
Abstract
Key message
Our study on tree bark thickness across the major forest types of Central Himalayas indicates that the higher relative bark thickness at an early age is a key fire adaptational feature.
Abstract
In this study, we have primarily examined the relationship between fire incidence, bark thickness, relative bark thickness and related characters of the dominant tree species across the major forest type of Central Himalayas (29° 20′–29° 23′ N latitude and 79° 23′–79° 30′ E longitude) differing in fire incidences. The forest fires are human-ignited, small, and patchy surface fires and their incidences vary considerably across the forest types: 472.4 (fires/year) in Pinus roxburghii forests (between 1000 and 2000 m); 50.2 (fires/year) in Shorea robusta forests in the foothills; 7.4 (fires/year) in Quercus leucotrichophora forests (1200–2700 m); and rare in P. wallichiana forests (> 1800 m). With regard to bark traits, we focus on absolute bark thickness, and relative bark thickness. Our analysis of bark-related traits showed that bark thickness and bark development at an early age are directly correlated with fire exposure, indicating the role of fire in the species dominance. With increasing diameter of trees, bark thickness increases but relative bark thickness decreases in all tree species, indicating that allocation to bark decreases as trees become big hence less vulnerable to fire. The relative bark thickness declines more with tree size in species exposed more to fires than those less expose to fire. Our analysis has contributed to highlighting the bark thickness as a functional feature, with implication for community composition.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Bark as an outermost cover is of key importance in tree adaptation to adverse environmental conditions (Ferrenberg and Mitton 2014). Frequent fires are supposed to have selected for several fire-adaptive traits, such as bark thickness in many species, serotiny in Pinus spp. (Barden 1978), and re-sprouting in Quercus and several other species (Little 1974). A thick bark of stems is considered as the main protection against under-canopy or ground surface fires (Lawes et al. 2013; Pausas 2015).
Variation in bark thickness across species is overlain on a positive relationship between stem diameter and bark thickness (Paine et al. 2010; Poorter et al. 2014). The rate of bark accumulation will depend not only on the relative investment in bark, but also on the rate of stem growth (Hoffmann et al. 2012). Ecologists measure several bark characters, such as bark density, moisture content and thermal conductivity (Uhl and Kauffman 1990; Cornelissen et al. 2003). Here, we have analyzed how dominant tree species of Central Himalayan forests varying in fire incidences differ in bark thickness, relative bark thickness, and other related characters.
Although most tree species of fire-prone habitats are characterized by thick bark, it is not clear whether they invest relatively more in bark on the portions of their stems that experience the highest fire intensities. Because fire regime varies world-wide (Pausas and Keeley 2009; Pausas and Ribeiro 2013), bark thickness should also vary across ecosystems in a predictable manner.
The stand replacing infrequent but severe fires are most common in moist coniferous forests, such as Cascade western hemlock, and Pacific silver fir forests (Agee 1993). A wide variety of fire-dependent pine forests and pine savannas across Central America and the Caribbean, and extensive open pine forests and woodlands in the tropical and subtropical environments of Southeast and South Asia face frequent fires; here fires act as a key ecosystem driver in maintaining stand characteristics (Troup 1921; Singh and Singh 1992; Kraus and Goldammer 2007; Singh et al. 2016). Surface fires, such as those in many savannas, damage trees principally near the ground due to low flame heights, long fire residence times, and high temperatures (Franklin et al. 1997). Most comparative studies on bark thickness have been conducted in savannas and open dry forests, where the effects of fire are likely to overwhelm any other factors that affect bark thickness (e.g., Hoffmann et al. 2003). As fire frequency and intensity can select for higher investment in bark thickness (Lawes et al. 2013; Pausas 2015), it can be hypothesized that bark thickness is directly related to fire incidences. As fire frequency increases, relatively thicker bark and a greater rate of bark accumulation should be beneficial for protecting the vascular cambium during fire (Lawes et al. 2011).
