
Abstract
To date, few attempts have been made to assess the influence of climate change on forest ecosystems and on the relationship between tree growth and climate in humid areas of low latitudes. In this paper, we studied the response of tree growth and forest ecosystem to climate change by using Fokienia hodginsii tree-ring cores from the northern Yunnan-Guizhou Plateau, southwest of China. Tree growth correlates the highest (r = −0.64, p < 0.01) with mean temperature (July–September), but the coefficients were changing with time as revealed by a moving correlation analysis. Tree growth is significantly (p < 0.05) and positively correlated with January–April mean temperature from AD 1961–1987, while correlations with precipitation are insignificant. In contrast, from 1988 to 2014, tree growth correlated negatively with mean temperature of previous summer and positively with precipitation of previous August–September. This indicated that the limiting factors for tree growth have changed under different climate conditions. The meteorological data suggested that from 1961 to 1987 it was cold and wet in the study area and radial growth is limited by winter and spring temperatures. This restriction is weaker if the climate is appropriate in general. However, from 1988 to 2014, the combined effects of recent warming and decreasing precipitation have led to an increasing response of tree-ring width to drought. In addition, a large proportion of mature F. hodginsii mortality occurred from 2007 to 2013, which corresponds with a drastic reduction of radial growth (narrowest in recent 100 years). The recent drought, induced by decreasing precipitation and increasing temperature, may have passed the threshold which F. hodginsii could tolerate, causing tree growth reduction, tree growth-climate relationship change, as well as catastrophic tree mortality. All these changes may lead to further responses of the local ecosystem to climate change which should be highly regarded.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Global surface mean temperature has increased by 0.85 ± 0.20 °C in about the last 130 years (1880–2012) (IPCC 2013), and this trend is supposed to continue in the near future. Global warming has caused a large number of extreme weather and climate events (drought, flood, snow disaster) across the world (Alexander et al. 2006; Anderegg et al. 2015; Easterling et al. 2000; Min et al. 2015). In recent years, a severe drought struck the southwest of China, which has brought enormous socio-economical losses. The impact of extreme climate events on regional societies and environments has received widespread attention (Allen et al. 2010; Easterling et al. 2000; Walther et al. 2002).
Tree rings have been widely used in paleoclimatic studies as a high-resolution proxy around the world. However, tree-ring research in China is focused on northwest China, Tibetan Plateau, and the surrounding area (Fang et al. 2010b; Gao et al. 2015; Gou et al. 2014; Liang et al. 2008, 2009; Yang et al. 2014; Zhang et al. 2015), but it is still scarce in the low latitudes and humid southern China. There are many difficulties for tree-ring research in humid regions, cause by vegetation deterioration, richness in tree species, and an only weak climate limitation of tree growth. Tree growth may also reflect information about microclimate, competition between individuals and species, as well as human and other biotic activities. To date, there are some tree-ring-width and isotope studies in southern China (An et al. 2012; Duan et al. 2013; Li et al. 2016; Qian et al. 2002; Shi et al. 2013, 2015). However, still few attempts have been made to assess the influence of climate change (especially the recent drought in southwest China) on forest ecosystem and the relationship between tree growth and climate.
A stable relationship between tree-ring width and climate is a basic principle of dendroclimatology. However, a “divergence phenomenon,” expressing a remarkable instability in the climate sensitivity of tree growth, was found in recent decades. For example, D’Arrigo et al. (2008) reported that the temperature signal in white spruce ring width and maximum latewood-density chronologies from altitudinal and latitudinal tree-line sites in interior and northern Alaska has weakened in recent decades. Schneider et al. (2014) found that the strong coherence between tree-ring data and instrumental climate data in northern Fennoscandia is interrupted by a short, but significant correlation decrease in the early twentieth century; they ascribed this phenomenon to a substantial reduction in interannual summer temperature variability during that time period. D’Arrigo et al. (2008) reviewed the literature on the “divergence problem” and concluded that a clear identification of a sole cause for the divergence is probably unlikely due to the combination of reasons, such as the varying locations, species, and other factors. The divergence problem has caused extensive concern and challenged the traditional tree-ring-based climate reconstructions. This problem may have potentially significant implications for large-scale patterns of forest growth, the paleoclimatic reconstructions based on tree-ring records, as well as the global carbon cycle. However, on a global scale, the divergence problem research is focusing on middle and high latitudes (Briffa et al. 1998; Carrer and Urbinati 2006; Esper and Frank 2009; Jacoby and D’Arrigo 1995; Schneider et al. 2014); In China, it focuses on arid areas in the northwest of the country, on the Tibetan Plateau, and in surrounding areas (Gao et al. 2011; Jiao et al. 2015; Yu et al. 2013; Zhang et al. 2009), but it is very scarce in humid low latitudes (Fang et al. 2010a; Li et al. 2016). Therefore, it is meaningful to carry out such research in these areas.
