Res 17:1486–1495. https://doi.org/10.1101/gr.6649807
Zadik PM, Chapman PA, Siddons CA (1993) Use of tellurite for the selection of verocytotoxigenic Escherichia coli O157. J Med Microbiol 39:155–158. https://doi.org/10.1099/00222615-39-2-155
Moreno Switt AI, Orsi RH, den Bakker HC et al (2013) Genomic characterization provides new insight into Salmonella phage diversity. BMC Genomics 14:481. https://doi.org/10.1186/1471-2164-14-481
Taylor DE (1999) Bacterial tellurite resistance. Trends Microbiol 7:111–115. https://doi.org/10.1016/S0966-842X(99)01454-7
Walker PJ, Siddell SG, Lefkowitz EJ et al (2020) Changes to virus taxonomy and the Statutes ratified by the International Committee on Taxonomy of Viruses (2020). Arch Virol 165:2737–2748. https://doi.org/10.1007/s00705-020-04752-x
Acknowledgements
We kindly thank Gesa Gehrke and Inka Lammertz for technical assistance, and Angela Back for her technical support of the electron microscopy. The authors sincerely acknowledge the help of Mareike Wenning and Christopher Huptas from the Chair of Microbial Ecology, ZIEL - Institute for Food & Health, Technische Universität München, Germany, in defining the Pseudomonas species from the genome sequence data.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
None of the authors has any conflict of interest to declare.
Ethical approval
This article does not contain any studies with human participants or animals performed by any of the authors.
Additional information
Handling Editor: Johannes Wittmann.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Sprotte, S., Brinks, E., Wagner, N. et al. Characterization of the first Pseudomonas grimontii bacteriophage, PMBT3. Arch Virol 166, 2887–2894 (2021). https://doi.org/10.1007/s00705-021-05173-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-021-05173-0