
Abstract
Key message
Bark allocation patterns did not follow fire-tolerance grouping of tree species in all cases; more studies are needed on bark development from juvenile to adult trees.
Abstract
Thick bark is presumed to be a trait of fire-tolerant trees that were historically subjected to frequent surface fires. To be recruited into the forest canopy, a tree must be able to survive fire and grow thick enough bark during the fire-free interval. Previous studies have also suggested that fire-tolerant species grow disproportionately thick bark lower on the stem, where the threat of fire is greatest. The objectives of this study were to quantify how bark develops in six juvenile hardwood species of the Appalachian Piedmont and compare bark allocation between fire-tolerant (Carya tomentosa, Quercus alba, and Quercus velutina) and mesophytic species (Acer rubrum, Fagus grandifolia, and Liriodendron tulipifera). We sampled up to 32 individuals of each species and modeled bark area as a function of wood area and measure height using mixed effect models. Bark area increased approximately 0.4 cm2 for Q. velutina and C. tomentosa, 0.2 cm2, for L. tulipifera and Q. alba, and 0.1 cm2 for A. rubrum and F. grandifolia for each cm2 increase in wood area, holding measure height constant. For Q. velutina, C. tomentosa, and L. tulipifera, the measure height was found to decrease bark area by approximately 0.02, 0.01, and 0.01 cm2 for each cm increase in height, holding diameter constant. A paired sample Wilcoxon test comparing bark:wood ratio at 0 and 140 cm heights yielded similar results. Differences in bark allocation among the six species were not consistent with current groupings of “fire-tolerant” and “mesophytic” species.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Trees in their seedling and sapling stage can be readily killed or top-killed even by fire of low intensity. Consequently, a high frequency (once per 1–3 year) surface fire regime often supports treed savanna or grassland (Van Lear and Waldrop 1989; Frost 1998). To be recruited into the forest canopy, a stem must be able to survive fire and grow thick enough bark during the fire-free interval to protect the cambium from the potentially lethal temperatures of the next fire (Hare 1965). Thus, fire regime can serve as a selective force, favoring thick-barked species under frequent surface fires (Pausas 2015).
It has been well established that bark thickness increases with stem size. As a result, seedlings and saplings have the greatest potential to die or be topkilled from surface fires. To quickly gain the protection that bark offers (i.e., reach a critical bark thickness), fire-tolerant species would benefit from a strategy that makes bark growth a high priority during the seedling and sapling stages. Allometric studies have found that early investment in bark (negative allometry) is associated with species that inhabit drier sites with open canopies that experience frequent low-intensity fires, while species with late investments in bark are associated with wetter sites, closed canopies, and infrequent, higher intensity fires (Jackson et al. 1999; Schwilk et al. 2013). Thus, bark growth for species with negative allometry is reduced in larger individuals, while species with constant bark allocation may eventually develop bark as thick as, or even thicker than, fire-tolerant species in the absence of fire. Failure to incorporate juvenile stems in a bark study may result in the erroneous conclusion that there is no difference in bark thickness between fire-tolerant and fire-intolerant species.
Many tree species in the Carya and Quercus genera are considered fire-tolerant species that dominated eastern deciduous forests under historical fire regimes (Abrams 1992). Quercus-dominated forests are thought to have arisen due to intermediate, low-intensity surface fires (Frost 1998). Fire suppression since the early 1900s is causing a positive feedback loop of ecological succession in which fire-tolerant species are competitively excluded by shade-tolerant, fire-intolerant species such as Acer spp. and Fagus grandifolia. The increased dominance of these so-called “mesophytic” species would, in turn, reduce fire occurrence, a process known as “mesophication” (Nowacki and Abrams 2008). In the absence of fire, these mesophytic species create a densely shaded, mesic understory with a compacted leaf litter layer that is less flammable (Kreye et al. 2013), creating microenvironments that are unconducive to oak recruitment to the understory (Nowacki and Abrams 2008). To rectify this trend, prescribed fire has been widely studied in recent decades for their role in promoting Quercus regeneration (e.g., Brose and Van Lear 1998; Brose et al. 2001, 2006). Review of the fire-Quercus literature reveals mixed results, with studies reporting that fire improved, hindered, or had no effect on Quercus spp. regeneration (Brose et al. 2006). To improve Carya spp. and Quercus spp. understory dominance with prescribed fire, the mechanisms that Carya spp. and Quercus spp. to survive surface fire in the seedling and sapling stage must be examined.
