
Abstract
Porcine circovirus 3 (PCV3) is a recently identified virus that is associated with reproductive failure, porcine dermatitis and nephropathy syndrome, and multi-systemic inflammation. To investigate the molecular epidemic characteristics and genetic evolution of PCV3 in northern China, a commercial TaqMan-based real-time quantitative PCR kit was used to detect PCV3 in 435 tissue specimens collected from pigs with various clinical signs from 105 different swine farms in northern China. The results showed that 48 out of 105 (45.7%) farms and 97 out of 435 (22.3%) samples tested positive for PCV3. Of the 97 PCV3-positive samples, 80 (82.5%) tested positive for other pathogens. PCV3 was found more frequently in pigs with reproductive failure than in those with other clinical signs. This study is the first to detect PCV3 in Tianjin. The complete genome sequences of six PCV3 isolates and the capsid (Cap) protein gene sequences of 11 isolates were determined. Based on the predicted amino acids at positions 24 and 27 of the Cap protein and their evolutionary relationships, the 17 PCV3 strains obtained from northern China and 49 reference strains downloaded from the GenBank database were divided into four major groups (3a-3d). An analysis of selection pressure and polymorphism indicated that the PCV3 Cap protein seems to be evolving under balancing selection, that the population is in dynamic equilibrium, and that no population expansion occurred during the study period. Our results provide new information about the molecular epidemiology and evolution of PCV3.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Porcine circovirus 3 (PCV3) was first discovered in pigs with multi-systemic disease in the United States [1] in 2016 and has since been shown to be associated with reproductive failure and porcine dermatitis and nephropathy syndrome (PDNS) in swine [2]. To date, PCV3 has been reported in Brazil [3], South Korea [4], Thailand [5], and several European countries [6, 7]. In China, PCV3 was first detected from piglets with respiratory dyspnea and sows with reproductive failure in 2016 [8]. However, in a retrospective study, the presence of PCV3 in China could be traced back to 1996, indicating that the virus was present before the first outbreak [9]. Recently, PCV3 has been reported in sows and liveborn piglets without signs of clinical infection in China [10], indicating the potential threat of PCV3 in pig herds.
Like PCV1 and PCV2, PCV3 belongs to the family Circoviridae, genus Circovirus, and has a single-stranded circular DNA genome. The PCV3 genome contains 2,000 nucleotides and has two major open reading frames (ORFs): ORF1 and ORF2 [1, 2]. Studies on PCV2 have shown that ORF1 encodes the replication-associated protein and is relatively conserved. ORF2 encodes the major structural capsid protein (Cap), which may be associated with antigenicity and virulence. Previous studies have shown that Cap is prone to variation, and the antigenicity and virulence of ORF2 can be affected by mutation of only one or two amino acids [11,12,13]. Therefore, ORF2 is commonly used for phylogenetic and epidemiological analysis of PCV2. However, PCV3 and PCV2 have a low level nucleotide sequence identity in the Cap gene (37%).
It has been reported that the positive rates for PCV3 in pig populations range from 21.9% to 31.1% and 20% to 42.86% in southern and central China, respectively, suggesting that PCV3 is widespread in those areas [4, 14]. In the present study, we screened for PCV3 in two municipalities and four provinces of northern China and analyzed the evolutionary relationship of the epidemic strains. Our results should contribute to a better understanding of the circulation and genetic variation of PCV3 in China.
Materials and methods
Sample collection and pre-treatment
In total, 435 tissue specimens, including lymph nodes, spleens, kidneys, and lungs, were collected from stillborn fetuses, piglets, and sows with various clinical signs (diarrhea, respiratory disease, reproductive failure, etc.) in two municipalities (Beijing, Tianjin) and four provinces (Hebei, Shandong, Shanxi and Liaoning) from December 2016 to July 2018. Each sample was collected from a different pig. All samples were sent to the Animal Infectious Disease Laboratory at Hebei Agricultural University, Baoding, China, for pathogen detection. Among the 435 samples, 310 were from 60 farms in Hebei, 13 were from five farms in Beijing, 25 were from nine farms in Tianjin, 23 were from 10 farms in Shanxi, 27 were from nine farms in Shandong, and 37 were from 12 farms in Liaoning. Detailed information about these samples is provided in Table 1. The tissue samples were homogenized and stored at −20°C.
