
Abstract
Working memory impairment in schizophrenia has been strongly associated with abnormalities in gamma oscillations within the dorsolateral prefrontal cortex (DLFPC). We recently published the first ever study showing that anodal transcranial direct current stimulation (tDCS) to the left DLPFC was able to significantly improve working memory in schizophrenia and did so seemingly via restoring normal gamma oscillatory function. Transcranial alternating current stimulation (tACS) is a form of electrical brain stimulation that delivers stimulation at a specific frequency and has been shown to entrain endogenous cortical oscillations. Therefore, gamma (γ) tACS may be even more effective than tDCS in improving working memory in schizophrenia. In a randomized repeated-measures study we compared the effects of γ-tACS, tDCS and sham stimulation on the performance of the two back working memory tasks in ten patients with schizophrenia. There was a significant time by stimulation interaction, with tDCS and sham showing trend-level improvements in working memory, while γ-tACS, contrary to our hypothesis, showed no change. The results are discussed in light of posited divergent effects of tACS and tDCS on the pathophysiology of working memory impairment in schizophrenia.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Working memory impairment in schizophrenia is common and associated with functional disability in areas such as work and educational functioning (Nuechterlein et al. 2011; Shamsi et al. 2011). This is not surprising, as working memory capacity has been shown to be strongly related to broader cognitive impairment in schizophrenia, including in areas such as visual/verbal learning, problem solving, and social cognition (Johnson et al., 2013). The current treatment approaches for these disabling cognitive symptoms consist of pharmacotherapy, shown to have very limited effects, or cognitive remediation which has resulted in modest improvements at best (Elie et al. 2010; Lett et al. 2014; Wykes et al. 2011). The lack of efficacy of current treatments is likely due to the fact that they do not address the underlying pathophysiology of these symptoms (Insel 2010).
Working memory impairment in schizophrenia has been related to dysfunctions in many areas from genetic expression to neurotransmitter function (Millan et al. 2012). However, one of the more promising theories to date suggests a framework for linking these findings and provides a viable target for treatment (Dobbs 2010). Essentially, the theory proposes that a potential point of convergence of these dysfunctions is a ‘weakness’ in GABA-ergic inhibitory neurons in the prefrontal cortex which results in an inability to generate the coordinated firing of neurons, known as cortical oscillations, which are required for successful working memory (Dobbs 2010; Chen et al. 2014). Indeed, abnormalities in GABA-ergic activity are a consistent finding in schizophrenia and have been reliably associated with impaired cortical oscillatory activity within the dorsolateral prefrontal cortex (DLFPC), specifically in the gamma frequency range (>40 Hz), as well as working memory impairment (Chen et al. 2014; Lett et al. 2014). Cognitive functions such as working memory are complex and as such engage cortical oscillations from multiple frequency bands independently (i.e. theta, alpha, beta, and gamma), as well as frequency-coupled oscillatory activity (i.e. theta-nested-gamma) (Howard et al. 2003; Sauseng et al. 2009). There is considerably greater evidence, however, for impaired gamma oscillations being associated with working memory dysfunction in schizophrenia (Gonzalez-Burgos et al. 2011).
The body of research with respect to impaired gamma oscillations in schizophrenia is quite mixed with respect to whether oscillations are reduced or excessive. A comprehensive review of this literature Sun et al. (2011) concluded that these seemingly inconsistent findings reflect the situation where gamma is “not optimally regulated in schizophrenia”, thus leading to insufficient activity when gamma is required and overactivation when gamma should be suppressed. Therefore, it appears to be an impairment in the modulation of gamma oscillations, likely due to impaired GABA-ergic activity, which is associated with working memory dysfunction in schizophrenia. Such abnormalities, i.e. dysfunctional cortical inhibition and gamma activity, are therefore promising treatment targets and have been shown to be amendable to therapeutic approaches using brain stimulation techniques (Barr et al. 2012; Hoy et al. 2014, 2015a , b).
