
Abstract
Key message
The influence of climate in the previous year on radial growth was stronger than that in the current year and climatic influence became weaker as the altitude increased.
Abstract
The climate conditions during the previous year often influence the radial growth of trees in the current year, which is known as the lingering effect of climate. To explore this lingering effect in depth, we compare the influence of climate in the previous and current year on radial growth and find variations in climatic effects on growth along the altitudinal gradient. We also relate radial growth of Picea crassifolia to climate variables from 24 different timescales (from 1 month to the previous 24 months) at different altitudes in the middle of the Qilian Mountains. In the study area, accumulated monthly precipitation before November in the previous year benefited radial growth in the current year at all altitudes. At lower altitudes, July could be seen as a demarcation point that precipitation from the current May–July significantly affected radial growth in the current year, but precipitation from the current July–October significantly affected radial growth in next year. At the same time, radial growth there responded stronger to precipitation from the previous July–October than that from the current May–July. Radial growth at higher altitudes responded weaker to climatic factors than that at lower altitudes and rarely correlated with precipitation from the current year.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Tree growth is directly influenced by the current environment, but environmental conditions during the previous year often have lingering effects on current growth as a result of the induction of physiological preconditioning (Fritts 2012). With regard to arid-site conifers, physiological preconditioning for the current year’s radial growth includes physiological processes (e.g., net photosynthesis, respiration and transpiration) from the previous growing season and the accumulation and storage of carbohydrates and other substances during the dormant period (Helle and Schleser 2004; Gruber et al. 2013). For example, some studies found that carbohydrates accumulated at the end of the growing season during the previous year were associated with winter freezing tolerance (Gruber et al. 2013), which may affect growth in the current year. Temperature and moisture conditions from the previous year usually effect growth in the current year by influencing these physiological process and carbohydrate reserves. For example, high temperature and low moisture during the midsummer of the previous year, when cambial activity slows down, impinged more on the accumulation of stored food than on growth at that time, which in turn affected growth in the current year (Fritts 2012).
The phenomenon that the climate in the previous and current years influences current radial growth has been reflected in tree-ring analysis, i.e., tree-ring width indexes in the current year show significant correlation with monthly climatic data in the previous and current growing seasons (Fritts 2012). According to previous studies, the reason may be that the climate from the previous growing season partially influences the first part of earlywood formation in the current year, whereas the climate in the current growing season affects partial earlywood and latewood formation in the current year (Kuptz et al. 2011; Sidor et al. 2015). However, previous studies have not focused on the comparison between the effect of climate in the previous and current years. It is not known which influence is stronger for radial growth.
The influences of climate in the previous and current years on current growth usually vary with altitude (Ettl and Peterson 1995; Savva et al. 2006; Cailleret and Davi 2011). In some humid areas, trees at high altitude were mainly limited by low temperature rather than moisture, whereas trees at low-altitude sites tended to suffer from high water stress accompanied by high temperature (Affolter et al. 2010; Chen et al. 2011; Wang et al. 2013; Jiang et al. 2014). In some relatively arid regions, climate–radial growth relationships showed altitude independence and growth was mainly controlled by precipitation at different elevations (Wang et al. 2005; Esper et al. 2007; Yang et al. 2013). Picea crassifolia at lower altitudes in northwest China was found significantly correlated with climatic factors during the previous year, including the total precipitation in July, August and September and the mean temperature of July and August (Liang et al. 2009; Liu et al. 2013). Picea crassifolia at higher altitudes was found significantly correlated with the total precipitation and the mean temperature during September and October in the previous year (Liang et al. 2010; Zhang et al. 2012; Bond-Lamberty et al. 2014). This indicates the complexity and regularity of the relationship between climate and radial growth at different altitudes.
The Qilian Mountains are located on the northeastern margin of the Tibet Plateau in northwest China. Because they are in a climate transition zone between the East Asian Monsoon and the Westerlies, the region is highly sensitive to climate (Zheng 1996). Picea crassifolia is endemic and widespread at the northeastern margin of the Tibet Plateau, and more than 90 % of Picea crassifolia forests are distributed in the Qilian Mountains (Liu 1992). There have been many dendrochronological studies of Picea crassifolia carried out on the Qilian Mountain because of its climatic sensitivity. Nearly half of these studies focused on the reconstruction of past climate changes (Liang et al. 2009; Xu et al. 2012; Chen et al. 2012, 2013). Other studies focusing on the climate–radial growth relationships found an impact of climate from the previous year, which provided conditions for researching the lingering effect of climate in this area (Liang et al. 2006, 2010). In this study, we carried out sampling in the middle of the Qilian Mountains to analyze the climate–radial growth relationship at different altitudes for Picea crassifolia to test our hypothesis: radial growth responded stronger to climate in the current year than that in the previous year and responded weaker and weaker to climate along with increasing altitude. Specifically, this study aimed at (1) analyzing the influence of climate in the previous and current years on the current radial growth at different timescales, especially the lingering effect of previous climate; (2) comparing the influence of climate in the previous and current years on current radial growth; (3) exploring the variation of climate–radial growth relationships along the altitude gradients.