DNA extraction and virus detection
DNA from tissue samples was extracted using a MiniBEST Universal Genomic DNA Extraction Kit (TaKaRa, Dalian, China) and stored at −20°C. DNA from the 435 tissue specimens was used for PCV3 detection using a commercial TaqMan-based real-time quantitative PCR detection kit (Beijing Anheal Laboratories Co., Ltd., Beijing, China). The PCV3 genomic titer of the specimens was calculated as described previously [15]. Moreover, other pathogens, including classical swine fever virus (CSFV), porcine reproductive and respiratory syndrome virus (PRRSV), pseudorabies virus (PRV), PCV2, porcine parvovirus (PPV), porcine epidemic diarrhea virus (PEDV), and transmissible gastroenteritis virus (TGEV), were also detected using commercial fluorescence-based quantitative polymerase chain reaction (PCR) detection kits or reverse transcription PCR (RT-PCR) detection kits (Beijing Anheal Laboratories Co., Ltd.).
Sequencing and phylogenetic analysis
Three pairs of primers were designed based on a multiple alignment of currently available sequences from PCV3 isolates. Primers PCV3-F2 (5’-GGAGGGAAAGCCCGAAAC-3’) and PCV3-R2 (5’-ACCCGCCTAAACGAATGG-3’), and PCV3-F3 (5’-TCCGCATAAGGGTCGTCTT-3’) and primers PCV3-R3 (5’-AGGCATCTTCTCCGCAAC-3’) were designed to amplify the full genome sequence. Another pair of primers, ORF-F (5’-ATGAGACACAGAGCTATATTC-3’) and ORF-R (5’-TTAGAGAACGGACTTGTAACG-3’), was designed for amplification of the full-length ORF2. The primers were synthesized by Beijing Sunbiotech Co. Ltd. (Beijing, China). The PCR products were purified using a Gel Band Purification Kit (Tiangen Biotech, Beijing, China) and then cloned into the pMD-19T vector. Competent Escherichia coli DH5α was transformed with the recombinant plasmids, which were then sequenced by a commercial company (Sangon Biotech, Shanghai, China). A total of 17 PCV3 sequences, including six complete genome sequences and 11 complete Cap gene sequences were obtained and deposited in the GenBank database (Appendix 1). In addition, 40 complete genome sequences and 49 complete Cap gene sequences of PCV3 were downloaded from GenBank and used for phylogenetic analysis. Phylogenetic trees based on the complete PCV3 genome and the complete Cap gene were constructed using the maximum-likelihood method (10,000 bootstrap replicates) in the MEGA 7 program.
A similarity plot analysis of the whole genome sequences of PCV3 and various circoviruses, including unique sequences of PCV1 (NC001792), canine (NC020904), duck (NC007220), swan (NC025247), and goose (NC003054) CVs, four PCV2 isolates (AF055392, EU148503, KJ187306, AF055394), and five bat CVs (KJ641711, KJ641727, KJ641716, KJ641724, KJ641723). The analysis was done using SimPlot version 3.5.1.
The selection pressure over the Cap gene sequence was analyzed using Tajima’s D, Fu and Li’s F, and Fu and Li’s D tests of neutrality, which are available in DnaSP version 5.10 [16]. These methods evaluate whether the number of segregating sites in a sequence alignment departs significantly from the neutral expectation. Selective pressures acting on each codon of the Cap gene nucleotide sequences were analyzed using the fixed-effects likelihood (FEL), mixed effects model evolution (MEME), and single-likelihood ancestor counting (SLAC) methods [17], which are available on the Datamonkey web server (http://www.datamonkey.org/) [18].