Drought-related tree mortality has also given wide attention. Increasing tree mortality rates have been reported for boreal forests, temperate forests, tropical forests, and savannas (Peng et al. 2011; Phillips et al. 2010; van Mantgem et al. 2009). Suresh et al. (2010) analyzed the mean annual mortality rate over a 19-year period (1988–2007) of all trees with stems thicker than 1 cm dbh, and found that tree mortality can be categorized into several major causes. Fire, elephant-herbivory and climate change were the main agents for the mortality of trees with a stem of 1–5 cm, 5–10 cm, and thicker than 10 cm of dbh. Additionally, tropical dry forests growing in regions of a high rainfall variability may have developed greater resistance to rainfall deficit in comparison with tropical moist or temperate forests. Drought may tend to impact some tree species, as Fensham and Holman (1999) found in northeast Australia. Species with a shallow root system will have a high-risk to suffer severe water stress in semi-arid environments, while competition for soil moisture of different species resulted in a patchiness of drought-induced tree death. Moreover, Slik (2004) proposed that severe drought can regulate the structure of a plant community and affect the recruitment and growth conditions of small and immature trees. Tree death, as a fundamental process in ecosystems, has a substantial leverage in forest ecosystem responses to global environment change (Allen et al. 2010; Franklin et al. 1987). Recently, a severe drought impacted southwestern China, but to date, no attempt has been made to assess the influence of this extreme climate event on forest ecosystems.
Here, we present the first tree-ring-width chronology from a long-lived conifer, Fokienia hodginsii in the Xishui National Nature Reserve of the northern Yunnan-Guizhou Plateau. This paper explores the temporal stability of the relationship between tree growth and climate, the possible reason of drastic tree growth reduction and catastrophic tree mortality. This study is an expansion of tree-ring research in humid areas, and it will provide important information for policy decisions, rare species protection, and forest management.
Materials and methods
Research area
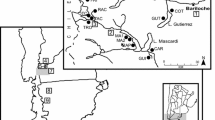
Our study was conducted in the Xishui National Nature Reserve, in the northwestern Guizhou province, China (105° 50′~106° 29′ E, 28° 07′~28° 34′ N, 420–1756 m asl) (Li et al. 2015) (Fig. 1). With an undulating topography with steep hills, this national wildlife refuge is trackless. This area features a subtropical monsoon climate, and the 56-year-long records of monthly mean temperature and monthly total precipitation at the Xishui weather station (Fig. 2) represent the general climate of the study region. The mean annual precipitation is 1096 mm, 58.8% of which fall in the monsoon months from May to August. The average annual temperature is 13.1 °C, and the hottest season is July to August. Though not found in pure stands, F. hodginsii is one of several canopy species such as Masson pine, Pinus fenzeliana, and Gordonia acuminata, which are scattered throughout dense and tall mixed forest stands with multiple canopy layers consisting of both evergreen and deciduous tree species. Our F. hodginsii community in this study area is known as one of the highest latitude natural community for this species. This species is mainly distributed in Vietnam and south-southwest China (110°~118° E, 24° 30′~26° 30′ N) (Wang et al. 1984).

Location of the study region in China (inset); the location of the sampling site, the nearby meteorological station, and scPDSI grid point are shown in the detailed map

Monthly total precipitation and monthly mean temperature of Xishui from 1959 to 2014
Climate data
The Xishui station (28° 19′ 48″ N, 106° 13′ 12″ E, 1180.2 m asl) (Fig.1) is the closest meteorological station to the sampling site (~30 km). Climate variables for climate-growth analyses include monthly mean temperature and total precipitation from 1959 to 2014. In addition, the nearest grid point (28.75° N, 106.25° E) of the monthly self-calibrating Palmer Drought Severity Index (scPDSI) dataset was downloaded from http://climexp.knmi.nl. This drought index was first proposed by Wells et al. (2004) to quantitatively measure the accumulated deficit of local mean moisture conditions, allowing comparisons of drought severities across regions and time.
We use the linear regression method to analyze the variations of climate recorded at the Xishui station (Fig. 3) since 1961. Precipitation has decreased by 31.2 mm/10a (p < 0.01), while the mean temperature has increased by 0.16 °C/10a (p < 0.01). The temperature change is not evident from 1961 to 1987 but rapid from 1988 to 2014 (0.38 °C/10a, p < 0.01). The precipitation decreased, but not highly significantly in all four seasons, whereby the maximum decrease occurred in autumn by a rate of 19.3 mm/10a (p < 0.01). The mean temperature has increased in all four seasons, with a maximum increase by 0.23 °C/10a (p < 0.01) in autumn. Those characteristics of climate change in the Xishui are similar to other places in southwestern China (Wang et al. 2015; Zhang et al. 2013a, b), which means that the Xishui station represents large-scale climate patterns. With the precipitation decreasing and temperature rising, this area is getting drier. The scPDSI values showed evidence of a significant trend of moisture decreasing after 2004, when the mean annual scPDSI value is −4.8, which is far below its long-term average (−0.83) (Table 1).