Materials and methods
Study area and climate
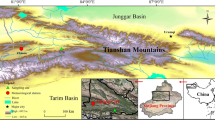
The Qilian Mountains (36.433°–40.017°N; 94.867°–103.150°E), on the northeastern margin of the Tibet Plateau, lie in a transition zone between the East Asian Monsoon and the Westerlies climates. A mix of these two weather patterns occurs in the Heihe River catchment area in the middle of the Qilian Mountains where primary forests of Picea crassifolia are distributed (Li and Liu 2000). The study area is located at the Sidalong Forest Farm (SDL) in the reaches of the Heihe River (Fig. 1). The total annual precipitation (1959–2011) increases with increasing altitude from 128.3 mm at the Zhangye meteorological station (38.933°N, 100.433°E, 1482 m) to 405.9 mm at the Qilian meteorological station (38.183°N, 100.25°E, 2787 m) and 415.2 mm at the Yeniugou meteorological station (38.417°N, 99.583°E, 3320 m). The mean annual temperature decreases with altitude from 7.5 °C at Zhangye to 1.1 °C at Qilian and −2.9 °C at Yeniugou (Fig. 2). The maximum value of precipitation and temperature occurred in July, and 63.6 % of the precipitation is concentrated between June and August. The study area is on a wet, shaded slope where the forest belt ranges from 2600 m to 3300 m a.s.l. and is dominated by Picea crassifolia (including moss-spruce forests, shrub-spruce forests, and meadow-spruce forests along the altitude gradient). The understory vegetation includes Salix oritrepha, Caragana jubata, Potentilla fruticosa, Lonicera microphylla, and Potentilla glabra (Wang et al. 2001).

Locations of the sampling site and three nearest meteorological stations in the study area (the Qilian Mountains)

Total monthly precipitation (a) and mean monthly temperature (b) at the meteorological stations of Zhangye (1482 m a.s.l.), Qilian (2787 m a.s.l.) and Yeniugou (3320 m a.s.l.) along the altitudinal gradients in the Qilian Mountains, calculated for the periods of 1959–2011
Tree-ring materials and characteristics
Tree-ring cores were sampled from a continuous north-facing slope at four elevations, named HE (i.e., highest, 3200 m a.s.l.), SH (i.e., sub-higher, 3000 m a.s.l.), SL (i.e., sub-lower, 2765 m a.s.l.) and LE (i.e., lowest, 2649 m a.s.l.), along the altitude gradient in August 2012 (Table 1). The sampling sites were chosen by maximizing forests’ representation of altitudinal distribution and minimizing habit difference induced by micro-geomorphological factors. In all, 200 cores from 100 trees (two cores from each tree) were sampled at the four sites using the 5.15-mm-diameter increment borers. Two cores from the same tree were perpendicular to each other, and one was parallel to the contour line. The cores were taken to the laboratory and treated according to the standard dendrochronological procedures (Fritts 2012): cores were fixed in wood tanks, burnished, cross-dated and measured with TSAP (Rinn 2003). The COFECHA program (Holmes 1983) was used to examine the results of the measurement and eliminate the short cores and those containing too many errors. Residual chronologies were calculated by detrending the tree-ring series with a 50-year cubic smoothing spline function in the ARSTAN program (Cook 1985).
The quality of chronologies was tested using parameters including the standard deviation (SD), mean sensitivity (MS), mean correlation between all series (Rbar), signal to noise ratio (SNR), variance in the first principal component (PC1) (Fritts 2012) and expressed population signal (EPS) (Wigley et al. 1984). The SD of the tree-ring width indexes in each series evaluates the variability in measuring the tree-ring series. The MS indicates the relative variability in ring width indexes between consecutive years and evaluates the climatic influence on tree-rings. The Rbar evaluates the common signal in tree-rings at one site. The SNR indicates the signal strength of the chronologies, and the PC1 explains the percent variance. An EPS level of 0.85 is used to indicate a satisfactory quality of chronologies (Wigley et al. 1984).