Results
Detection of PCV3 in northern China
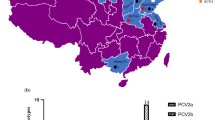
In this study, 435 tissue samples collected from two municipalities and four provinces of northern China were tested. The results showed that 97 out of 435 samples (22.3%) and 48 out of 105 farms (45.7%) tested positive for PCV3 (Table 1). For the samples collected from December 2016 to December 2017 in Beijing, Hebei, and Tianjin, the PCV3 farm positive rate was 37.5% and the sample positive rate was 14.5%. For the samples collected from January 2018 to July 2018 in these districts, the PCV3 farm and samples positive rates were 65.3% and 40.6%, respectively. This increase in the positive detection rate of PCV3 may indicate an increasing threat to pig herds.
Differences in positive detection rates were also observed between samples from different regions of northern China (Tables 1 and 2). Samples collected from Tianjin showed a higher PCV3-positive rate (44.0%, 11/25) than those from Hebei (16.5%, 51/310), Beijing (30.8%, 4/13), Shandong (40.7%, 11/27), Shanxi (34.8%, 8/23), and Liaoning (32.4%, 12/37). PCV3 was detected at a higher rate (37.1%; 23/62) in pigs with reproductive failure than in those with other clinical signs (Table 2), and the mean viral load of PCV3 was higher in the group with reproductive failure (4.68 ± 0.11) than in the groups with diarrhea (3.88 ± 0.07) or respiratory disease (3.48 ± 0.11) (Table 2).
Further analysis showed that coinfection of PCV3 with other pathogens was common.
The rates of dual infection with PCV3 and PCV2, CSFV, PRV, PEDV, PPV, PRRSV and TGEV, were 14.43% (14/97), 11.34% (11/97), 7.22% (7/97), 7.22% (7/97), 5.15% (5/97), 2.06% (2/97) and 2.06% (2/97), respectively. The highest rate of triple infection was with PCV3, PCV2 and PPV 10.31% (10/97). The rate of single infection with PCV3 in the samples was 17.5% (17/97) (Table 3). These results provide a further indication of the potential threat of PCV3 to the swine industry.
Phylogenetic analysis
Sequence analysis showed that the complete genomes of the six PCV3 strains detected in this study shared 98.6%-99.4% nucleotide sequence identity with each other and 98.8%-100% nucleotide sequence identity with those of the other 40 reference strains from various countries. The alignment of Cap gene sequences showed that the 17 complete PCV3 Cap genes shared 97.2% to 99.7% nucleotide sequence identity with each other and 97.2%-99.8% nucleotide sequence identity with those of the other 49 reference strains. No deletions or insertions were found in the Cap gene region.
A phylogenetic tree based on the complete genome sequence of PCV3 showed that the PCV3 isolates were divided into two clusters, designated as 3a and 3b. Five strains detected in this study (PCV3/CH/HB/CZ-2/2017 MG727537, PCV3/CH/HB/BD/2017 MG727538, PCV3/CH/HB/CZ-1/2017 MG727539, PCV3/CH/HB/HD/2017 MH101645, and PCV3/CH/Tianjin-1/2018 MH547276) belong to the 3a cluster, and one strain (PCV3/CH/HB/SJZ/2017 MG727540) was in the 3b cluster. The strains PCV3/CH/HB/BD/2017 MG727538, PCV3/CH/HB/HD/2017 MH101645, and PCV3/CH/HB/CZ-1/2017 MG727539 were found to be closely related to strain 2164/USA KX458325 (Fig. 2a).
A separate phylogenetic analysis was carried out based on a multiple sequence alignment of the deduced amino acid sequences of the 66 PCV3 Cap protein genes mentioned above. These strains were divided into four different clades, groups 3a-3d, based on the amino acids present at positions 24 and 27 of the Cap protein, as described previously [9, 19]. Group 3a consisted of strains with alanine and arginine at positions 24 and 27, respectively, group 3b consisted of strains with alanine and lysine at these positions, and group 3c consisted of strains with valine and lysine at these positions. More importantly, a new group, 3d, was identified in our study, with valine at position 24 and arginine at position 27. Of the 17 PCV3 strains identified in the present study, eight were in group 3a, seven were in group 3c, two were in group 3d, and none were in group 3b (Figs. 1 and 2b).