Variations of a annual mean temperature, b annual total precipitation, and c annual mean scPDSI over the period from 1961 to 2014. The dashed straight lines represent the temporal trends
Tree-ring data
The tree-ring samples were collected from old-growth F. hodginsii trees, at an altitude range from 1200 to 1270 m in the Sancha River scenic area (SCH), Dalou Mountains, northern Guizhou province (Fig. 1). The samples were collected from two sites, including both dead trees and living trees (Table 2). Dead trees (25 cores/12 trees) were obtained from a scenic spot locally known as Wangxiantai (SCH01), in which almost all mature F. hodginsii were dead (Li et al. 2013, 2015); a similar situation was observed in Tiantanggou (Li et al. 2015; Wang et al. 1984), another scenic site in this Nature Reserve. Only 10 trees out of 17 dead trees were precisely cross-dated by attributing calendar years to the outermost annual rings because of the rapid decay of dead trees in this warm and humid environment. The living trees (30 cores/17 trees) were obtained from SCH02 of a similar elevation and slope direction as SCH01, and no standing dead trees were found in this site. The two places are about 3 km apart. Two or three cores per tree were obtained by an increment borer at breast height (~1.3 m).
The tree-ring samples were processed following standard dendrochronological practices (Fritts 1976). All cores were air dried and glued onto wooden mounts with the transverse surface exposed and polished with progressively fining sandpapers until the cellular structure became clearly visible. All cores were visually cross-dated using the skeleton plot methodology (Fritts 1976) to assign an exact calendar year to each growth ring. Then, the ring widths were subsequently measured to a precision of 0.001 mm using a Velmex measuring system. The quality of the cross-dating was checked using the COFECHA program (Holmes 1983). Cores that were too short, profoundly corrupt, or not suitable for cross-dating due to uneven radial growth and frequent “wedging” of rings and “false rings” (Buckley et al. 2007; Sano et al. 2007, 2008) were excluded from further analysis. Because the samples from the two sites cross-dated well, we merged them into one chronology.
This chronology was developed from 40 cores from 21 trees. The ring-width series were standardized using ARSTAN (Cook 1985). Thirty-five series were de-trended with a negative exponential curve or a straight line (negative slope or horizontal), and the other remaining five series were fitted by a cubic smoothing spline of 67% of the series length (Cook and Kairiukstis 1990). The de-trended individual series were finally combined into a standard chronology (Fig. 4a) using a bi-weight robust mean methodology.

a Tree-ring-width standard chronology (solid line) and the corresponding sample size (dotted line); b the number of dead trees (black columns) with the years of death; the white star shows the number of missing rings with the exact timing; c 5-year moving average of annual precipitation (solid line) and annual mean temperature (dotted line)
Summary statistics for the STD chronology
Mean sensitivity (MS) is a measure for the change of ring width from 1 year to the next and thus of the high-frequency variance in a given tree-ring series (Speer 2010). A tree-ring series with a high MS is believed to contain more climate information than a tree-ring series with lower MS; the mean sensitivity in this site is 0.27 (Table 3). The chronology has a low percentage of absent ring (0.12%), which are mainly associated with extremely narrow ring in other trees. In general, tree-ring series in arid and other harsh environments have higher percentages of absent rings than series in humid and habitable environments. The first-order autocorrelation quantifies the dependence of the tree-ring widths from their directly preceding tree-ring width hence reflecting the influence of previous climate factors on the current ring growth. This tree-ring series has a fairly high first-order autocorrelation (0.74), which indicates that tree growth was influenced by the climate of the previous year and lag effects may exist in the time series (Wu 1990). The subsample signal strength (SSS) was used to determine the most reliable period within a chronology, with SSS above 0.85 as a desirable level (Wigley et al. 1984). This chronology has a high correlation within trees and a low correlation between trees, suggesting that tree growth may be influenced by a multiple of climate factors, different habitats, as well as other interferences.
Assessment of the relationship between tree growth and climate
Correlations between tree-ring chronology and the climatic variables (monthly temperature, total monthly precipitation and monthly scPDSI)—extending over a 20-month window from March of the previous year to October of the current year were examined via Pearson correlation (Briffa and Jones 1990). Given that the relationship between tree-ring chronology and climate factors may be influenced by the remarkable change of hydroclimate condition of study area in recent decades, a moving correlation analysis was applied to the chronology and the meteorological data to explore the possible temporal instability of the relationship. In addition, as the correlation coefficient was influenced by the linear tendency as discovered in another section below, we therefore used the first difference of the meteorological data and the tree-ring data.
Results
Correlation between the tree-ring chronology and climate variation
Figure 5a shows the Pearson correlations between the SCH chronology and monthly precipitation and temperature in Xishui station and the scPDSI grid point nearest to the study area. There were significant negative correlations (p < 0.05) with the mean temperatures in previous June–September and November, with previous September (−0.47, p < 0.01) being the strongest (Fig. 5a). In contrast, for monthly precipitation, no significant correlations existed with the tree-ring chronology. All monthly scPDSI from previous March to current October are positively correlated with the ring-width chronology, with the highest in previous October (0.6, p < 0.01).

a Correlation between the tree-ring chronology and monthly mean temperature, precipitation, and scPDSI (1961–2014) from March of the previous year to October of the current year. b The same as (a) but using first-difference records. Dotted and dashed horizontal lines indicate p = 0.05 and p = 0.01 significance levels, respectively
Despite evidence for strong correlations, the correlation coefficient may be influenced by the tendency of the linear variation of climate and chronology. This is why the correlation coefficient distinctly declined with scPDSI after applying the first-difference methodology. Accordingly, the chronology is significantly correlated with scPDSI only from previous July to current January (Fig. 5b). However, the correlation coefficients between tree-ring data and mean temperature do not change significantly and neither did the relationship with precipitation.