Pointer years from 1865 to 2011 were selected to evaluate the growth variability of trees at each site. We defined 1 year as a pointer year when the tree-ring width index of that year was one standard deviation higher or lower than the mean ring width index value during 1865–2011 (Liang et al. 2006). An ANOVA F test was calculated between the tree-ring width indexes of positive pointer years (and negative pointer years) at the four altitudes.
Analysis of climate–radial growth relationships
The corresponding climate data at each sampling site were calculated according to data of the two nearest meteorological stations in the vertical distance. Precipitation at each site was calculated according to the proportion of vertical distance to the two nearest meteorological stations which was similar to inverse distance weighted interpolation (Rahman and Lateh 2015). For example, the vertical distance of site HE (3200 m a.s.l) to the Yeniugou station (3320 m a.s.l) was 120 m, whereas the vertical distance to the Qilian station (2787 m a.s.l) was 413 m. Precipitation at HE was calculated according to the following equation:
where Pre(HE) indicates precipitation at site HE, Pre(Yeniugou) indicates precipitation at the Yeniugou station and Pre(Qilian) indicates precipitation at the Qilian station.
The temperature at each site was calculated based on the standard formula that temperature drops 0.4 °C with each 100 m increase in altitude. We computed the total precipitation and mean temperature as climatic factors for 24 different timescales, i.e., from 1 month up to the previous 24 months, similar to the method used to calculate the standardized precipitation evapotranspiration index (Vicente-Serrano et al. 2014). For example, the precipitation or temperature during July at the 5-month timescale was the total precipitation or mean temperature during March–July.
For the common period (1959–2011), Pearson correlations (abbreviated as “correlation”) were calculated between the residual chronologies at different altitudes and climatic factors from June during the previous year to October of the current year for tree-ring formation for the 24 different timescales using SPSS (version 19.0; SPSS Inc., Chicago; USA). High and significant correlation coefficients in the previous and current growing season at each elevation were selected to compare with each other. The partial correlation analysis was commonly used to find correlation between two variables after removing the effects of other variables (Yang et al. 2015). To compare the influence of climate in the previous and current year on current radial growth, a first-order partial correlation analysis was carried out on tree-ring width indexes and climatic factor in the previous year (or current year) by removing the effects of climatic factor in the current year (or previous year) using SPSS. In this study, the total precipitation in the previous July–October and current May–July was, respectively, selected as climatic factor in the partial correlation according to the results of the Pearson correlation tests.
Results
Characteristics of residual chronologies at different altitudes
The chronologies dated to 1796, 1795, 1813 and 1819 were at the highest altitude (HE), the sub-higher altitude (SH), the sub-lower altitude (SL) and the lowest altitude (LE), respectively. The high SNR values (25.514–43.842) and EPS values (0.962–0.978) that exceeded 0.85 indicated a high reliability of chronologies (Wigley et al. 1984). High values of the SD, MS and SNR showed that the four chronologies were all qualified for exploring climate–radial growth relationships. A high PC1 indicated a common response of growth to climatic factors in the study area. The chronologies at the highest site and the sub-higher site had lower values of the SD, MS, Rbar, PC1 and SNR (Table 2), reflecting that radial growth there was less sensitive to climate than that at the two lower altitudes in the context that microenvironment at the four sites was similar to each other.
The average tree-ring width indexes of positive and negative pointer years during 1865–2011 at the highest and sub-higher site did not have obvious differences, but did have significant differences with those at the sub-lower and lowest site (Fig. 3). The average tree-ring width indexes of positive pointer years at the highest and sub-higher site were lower and the average indexes of negative pointer years were higher than those at the other two sites.

Average tree-ring indexes of positive and negative pointer years during 1865–2011 at different altitudes. Bars for each parameter labeled with different letters differed significantly between sites (P < 0.05, ANOVA)
Precipitation–radial growth relationships at different altitudes
Radial growth at each altitude significantly correlated not only with total precipitation of 1 month but also with that of several months (Fig. 4). The intensity of color in Fig. 4a–d shows that the chronology at the higher altitude was less limited by precipitation. The most significant correlation coefficients between the previous and current growing season at different altitudes were selected (Table 3a). In the previous growing season, radial growth at the highest site showed the most significant correlation with the total precipitation during October (r = 0.331) and at the sub-higher site displayed the highest correlation with the total precipitation during October on the 2-month timescale (i.e., September–October, r = 0.418). The radial growth at the sub-lower and lowest site was most significantly correlated with total precipitation during October at the 4-month timescale (i.e., July–October, r = 0.477 and 0.496). In the current growing season, the radial growth at the highest and sub-higher site was not significantly correlated with precipitation, whereas growth at the sub-lower and lowest site showed the highest correlation with the total precipitation during July at the 3-month timescale (i.e., May–July, r = 0.284 and 0.453). Both the most significant Pearson correlation coefficients with precipitation in the previous growing season and in the current growing season showed downward trends over increasing altitudes.