Molecular features of the PCV3 Cap protein. An alignment of the deduced amino acid sequences of PCV3 Cap proteins was performed using Megalign software (DNASTAR). The major amino acid mutations are displayed in a box, and based on two amino acid mutations at positions 24 and 27 of the Cap protein (A24V and R27K), each amino acid mutation is labeled in blue, yellow, red or green. Red boxes indicate the strains identified in this study

(a) Phylogenetic tree based on complete PCV3 genome sequences. (b) Phylogenetic tree of PCV3 strains based on the deduced amino acid sequences of PCV3 Cap proteins. All of the reference sequences used in this study were obtained from the GenBank database. The tree was constructed by the maximum-likelihood method with 10,000 bootstrap replicates using MEGA 7.0 software. PCV3 strains are identified by virus name, country of origin, isolation year, and GenBank accession no. Black solid circles indicate the strains identified in this study
Group 3a (24A and 27R) was further divided into three subgroups (3a1, 3a2 and 3a3) on the basis of the molecular features and variations in the Cap protein. The strains in subgroup 3a1 contained the S77T and I150L mutations, except for the PCV3/CH/HB/CZ-2/2017 MG727537 strain, which had only the I150L mutation in the Cap protein. Strains with the N56D mutation clustered in subgroup 3a2; however, the PCV3/CH/Liaoning-2/2018 MH548438 strain had both N56D and F104Y mutations. Subgroup 3a3 consisted of the seven strains with the F104Y mutation. These differences might be useful for understanding the evolutionary history of these strains. Notably, the formation of this subgroup was more pronounced when the amino acid sequence of the Cap protein was used for analysis (Fig. 1).
Recombination analysis
Similarity plots comparing the whole genome sequence of PCV3 with those of other circoviruses were made using SimPlot software. The results showed that only bat circoviruses and PCV2 showed nucleotide sequence similarity to PCV3. There were three main peaks of sequence similarity between PCV3 and bat circoviruses, but only one between PCV3 and PCV2 (Fig. 3). Recombination analysis was also performed using the RDP software, but no recombination signal was found.

Similarity plot of the whole genome sequences of 54 different circoviruses, including unique sequences of PCV1, canine, duck, swan, and goose CVs, as well as 40 PCV3 isolates, four PCV2 isolates, and five bat CV isolates. The similarity plot was constructed using the Kimura two-parameter distance model with a sliding window of 200 bp and a step size of 20 bp. The vertical and horizontal axes indicate the similarity (%) and the position (bp) of the alignment, respectively
Neutrality tests and selection pressure analysis
Neutrality tests yielded a negative value for Tajima’s D and a positive value for Fu and Li’s F, but neither of these values was statistically significant. However, the value of Fu and Li’s D was positive and statistically significant (Table 4). The positive value for Fu and Li’s D indicated that the Cap gene of PCV3 should be under balancing selection.
The Ka/Ks ratio is a measure of the selection pressure at the protein level. For the PCV3 Cap gene, the value of Ka/Ks was 0.1559. No positive or negative selection sites were detected by the MEME and FEL methods. Only the SLAC method detected six codons (2.80%) that appear to be under negative selection pressure (Table 5). In summary, the Cap protein of the PCV3 isolate from this study has been subjected to balancing selection.
Further mismatch distribution analysis showed that the mismatch curve for individual sequences contained multiple peaks, showed a multipeak curve, indicating that the population is in dynamic equilibrium and that no population expansion has occurred recently (Fig. 4).

Mismatch distribution of Cap gene sequences
Discussion
The novel virus PCV3 has been identified and reported worldwide, and has been found in more than two-thirds of the provinces in China [8, 20]. In the present study, a TaqMan-based real-time quantitative PCR was used for detection of PCV3, and 435 tissue samples collected from two municipalities (Beijing, Tianjin) and four provinces (Hebei, Shandong, Shanxi and Liaoning) in northern China between December 2016 and July 2018 were analyzed. The results showed that 97 (22.3%) samples and 48 (45.7%) farms were PCV3 positive, demonstrating the wide spread of the virus in this area. The rate of PCV3 detection was lower in Hebei province than in Beijing, Tianjin, Shandong, Liaoning, and Shanxi. One reason for these differences may be that the rates were calculated without using epidemiological tools. Another probable reason is that Hebei is a pig-exporting province and therefore less affected by infections. The PCV3 positive rate in 2018 was higher than that from December 2016 to December 2017, indicating that the infection rate in northern China has been gradually increasing. Notably, this is the first time that PCV3 has been detected in Tianjin, suggesting that the virus has been circulating in swine herds in this district. More specimens need to be tested to evaluate the epidemic state of PCV3 in these regions.