Bark development requires an investment in carbon resources, which may otherwise be used for height or stem wood growth. An optimal strategy for species in ecosystems that experience frequent low-intensity surface fires is to develop thicker basal bark, where protection from surface fires is most needed, and devote fewer resources to bark growth higher on the stem, where damage from surface fire would be less severe or less likely. There is increasing evidence that some fire-tolerant species have disproportionately thick basal bark. Graves et al. (2014) demonstrated that Floridian Quercus species in fire prone habitats decreased their relative and absolute bark thickness with height, whereas Quercus species in non-fire prone areas did not. Odhiambo et al. (2014) found that tapering of outer bark with height occurred more in Pinus species with higher heat resistance capacity (P. pinaster and P. elliottii), as opposed to the thin barked P. radiata. Hammond et al. (2015) documented the bark taper along the stem between hardwood species in Pinus palustris savannas. They found that species that commonly co-occur with frequently burned P. palustris, particularly Q. marilandica, had greater bark taper and higher bark:wood ratio than mesophytic species such as Acer rubrum. There is a paucity of knowledge of bark thickness development in species of Carya and Quercus found in the Appalachian Piedmont, where a frequent surface fire regime was historically dominant (Van Lear and Waldrop 1989).
The objectives of this study were (1) to quantify how bark develops in juvenile hardwood species of the Appalachian Piedmont, and (2) to compare bark allocation between fire-tolerant and mesophytic species. We hypothesized that fire-tolerant species devote more resources to bark growth compared to mesophytic species during the juvenile stage. We also hypothesized that bark allocation decreases with height in fire-tolerant species, while mesophytic species have constant bark to wood ratio.
Materials and methods
Study site
This study was conducted in the Clemson Experimental Forest (CEF), located in Clemson, South Carolina. The CEF consists of 7082 ha of primarily Quercus spp. and Pinus taeda dominated forests. Most of the pine stands were planted, while oak-dominated hardwood stands originated naturally in the 1930s as a result of a land reclamation project in which eroded agriculture lands were stabilized and converted into forest. As with much of the southern Piedmont region of the United States, soil orders in the CEF include Ultisols, Entisols, and Inceptisols (USDA-NRCS 2015). The majority of the soils in the CEF are in the Cecil, Pacolet and Madison series, in the thermic family.
Sampling methods
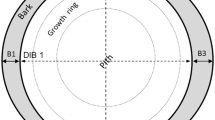
Juvenile hardwood species, defined as ground-line diameter less than 10 cm, were selected at various locations in the CEF based on the availability of a range of small diameter stems and those that were considered in the literature to be either fire-tolerant or mesophytic. Species sampled included Acer rubrum L., Fagus grandifolia Ehrh., Carya tomentosa [Poir.] Nutt., Liriodendron tulipifera L., Quercus. alba L., and Q. velutina Lam. Given their pre-colonial dominance, we considered C. tomentosa, Q. alba, and Q. velutina to be fire-tolerant, while A. rubrum, F. grandifolia, and L. tulipifera were considered mesophytic (Abrams 1992; Nowacki and Abrams 2008). Stems were initially cut in the field with handsaws as close to the ground as possible. Acer rubrum, Q. alba, and Q. velutina were sampled from the same oak shelterwood stand which had recently been harvested. Carya tomentosa and F. grandifolia were both sampled from a mixed hardwood stand under a canopy of mature trees. Liriodendron tulipifera was sampled from a stand that seeded into a former Pinus taeda clearcut. Although prescribed fire is occasionally conducted in the CEF, none of the stands sampled were frequently burned to our knowledge. In the lab, each stem was cut at approximately 0, 30, 60, 90 and 140 cm using a band saw. For each cross section, three diameter outside of bark (DOB) and three diameter inside of bark (DIB) measurements were taken approximately 60° apart using digital calipers. Double bark thickness was determined by subtracting DIB from DOB. The three double bark measurements were then added together and divided by six to estimate the average bark thickness (BT) for the cross section. The three DOB and DIB measurements were also averaged to get an estimate for the average DOB and DIB, respectively. We did not distinguish between inner and outer bark, thus BT in this study consists of total bark thickness. Some stems (especially the smaller ones) were sanded to help distinguish between bark and wood, but the initial cut with the band saw was sufficient for most of the larger stems. A hand lens was also used on some of the smallest stems where bark and wood were difficult to distinguish. Table 1 shows a summary of stems measured at 0 and 140 cm heights.