Pearson correlation analysis of tree-ring width indexes at each site with (a–d) total precipitation and (e–h) mean temperature of previous June to current October from 24 different timescales (from 1 month to the previous 24 months). p6 to p12 represented previous June to previous December; c1 to c10 represented current January to current October. The black lines (a–d) and white lines (e–h) indicated a significance level of 0.05
Significant Pearson correlation and partial correlation coefficients of residual chronology with total precipitation during the previous July–October and the current May–July were mainly reflected at the two lower altitudes (Table 3). Both Pearson correlation and partial correlation coefficients of the residual chronology with the total precipitation from the previous July–October were higher than those with the total precipitation from the current May–July.
Temperature–radial growth relationships at different altitudes
Radial growth at each altitude was significantly and negatively correlated with the mean temperature of the previous July–August (Fig. 4) and the correlations became weaker along with increasing altitude (Fig. 5). Radial growth rarely showed a significant correlation with the mean temperature during the current year in the study area except at the highest site (Fig. 4).

Pearson correlation coefficients of tree-ring width indexes with mean temperature of previous July–August at each altitude. The dashed lines indicate a significance level of 0.05
Discussion
Lingering effect of previous precipitation on radial growth
The growing season of Picea crassifolia begins in the middle of March and ends in early November of the same year (Liu 1992). We defined 1–4 months before November as the preseason (Yang et al. 2015). Our study found that preseason accumulated precipitation in the previous year was closely and positively correlated to radial growth in the current year. We considered that more preseason accumulation of precipitation could increase soil moisture in the previous autumn and winter. The previous high soil moisture could favor high photosynthesis in the current spring when the temperature was favorable and growth began (Fritts 2012). However, more accumulated preseason precipitation may correspond to a later end of the growing season due to the mitigation of drought stress in late autumn (Yang et al. 2015). The delayed end of the growing season could facilitate the carbohydrate storage (Toromani et al. 2011). The abundantly stored carbohydrates during the previous autumn positively influenced tree-ring formation in the current year (Rossi et al. 2009; Huang et al. 2014) because initial tree growth in the current year may partially make use of stored carbohydrates (Kozlowski 1992).
Yang et al. (2015) also found that higher preseason precipitation was associated with a later end of the growing season in the area where the preseason mean temperature was below 13 °C, which agreed with our study area. By reviewing all of the dendrochronological studies regarding Picea crassifolia in the Qilian Mountains, we found that radial growth significantly and positively correlated with precipitation during the previous July, August and September from 2650 m to 3000 m a.s.l. and with precipitation in the previous September and October from 3100 to 3400 m a.s.l (Gou et al. 2005; Liang et al. 2010, 2016). In our study, the effect of preseason cumulative precipitation over long timescales was more obvious. The findings from former studies and our study all emphasize the lingering effect of preseason precipitation from the previous year on radial growth of the current year, which should be addressed in forest protection and management.
Comparison between influence of precipitation in the previous and current year on radial growth at lower altitudes
Both Pearson correlation and partial correlation coefficients between radial growth and total precipitation from the previous July–October and current May–July were significant at the level of 0.05 at the two lower altitudes. Interestingly, the two time periods of precipitation had a junction point (Fig. 6). July could be regarded as a demarcation point. In the growing season, the total precipitation before July (May–July) mainly influenced radial growth of the current year, whereas the total precipitation after July (July–October) had lingering effects on the growth for the next year. July may not be a highly precise time point, but our previous study in the middle of the Qilian Mountain also highlighted the demarcation significance of July (unpublished data).