We recently showed that transcranial direct current stimulation (tDCS) in particular was able to enhance working memory in schizophrenia and appeared to do so by restoring the patient’s ability to modulate gamma activity in response to task demands (Hoy et al. 2014, 2015a, b). TDCS involves the application of a weak electrical current between two electrodes (an anode and a cathode) applied to the scalp. The current alters the excitability of brain cells by shifting their membrane potentials in a de- or hyperpolarizing direction, making them more or less likely to fire (Nitsche and Fregni 2007). Stimulation of brain cells under the anode appears to increase the brain’s natural firing rate, whereas stimulation under the cathode generally has the opposite effect (Jacobson et al. 2012). Magnetic resonance spectroscopy (MRS) findings indicate that the excitatory effects of anodal tDCS are due, in part, to the direct modulation of GABA-ergic activity (Stagg et al. 2009, 2011). Therefore, we theorized that the improvements we saw in patients’ ability to modulate gamma, and thus improve their working memory performance, were due to the GABA-ergic effects of anodal tDCS (Hoy et al. 2015a, b). Specifically, by modulating GABA-ergic activity, tDCS was able to induce a more optimal excitation (E)/inhibition (I) balance, subsequently providing the required cortical ‘environment’ for normal gamma modulation in patients with schizophrenia (Hoy et al. 2015a, b; Krause et al. 2013). These findings clearly indicate that tDCS has potential for treating working memory impairment in schizophrenia. However, there are also stimulation techniques that can directly entrain specific bands of synchronous activity, such as transcranial alternating current stimulation (tACS), which may hold even greater therapeutic promise.
tACS differs from tDCS in that the electrical current delivered alternates at a specific frequency back and forth between the electrodes, as opposed to tDCS where the current is unidirectional (Helfrich et al. 2014). A number of studies have shown that stimulation with tACS can result in entrainment of endogenous oscillations at the frequency of stimulation (Herrmann et al. 2013). The ability to entrain cortical oscillations theoretically allows for more direct enhancement of the impaired neural activity underlying the dysfunction, in this case gamma (γ)-tACS to entrain gamma activity in schizophrenia, which could in turn produce more robust behavioural improvements (Herrmann et al. 2013). To date, there has been no research looking at the effect of γ-tACS on working memory performance in schizophrenia.
Determining the optimal brain stimulation techniques and parameters for enhancing working memory in schizophrenia is crucial prior to the conduct of large-scale clinical trials. Therefore, the aim of the current study was to investigate the effect of γ-tACS on working memory performance in patients with schizophrenia compared to an active comparator (standard tDCS) and a placebo control (sham tDCS). It was hypothesized that γ-tACS and tDCS would result in significantly greater overall improvements in WM than sham, and that the improvements seen with γ-tACS would be superior to tDCS.
Method
Participants
Eleven patients with a diagnosis of schizophrenia or schizoaffective disorder were recruited into the study (See Tables 1, 2 for demographic and clinical data). Diagnosis was confirmed using the Mini-International Neuropsychiatric Interview (MINI) (Sheehan et al. 1998). The MINI was administered by trained researchers experienced in its use (KH/DW). All 11 participants were regularly taking antipsychotics (i.e. 2 aripiprazole; 2 risperidone; 2 olanzapine; 1 amisulpride; 1 clozapine + aripiprazole; 1 quetiapine + ziprasidone; 1aripiprazole + quetiapine; 1 clozapine + olanzapine + aripiprazole). There are numerous known, and likely unknown, interaction effects between medication and tDCS (Brunoni et al. 2012). In light of this, participants were required to remain on their current dose and type of medication throughout participation in the study (Hoy et al. 2014, 2015a, b). Participants were excluded if they had a history of any neurological or serious medical conditions, or were currently pregnant. Written consent was obtained from participants prior to commencement of the study. Ethical approval was granted by the Monash University and the Alfred Hospital ethics committees.
Procedure
This was a randomized repeated-measures single-blind study design. Participants attended the research centre on three separate occasions at least 72 h apart. Each testing session involved 20 min of stimulation (either γ-tACS, tDCS or sham tDCS), with working memory assessed using the 2-back task, both prior to and immediately following stimulation. The active/anodal electrode was located over F3 and the reference/cathode over the right supraorbital region, which is the standard montage for left DLFPC stimulation (Nitsche and Fregni 2007). Concurrent with stimulation, participants undertook two 5 min blocks of the 2-back task (from 2.5 to 7.5 and from 12.5 to 17.5 min), See Fig. 1. The stimulation conditions for the three repeated sessions were randomized and counterbalanced.