PCV3 has been reported to be associated with reproductive failure, PNDS, diarrhoea, respiratory disease, and congenital tremors [1,2,3, 5, 21]. In this study, we examined the association of various clinical signs with PCV3 detection, as was also done in 2017 [19]. Positivity for PCV3 was more frequently associated with reproductive failure (23/67, 34.3%) than with any of the other clinical signs investigated, and the mean viral load in this group was higher than in the other groups, suggesting that PCV3 might be an important factor in reproductive failure and a significant threat to pig production. Further research on the pathogenesis of PCV3 and coinfection with other viruses should be performed.
Of the 97 PCV3-positive samples, only 17.5% were infected by PCV3 alone, and 82.5% were positive for other viruses. In the cases of coinfection, the most commonly detected coinfecting virus was PCV2 (31/97). PCV2 has been reported to be the primary causative agent of PCV-associated disease, and it causes immunosuppression in infected pigs [22]. The 17.5% single infection rate for PCV3 suggests that PCV3 might also play a similar role in infected pigs. Further research on the pathogenesis of PCV3 and coinfection with other viruses should be performed.
The complete genome sequences of the six PCV3 isolates from this study were 98.6%-99.4% identical, and the sequences of 645-nt fragments from 17 PCV3 isolates from this study were 97.2%-99.7% identical, indicating that the PCV3 strains in northern China remained genetically stable during the 2-year period of the study. Li et al. used maximum-likelihood (ML), maximum-clade-credibility (MCC), and neighbor-joining (NJ) methods to construct phylogenetic trees for PCV3 using the full genome sequence, which revealed the existence of two clades [23]. In this study, PCV3 strains were also divided into two different clades using the ML method, PCV3a and PCV3b, based on complete genome sequences. SimPlot and RDP analysis indicated that there has been no recombination between PCV3 and other known circoviruses. Neutrality tests and selection pressure analysis showed that the Cap protein gene of PCV3 is likely to be subjected to balancing selection. Selection pressure analysis on the ORF2 sequences of PCV3 in Brazil was performed by Saraiva et al. [24]. In that study, 17% of the codons were predicted to be evolving under purifying selection due to negative selection. In this study, six codons (2.80%) were found to be under negative selection. These results show that the Cap protein sequence of PCV3 is highly conserved and does not seem to permit high levels of variation.
Mutations at positions 24 and 27 in the Cap protein have been speculated to be associated with immune escape, and on the basis of two amino acid mutations (A24V and R27K) in the Cap protein, PCV3 strains have been divided into three groups (3a-3c), as described previously [9, 19]. In current study, based on A24V and R27K mutations, the 66 PCV3 strains were clustered into four groups (3a-3d) (Figs. 1 and 2b). A new group with the mutation 24VRRR27 in Cap gene, named 3d, was found in our study, and the strains PCV3/CH/Tianjin-2/2017 MH579786 and PCV3/CH/HB-TS/2018 MH636590 were found to belong to this group. A recent report on the evolutionary history of PCV3 has suggested that amino acids 24 and 27 of the Cap protein in PCV3 have mutated from 24ARRR27 to 24ARRK27, and then to 24VRRK27 [9]. Although the number of strains with the 24VRRR27 mutation was limited, the discovery might contribute to our understanding of the evolution of this virus and should be investigated further.
In conclusion, our research provides evidence of an epidemic of PCV3 in part of northern China and analysis of the genetic evolution of PCV3 strains over a 2-year period. The results could contribute to future research on the molecular epidemiology and evolution of PCV3 in China. Further studies should be performed to determine the pathogenic characteristics, epidemiological status, and evolutionary relationships of PCV3 and its association with various clinical signs in pigs.