Influence of total precipitation in May–July and July–October on radial growth. p7, p10 and p12 indicate July, October and December in the previous year; c5, c7, c10 and c12 indicate May, July, October and December in the current year; n5 and n7 indicate May and July in the next year
A possible reason for the occurrence of break point in July may be that the total precipitation during July–October of the previous year changed the moisture conditions and stored carbohydrates before the beginning of the growth in the current spring and influenced the first part of earlywood growth in the current year (Rossi et al. 2009; Huang et al. 2014). A suitable moisture environment during May–July could increase the cell formation rate and number of cells that delay the earlywood-to-latewood transition time (Xu et al. 2014) and the end of xylem maturation (Lupi et al. 2010). Good growth of needles and roots during this period also provides suitable conditions for the formation of latewood, which begins after needle and root elongation (Rossi et al. 2009). Therefore, high precipitation during the current May–July may benefit partial earlywood growth and latewood growth during the current year. Maximal mean monthly temperature emerges in July. We inferred that the high temperature in July impinged more on the accumulation of stored carbohydrate than on growth at that time when cambial activity slowed down. The stored carbohydrate would affect radial growth in the next year in turn (Fritts 2012). To find the more accurate reason, deeper physiological experiments were needed to carry out on Picea crassifolia. Radial growth of Picea crassifolia also showed a significant correlation with precipitation from the current May and June on the Animaqing Mountains and Daqing Mountains in northwest China (Liang et al. 2006; Peng et al. 2007). Juniperus przewalskii of the northeastern Tibetan Plateau also responded significantly to precipitation from May to July during the current year because it plays an important role in the onset of xylogenesis, which affected xylem cell production (Ren et al. 2015; Zhang et al. 2015).
In our study, the correlations of growth with precipitation during the previous year (r = 0.496, P < 0.01 at 2649 m a.s.l.; r = 0.477, P < 0.01 at 2765 m a.s.l.) were higher than those in the current year (r = 0.453, P < 0.01 at 2649 m a.s.l.; r = 0.284, P < 0.05 at 2765 m a.s.l.) at the two lower altitudes. When we used partial correlation to remove the effect of the other factor in analyzing the relationship between radial growth and one precipitation factor above, we found similar results (Table 3). In other studies, radial growth of Picea crassifolia showed a higher correlation with total monthly precipitation in the previous growing season than in the current growing season at low altitudes in the Qilian Mountains (Chen et al. 2013; Liang et al. 2016). We could infer that influence of precipitation from the previous year was stronger than that in the current year, which was unexpected. A potential explanation is that the climate condition of the current year for radial growth was relatively suitable, so the role of stored carbohydrates and moisture conditions before the current growing season was highlighted instead. To affirm this inference and make an exact conclusion, deeper analysis should be performed by carrying out more physiological experiments.
Variation of climate–radial growth relationships along the altitude gradients
The most effective timescales of preseason cumulative precipitation varied with elevation and were the 1-month timescale of October at the highest site, 2-month timescale of October at the sub-higher site and 4-month timescale of October at the sub-lower and lowest site. In our study area, precipitation declines along with the decreasing altitude. The environment at the sub-lower and lowest site was drier than that at the two higher altitudes, so more accumulated precipitation may be needed to relieve water stress, increase soil moisture and put off the end of growing season effectively. The effective timescales of preseason cumulative precipitation also became longer with the decreasing altitudes. The influence degree of the preseason cumulative precipitation from the previous year declined with increasing elevation. The effect from total precipitation of the current May–July and mean temperature of the previous July–August also showed this trend. Furthermore, the significant influence of total precipitation during the current May–July diminished at the two higher altitudes. This phenomenon indicated that the drought that inhibited growth declined with rising altitudes because of increasing precipitation and decreasing temperature. Picea crassifolia below 3000 m a.s.l. seemed to be more limited by climate than that above 3000 m a.s.l.
The parameters of chronologies showed that the chronology above 3000 m a.s.l. had lower MS, SD, Rbar, PC1 and SNR values than that below 3000 m a.s.l., which was similar to a previous study at Sidalong Forest Farm (Gou et al. 2005). The study of Liang et al. (2010) also found this phenomenon in the Qilian Mountains. The parameters of chronology tended to decrease along with weakened stress from the environmental factors (Splechtna et al. 2000; Shao et al. 2009), so radial growth was less strongly affected by climate factors at higher altitudes than at lower altitudes. Tree-ring indexes of positive and negative pointer years above 3000 m a.s.l. were significantly lower and higher, respectively, than those at the two lower altitudes, which indicated that growth fluctuation was smaller. Lower growth variability above 3000 m a.s.l. also indicated that these trees were less limited by aridity than trees below 3000 m a.s.l. (Wang et al. 2005).
Both correlations between radial growth and climatic factors and characteristics of chronologies showed that radial growth of Picea crassifolia below 3000 m a.s.l. responded more to water deficit. This phenomenon was also observed in the whole Qilian Mountains when comparing other studies on Picea crassifolia, so this phenomenon was not unique to our study area but universal in the Qilian Mountains (Liang et al. 2009, 2010; Deng et al. 2013; Lu et al. 2015). Picea meyeri from the Luya Mountain in north-central China (Zhang et al. 2012) and Pinus koraiensis from the Changbai Mountain in northeastern China (Wang et al. 2013) were also more limited by drought at lower altitudes. This trend suggested that Picea crassifolia below 3000 m a.s.l in the Qilian Mountains could easily suffer from growth reduction and high mortality induced by drought under the context of global warming. Intensive protection and careful management should be undertaken for the Picea crassifolia forests at lower altitudes.