In Himalayas, forests can grow up to over 4000 m elevation, encompassing forest types varying from tropical rainforest to dry alpine scrubs (Singh and Singh 1992). Frequent human-made low-intensity fires are an integral part of chir pine (Pinus roxburghii)—banj oak (Quercus leucotrichophora) forest belt (generally, between 800 and 2000 m altitude) in Uttarakhand (Singh and Singh 1986, 1992; Singh et al. 2016). There are evidences indicating that fires have promoted the regional domination of chir pine at the expense of broadleaf oak forests (Singh and Singh 1986, 1992; Singh et al. 2016).
Human-ignited fires have been shaping the Central Himalayan forests from foothills to 1800 m elevation or more for thousands of years, modifying their structural and compositional features (Singh and Singh 1986). To the best of our knowledge, no exercise between incidence of forest fire and bark characters of dominant forest species has been done before for this part of the world.
The objectives of this study were: (1) to examine the relationship between exposure to surface forest fires and bark thickness of dominant forest tree species of the central Himalayas, and (2) to analyze variation in absolute and relative bark thickness in relation to diameter/age of trees from the forests. Since fire has been an integral evolutionary agent for millions of years in Himalayas (Parashar and Biswas 2003), we hypothesize parameters of bark thickness in stems of dominant species and fire frequency would be correlated. By examining the dominant tree species of Central Himalayan region (two pines and two broadleaved species) of forests with a closed canopy, we expect that a more balanced understanding of bark’s functions would be obtained in relation to forest fires. As frequent fires can impose a substantial selection pressure on bark thickness (Stephens and Libby 2006), species whose range includes frequently burned habitats would be expected to have thicker bark than those whose range includes only habitats in which fires are infrequent.
Materials and methods
Study species and region
The study species from foothills to about 2000 m elevation were sal (Shorea robusta Gaertn.), chir pine (P. roxburghii Sarg.), banj oak (Q. leucotrichophora A. Camus), and blue pine (P. wallichiana A. B. Jacks) (Table 1), which dominate in 12.1%, 15.3%, 14.8%, and 0.7% forest area, respectively, in Uttarakhand (Uttarakhand Forest Statistics 2018). All study species are tall (> 25 m) canopy-forming forest species. The banj oak and chir pine (occurring between 800 and 2000 m) are among the top 10 forest species of the Indian subcontinent in terms of growing stock (FSI 2019). Shorea robusta (from 300 to 1200 m), the most important species of India in terms of growing stock (contributing 10.6% to total growing stock in country’s forests, FSI (2019)) forms forests in the foothills along most of the Himalayan Arc. Pinus wallichiana is generally distributed between 1800 and 4300 m in Himalayas (Troup 1921; Champion and Seth 1968; Singh and Singh 1992). Among the study species, P. roxburghii and S. robusta are regarded as fire-tolerant species, and P. wallichiana and Q. leucotrichophora as vulnerable to fire (Troup 1921; Singh and Singh 1992).
The present study area extends from foothills (200–300 m) to 2300 m in the Nainital catchment (29° 20′ 11′′ to 29° 23′ 25.8′′ N latitude and 79° 23′ 15.8′′ to 79° 30′ 50.2′′ E longitude) of Kumaun region, Uttarakhand. The rainfall pattern of the region is characterized by a typical monsoon season (mid-June to mid-September), which accounts for about three-fourths of the annual rainfall. The average annual precipitation in Nainital was 1271 mm (for 2004–2016) of which 347 mm was during pre-monsoon months (March–May) when fires generally occur (https://power.larc.nasa.gov/data-access-viewer/). The occasional and sporadic pre-monsoon rainstorms largely influence the fire intensity.