References
Phan TG, Giannitti F, Rossow S, Marthaler D, Knutson TP, Li L, Delwart E (2016) Detection of a novel circovirus PCV3 in pigs with cardiac and multi-systemic inflammation. Virol J 13:184
Palinski R, Pineyro P, Shang P, Yuan F, Guo R, Fang Y, Hause BM (2017) A novel porcine circovirus distantly related to known circoviruses is associated with porcine dermatitis and nephropathy syndrome and reproductive failure. J Virol 91(1):e01879-16
Tochetto C, Lima DA, Varela APM, Loiko MR, Paim WP, Scheffer CM, Roehe PM (2018) Full-Genome Sequence of Porcine Circovirus type 3 recovered from serum of sows with stillbirths in Brazil. Transbound Emerg Dis 65:5–9
Kwon T, Yoo SJ, Park CK, Lyoo YS (2017) Prevalence of novel porcine circovirus 3 in Korean pig populations. Vet Microbiol 207:178–180
Kedkovid R, Woonwong Y, Arunorat J, Sirisereewan C, Sangpratum N, Lumyai M, Thanawongnuwech R (2018) Porcine circovirus type 3 (PCV3) infection in grower pigs from a Thai farm suffering from porcine respiratory disease complex (PRDC). Vet Microbiol 215:71–76
Faccini S, Barbieri I, Gilioli A, Sala G, Gibelli LR, Moreno A, Nigrelli A (2017) Detection and genetic characterization of Porcine circovirus type 3 in Italy. Transbound Emerg Dis 64:1661–1664
Stadejek T, Wozniak A, Milek D, Biernacka K (2017) First detection of porcine circovirus type 3 on commercial pig farms in Poland. Transbound Emerg Dis 64:1350–1353
Ku X, Chen F, Li P, Wang Y, Yu X, Fan S, He Q (2017) Identification and genetic characterization of porcine circovirus type 3 in China. Transbound Emerg Dis 64:703–708
Sun J, Wei L, Lu Z, Mi S, Bao F, Guo H, Gong W (2018) Retrospective study of porcine circovirus 3 infection in China. Transbound Emerg Dis 65:607–613
Zheng S, Wu X, Zhang L, Xin C, Liu Y, Shi J, Wang J (2017) The occurrence of porcine circovirus 3 without clinical infection signs in Shandong Province. Transbound Emerg Dis 64:1337–1341
Ge M, Yan A, Luo W, Hu YF, Li RC, Jiang DL, Yu XL (2013) Epitope screening of the PCV2 Cap protein by use of a random peptide-displayed library and polyclonal antibody. Virus Res 177:103–107
Guo L, Lu Y, Huang L, Wei Y, Liu C (2011) Identification of a new antigen epitope in the nuclear localization signal region of porcine circovirus type 2 capsid protein. Intervirology 54:156–163
Karuppannan AK, Opriessnig T (2017) Porcine Circovirus Type 2 (PCV2) Vaccines in the context of current molecular epidemiology. Viruses 9(5):99
Xu PL, Zhang Y, Zhao Y, Zheng HH, Han HY, Zhang HX, Zheng LL (2018) Detection and phylogenetic analysis of porcine circovirus type 3 in central China. Transbound Emerg Dis 00:1–7
Xiao CT, Halbur PG, Opriessnig T (2015) Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. J Gen Virol 96:1830–1841
Rozas J, Ferrer-Mata A, Sánchez-DelBarrio JC, Guirao-Rico S, Librado P, Ramos-Onsins SE, Sánchez-Gracia A (2017) DnaSP 6: DNA sequence polymorphism analysis of large data sets. Mol Biol Evol 34:3299–3302
Kosakovsky Pond SL, Frost SDW (2005) Not so different after all: a comparison of methods for detecting amino acid sites under selection. Mol Biol Evol 22:1208–1222
Weaver S, Shank SD, Spielman SJ, Li M, Muse SV, Kosakovsky Pond SL (2018) Datamonkey 2.0: A modern web application for characterizing selective and other evolutionary processes. Mol Biol Evol 35:773–777