Data analysis
Bark area (B a ) was calculated for each cross section by subtracting the wood basal area from the basal area of the bark using the formula
We conducted multiple mixed-effect models for each species to compare bark growth. Because multiple measurements along the same stem are not independent, each individual tree was assigned a unique number and that number was included as a random effect (Tree). For each species we fit two models where the full model included bark area in cm2 as a function of wood area (W a) in cm2 and measure height (MHT) in cm, while the reduced model includes bark area as a function of wood area only:
Models were fit in R (version 3.3.1, R Core Team 2016) using the lmer function in the lme4 package (Bates et al. 2015). Second-order Akaike Information Criterion (AICc) was used for model selection (Hurvich and Tsai 1989). Pseudo-R 2 values and AICc were calculated with the r.squaredGLMM and AICc functions in the MuMIn package (Barton 2016).
To test if species allocate more resources to bark at lower stem heights, we compared the difference in bark:wood ratios from the 0 cm (B:W 0 ) and the 140 cm (B:W 140 ) cross sections,
We used a paired sample Wilcoxon test for each species to see if the difference in bark:wood ratio from 0 to 140 cm was significantly greater than zero, which would indicate taper in bark:wood ratio with height.
Results
The full mixed-effect model, which included measure height, was a better fit for C. tomentosa, Q. velutina, and L. tulipifera, while the reduced model was more appropriate for all other species (Table 2). Pseudo-R 2 values were high for all models, with fixed effects explaining 85, 92, 77, 96, 91, and 96% of the variance for C. tomentosa, Q. alba, Q. velutina, A. rubrum, F. grandifolia, and L. tulipifera, respectively. Bark allocation was highest for Q. velutina, followed by C. tomentosa, L. tulipifera, Q. alba, A. rubrum, and F. grandifolia. For these species, bark area increased approximately 0.4 cm2 for Q. velutina and C. tomentosa, 0.2, for L. tulipifera and Q. alba, and 0.1 cm2 for A. rubrum and F. grandifolia for each cm2 increase in wood area, holding MHT constant. For the species where the full model was more appropriate, MHT was found to decrease bark area by approximately 0.02 cm2 for each cm increase in height, holding diameter constant, for Q. velutina and 0.01 cm2 for C. tomentosa hickory and L. tulipifera.
The results of the paired sample Wilcoxon test agreed with our mixed models. Carya tomentosa, Q. velutina, and L. tulipifera all showed significant (P < 0.01) taper in bark:wood ratio, with median ratio difference of 0.26, 0.18, and 0.08, respectively, between the two compared heights (Fig. 1). Although the A. rubrum median ratio difference was significant from zero (P = 0.03), the median difference was only 0.01. Fagus grandifolia and Q. alba did not show significant bark taper.

Difference in bark:wood ratios from 0 to 140 cm heights for six common hardwood species in the Clemson Experimental Forest. Boxes represent the interquartile range (IQR) with whiskers extending to 1.5 × IQR. Points are trees with bark:wood ratio differences beyond 1.5 × IQR. Bars within each box represent the median difference. *Species with changes significantly different than 0 at P < 0.001 in a Wilcoxon rank sum test
Discussion
Developing thick bark requires an investment in resources and a cost of limiting the diffusion of gases and water (Pausas 2015). Thus, producing thick bark is only advantageous if its benefits outweigh the costs, as may be the case of fire prone areas. To limit the costs of producing thick bark, a species may reduce bark allocation after reaching a critical thickness. To illustrate any potential changes of bark allometry, future studies must collect data over a wide range of tree sizes because any change in bark allometry may not occur until a tree reaches a stem size with bark thick enough to protect cambium from fires.
Another way to limit the cost of producing thick bark is to limit the production to areas that would need more protection during a surface fire: the lower portion of the stem. Our findings only partially agree with this hypothesis. Carya tomentosa and Q. velutina displayed bark taper, but Q. alba did not. Liriodendron tulipifera, which is traditionally grouped with the mesophytic species (Abrams 1992; Nowacki and Abrams 2008), did show evidence of bark:wood ratio taper. Hammond et al. (2015) found similar mixed results for hardwoods in Pinus palustris savannas. They found that Q. marilandica, a common associate of P. palustris and presumably a fire-tolerant species, had the highest absolute bark thickness as well as the highest degree of bark:wood ratio taper. Carya pallida, another species common to the ecosystem, did not have significant bark taper (Hammond et al. 2015). Midgley and Lawes (2016) suggest that the decline of bark allocation with height is so small that it should not be taken into account except for a few cases where thick basal bark is retained as in Eucalyptus and Corymbia species. Based on our results, C. tomentosa, L. tulipifera, and Q. velutina all had significant taper with height, having slightly thicker basal bark relative to size. The degree to what these gains in bark thickness at lower stem heights contributes to survival in fire prone systems is still unclear.