Fire regime
Human-ignited fires during pre-monsoon (March to mid-June) drought have been almost an annual feature in much of the western and central Himalayas, below 2000 m during last two centuries or so (Troup 1921; Singh and Singh 1992; Singh et al. 2016). They are small, and patchy surface fires, flames seldom damaging tree crowns at a stand scale. On an average, 1858 ha of the total forest area in Uttarakhand was burnt annually during 2007–2017, and the area affected per fire incidence was less than 30 ha, in about 90% cases, it was below 10 ha, and some affected only a fraction of hectare. About 65% fire incidences occurred below 1000 m elevation and 85% below 2000 m. During 2013–2017, the fire density was highest in P. roxburghii forests (599 fires/1000 km2), followed by S. robusta forests (80 fires/1000 km2) and Q. leucotrichphora forests (10 fires/1000 km2). Fire was rarely observed in areas under P. wallichiana. During the five study years, P. roxburghii and S. robusta forests experienced forest fires annually, whereas in Q. leucotrichophora forests, the recurrence interval is of 3–4 years.
The high fire incidences in P. roxburghii forest is associated with heavy ground fuel loads along with long needles rich in resin. The loose litter of long needle is more flammable because it holds more air. As per the State Forest Department records, monthly mean fire incidence (across the study years) followed a unimodal curve with a peak in May (381). The total fire affected area per month increased from March (28.60 ha) to May (737.91 ha) and then decreased in June with the arrival of monsoon (227.07 ha). The monthly average area burned per fire incident ranged between 1.56 ha and 7.25 ha. Prescribed burning is used by the forest managers to promote even-aged young stands of P. roxburghii and S. robusta at the expense of other tree species (Troup 1921; Singh and Singh 1992).
Methods
For each study species, trees of the entire stem diameter range available were sampled for bark in a representative forest stand. We sampled 125 trees of P. roxburghii, 108 of S. robusta, 76 of P. wallichiana and, 33 of Q. leucotrichophora, which were distributed across a wide diameter levels.
The data on forest fires which pertained to the date and site of fire incidence, and area affected by fire were collected from the Uttarakhand State Forest Department.
Bark thickness was measured using Swedish Bark Gauge. Where bark was fissured, we measured thickness from the highest point. We took four evenly spaced measures of bark thickness around the circumference of the stem at breast height (1.37 m) per individual. The diameter at breast height of each tree was also measured. For every sampled tree, tree height, clean bole height, and the maximum char height were measured by Ravi Altimeter. Char height is an assumed proxy for flame height, which corresponds to fire line intensity (Rothermel and Deeming 1980).
For each species, we used linear regression to analyze the relationship between stem diameter and bark thickness, and stem diameter and relative bark thickness. Relative bark thickness was calculated for each individual as the ratio of bark thickness to diameter. By assuming that the cost of thick bark is a function of the cross-sectional area of bark, the relative costs of bark growth among individuals with different relative bark thickness were calculated by:
where BA, cross-sectional area of the bark (cm2); r, radius of the stem (cm); BT, bark thickness (cm).
According to Dickinson and Johnson (2001), fire kills a stem by damaging cambial tissue by conductive heat transfer and bark protects the tree stem from fire and prevents cambial necrosis (Lawes et al. 2011). The bark insulating ability is given by the critical time to cambium kill which was computed from bark thickness. Time to kill the cambium is directly proportional to the bark thickness squared (Peterson and Ryan 1986; Lawes et al. 2011). We use the simplified formula of Peterson and Ryan (1986) based on a fire with a constant temperature and α = 0.060 cm2 min−1 (α is the bark thermal diffusivity):
where \({\tau }_{C}\), the critical time (minutes) for cambium kill; BT, bark thickness (cm).
Polynomial regression was conducted to test the relationship between cross-sectional bark area and stem diameter; \({\tau }_{C}\) and bark thickness; char height and stem diameter. Differences among species in bark thickness, relative bark thickness, and the time required to kill the cambium were analyzed with one-way ANOVA with species as a fixed factor. A quadratic model was also developed to test the relationship between annual fire incidents and maximum bark thickness of each study tree species.