Fu X, Fang B, Ma J, Liu Y, Bu D, Zhou P, Zhang G (2018) Insights into the epidemic characteristics and evolutionary history of the novel porcine circovirus type 3 in southern China. Transbound Emerg Dis 65(2):e296–e303
Qi S, Su M, Guo D, Li C, Wei S, Feng L, Sun D (2019) Molecular detection and phylogenetic analysis of porcine circovirus type 3 in 21 Provinces of China during 2015-2017. Transbound Emerg Dis 66:1004–1015
Zhai SL, Zhou X, Zhang H, Hause BM, Lin T, Liu R, Wang D (2017) Comparative epidemiology of porcine circovirus type 3 in pigs with different clinical presentations. Virol J 14:222
Chae C (2016) Porcine respiratory disease complex: Interaction of vaccination and porcine circovirus type 2, porcine reproductive and respiratory syndrome virus, and Mycoplasma hyopneumoniae. Vet J 212:1–6
Li G, He W, Zhu H, Bi Y, Wang R, Xing G, Zhang C, Zhou J, Yuen KY, Gao GF et al (2018) Origin, genetic diversity, and evolutionary dynamics of novel porcine circovirus 3. Adv Sci 5:1800275
Saraiva GL, Vidigal PMP, Assao VS, Fajardo MLM, Loreto ANS, Fietto JLR et al (2019) Retrospective detection and genetic characterization of porcine circovirus 3 (PCV3) strains identified between 2006 and 2007 in Brazil. Viruses 11(3):102
Acknowledgements
This research was supported by the Program of the Modern Agriculture Industry Technology System Foundation of Hebei Province (HBCT2018110207), Science and Technology Innovation Program of Hebei Province for Graduate Students (CXZZBS2019095), and Key R & D Projects in Hebei Province, China (19226622D).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Handling Editor: Ana Cristina Bratanich.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendix 1
Appendix 1
No. | Strain name | GenBank accession no. | Collection location | Collection date | Gene type |
|---|---|---|---|---|---|
1 | PCV3/CH/HB/CZ-2/2017 | MG727537 | Hebei | 2017 | Complete genome |
2 | PCV3/CH/HB/BD/2017 | MG727538 | Hebei | 2017 | Complete genome |
3 | PCV3/CH/HB/CZ-1/2017 | MG727539 | Hebei | 2017 | Complete genome |
4 | PCV3/CH/HB/SJZ/2017 | MG727540 | Hebei | 2017 | Complete genome |
5 | PCV3/CH/HB/HD/2017 | MH101645 | Hebei | 2017 | Complete genome |
6 | PCV3/CH/Tianjin-1/2018 | MH547276 | Tianjin | 2018 | Complete genome |
7 | PCV3/CH/Liaoning-1/2018 | MH547277 | Liaoning | 2018 | Full-length cap gene |
8 | PCV3/CH/Beijing-1/2016 | MH547278 | Beijing | 2016 | Full-length cap gene |
9 | PCV3/CH/Beijing-2/2018 | MH547279 | Beijing | 2018 | Full-length cap gene |
10 | PCV3/CH/Shanxi-1/2018 | MH547280 | Shanxi | 2018 | Full-length cap gene |
11 | PCV3/CH/Shanxi-2/2018 | MH548436 | Shanxi | 2018 | Full-length cap gene |
12 | PCV3/CH/Shandong-1/2018 | MH548437 | Shandong | 2018 | Full-length cap gene |
13 | PCV3/CH/Liaoning-2/2018 | MH548438 | Liaoning | 2018 | Full-length cap gene |
14 | PCV3/CH/Tianjin-2/2017 | MH579786 | Tianjin | 2017 | Full-length cap gene |
15 | PCV3/CH/HB-CD/2018 | MH636587 | Hebei | 2018 | Full-length cap gene |
16 | PCV3/CH/HB-ZJK/2018 | MH636589 | Hebei | 2018 | Full-length cap gene |
17 | PCV3/CH/HB-TS/2018 | MH636590 | Hebei | 2018 | Full-length cap gene |
Rights and permissions
About this article

Cite this article
Gu, W., Shi, Q., Wang, L. et al. Detection and phylogenetic analysis of porcine circovirus 3 in part of northern China from 2016 to 2018. Arch Virol 165, 2003–2011 (2020). https://doi.org/10.1007/s00705-020-04709-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-020-04709-0