The “response divergence”
Before early 1990s, the 21-year running correlation calculated between chronology and climate facts (except temperature) were most of the time not significant and decreasing (Fig. 6). However, a different situation was found in the early 1990s that the correlation coefficients of the chronology with precipitation and scPDSI increased quickly and gradually reached the significance level, and the correlation coefficients of the chronology with temperature was consistently highly negative and suddenly increased. This huge change was coincidence with a tremendous difference in the variation of climate patterns that the temperature showed a decreasing trend while precipitation and scPDSI maintained higher than mean before the early 1990s, but a rapid increase while precipitation and scPDSI showed a decreasing trend in the early 1990s (Fig. 6).

Twenty-one-year running correlations between the chronology and the mean temperature (July–September of the previous year), precipitation (August–September of the previous year), and scPDSI (August–October of the previous year). For comparison, a 21-year running mean of the climate data (columns) was added. Dotted and dashed horizontal lines indicate the p = 0.05 and p = 0.01 significance levels, respectively
In order to quantify the temporal variation of tree-ring response to climate, we divided the meteorological data into two periods (1961–1987 and 1988–2014) (Fig. 7), taking the time of correlation coefficient change and the length of the two periods into consideration. In the 1961–1987 period, the chronology is positively correlated with mean temperature from January to April (r = 0.52; p = 0.05), while no persuasive relationship exists with monthly precipitation and scPDSI. This confirmed that the radial growth is limited by winter and spring temperature in this warm and humid climate, but their restriction on tree-ring growth is weak, therefore, the ring widths are wide and above average in this time period. In the 1988–2014 period, however, there is a highly negative correlation (p < 0.01) between tree-ring indices and previous July, August, and September temperature. The correlation between the chronology and precipitation is also noticeable in the late growing season of the previous year. The correlation between tree-ring chronology and previous August–September precipitation is 0.52 (p < 0.05), which is higher than the correlation (r = 0.45, p < 0.05) with current August–September monthly precipitation. These results suggest that moisture is the main limiting-factor for tree growth in the period from 1988 to 2014.

Correlation between tree-ring index and monthly mean temperature, precipitation, and scPDSI from March of the previous year to October of the current year. a Correlation over the 1961–1987 period. b Correlation over the 1988–2014 period. Dotted and dashed horizontal lines indicate the p = 0.05 and p = 0.01 significance levels, respectively
The above results suggest a clear “response divergence” phenomenon that shows up as limiting factor for tree growth is changing with time. From 1961 to 1987, the chronology is positively correlated with mean temperature in winter–spring, while no persuasive relationship existed with monthly precipitation. During this same time, the annual mean temperature decreased, with a modest decreasing rate in January–April (−0.13 °C/10a, p < 0.05). For annual precipitation, no obvious trend was observed during these year periods. Consequently, a relatively cold winter–spring restricted tree growth under the circumstance of plentiful precipitation, but this restriction is weaker if the climate is appropriate in general. The situation was notably different from 1988 to 2014, during which the chronology is negatively correlated with temperature and positively with precipitation. The correlation between the chronology and winter–spring temperature suddenly decreased with temperature rising after the early 1990s (Fig. 8). That is to say that the rapid increase of temperature by 0.43 °C/10a (p < 0.01) combined with a gradual decrease of precipitation may be the reason for the limiting factor shifting in the early 1990s and enhancing after the early 1990s.

Twenty-one-year running correlation between the chronology and the mean temperature (January–April of the current year). For comparison, a 21-year running mean of the climate data (columns) was added. Dotted and dashed horizontal lines indicate the p = 0.05 and p = 0.01 significance levels, respectively
The mortality of F. hodginsii
Meteorological data record that 2005, 2006, 2009, and 2011 were extreme drought years. We found a rapid reduction of radial growth from 2004 (narrowest in more than 100 years) coincided with recent drought and rapid warming (Fig. 4b) and tree mortality which concentrated from 2007 to 2013 occurred in extremely dry years or 1 to 3 years after a very dry summer or autumn. Following with sustained extreme drought during 2005 and 2006, most of the missing rings recorded by living younger trees in SCH02 occurred in 2007. Moreover, running correlation analysis (Fig. 5) shows high correlation coefficients of chronology with temperature, precipitation, and scPDSI, suggesting that the recent warming and decreasing precipitation may cause stress for F. hodginsii and thus may relate to the mortality and reduce growth of this species in this region (Fig. 5).
Reconstruction of the fate of selected tree individuals may help to understand the interdependence of growth decline and tree death. As an example, two neighboring F. hodginsii trees in the sampling site, one alive (young tree) and one dead (old tree), showed similar interannual growth variations over decades until the extremely dry and warm summer and autumn in 2007 (Fig. 9) when the young living tree had a missing ring and exhibited a sharp growth decline. The old tree died in 2007 after 10 years of very drastic reduction of radial growth from 1998 and the growth of the living tree also substantially reduced after 1998, but recovered afterwards, and this young tree still survives.