Illustration of experimental setup and protocol. a An anodal/active electrode was placed over F3 (left DLPFC) and cathodal/reference electrode over the right supraorbital space. b Each participant underwent three experimental sessions spaced at least 72 h apart. γ-tACS, tDCS and sham tDCS were randomly applied for 20 min (with concurrent 2-back performed from 2.5 to 7.5 and 12.5–17.5 min) across the 3 weeks. Working memory assessments pre- and post-stimulation used the 2-back. c Illustration of 2-back task
Electrical stimulation
The Elidth Stimulator Plus was used for all forms of stimulation (alternating current, direct current and sham). The settings required for the three different stimulation conditions does not allow experimenter blinding; therefore, the design was single blind. With respect to participant blinding, all participants were informed that they would receive two active and one sham stimulation session with no information as to the specific nature of the stimulation types. Blinding questionnaires were also conducted at the end of each session. Stimulation was delivered through two 35 cm2 (7 × 5 cm) electrodes covered with sponges soaked in a saline solution (0.09 %).
γ-tACS
tACS was delivered at a current of between −1000 to +1000 µA at 40 Hz for 48,000 cycles, or 20 min with no current offset to avoid the impact of static polarity effects. The active electrode was positioned over the left DLPFC (i.e. F3) and the reference electrode placed over the contralateral supraorbital area. The stimulator was set to fade in and out over the maximum period possible, which was 100 cycles.
tDCS
Anodal tDCS was applied at 2 mA for 20 min, with a fade in of 120 s and a fade out of 15 s. The anode was placed over the left DLPFC and the cathode over the contralateral supraorbital area.
Sham tDCS
Sham stimulation began with a fade in over 120 s to a peak of 2 mA, followed immediately by constant current stimulation for 30 s and a fade out of 15 s. This is the standard blinding procedure for tDCS (Nitsche and Fregni 2007). The anodal electrode was again placed over F3 and the cathode over the contralateral supraorbital area.
Working memory task: 2-back
Pre- and post-stimulation task
A series of quasi-random letters from A to J were presented consecutively in a 5-min block. Participants were required to respond with a button press if the letter was the same as that presented two trials earlier. Each block consisted of 130 trials containing 25 % targets. Letters were presented for 500 ms and there was a 1500 ms delay between stimuli presentations.
Intra-stimulation task
Participants also completed two blocks of the 2-back during stimulation, from 2.5 to 7.5 min (IS-1) and 12.5–17.5 min (IS-2). The intra-stimulation blocks used the same task design as the pre- and post-stimulation task described above.
Therefore, each participant undertook the 2-back four times per session, over the three sessions and this equated to a total of 12 blocks of 2-back. Alternate stimuli were used for each of these blocks.
Statistical analysis
Data were analysed from 10 out of the 11 participants (one participant’s data was excluded as they did not engage in any of the working memory assessments over the three sessions). The dependent variables were d prime and accurate reaction time. d prime is a discriminability index which takes into account the ability to correctly identify targets and to minimize false alarms. Specifically, it is calculated using the formula d prime = ZHit−ZFA, where Hit = hits/(hits + misses), known as the hit rate, and FA = false alarms/(false alarms + correct negative), known as the false alarm rate (Haatveit et al. 2010). Therefore, d prime is the difference between the Z transforms of these two rates and has been shown to have high sensitivity in schizophrenia (Haatveit et al. 2010). We conducted repeated-measures ANOVAs for both dependent variables with stimulation type (γ-tACS, tDCS, Sham) and time (pre, post) as within subject factors. Post hoc analysis of simple main effects was conducted using pairwise comparisons, while significant interactions were further explored using one-way ANOVAS and t tests measuring the effect of stimulation at each time point as well as change over time for each stimulation condition. We used the same statistical approach for analyses of performance on the intra-stimulation task at the two time points, i.e. IS-1 and IS-2. Results were assessed using two-tailed tests with a significance alpha level of 0.05, with less than 0.07 considered a trend.
Results
Order effects analysis confirmed the effectiveness of the counterbalancing of sessions, with no significant session order effects seen in either d prime (F (2, 18) = 0.594, p = 0.594) or accurate reaction time (F (2, 18) = 1.095, p = 0.356). With respect to blinding, participants were not able to guess the stimulation condition better than chance across the three sessions: session one [χ 2(1, N = 10) = 0.500, p = 1.000], session two (χ 2(1, N = 10) = 0.500, p = 1.000] and session three [χ 2(1, N = 10) = 0.600, p = 0.527].