Although there is general agreement that fire historically maintained oak-hickory dominance in hardwood forests, the evidence in the literature on the species in our study is mixed. For instance, Q. velutina had the highest relative bark thickness and largest amount of bark:wood ratio taper with height, but is considered only moderately fire-resistant (Carey 1992). Sander (1990) suggests that wildfires can easily kill Q. velutina stems up to pole size (10 cm diameter at 137 cm height), which includes the range of our sample, by damaging the cambium and allowing entry for fungal decay. Elliott et al. (1999) reported that Q. alba and Q. velutina saplings suffered high mortality and did not regenerate in the understory after a prescribed burn in the southern Appalachians in 1995, while other oak species increased in importance. Yet fire is said to be the responsible factor for the establishment of Q. alba and Q. velutina dominated stands (Tirmenstein 1991). Similarly, the evidence of fire resistance is mixed in Cary tomentosa, which had the second thickest absolute bark for any given diameter and showed evidence of bark:wood ratio taper with height. Carya tomentosa bark has been cited as having low insulating capacity and as being easily killed by fire (Smith 1990; Coladonato 1992). However, Brose and Van Lear (1998) found that advanced regeneration of hickory (C. pallida and C. tomentosa combined) had significantly lower mortality than oak (Q. alba, Q. coccinea, Q. prinus, Q. rubra, and Q. velutina combined), A. rubrum, and L. tulipifera in varying intensity prescribed burns in the Piedmont of Virginia. It is important to note that Brose and Van Lear (1998) did not separate species within Carya and Quercus in their study. Our study, as well as others show that bark development (and possibly fire resistance) in the oak genus varies significantly by species (Graves et al. 2014; Hammond et al. 2015). It is likely that a large variation in bark development also exists in the Carya genus as well. Future studies would benefit from analysis at the species level to possibly resolve the discrepancies in fire resistance currently described in the literature.
Our results suggest that dichotomy of fire-tolerant (i.e., oaks and hickories) and fire-sensitive (i.e., mesophytic species) is somewhat simplistic if bark thickness is the critical trait determining fire survival of juvenile trees. Bark thickness is part of a suite of traits, including flammable litter and fast wound closure, which can help certain species persist in fire prone areas (Varner et al. 2016). Some of the species found in the CEF exhibit traits in common with those of other fire-tolerant species around the world. Documenting these traits is important in furthering our understanding of how fire has influenced the distribution of tree species.
Author contribution statement
TMS: field data collection, lab data preparation, data analysis, and preparing the manuscript; GGW: research idea initiation, design of field methods, design of lab methods, and contribution to manuscript; PTM: field data collection, lab data preparation, and contribution to manuscript; SG: field data collection and lab data preparation.