Ring-width chronology of a living Fokienia hodginsii tree and a tree that died in 2007. The samples were taken in November 2014
Discussion
It is generally acknowledged that the climate limitation on tree growth is weak in humid areas and low latitudes. But in this research, we found the response divergence and the response of tree growth to climate change is enhancing as climate is getting drought and warm. During 1961 to early 1990s, a relatively cold winter–spring restricted the tree growth under the circumstance of plentiful precipitation, since a warm winter–spring can protect trees from frost damaging and triggering the cambium to activate earlier, which may enhance growth (Shi et al. 2010, 2013; Yang 2012), but this restriction is relatively weaker as the climate is appropriate in general. However, after the early 1990s, the excessively high temperatures in late summer and early autumn during the previous year would reduce the water reserve in the soil, which may have enhanced the transpiration and respiration of trees, and therefore can constraint tree growth in the following year by reducing the carbohydrate storages that trees use for growth during the next year. This phenomenon was also found in some other tree-ring studies (Chen et al. 2012; Rolland 1993; Shang et al. 2011).
The reduced sensitivity of recent tree growth to temperature at Northern high latitudes may be due to the reduced limitation of temperature on tree growth under the circumstances of global warming (Briffa et al. 1998). In middle latitudes, the combined effects of recent warming and increasing intensity of climate extremes are enhancing synchrony of tree growth in Spain (Shestakova et al. 2015). Intense droughts and warming have led to decreasing tree growth and increasing mortality in southwestern USA, and tree growth will experience a substantial decrease during the twenty-first century if temperature and aridity rise (Williams et al. 2010). In general, there is still lack of direct evidence that growth–climate relationship has been changed in mid-high latitudes. However, we found that the association between climate and tree growth is changing as climate is getting warmer and drought. As a case study of the relationship between tree growth and climate under climate change in south China, this research could be a message that a large scale of tree-ring-climate patterns may be changing in humid areas of low latitudes. Unfortunately, it is unknown whether this regional tree-ring chronology which estimates response divergence can correlate with potentially more representative forest growth, as well as it is not clear whether this tree growth and climate patterns existed before since the instrumental records were too short. Consequently, more research is needed to confirm our observations.
The second objective of this study is to determine if drought or global warming were related to the drastic reduction of radial growth and tree mortality. Among endogenous processes, perhaps the best-known cause of increasing tree mortality rates is increasing competition resulting from increasing forest density and basal area. However, consistent with our observations of catastrophic mortality without compensating increases in recruitment, radial growth declined quickly during the study period. Thus, tree mortality is consistent with a quick decline of radial growth rather than an increase in potential for competition, which suggests that the catastrophic tree mortality cannot be attributed to forest dynamics. Furthermore, competition will be responsible for the gradual increase in tree mortality in general; however, consistent with recent drought and rapid warming, radial growth declined quickly since 2004 (narrowest in more than 100 years) and the mature dead F. hodginsii concentrated in a few years, and we did not find any missing rings and tree mortality before this time period. Thus, tree mortality is consistent with climate change rather than an increase in potential for competition.
Though not all samples from dead trees were collected since there are less F. hodginsii in this area, and is difficult to collect all of the outermost rings because of the rapid decay of dead trees in this warm and humid environment, it should generally represent the phase of recent tree death in this area. We observed that the trunks of mature F. hodginsii exposed to catastrophic tree mortality were tall and straight, and no dead young trees were found. It could be that big trees were more vulnerable to climate change, carbon starvation, as well as biotic diseases than smaller trees, as drought limits the plant leaf photosynthesis and carbon storage (McDowell et al. 2008; Phillips et al. 2010) and moisture determines the height of the tree (Niklas and Spatz 2004). Moreover, drought is more likely to hurt and kill mature F. hodginsii possibly (Fig. 9) because of its tall and straight trunk and less leaf features (Fensham and Fairfax 2007; McDowell et al. 2008). In addition, the occurence of missing rings provides compelling evidence for a strong sensitivity of the trees to global warming and local drought. Missing rings, i.e., the failure to produce stem wood in a particular year, represent an adaptive mechanism that may increase the survival rate of trees under extreme drought stress, when the trees have to reduce the respiratory demand and to decrease the risk of hydraulic failure (Liang et al. 2015). The occurrence of missing rings indicates that the environmental conditions are close to the survival limit of the trees. Thus, the crop up of missing rings may serve as an excellent indicator of the severity of drought stress effects on tree growth in this humid region of low latitudes. We suggest that the presence of recent drought and temperature stress thresholds beyond what F. hodginsii could suffer may operate as a trigger that caused the reduction of basal area increment and may ultimately result in catastrophic tree mortality. The implication is that if warming continues and effective moisture continue to increasingly become less available, populations of these species may become increasingly sensitive to drought and at risk for mortality. The above suggestions is supported by many studies. For example, Liang et al. (2015) suggested that recent climate warming has increased missing ring frequency and tree mortality rates of Qinghai spruce in north-eastern Tibetan plateau. Anderegg et al. (2012) provides insight into the physiological mechanisms of drought on tree death with modeling approaches. Van Mantgem et al. (2009) inferred that regional warming and consequent increases in water deficits may be the dominant contributor of the increases in tree mortality rates. Pedersen (1998) noted that the decreases in basal area growth and increases in tree mortality of red oak in America followed the drought event. However, the patterns and causes of tree mortality typically are complex, thus, it is hard to come to a conclusion at this point in time of whether climate change itself or other factors were responsible for tree mortality. More work should be done to test our hypotheses in the future.