Results
Bark thickness
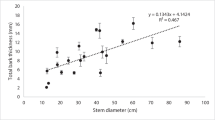
The stem bark thickness was positively correlated with stem diameter and varied greatly within and among species. Pinus roxburghii, S. robusta, Q. leucotrichophora and P. wallichiana showed ranges of stem bark thickness from 1.06 to 3.33 cm, 1.15 to 2.95 cm, 0.30 to 2.5 cm and 0.25 to 1.85 cm, respectively. The stem diameter and bark thickness were significantly positively correlated for both: P. roxburghii (R2 = 0.73; p < 0.01) and P. wallichiana (R2 = 0.57; p < 0.01) (Fig. 1), but at early age, bark thickness was much less in the latter. At about 60 cm stem girth, the bark thickness was 1.08 cm in P. wallichiana and 3.15 cm in P. roxburghiii. There was also a positive trend of increasing tree size (DBH) and bark thickness in the broadleaved species viz. S. robusta (R2 = 0.19; p < 0.01) and Q. leucotrichophora (R2 = 0.54; p < 0.01) (Fig. 1). Pinus roxburghii had about 20% thicker bark than S. robusta and 43.2% than Q. leucotrichophora. For individuals < 25 cm DBH, which are particularly susceptible to top kill during fire, bark thickness ranked as following: S. robusta (1.74 cm) > P. roxburghii (1.59 cm) > Q. leucotrichophora (0.75 cm) > P. wallichiana (0.27 cm).

Relationship between bark thickness (cm) and stem diameter (cm) for Himalayan tree species. The slopes of the regression line are given in the parentheses for each species
The benefit of relatively thicker bark, in terms of protecting the vascular cambium and epicormic buds from heating is achieved at smaller stem diameters (> 10 cm diameter) for P. roxburghii and S. robusta trees than for Q. leucotrichophora and P. wallichiana (> 30 cm diameter). Relative bark thickness was significantly different among species. On an average, relative bark thickness was higher for S. robusta (13.45%) and P. roxburghii (12.05%) than for Q. leucotrichophora, 8.50% and P. wallichiana, 4.35% (Fig. 2). Relative bark thickness declined with tree size for all species (Fig. 3). In all species, bark thickness and relative bark thickness were significantly different (F3,128 = 62.19, p < 0.01; F3,128 = 40.76, p < 0.01, respectively). The cross-sectional bark area was significantly correlated with stem diameter for all the species: P. roxburghii (R2 = 0.96; p < 0.01), S. robusta (R2 = 0.88; p < 0.01), Q. leucotrichophora (R2 = 0.79; p < 0.01), and P. wallichiana (R2 = 0.78; p < 0.01) (Fig. 3).

Boxplots of relative bark thickness (%) for studied tree species

Relationship of cross-sectional bark area (cm2) and relative bark thickness (%) with respect to stem diameter (cm) for (a) P. roxburghii, (b) S. robusta, (c) Q. leucotrichophora and (d) P. wallichiana
The critical time (\({{\varvec{\tau}}}_{{\varvec{C}}}\)) required to kill the cambium
There was a significant difference among the species in \({\tau }_{C}\) (F3,128 = 51.28, p < 0.01)(Fig. 4).The mean time required to kill the cambium by burning a bark > 1.5 cm thickness, which is sufficient to insulate the cambium from low-intensity surface fires (van Mantgem and Schwartz 2003) was highest (~ 15 min) for P. roxburghii individuals, followed by S. robusta (~ 13 min). It was approximately 10 min for both Q. leucotrichophora and P. wallichiana to transfer heat from the bark surface (with > 1.5 cm thickness) to the cambium. For individuals < 25 cm DBH, S. robusta with an average bark thickness of 1.74 cm would require nine minutes to kill the cambium. It was high (7.5 min) also for P. roxburghii and low (2.5 min) and very low (0.2 min) for, Q. leucotrichophora, and P. wallichiana, respectively.