Conclusions
Variable relationships between radial growth and climate in previous and current growing seasons along altitude gradients for Picea crassifolia were clearly found through our dendrochronological analysis. Accumulated monthly precipitation before November during the previous year had lingering effects on radial growth in the current year. July could be seen as a demarcation point between climatic factors having an effect on growth in the current year and the next year at lower altitudes. Both the total precipitation during the previous July–October and total precipitation during the current May–July affected current radial growth below 3000 m a.s.l. Radial growth responded strongly to precipitation in the previous year than that in the current year. The response degree of radial growth to climate from previous and current years declined with increasing elevation. Radial growth below 3000 m a.s.l. was more sensitive to precipitation factors than that above 3000 m a.s.l.
Author contribution statement
Study conception and sampling design: L.Z. and Y.J. Field sampling: L.Z., S.Z., W.Z. and T.L. Laboratory experiment: L.Z., and S.Z. Analysis and interpretation of data: L.Z., X.K. and J.Y. Writing of manuscript: L.Z. and J.Y.
References
Affolter P, Buntgen U, Esper J, Rigling A, Weber P, Luterbacher J, Frank D (2010) Inner Alpine conifer response to 20th century drought swings. Eur J Forest Res 129:289–298. doi:10.1007/s10342-009-0327-x
Bond-Lamberty B, Rocha AV, Calvin K, Holmes B, Wang CK, Goulden ML (2014) Disturbance legacies and climate jointly drive tree growth and mortality in an intensively studied boreal forest. Glob Change Biol 20:216–227. doi:10.1111/gcb.12404
Cailleret M, Davi H (2011) Effects of climate on diameter growth of co-occurring Fagus sylvatica and Abies alba along an altitudinal gradient. Trees Struct Funct 25:265–276. doi:10.1007/s00468-010-0503-0
Chen L, Wu SH, Pan T (2011) Variability of climate-growth relationships along an elevation gradient in the Changbai Mountain, northeastern China. Trees Struct Funct 25:1133–1139. doi:10.1007/s00468-011-0588-0
Chen F, Yuan YJ, Wei WS, Yu SL, Fan ZA, Zhang RB, Zhang TW, Li Q, Shang HM (2012) Temperature reconstruction from tree-ring maximum latewood density of Qinghai spruce in middle Hexi Corridor, China. Theor Appl Climatol 107:633–643. doi:10.1007/s00704-011-0512-y
Chen F, Yuan YJ, Wei WS, Zhang RB, Yu SL, Shang HM, Zhang TW, Qin L, Wang HQ, Chen FH (2013) Tree-ring-based annual precipitation reconstruction for the Hexi Corridor, NW China: consequences for climate history on and beyond the mid-latitude Asian continent. Boreas 42:1008–1021. doi:10.1111/bor.12017
Cook ER (1985) A time series analysis approach to tree ring standardization. University of Arizona, Arizona
Deng Y, Gou XH, Gao LL, Zhao ZQ, Cao ZY, Yang MX (2013) Aridity changes in the eastern Qilian Mountains since AD 1856 reconstructed from tree-rings. Quatern Int 283:78–84. doi:10.1016/j.quaint.2012.04.039
Esper J, Frank DC, Wilson RJS, Buntgen U, Treydte K (2007) Uniform growth trends among central Asian low- and high-elevation juniper tree sites. Trees Struct Funct 21:141–150. doi:10.1007/s00468-006-0104-0
Ettl GJ, Peterson DL (1995) Growth response of subalpine fir (Abies lasiocarpa) to climate in the Olympic Mountains, Washington, USA. Glob Change Biol 1:213–230. doi:10.1111/j.1365-2486.1995.tb00023.x
Fritts H (2012) Tree rings and climate. Academic Press, London
Gou XH, Chen FH, Yang MX, Li JB, Peng JF, Jin LY (2005) Climatic response of thick leaf spruce (Picea crassifolia) tree-ring width at different elevations over Qilian Mountains, northwestern China. J Arid Environ 61:513–524. doi:10.1016/j.jaridenv.2004.09.011
Gruber A, Pirkebner D, Oberhuber W (2013) Seasonal dynamics of mobile carbohydrate pools in phloem and xylem of two alpine timberline conifers. Tree Physiol 33:1076–1083. doi:10.1093/treephys/tpt088