Baseline differences
There was no significant difference at baseline between the three stimulation conditions for either d prime (F (2, 18) = 1.831, p = 0.19) or reaction time (F (2, 18) = 0.104, p = 0.90).
Pre- to post-stimulation performance
d prime
Means and standard deviations for 2-back d prime are provided in Table 3.
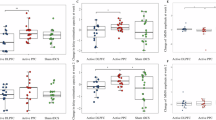
There was no main effect of stimulation on performance (F (2, 18) = 1.451, p = 0.26); there was however a significance time by stimulation interaction (F (2, 18) = 4.404, p = 0.03) (see Fig. 2).

Means and standard errors of d prime pre-and post-stimulation across the three conditions; γ-tACS, tDCS and sham
Post hoc analyses within stimulation conditions revealed a trend level improvement following tDCS (t (9) = 2.078, p = 0.06; d = 0.65) and sham (t (9) = 2.024, p = 0.07), with no change over time for γ-tACS (t (9) = −0.777, p = 0.45). Analysis of the post-stimulation time point revealed significantly greater d prime following tDCS condition compared to γ-tACS (mean difference = 0.61; p = 0.005), with no difference between tDCS and sham (mean difference = 0.42, p = 0.17) or sham and γ-tACS (mean difference = 0.193, p = 0.51). There was no effect of stimulation at the pre-stimulation time point.
Reaction time
Means and standard deviations for reaction time are provided in Table 3.
There was no main effect of stimulation on reaction time (F (2, 18) = 0.029, p = 0.97), nor was there a stimulation by time interaction (F (2, 18) = 0.37, p = 0.69).
Intra-stimulation performance
d prime
Means and standard deviations for 2-back d prime during stimulation are provided in Table 4.
There was no main effect of stimulation on reaction time (F (2, 18) = 0.671, p = 0.52), nor was there a stimulation by time interaction (F (2, 18) = 0.647, p = 0.54).
Reaction time
Means and standard deviations for intra-stimulation reaction time are provided in Table 4.
There was no main effect of stimulation on reaction time (F (2, 18) = 0.211, p = 0.81), nor was there a stimulation by time interaction (F (2, 18) = 1.600, p = 0.23).
Tolerability
Both tACS and tDCS were generally well tolerated. 64 % (7/11) of participants reported mild site discomfort described as ‘itch’ or ‘warmth’; 27 % (3/11) reported no side effects; and 9 % (1/11) described moderate site discomfort during the initial 5 min of stimulation.
Discussion
We found a significant interaction between stimulation type and time on working memory accuracy as assessed using the 2-back. Specifically, patients showed a trend towards significantly improved performance following tDCS and sham, while, contrary to our hypothesis, there was no change in performance following γ-tACS. There was no difference in performance at baseline between the three stimulation sessions, and at the post-stimulation time point tDCS resulted in significant better performance than γ-tACS. There was no effect of stimulation on accurate reaction time for the 2-back. With respect to intra-stimulation performance, there was no significant difference in performance in either d prime or accurate reaction time in the 2-back between stimulation conditions or across time. The results of this preliminary investigation do not support our hypothesis. While tDCS did show a positive effect over time, essentially inducing a slightly greater learning effect than what was seen in sham, γ-tACS did not result in any change in performance over time.