Relationship between critical time required to kill the cambium (minutes) and bark thickness (cm) of a tree for a P. roxburghii, b S. robusta, c Q. leucotrichophora and d P. wallichiana
Char height and tree mortality
Char height on individual boles was explored as predictors of stem mortality. Even having scars up to an average height of 1.4 m, the thin-barked species viz. Q. leucotrichophora and P. wallichiana survived during fire. Among the P. roxburghii trees, bark char does not exceed 2 m and no crown damage was found.
Discussion
We examined both absolute bark thickness and relative bark thickness across four trees species growing in forests differing in fire exposure. Our study on bark thickness of the four dominant tree species of Central Himalayan forests differing in under-canopy fire incidences shows that absolute bark thickness was greater in the species (P. roxburghii and S. robusta) of frequently burnt forests than in those (Q. leucotrichophora and P. wallichiana) subjected to infrequently burnt forests (Fig. 5). Pellegrini et al. (2017) have shown that in savannas (which are fire-driven ecosystems), species have thicker bark than forest species. The significantly positive correlation between bark thickness of Central Himalayan tree species and fire incidences (R2 = 0.65, p < 0.05) (Fig. 5), is an evidence that defense against fire is one of the primary factors shaping bark thickness (Uhl and Kauffman 1990; Van Nieuwstadt and Sheil 2005). The bark of P. roxburghii and S. robusta was about 1.5 cm thick right from sapling stage, which is enough to protect its cambium from surface fires of three minutes when temperature reaches 400 °C (He et al. 2012). Pines are known to show a wide range of traits associated with different fire regimes (He et al. 2012; Keeley 2012). The pine species with basal thick bark mostly live in surface-fire ecosystems (Jackson et al. 1999; Fig. 5). Similar to pines, there are other conifers that live in surface-fire affected ecosystems and have a very thick bark, grow very tall and tend to self-prune the lower branches. Prominent examples are Araucaria araucana and Fitzroya cupressoides in the Andes, and the giant sequoia (Sequoiadendron giganteum) and Calocedrus decurrens in western North America (Beaty and Taylor 2007; Gonzalez et al. 2010).

Relationship between maximum bark thickness (cm) and annual fire incidences in study forest types
Fires are so frequent in some areas of P. roxburghii forests that almost all under-canopy trees and shrubs are exterminated and only fire-adapted grasses grow. The thin-barked under-canopy tree species, such as Myrica esculenta (0.48 cm), and Litsea umbrosa (0.55 cm), are first to disappear. Rhododendron arboreum (1.23 cm) with relatively thicker bark may persist but not for long. In oak savannas, bark thickness is shown to be correlated with the regime of surface fires (Cavender-Bares et al. 2004). In Central Himalayas, oaks form forests with a close canopy, crown density generally in the range of 50–80% (Singh and Singh 1987). In them, the young oak trees have thinner bark, hence fail to regenerate in areas around pine forests where fires are common. However, being efficient coppicers, Himalayan oaks survive when fire is less frequent (Ralhan et al. 1985). A fire regime characterized by frequent surface fires for at least two centuries is likely to create more opportunities for establishment of invasive species (Hellmann et al. 2008). In repeatedly burnt forests of the present study area Lantana camara, Anaphalis spp., Eupatorium odoratum, and Imperata cylindrica are common invasive species which profusely proliferate after forest fires (Negi 2019; Singh et al. unpubl.).
In all the four species of our study, bark thickness increased with stem diameter, but it increased more in species living in forests with more frequent fires (Fig. 1). Contrary to this, the relative bark thickness which indicates proportional allocation to bark declined with increase in tree diameter (and age) in all study species (Fig. 3). Allocation to bark declined in the study species, enabling stems to grow bigger when they are away from the reach of the fire. The decrease in the relative bark thickness was more in fire-exposed species than those occurring in infrequently burnt forests. Developing a thick stem bark has a cost both in terms of resources and opportunity as a thick bark limits the diffusion of water and carbon dioxide and other through the stem (Pausas 2015).