Conclusion
We present the first F. hodginsii tree-ring-width chronology in the northern Yunnan-Guizhou Plateau. The response of tree-ring width to climate change and the impact of climate change on forest ecosystem were studied. This study is an important supplement to tree-ring research in the subtropical humid areas in China. We found a clear response divergence phenomenon that the limiting factors for tree growth changed under different hydroclimate conditions. The catastrophic tree mortality at the edge of F. hodginsii’s natural distribution informs us that special attention should be paid to the ecological impact of global warming and extreme climate events, especially at the border area of tree species.
References
Alexander LV, Zhang X, Peterson TC, Caesar J, Gleason B, Klein Tank AMG, Haylock M, Collins D, Trewin B, Rahimzadeh F, Tagipour A, Rupa Kumar K, Revadekar J, Griffiths G, Vincent L, Stephenson DB, Burn J, Aguilar E, Brunet M, Taylor M, New M, Zhai P, Rusticucci M, Vazquez-Aguirre JL (2006) Global observed changes in daily climate extremes of temperature and precipitation. J Geophys Res 111:1042–1063
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzalez P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J-H, Allard G, Running SW, Semerci A, Cobb N (2010) A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manag 259:660–684
An W, Liu X, Leavitt SW, Ren J, Sun W, Wang W, Wang Y, Xu G, Chen T, Qin D (2012) Specific climatic signals recorded in earlywood and latewood δ18O of tree rings in southwestern China. Tellus B 64:18703
Anderegg WRL, Berry JA, Field CB (2012) Linking definitions, mechanisms, and modeling of drought-induced tree death. Trends Plant Sci 17:693–700
Anderegg WRL, Flint A, Huang C-Y, Flint L, Berry JA, Davis FW, Sperry JS, Field CB (2015) Tree mortality predicted from drought-induced vascular damage. Nat Geosci 8:367–371
Briffa KR, Jones PD (1990) Basic chronology statistics and assessment. In: Cook ER, Kairiukstic LA (eds) Methods of dendrochronology. Kulwer Academic Publishers, Boston, MA, pp 137–152
Briffa KR, Schweingruber FH, Jones PD, Osborn TJ, Shiyatov SG, Vaganov EA (1998) Reduced sensitivity of recent tree-growth to temperature at high northern latitudes. Nature 391:678–682
Buckley BM, Palakit K, Duangsathaporn K, Sanguantham P, Prasomsin P (2007) Decadal scale droughts over northwestern Thailand over the past 448 years: links to the tropical Pacific and Indian Ocean sectors. Clim Dyn 29:63–71
Carrer M, Urbinati C (2006) Long-term change in the sensitivity of tree-ring growth to climate forcing in Larix decidua. New Phytol 170:861–871
Chen F, Yuan YJ, Wei WS, Yu SL, Zhang TW (2012) Reconstructed temperature for Yong’an, Fujian, Southeast China: linkages to the Pacific Ocean climate variability. Glob Planet Chang 86-87:11–19
Cook, ER., 1985. A time series analysis approach to tree-ring standardization. PhD dissertation, University of Arizona, Tucson, 1–171
Cook ER, Kairiukstis LA (1990) Methods of dendrochronology: applications in the environmental sciences, vol 414. Kluwer Academic Publishers, Dordrecht, Netherlands
D’Arrigo R, Wilson R, Liepert B, Cherubini P (2008) On the ‘divergence problem’ in northern forests: a review of the tree-ring evidence and possible causes. Glob Planet Chang 60:289–305
Duan J, Zhang Q-B, Lv L-X (2013) Increased variability in cold-season temperature since the 1930s in subtropical China. J Clim 26:4749–4757
Easterling DR, Evans JL, Groisman PY, Karl TR, Kunkel KE, Ambenje P (2000) Observed variability and trends in extreme climate events: a brief review. Bull Am Meteorol Soc 81:417–425
Esper J, Frank D (2009) Divergence pitfalls in tree-ring research. Clim Chang 94:261–266
Fang K, Gou X, Chen F, Li J, D’Arrigo R, Cook E, Yang T, Liu W, Zhang F (2010b) Tree growth and time-varying climate response along altitudinal transects in central China. Eur J For Res 129:1181–1189
Fang KY, Gou XH, Chen FH, Li JB, D’Arrigo R, Cook E, Yang T, Davi N (2010a) Reconstructed droughts for the southeastern Tibetan plateau over the past 568 years and its linkages to the Pacific and Atlantic Ocean climate variability. Clim Dyn 35:577–585
Fensham RJ, Fairfax RJ (2007) Drought-related tree death of savanna eucalypts: species susceptibility, soil conditions and root architecture. J Veg Sci 18:71–80
Fensham RJ, Holman JE (1999) Temporal and spatial patterns in drought-related tree dieback in Australian savanna. J Appl Ecol 36:1035–1050
Franklin JF, Shugart HH, Harmon ME (1987) Tree death as an ecological process. Bioscience 37:550–556
Fritts HC (1976) Tree rings and climate. Q Rev Biol 52:226–567