References
Abrams MD (1992) Fire and the development of oak forests. BioScience 42:346–353
Barton K (2016) MuMIn: multi-model inference. R package version 1.15.6
Bates D, Maechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Brose PH, Van Lear DH (1998) Responses of hardwood advance regeneration to seasonal prescribed fires in oak-dominated shelterwood stands. Can J For Res 28:331–339
Brose PH, Schuler TM, Van Lear DH, Berst J (2001) Bringing fire back: the changing regimes of the Appalachian mixed-oak forests. J For 99:30–35
Brose PH, Schuler TM, Ward JS (2006) Responses of oak and other hardwood regeneration to prescribed fire: what we know as of 2005. In: Dickenson MB (ed.) 2006. Fire in eastern oak forests: delivering science to land managers, proceedings of a conference; 2005 November 15–17; Columbus, OH. Gen. Tech. Rep. NRS-P-1. Newtown Square, PA: U.S. Department of Agriculture, Forest Service, Northern Research Station: 123–135
Carey JH (1992) Quercus velutina. In: Fire effects information system, [Online]. US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). http://www.fs.fed.us/database/feis/. Accessed 1 Oct 2016
Coladonato M (1992) Carya tomentosa. In: Fire effects information system, [Online]. US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). http://www.fs.fed.us/database/ feis/. Accessed 1 Oct 2016
Elliott KJ, Hendrick RL, Major AE, Vose JM, Swank WT (1999) Vegetation dynamics after a prescribed fire in the southern Appalachians. For Ecol Manage 114:199–213
Frost CC (1998) Presettlement fire frequency regimes of the United States: a first approximation. In: Pruden TL, Brennan LA (eds) Fire in ecosystem management: shifting the paradigm from suppression to prescription, pp 70–81
Graves SJ, Rifai SW, Putz FE (2014) Outer bark thickness decreases more with height on stems of fire-resistant than fire-sensitive Floridian oaks (Quercus spp.; Fagaceae). Am J Bot 101:2183–2188
Hammond DH, Varner JM, Kush JS, Fan Z (2015) Contrasting sapling bark allocation of five southeastern USA hardwood tree species in a fire prone ecosystem. Ecosphere 6:1–13
Hare RC (1965) Contribution of bark to fire resistance of southern trees. J For 63:248–251
Hurvich CM, Tsai CL (1989) Regression and time series model selection in small samples. Biometrika 76:297–307
Jackson JF, Adams DC, Jackson UB (1999) Allometry of constitutive defense: a model and a comparative test with tree bark and fire regime. Am Nat 153:614–632
Kreye JK, Varner JM, Hiers JK, Mola J (2013) Toward a mechanism for eastern North American forest mesophication: differential litter drying across 17 species. Ecol App 23:1976–1986
Midgley JJ, Lawes MJ (2016) Relative bark thickness: towards standardised measurement and analysis. Plant Ecol 217:677–681
Nowacki GJ, Abrams MD (2008) The demise of fire and “mesophication” of forests in the eastern United States. Bioscience 58:123–138
Odhiambo B, Meincken M, Seifert T (2014) The protective role of bark against fire damage: a comparative study on selected introduced and indigenous tree species in the Western Cape, South Africa. Trees 28:555–565
Pausas JG (2015) Bark thickness and fire regime. Func Ecology 29:315–327
R Core Team (2016) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/. Accessed 1 Oct 2016
Sander IL (1990) Quercus velutina Lam. (Black oak). In: Burns RM. Honkala BH, technical coordinators. Silvics of North America. Vol. 2. Hardwoods. Agricultural Handbook 654. https://www.na.fs.fed.us/spfo/pubs/silvics_manual/volume_2/quercus/velutina.htm. Accessed 1 Oct 2016
Schwilk DW, Gaetani MS, Poulos HM (2013) Oak bark allometry and fire survival strategies in the Chihuahuan Desert Sky Islands, Texas, USA. PLoS ONE 8:e79285
Smith HC (1990) Carya tomentosa (Poir.) Nutt. (Mockernut hickory). In: Burns RM, Honkala BH, technical coordinators. Silvics of North America. Vol. 2. Hardwoods. Agricultural Handbook 654. https://www.na.fs.fed.us/spfo/pubs/silvics_manual/volume_2/carya/tomentosa.htm. Accessed 1 Oct 2016
Tirmenstein DA (1991) Quercus alba. In: Fire effects information system, [Online]. US Department of Agriculture, Forest Service, Rocky Mountain Research Station, Fire Sciences Laboratory (Producer). http://www.fs.fed.us/database/ feis/. Accessed 1 Oct 2016
USDA-NRCS (2015) Natural resources conservation service, United States Department of Agriculture. Web Soil Survey. http://websoilsurvey.nrcs.usda.gov/. Accessed 1 Oct 2016
Van Lear DH, Waldrop TA (1989) History, uses, and effects of fire in the Appalachians. USDA Forest Service Gen. Tech. Rep. SE-54
Varner JM, Kane JM, Hiers JK, Kreye JK, Veldman JW (2016) Suites of fire-adapted traites of oaks in the southeastern USA: multiple strategies for persistence. Fire Ecol 12:48–64
Acknowledgements
This study was partially funded by the Clemson Foundation. We thank Bridget Blood for reviewing an early version of this manuscript. We also thank the two anonymous reviewers, whose comments greatly improved the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Y. Sano.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Shearman, T.M., Wang, G.G., Ma, P.T. et al. Patterns of bark growth for juvenile trees of six common hardwood species in the eastern United States and the implications to fire-tolerance. Trees 32, 519–524 (2018). https://doi.org/10.1007/s00468-017-1649-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-017-1649-9