There has been a small amount of research to date looking at the cognitive effects of tACS, all of which has been in healthy populations. These studies have shown significant improvements in cognition following tACS in the alpha, theta and gamma ranges, with the majority showing positive effects on the more robust measure of accuracy rather than reaction time (Helfrich et al. 2014; Jaušovec and Jaušovec 2014; Jaušovec et al. 2014; Santarnecchi et al. 2013). We also recently showed, in a similar protocol to the current study conducted in healthy controls, that γ-tACS significantly enhanced working memory as a function of load, with significant and large improvements seen only at higher working memory loads (Hoy et al. 2015a, b). Therefore, the current findings, i.e. a lack of improvement following γ-tACS, are inconsistent with the previous healthy control literature. This may be due to the divergent effects of tACS on the neurophysiology of the healthy brain as compared to schizophrenia, a theory we discuss in more detail below. With respect to the effects of tDCS, our data largely supports previous research, which consistently shows improvement in cognitive functioning in patients with schizophrenia following anodal tDCS (Hoy et al. 2014; Rassovsky et al. 2015; Reinhart et al. 2015; Smith et al. 2015; Vercammen et al. 2011). While there was improvement in 2-back performance in the current study following tDCS, in contrast with our previous findings, this improvement did not reach significance (Hoy et al., 2014). This is likely due to the fact that in the current study, the ‘natural’ learning effect (as measured in the sham condition) was greater than that seen in our original studies and subsequently the improved performance in the tDCS condition did not reach significance. There are a number of potential explanations for this. Firstly, the experimental paradigm employed across the two studies was markedly different; namely the current study involved performance of the 2-back on four occasions at intervals of between 2.5 and 5 min, while the 2014 study required participants to perform the task three times at intervals of between 10 and 15 min. Both the reduced time between practices and the additional performance could have resulted in the greater ‘natural’ learning effect (Goldberg et al. 2010). Additionally, the current study utilized an online paradigm, whereby the cognitive task was conducted during stimulation, as well as pre and post, which may have influenced the magnitude of effect in the tDCS group. This is consistent with our recently published findings of the essentially the same methodology in healthy controls (Hoy et al. 2015a, b).
The apparent inconsistency in the effects of tDCS and γ-tACS on working memory performance in schizophrenia may be related to divergent effects on pathophysiology.
We have previously shown that tDCS enhanced working memory in schizophrenia seemingly by restoring normal gamma oscillatory function, most likely via indirect effects of tDCS on GABA-ergic activity (Hoy et al. 2014, 2015a, b). Therefore, in light of the importance of gamma activity for working memory in schizophrenia, we hypothesized that tACS applied within the gamma range (γ-tACS) would result in significantly greater improvements through a process of direct gamma entrainment. We propose that the lack of effect seen in the current study is most likely due to an inability of tACS to ‘entrain’ oscillatory activity in the absence of an innate ability to generate/modulate endogenous oscillations, as is the case in schizophrenia (Sun et al. 2011). Studies have shown that abnormalities in gamma activity in schizophrenia are essentially a downstream consequence of impairments in cortical inhibition; therefore, it may be that alteration of GABA-ergic activity is required for restoration of gamma modulation in this patient group—as is the posited mechanism of action of tDCS (Hoy et al. 2015a, b).
In light of the absence of a learning effect in the γ-tACS condition it could be also argued that this stimulation actively interfered with the patient’s learning ability. Either through disruption of the patient’s ability to intrinsically entrain gamma to a degree to meet task demands; or, from a more practical standpoint, that the visual phosphenes induced with γ-tACS interfered with patient’s ability to engage in the intra-stimulation task lessening the behavioural ‘learning opportunity’ that was provided in the sham and tDCS conditions. However, such an explanation is not supported by the data. There was no effect of stimulation, nor a stimulation by time interaction, for performances on the intra-stimulation task. Indeed, looking at the means γ-tACS showed the same pattern of performance across these two time points as was seen in sham (i.e. non-significant decrease), while tDCS showed a small mean improvement. These data are more supportive of tDCS enhancing an intrinsic learning effect and γ-tACS showing an absence of effect.
This preliminary investigation of the effects of γ-tACS on working memory enhancement in schizophrenia indicates that this form of stimulation is not superior to tDCS in inducing behavioural change. These findings need to be considered in light of a number of limitations. Firstly, as stated, this is a preliminary investigation and included only ten patients. These findings would need to be confirmed in a larger sample size. Also, in the current study we applied γ-tACS without an offset, to avoid the impact of static polarity effects; however, it may be that the provision of γ-tACS with a DC offset to induce excitability changes is required for gamma entrainment with tACS in schizophrenia. In addition, there is the potential that the ‘reference’ electrode, particularly in the tACS condition, has active effects. Therefore, the results could be contributed to modulation of activity in regions such as the orbitofrontal cortex. It is not possible to ascertain such affects without utilizing neurophysiological outcomes and field modelling in future studies. Indeed, inclusion of neurobiological measures in future research would provide important information as to the mechanisms of actions of these varying stimulation approaches.