Several studies support the finding that the duration of heating required to kill the cambium of a tree is directly proportional to bark thickness squared (Uhl and Kauffman 1990; van Mantgem and Schwartz 2003). For the present Himalayan study species, correlation between bark thickness and the duration of heat required to kill the cambium was significant. In Himalayas, chir pine forests burn more frequently than adjacent broadleaved forests, so higher relative bark thickness should be beneficial for the survival of young trees. Our study indicates that when young (< 25 cm tree diameter) the relative bark thickness in S. robusta and P. roxburghii was similar, and 37% more than that of Q. leucotrichophora and 67% more than that of P. wallichiana. Several studies (Hoffmann et al. 2003, 2009; Lawes et al. 2011) have earlier reported higher relative bark thickness in trees of frequently burned environments. A study by Balch et al. (2011) in Brazilian Amazon forest, shows the survival of two commonly thin-barked species. In a way, bark thickness has already being used as a functional trait in managing forest structure. However, understanding of bark as a functional trait is still in infancy. For example, the relative bark thickness as a metric has hardly been used to understand community ecology and measurement.
Conclusion
Our study on the relationship between fire incidence (man-made fires) and bark thickness based on four dominant tree species of Central Himalayas indicates that bark thickness in the stems near the ground is a principal adaptation to small surface forest fires. Apart from the absolute bark thickness, higher relative bark thickness at an early age is a key fire adaptation. A rapid thickening of the bark when tree individuals are young, and more vulnerable to fire plays a key role in their survival. Species adapted to frequent surface fires allocate a substantial proportion of reserves to stem bark development, when young, and then decrease the bark allocation when trees are large and out of reach of surface fires.
Author contribution statement
SPS, SG and RDS conceived the idea for this article and designed the methodology. SG and RDS collected, and analyzed the data under the guidance of AT and JR. SG and RDS prepared the first draft of the manuscript under the supervision of SPS. All authors provided inputs and commented on subsequent versions of the manuscript. All authors have read and agreed to the final version of the manuscript.
Data accessibility
We have not included any data in a publicly accessible repository.
References
Agee JK (1993) Fire ecology of Pacific Northwest forests. Island Press, Washington, D.C
Balch JK, Nepstad DC, Curran LM, Brando PM, Portela O, Guilherme P, Reuning-Scherer JD, de Carvalho JO (2011) Size, species, and fire behavior predict tree and liana mortality from experimental burns in the Brazilian Amazon. For Ecol Manag 261(1):68–77
Barden LS (1978) Serotiny and seed viability of Pinus pungens in the Southern Appalachians. Castanea 44:44–47
Beaty RM, Taylor AH (2007) Fire disturbance and forest structure in old-growth mixed conifer forests in the northern Sierra Nevada, California. J Veg Sci 18:879–890
Cavender-Bares J, Ackerl DD, Baum DA, Bazzaz FA (2004) Phylogenetic over dispersion in Floridian oak communities. Am Nat 163:823–843
Champion HG, Seth SK (1968) A revised survey of the forest types of India. Government of India Press, New Delhi
Cornelissen JHC, Lavorel S, Garnier E, Diaz S, Buchmann N, Gurvich DE, Reich PB, Ter Steege H, Morgan HD, Van Der Heijden MGA, Pausas JG, Poorter H (2003) A handbook of protocols for standardised and easy measurement of plant functional traits worldwide. Aust J Bot 51:335–380