Gao L, Gou X, Deng Y, Yang M, Zhang F, Li J (2015) Dendroclimatic reconstruction of temperature in the eastern Qilian Mountains, northwestern China. Clim Res 62:241–250
Gao L, Gou X, Deng Y, Zhang F, Fang K (2011) An overview of the divergence phenomenon in dendroclimatology (in Chinese abstract review). J Glaciol Geocryol 33:454–460
Gou X, Deng Y, Gao L, Chen F, Cook E, Yang M, Zhang F (2014) Millennium tree-ring reconstruction of drought variability in the eastern Qilian Mountains, northwest China. Clim Dyn 45:1761–1770
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 44:51–67
IPCC (2013) Summary for policymakers of climate change 2013: the physical science basis. Contribution of working group I to the fifth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge
Jacoby GC, D'Arrigo RD (1995) Tree ring width and density evidence of climatic and potential forest change in Alaska. Glob Biogeochem Cycles 9:227–234
Jiao L, Jiang Y, Zhang WT, Wang MC, Zhang LN, Zhao SD (2015) Divergent responses to climate factors in the radial growth of Larix sibirica in the eastern Tianshan Mountains, northwest China. Trees 29:1673–1686
Li M, Deng L, Jiang Y, Yang C, Mu J (2013) Community composition and structure of Fokienia hodginsii community in Xishui Nature Reserve in Guizhou (in Chinese). J Northwest Forestry University 28:46–50
Li M, Deng L, Pan D, Mu J, Li C, Yang C (2015) Study on growth process of Fokienia hodginsii community in Xishui Nature Reserve in Guizhou (in Chinese). J Anhui Agriculture:199–202
Li Y, Fang K, Cao C, Li D, Zhou F (2016) A tree-ring chronology spanning 210 years in the coastal area of southeastern China, and its relationship with climate change. Clim Res 67:209–220
Liang E, Eckstein D, Liu H (2009) Assessing the recent grassland greening trend in a long-term context based on tree-ring analysis: a case study in North China. Ecol Indic 9:1280–1283
Liang E, Leuschner C, Dulamsuren C, Wagner B, Hauck M (2015) Global warming-related tree growth decline and mortality on the north-eastern Tibetan plateau. Clim Chang 134:163–176
Liang E, Shao X, Qin N (2008) Tree-ring based summer temperature reconstruction for the source region of the Yangtze River on the Tibetan plateau. Glob Planet Chang 61:313–320
McDowell N, Pockman WT, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, Yepez EA (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? The New Phytologist 178:719–739
Min S-K, Son S-W, Seo K-H, Kug J-S, An S-I, Choi Y-S, Jeong J-H, Kim B-M, Kim J-W, Kim Y-H, Lee J-Y, Lee M-I (2015) Changes in weather and climate extremes over Korea and possible causes: a review. Asia-Pac J Atmos Sci 51:103–121
Niklas KJ, Spatz HC (2004) Growth and hydraulic (not mechanical) constraints govern the scaling of tree height and mass. Proc Natl Acad Sci U S A 101:15661–15663
Pedersen BS (1998) The role of stress in the mortality of midwestern oaks as indicated by growth prior to death. Ecology 79:79–93
Peng CH, Ma ZH, Lei XD, Zhu Q, Chen H, Wang WF, Liu SR, Li WZ, Fang XQ, Zhou XL (2011) A drought-induced pervasive increase in tree mortality across Canada’s boreal forests. Nat Clim Chang 1:467–471
Phillips OL, van der Heijden G, Lewis SL, Lopez-Gonzalez G, Aragao L, Lloyd J, Malhi Y, Monteagudo A, Almeida S, Davila EA, Amaral I, Andelman S, Andrade A, Arroyo L, Aymard G, Baker TR, Blanc L, Bonal D, de Oliveira ACA, Chao KJ, Cardozo ND, da Costa L, Feldpausch TR, Fisher JB, Fyllas NM, Freitas MA, Galbraith D, Gloor E, Higuchi N, Honorio E, Jimenez E, Keeling H, Killeen TJ, Lovett JC, Meir P, Mendoza C, Morel A, Vargas PN, Patino S, Peh KSH, Cruz AP, Prieto A, Quesada CA, Ramirez F, Ramirez H, Rudas A, Salamao R, Schwarz M, Silva J, Silveira M, Slik JWF, Sonke B, Thomas AS, Stropp J, Taplin JRD, Vasquez R, Vilanova E (2010) Drought-mortality relationships for tropical forests. New Phytol 187:631–646
Qian J, Lu J, Qipu T, Suming W (2002) Reconstruction of the climate in the Tianmu Mountain area, Zhejiang Province, in the last 160 years by delta C-13 sequence of tree ring alpha-cellulose. Sci China Ser D Earth Sci 45:409–419
Rolland C (1993) Tree-ring and climate relationships for Abies alba in the Internal Alps. Tree-Ring Bull 53:1–11
Sano M, Buckley BM, Sweda T (2008) Tree-ring based hydroclimate reconstruction over northern Vietnam from Fokienia hodginsii: eighteenth century mega-drought and tropical Pacific influence. Clim Dyn 33:331–340
Sano M, Tinh PH, Sweda T (2007) Dendroclimatic potential of Fokienia trees in the Indochina peninsula. J Japanese Forest Soc 89:144–148
Schneider L, Esper J, Timonen M, Büntgen U (2014) Detection and evaluation of an early divergence problem in northern Fennoscandian tree-ring data. Oikos 123:559–566