References
Barr MS, Farzan F, Rajji TK, Voineskos AN, Blumberger DM, Arenovich T et al (2012) Can repetitive magnetic stimulation improve cognition in schizophrenia? Pilot data from a randomised controlled trial. Biol Psychiatry 73:510–517
Brunoni AR, Nitsche MA, Bolognini N, Bikson M, Wagner T, Merabet L, Edwards DJ, Valero-Cabre A, Rotenberg A, Pascual-Leone A, Ferrucci R (2012) Clinical research with transcranial direct current stimulation (tDCS): challenges and future directions. Brain Stimul 5(3):175–195
Chen CMA, Stanford AD, Mao X, Abi-Dargham A, Shungu DC, Lisanby SH et al (2014) GABA level, gamma oscillation, and working memory performance in schizophrenia. Neuroimage 4:531–539
Dobbs D (2010) The making of a trouble mind. Nature 468:154–156
Elie D, Poirier M, Chianetta JM, Durand M, Grégoire CA, Grignon S (2010) Cognitive effects of antipsychotic dosage and polypharmacy. J Psychopharmacol 24(7):1037–1044
Goldberg TE, Keefe RS, Goldman RS, Robinson DG, Harvey PD (2010) Circumstances under which practice does not make perfect: a review of the practice effect literature in schizophrenia and its relevance to clinical treatment studies. Neuropsychopharmacology 35(5):1053–1062
Gonzalez-Burgos G, Fish KN, Lewis DA (2011) GABA neuron alterations, cortical circuit dysfunction and cognitive deficits in schizophrenia. Neural Plast 2011:723184. doi:10.1155/2011/723184
Haatveit BC, Sundet K, Hugdahl K, Ueland T, Melle I, Andreassen OA (2010) The validity of d prime as a working memory index: results from the “Bergen n-back task”. J Clin Exp Neuropsychol 32(8):871–880
Helfrich RF, Schneider TR, Rach S, Trautmann-Lengsfeld SA, Engel AK, Herrmann CS (2014) Entrainment of brain oscillations by transcranial alternating current stimulation. Curr Biol 24(3):333–339
Herrmann CS, Rach S, Neuling T, Strüber D (2013) Transcranial alternating current stimulation: a review of the underlying mechanisms and modulation of cognitive processes. Front Hum Neurosci. doi:10.3389/fnhum.2013.00279
Howard MW, Rizzuto DS, Caplan JB, Madsen JR, Lisman J, Aschenbrenner-Scheibe R et al (2003) Gamma oscillations correlate with working memory load in humans. Cereb Cortex 13(12):1369–1374
Hoy KE, Arnold SL, Emonson MR, Daskalakis ZJ, Fitzgerald PB (2014) An investigation into the effects of tDCS dose on cognitive performance over time in patients with schizophrenia. Schizophr Res 155(1):96–100
Hoy KE, Bailey NW, Arnold SL, Fitzgerald PB (2015a) The effect of tDCS on gamma activity and working memory in schizophrenia. Psychiatry Res 228(2):191–196
Hoy KE, Bailey N, Arnold S, Windsor K, John J, Daskalakis ZJ, Fitzgerald PB (2015b) The effect of γ-tACS on working memory performance in healthy controls. Brain Cogn 101:51–56
Insel TR (2010) Rethinking Schizophrenia. Nature 468:187–193
Jacobson L, Koslowsky M, Lavidor M (2012) tDCS polarity effects in motor and cognitive domains: a meta-analytic review. Exp Brain Res 216:1–10
Jaušovec N, Jaušovec K (2014) Increasing working memory capacity with theta transcranial alternating current stimulation (tACS). Biol Psychol 96:42–47
Jaušovec N, Jaušovec K, Pahor A (2014) The influence of theta transcranial alternating current stimulation (tACS) on working memory storage and processing functions. Acta Psychol 146:1–6
Johnson MK, McMahon RP, Robinson BM, Harvey AN, Hahn B, Leonard CJ et al (2013) The relationship between working memory capacity and broad measures of cognitive ability in healthy adults and people with schizophrenia. Neuropsychology 27:220
Krause B, Márquez-Ruiz J, Kadosh RC (2013) The effect of transcranial direct current stimulation: a role for cortical excitation/inhibition balance? Front Hum Neurosci. doi:10.3389/fnhum.2013.00602
Lett TA, Voineskos AN, Kennedy JL, Levine B, Daskalakis ZJ (2014) Treating working memory deficits in schizophrenia: a review of the neurobiology. Biol Psychiatry 75(5):361–370