Dickinson MB, Johnson EA (2001) Fire effects on trees. In: Johnson EA, Miyanishi K (eds) Forest fires: behaviour and ecological effects. Academic Press, New York, pp 477–525
Ferrenberg S, Mitton JB (2014) Smooth bark surfaces can defend trees against insect attack: resurrecting a ‘slippery’ hypothesis. Funct Ecol 28:837–845
Franklin SB, Robertson PA, Fralish JS (1997) Small Scale fire temperature patterns in upland Quercus communities. J Appl Ecol 34:613–630
FSI (2019) Indian state of forest report. Forest Survey of India, Dehradun
Gamble JS (1985) A manual of Indian timbers: an account of the growth, distribution, and uses of the trees and shrubs of India and Ceylon, with descriptions of their wood-structure, 2nd edn. International Book Distributors, Dehra Dun
Gonzalez ME, Veblen TT, Sibold JS (2010) Influence of fire severity on stand development of Araucaria araucana–Nothofagus pumilio stands in the Andean cordillera of south-central Chile. Austral Ecol 35:597–615
He T, Pausas JG, Belcher CM, Schwilk DW, Lamont BB (2012) Fire-adapted traits of Pinus arose in the fiery Cretaceous. New Phytol 194:751–759
Hellmann JJ, Byers JE, Bierwagen BJ, Dukes JS (2008) Five potential consequences of climate change for invasive species. Conserv Biol 22:534–543. https://doi.org/10.1111/j.1523-1739.2008.00951.x
Hoffmann WA, Orthen B, Do Nascimento PKV (2003) Comparative fire ecology of tropical savanna and forest trees. Funct Ecol 17:720–726
Hoffmann WA, Adasme R, Haridasan M, de Carvalho MT, Geiger EL, Pereira MAB, Gotsch SG, Franco AC (2009) Tree topkill, not mortality, governs the dynamics of savanna-forest boundaries under frequent fire in central Brazil. Ecol 90:1326–1337. https://doi.org/10.1890/08-0741.1
Hoffmann WA, Geiger EL, Gotsch SG, Rossatto DR, Silva LCR, Lau OL et al (2012) Ecological thresholds at the savanna-forest boundary: how plant traits, resources and fire govern the distribution of tropical biomes. Ecol Lett 15:759–768
Jackson JF, Adams DC, Jackson UB (1999) Allometry of constitutive defense: a model and a comparative test with tree bark and fire regime. Am Nat 153:614–632
Keeley JE (2012) Ecology and evolution of pine life histories. Ann for Sci 69:445–453
Kraus D, Goldammer JG (2007) Fire regimes and ecosystems: an overview of fire ecology in tropical ecosystems. In: Schmerbeck J, Hiremath A, Ravichandran C (eds) Forest fires in India – workshop proceedings. ATREE, Bangalore, India and Institute of Silviculture, Freiburg, pp 9–13
Lawes MJ, Richards A, Dathe J, Midgley JJ (2011) Bark thickness determines fire resistance of selected tree species from fire-prone tropical savanna in north Australia. Plant Ecol 212:2057–2069. https://doi.org/10.1007/s11258-011-9954-7
Lawes MJ, Midgley JJ, Clarke PJ (2013) Costs and benefits of relative bark thickness in relation to fire damage: a savanna/forest contrast. J Ecol 101:517–524
Little S (1974) Effects of fire on temperate forests: north- eastern United States. In: Kozlowski TT, Ahlgren CE (eds) Fire and ecosystems. Academic Press, New York, pp 225–250
Miehe G, Pendry CA, Chaudhary RP (2015) Nepal: an introduction to the natural history, ecology and human environment of the Himalayas. Royal Botanic Garden Edinburgh, Edinburgh
Negi GCS (2019) Forest fire in Uttarakhand: causes, consequences and remedial measures. Int J Ecol Environ Sci 45(1):31–37
Paine CET, Stahl C, Courtois EA, Patino S, Sarmiento C, Baraloto C (2010) Functional explanations for variation in bark thickness in tropical rain forest trees. Funct Ecol 24:1202–1210
Parashar A, Biswas S (2003) The impact of forest fire on forest biodiversity in the Indian Himalayas (Uttaranchal). In: XII World Forestry Congress, 358:B-1. https://www.fao.org/3/XII/0358-B1.htm#fn1
Pausas JG (2015) Bark thickness and fire regime. Funct Ecol 29:315–327. https://doi.org/10.1111/1365-2435.12372