Helle G, Schleser GH (2004) Beyond CO2-fixation by Rubisco an interpretation of 13C/12C variations in tree rings from novel intra-seasonal studies on broad-leaf trees. Plant Cell Environ 27:367–380. doi:10.1111/j.0016-8025.2003.01159.x
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 43:69–78
Huang JG, Deslauriers A, Rossi S (2014) Xylem formation can be modeled statistically as a function of primary growth and cambium activity. New Phytol 203:831–841. doi:10.1111/nph.12859
Jiang Y, Zhang WT, Wang MC, Kang MY, Dong MY (2014) Radial growth of two dominant montane conifer tree species in response to climate change in North-Central China. PLoS One 9:e112537. doi:10.1371/journal.pone.0112537
Kozlowski TT (1992) Carbohydrate sources and sinks in woody-plants. Bot Rev 58:107–222. doi:10.1007/BF02858600
Kuptz D, Fleischmann F, Matyssek R, Grams TE (2011) Seasonal patterns of carbon allocation to respiratory pools in 60-yr-old deciduous (Fagus sylvatica) and evergreen (Picea abies) trees assessed via whole-tree stable carbon isotope labeling. New Phytol 191:160–172. doi:10.1111/j.1469-8137.2011.03676.x
Li DL, Liu DX (2000) Climate in Gansu. China Meteorological Press, Beijing (in Chinese)
Liang EY, Shao XM, Eckstein D, Huang L, Liu XH (2006) Topography- and species-dependent growth responses of Sabina przewalskii and Picea crassifolia to climate on the northeast Tibetan Plateau. Forest Ecol Manag 236:268–277. doi:10.1016/j.foreco.2006.09.016
Liang EY, Shao XM, Liu XH (2009) Annual precipitation variation inferred from tree rings since AD 1770 for the Western Qilian Mts, northern Tibetan Plateau. Tree Ring Res 65:95–103. doi:10.3959/2008-2.1
Liang E, Shao XM, Eckstein D, Liu XH (2010) Spatial variability of tree growth along a latitudinal transect in the Qilian Mountains, northeastern Tibetan Plateau. Can J For Res 40:200–211. doi:10.1139/X09-186
Liang E, Leuschner C, Dulamsuren C, Wagner B, Hauck M (2016) Global warming-related tree growth decline and mortality on the north-eastern Tibetan plateau. Clim Change 134:163–176. doi:10.1007/s10584-015-1531-y
Liu XC (1992) Picea crassifolia. Lanzhou University Press, Lanzhou (in Chinese)
Liu Y, Sun B, Song HM, Lei Y, Wang CY (2013) Tree-ring-based precipitation reconstruction for Mt. Xinglong, China, since AD 1679. Quatern Int 283:46–54. doi:10.1016/j.quaint.2012.03.045
Lu JX, Xu JM, Wu YQ, Li XJ, Evans R, Downes GM (2015) Climatic signals in wood property variables of Picea crassifolia. Wood Fiber Sci 47:131–140
Lupi C, Morin H, Deslauriers A, Rossi S (2010) Xylem phenology and wood production: resolving the chicken-or-egg dilemma. Plant Cell Environ 33:1721–1730. doi:10.1111/j.1365-3040.2010.02176.x
Peng JF, Gou XH, Chen FH, Liu PX, Zhang Y, Fang KY (2007) Characteristics of ring-width chronologies of Picea crassifolia and their responses to climate at different elevations in the Anyemaqen Mountains. Acta Ecol Sin 27:3268–3276
Rahman MR, Lateh H (2015) Spatio-temporal analysis of warming in Bangladesh using recent observed temperature data and GIS. Clim Dynam 46:2943–2960
Ren P, Rossi S, Gricar J, Liang EY, Cufar K (2015) Is precipitation a trigger for the onset of xylogenesis in Juniperus przewalskii on the north-eastern Tibetan Plateau? Ann Bot 115:629–639. doi:10.1093/aob/mcu259
Rinn F (2003) TSAPWin: time series analysis and presentation for dendrochronology and related applications. Heidelberg
Rossi S, Rathgeber CBK, Deslauriers A (2009) Comparing needle and shoot phenology with xylem development on three conifer species in Italy. Ann For Sci 66:206–213. doi:10.1051/forest/2008088
Savva Y, Oleksyn J, Reich PB, Tjoelker MG, Vaganov EA, Modrzynski J (2006) Interannual growth response of Norway spruce to climate along an altitudinal gradient in the Tatra Mountains, Poland. Trees Struct Funct 20:735–746. doi:10.1007/s00468-006-0088-9
Shao X, Wang S, Zhu H, Xu Y, Liang E, Yin Z, Xu X, Xiao Y (2009) A 3585-year ring-width dating chronology of Qilian juniper from the northeastern Qinghai–Tibetan Plateau. Iawa J 30:379–394. doi:10.1163/22941932-90000226