Millan MJ, Agid Y, Brüne M, Bullmore ET, Carter CS, Clayton NS et al (2012) Cognitive dysfunction in psychiatric disorders. Nat Rev Drug Discov 11:141–168
Nitsche MA, Fregni F (2007) TDCS—an adjuvant tool for the treatment of neuropsychiatric diseases? Curr Psychiatry Rev 3:222–232
Nuechterlein KH, Subotnik KL, Green MF, Ventura J, Asarnow RF, Gitlin MJ et al. (2011) Neurocognitive predictors of work outcome in recent-onset schizophrenia. Schizophr Bull 37(suppl 2):S33–S40
Rassovsky Y, Dunn W, Wynn J, Wu AD, Iacoboni M, Hellemann G et al (2015) The effect of transcranial direct current stimulation on social cognition in schizophrenia: a preliminary study. Schizophr Res 165(2):171–174
Reinhart RM, Zhu J, Park S, Woodman GF (2015) Synchronizing theta oscillations with direct-current stimulation strengthens adaptive control in the human brain. Proc Natl Acad Sci 112(30):9448–9453
Santarnecchi E, Polizzotto NR, Godone M, Giovannelli F, Feurra M, Matzen L et al (2013) Frequency-dependent enhancement of fluid intelligence induced by transcranial oscillatory potentials. Curr Biol 23(15):1449–1453
Sauseng P, Klimesch W, Heise KF, Gruber WR, Holz E, Karim AA, Hummel FC (2009) Brain oscillatory substrates of visual short-term memory capacity. Curr Biol 19(21):1846–1852
Shamsi S, Lau A, Lencz T, Burdick KE, DeRosse P, Brenner R et al (2011) Cognitive and symptomatic predictors of functional disability in schizophrenia. Schizophr Res 126:257–264
Sheehan DV, Lecrubier Y, Sheehan KH, Amorim P, Janavs J, Weiller E, Hergueta T, Baker R, Dunbar GC (1998) The Mini-International Neuropsychiatric Interview (MINI): the development and validation of a structured diagnostic psychiatric interview for DSM-IV and ICD-10. J Clin Psychiatry 59(Suppl 20):22–33
Smith RC, Boules S, Mattiuz S, Youssef M, Tobe RH, Sershen H et al (2015) Effects of transcranial direct current stimulation (tDCS) on cognition, symptoms, and smoking in schizophrenia: a randomized controlled study. Schizophr Res 126:257–264
Stagg CJ, Best JG, Stephenson MC, O’Shea J, Wylezinska M, Kincses ZT et al (2009) Polarity-sensitive modulation of cortical neurotransmitters by transcranial stimulation. The Journal of Neuroscience 29(16):5202–5206
Stagg CJ, Bachtiar V, Johansen-Berg H (2011) The role of GABA in human motor learning. Curr Biol 21(6):480–484
Sun Y, Farzan F, Barr MS, Kirihara K, Fitzgerald PB, Light GA et al (2011) Gamma oscillations in schizophrenia, mechanisms and clinical significance. Brain Res 1413:98–114
Vercammen A, Rushby JA, Loo C, Short B, Weickert CS, Weickert TW (2011) Transcranial direct current stimulation influences probabilistic association learning in schizophrenia. Schizophr Res 131(1):198–205
Wykes T, Huddy V, Cellard C, McGurk SR, Czobor P (2011) A meta-analysis of cognitive remediation for schizophrenia: methodology and effect sizes. Am J Psychiatry 168:472–485
Acknowledgments
KH is supported by a National Health and Medical Research Council (NHMRC) Career Development Fellowship. PF is supported by an NHMRC Practitioner Fellowship.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
PF has received equipment for research from Brainsway Ltd, Medtronic Ltd and MagVenture A/S, and funding for research from Cervel Neurotech and Neuronetics Ltd. PF has received consultancy fees as a scientific advisor for Bionomics. There are no other relevant conflicts of interest.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Hoy, K.E., Whitty, D., Bailey, N. et al. Preliminary investigation of the effects of γ-tACS on working memory in schizophrenia. J Neural Transm 123, 1205–1212 (2016). https://doi.org/10.1007/s00702-016-1554-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00702-016-1554-1