Pausas JG, Keeley JE (2009) A burning story: the role of fire in the history of life. Bioscience 59:593–601
Pausas JG, Ribeiro E (2013) The global fire-productivity relationship. Glob Ecol Biogeogr 22:728–736
Pellegrini AFA, Anderegg WRL, Paine CET, Hoffmann WA, Kartzinel T, Rabin SS, Sheil D, Franco AC, Pacala SW (2017) Convergence of bark investment according to fire and climate structures ecosystem vulnerability to future change. Ecol Lett 20:307–316. https://doi.org/10.1111/ele.12725
Peterson DL, Ryan KC (1986) Modelling of post-fire conifer mortality for long-range planning. Environ Manage 10:797–808. https://doi.org/10.1007/BF01867732
Poorter L, Mcneil A, Hurtado VH, Prins HHT, Putz FE (2014) Bark traits and life history strategies of tropical dry- and moist forest trees. Funct Ecol 28:232–242
Ralhan PK, Khanna RK, Singh SP, Singh JS (1985) Certain phenological characters of the shrub layer of Kumaun Himalayan forests. Vegetatio 63:113–119. https://doi.org/10.1007/BF00044061
Rothermel RC, Deeming JE (1980) Measuring and interpreting fire behaviour for correlation with fire effects. USDA Forest Service General Technical Report INT-93
Singh SP, Singh JS (1986) Structure and function of the Central Himalayan oak forests. Proc Plant Sci/indian Acad Sci 96:159–189
Singh JS, Singh SP (1992) Forests of Himalaya. Gyanodaya Prakashan, Nainital
Singh JS, Singh SP (1987) Forest vegetations of the Himalaya. Bot Rev 53:80–192
Singh RD, Gumber S, Tewari P, Singh SP (2016) Nature of forest fires in Uttarakhand: frequency, size and seasonal patterns in relation to pre-monsoonal environment. Curr Sci 111(2):398–440
Singh SP, Singh RD, Gumber S, Bhatt S (2017) Two principal precipitation regimes in Himalayas and their influence on tree distribution. Trop Ecol 58:679–691
Stainton JDA (1972) Forests of Nepal. John Murray, UK
Stephens SL, Libby WJ (2006) Anthropogenic fire and bark thickness in coastal and island pine populations from Alta and Baja California. J Biogeogr 33:648–652
Troup RS (1921) The silviculture of Indian trees, vol I. International Book Distributors, Dehradun
Uhl C, Kauffman JB (1990) Deforestation, fire susceptibility, and potential tree responses to fire in the eastern Amazon. Ecology 71:437–449. https://doi.org/10.2307/1940299
Uttarakhand Forest Statistics (2018) Uttarakhand forest statistics 2017–18. Forest Department Uttarakhand, Uttarakhand
van Mantgem P, Schwartz M (2003) Bark heat resistance of small trees in Californian mixed conifer forests: testing some model assumptions. For Ecol Manag 178:341–352
Van Nieuwstadt MGL, Sheil D (2005) Drought, fire and tree survival in a Borneo rain forest, East Kalimantan, Indonesia. J ECol 93:191–201
Acknowledgements
SG, RDS, AT and JR are grateful to Department of Forestry and Environmental Science, DSB Campus, Kumaun University for liberal support. All the authors acknowledge the research facilities provided by Central Himalayan Environment Association (CHEA), Nainital (Uttarakhand), India and Indian National Science Academy to provide Honorary Scientist Fellowship to one of the author (SPS). The inputs from the reviewers has helped us a lot in improving the manuscript. We have taken the liberty to acknowledge this.
Funding
The authors declare no specific funding for this work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Additional information
Communicated by H. Gärtner .
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Gumber, S., Singh, R.D., Ram, J. et al. Bark thickness analysis of four dominant tree species of Central Himalayan forests varying in exposure to surface fires. Trees 36, 685–695 (2022). https://doi.org/10.1007/s00468-021-02239-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-021-02239-z