Shang H, Wei W, Yuan Y, Yu S, Zhang T (2011) Variations of temperature during last 350 years at Xinyuan recorded by tree-ring (in Chinese). J Arid Land Resources and Environ 25:187–190
Shestakova TA, Gutiérrez E, Kirdyanov AV, Camarero JJ, Génova M, Knorre AA, Carlos J, Dios VRD, Sánchez-Salguero R, Voltas J (2015) Forests synchronize their growth in contrasting Eurasian regions in response to climate warming. Proc Natl Acad Sci 113
Shi J, Cook ER, Li J, Lu H (2013) Unprecedented January–July warming recorded in a 178-year tree-ring width chronology in the Dabie Mountains, southeastern China. Palaeogeogr Palaeoclimatol Palaeoecol 381-382:92–97
Shi J, Cook ER, Lu H, Li J, Wright WE, Li S (2010) Tree-ring based winter temperature reconstruction for the lower reaches of the Yangtze River in southeast China. Clim Res 41:169–176
Shi J, Lu H, Li J, Shi S, Wu S, Hou X, Li L (2015) Tree-ring based February–April precipitation reconstruction for the lower reaches of the Yangtze River, southeastern China. Glob Planet Chang 131:82–88
Slik JWF (2004) El Nino droughts and their effects on tree species composition and diversity in tropical rain forests. Oecologia 141:114–120
Speer J (2010) Fundamentals of tree-ring research. The University of Arizona Press, Tucson
Suresh HS, Dattaraja HS, Sukumar R (2010) Relationship between annual rainfall and tree mortality in a tropical dry forest: results of a 19-year study at Mudumalai, southern India. For Ecol Manag 259:762–769
van Mantgem PJ, Stephenson NL, Byrne JC, Daniels LD, Franklin JF, Fule PZ, Harmon ME, Larson AJ, Smith JM, Taylor AH, Veblen TT (2009) Widespread increase of tree mortality rates in the western United States. Science 323:521–524
Walther GR, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:389–395
Wang A, Liu Y, Liu S, Yang T (1984) A preliminary report on the geographic distribution of nature forest Fokienia hodginsii in China (in Chinese). Chinese J Ecol:19–23
Wang L, Chen W, Zhou W, Huang G (2015) Teleconnected influence of tropical Northwest Pacific sea surface temperature on interannual variability of autumn precipitation in Southwest China. Clim Dyn 45:2527–2539
Wells N, Goddard S, Hayes MJ (2004) A self-calibrating palmer drought severity index. J Clim 17:2335–2351
Wigley TML, Briffa KR, Jones PD (1984) On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Clim Appl Meteorol 23:201–213
Wu, X., 1990. Tree rings and climate change (in Chinese). China Meteorological Press
Williams AP, Allen CD, Millar CI, Swetnam TW, Michaelsen J, Still CJ, Leavitt SW (2010) Forest responses to increasing aridity and warmth in the southwestern United States. Proc Natl Acad Sci U S A 107:21289–21294
Yang B (2012) Spatial and temporal patterns of climate variations over the Tibetan plateau during the period 1300-2010 (in Chinese). Quaternary Sciences 32:81–94
Yang B, Qin C, Wang J, He M, Melvin TM, Osborn TJ, Briffa KR (2014) A 3,500-year tree-ring record of annual precipitation on the northeastern Tibetan plateau. Proc Natl Acad Sci U S A 111:2903–2908
Yu D, Liu J, Benard J, Zhou L, Zhou W, Fang X, Wei Y, Jiang S, Dai L (2013) Spatial variation and temporal instability in the climate–growth relationship of Korean pine in the Changbai Mountain region of Northeast China. For Ecol Manag 300:96–105
Zhang Y, Martin W, Gou X (2009) Changing relationships between tree growth and climate in Northwest China. Forest Ecol 201:39–50
Zhang M, He J, Wang B, Wang S, Li S, Liu W, Ma X (2013a) Extreme drought changes in Southwest China from 1960 to 2009. J Geogr Sci 23:3–16
Zhang W, Jin F-F, Zhao J-X, Qi L, Ren H-L (2013b) The possible influence of a nonconventional El Niño on the severe autumn drought of 2009 in Southwest China. J Clim 26:8392–8405
Zhang QB, Evans MN, Lyu L (2015) Moisture dipole over the Tibetan plateau during the past five and a half centuries. Nat Commun 6:8062
Acknowledgments
The authors would like to thank Mr. Chen Minghong (officer of the Forestry Bureau of Xishui country) for his kind help in the field. We also thank Fang Yang from New York University for her valuable help to improve the manuscript. This research is supported by the National Natural Science Foundation of China (No. 41475067 and No. 41401047), the Doctoral Fund of Ministry of Education of China (20120211110041), and the Fundamental Research Funds for the Central Universities (No. lzujbky-2016-163).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Su, J., Gou, X., Deng, Y. et al. Tree growth response of Fokienia hodginsii to recent climate warming and drought in southwest China. Int J Biometeorol 61, 2085–2096 (2017). https://doi.org/10.1007/s00484-017-1409-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00484-017-1409-y