Sidor CG, Popa I, Vlad R, Cherubini P (2015) Different tree-ring responses of Norway spruce to air temperature across an altitudinal gradient in the Eastern Carpathians (Romania). Trees Struct Funct 29:985–997. doi:10.1007/s00468-015-1178-3
Splechtna BE, Dobrys J, Klinka K (2000) Tree-ring characteristics of subalpine fir (Abies lasiocarpa (Hook.) Nutt.) in relation to elevation and climatic fluctuations. Ann For Sci 57:89–100. doi:10.1051/forest:2000105
Toromani E, Sanxhaku M, Pasho E (2011) Growth responses to climate and drought in silver fir (Abies alba) along an altitudinal gradient in southern Kosovo. Can J For Res 41:1795–1807
Vicente-Serrano SM, Camarero JJ, Azorin-Molina C (2014) Diverse responses of forest growth to drought time-scales in the Northern Hemisphere. Global Ecol Biogeogr 23:1019–1030. doi:10.1111/geb.12183
Wang JY, Chang XX, Ge SG, Miao YX, Chang ZQ, Zhang H (2001) Vertical distribution of the vegetation and water and heat conditions of Qilian Mountain (northern slope). J Northwest For Univ 16:1–3 (in Chinese, with English abstract)
Wang T, Ren HB, Ma KP (2005) Climatic signals in tree ring of Picea schrenkiana along an altitudinal gradient in the central Tianshan Mountains, northwestern China. Trees Struct Funct 19:736–742. doi:10.1007/s00468-005-0003-9
Wang H, Shao XM, Jiang Y, Fang XQ, Wu SH (2013) The impacts of climate change on the radial growth of Pinus koraiensis along elevations of Changbai Mountain in northeastern China. For Ecol Manag 289:333–340. doi:10.1016/j.foreco.2012.10.023
Wigley TM, Briffa KR, Jones PD (1984) On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Clim Appl Meteorol 23:201–213. doi:10.1175/1520-0450(1984)023<0201:OTAVOC>2.0.CO;2
Xu JM, Lu JX, Bao FC, Evans R, Downes G, Huang RF, Zhao YK (2012) Cellulose microfibril angle variation in Picea crassifolia tree rings improves climate signals on the Tibetan plateau. Trees Struct Funct 26:1007–1016. doi:10.1007/s00468-012-0678-7
Xu JM, Lu JX, Evans R, Downes GM (2014) Relationship between ring width and tracheid characteristics in Picea crassifolia: implication in dendroclimatology. Bioresources 9:2203–2213. doi:10.15376/biores.9.2.2203-2213
Yang B, He MH, Melvin TM, Zhao Y, Briffa KR (2013) Climate control on tree growth at the upper and lower treelines: a case study in the Qilian Mountains, Tibetan Plateau. PloS One 8:e69065. doi:10.1371/journal.pone.0069065
Yang YT, Guan HD, Shen MG, Liang W, Jiang L (2015) Changes in autumn vegetation dormancy onset date and the climate controls across temperate ecosystems in China from 1982 to 2010. Glob Change Biol 21:652–665. doi:10.1111/gcb.12778
Zhang WT, Jiang Y, Dong MY, Kang MY, Yang HC (2012) Relationship between the radial growth of Picea meyeri and climate along elevations of the Luyashan Mountain in North-Central China. Forest Ecol Manag 265:142–149. doi:10.1016/j.foreco.2011.10.017
Zhang H, Shao XM, Zhang Y (2015) Which climatic factors limit radial growth of Qilian juniper at the upper treeline on the northeastern Tibetan Plateau? J Geogr Sci 25:1173–1182. doi:10.1007/s11442-015-1226-3
Zheng D (1996) The system of physico-geographical regions of the Qinghai–Xizang (Tibet) Plateau. Sci China (Ser D) 39:410–417 (in Chinese, with English abstract)
Acknowledgments
This work was funded by the National Natural Science Foundation of China (Projects No. 41630750). The authors express their thanks to the Foundation, as well as to the forest rangers at Sidalong forestry station for providing assistance in sampling.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by E. Liang.
Rights and permissions
About this article

Cite this article
Zhang, L., Jiang, Y., Zhao, S. et al. Lingering response of radial growth of Picea crassifolia to climate at different altitudes in the Qilian Mountains, Northwest China. Trees 31, 455–465 (2017). https://doi.org/10.1007/s00468-016-1467-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-016-1467-5