
Abstract
Key message
Cambial activity of ~100-year-old Qilian juniper trees initiated before the middle of May at an elevation of 3200 m a.s.l.; June and July were the main stem radial increment period. Around the middle of August, all xylem cell differentiation periods are completed. Precipitation or relative humidity is the main limiting factor for tree radial growth on the northeastern Tibetan Plateau.
Abstract
Previous studies have found that annual tree-ring width series of Qilian juniper (Sabina przewalskii Kom.) in the northeastern Tibetan Plateau (TP) are mostly moisture controlled, irrespective of site elevation. Knowing precisely the cambial growth dynamics during a vegetation period can lead to a better understanding of the climatic factors driving regional tree growth patterns. We observed wood formation with micro-cores taken at weekly intervals from late April to early October 2013, and monitored daily stem radial changes with high-resolution electronic point dendrometers in 30-min intervals. Dormant cambium contained 5–6 cell layers in the cambium zone. Cambial activity initiated before the middle of May in all the monitored trees. About three quarters of the total cell production or radial growth formed during June and July. Lignification of secondary walls of new xylem cells continued from end of May to middle of August. After the middle of August, the cambium entered into the inactive period. Dendrometer measurements confirmed that June and July were the main stem radial increment period. Nonparametric Kendall’s Tau correlations indicate that daily relative humidity and precipitation significantly influence stem growth in May. No obvious relationships were detected in June. In July, significant negative influences of maximum temperature and positive effects of relative humidity prevailed. Overall, growing season precipitation or associated relative humidity is the limiting factor for tree stem radial increments on the intra-annual scale. Our results present a thorough understanding of Qilian juniper xylogenesis and its climate forcing within the whole growing season on the northeastern TP.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
During the past decades, dendroclimatology has been developed thoroughly, aiming to establish long-term relationships between tree growth and climatic variables by examining correlations at monthly, seasonal, and annual resolution, and to reconstruct climate series predating meteorological periods (Fritts 1976). Based on calibration functions derived from seasonal averages of climate data, millennium-long records of climate change are meanwhile available for different regions (Briffa et al. 2013; Büntgen et al. 2011; Cook et al. 2010). Due to its location at the margins of the Asian summer monsoon region and to its ability to grow in climatic sensitive arid and semi-arid regions, Qilian juniper (Sabina przewalskii Kom.) plays a prominent role for the reconstruction of climate history in the northeastern Tibetan Plateau (TP) (Gou et al. 2014; Qin et al. 2013; Sheppard et al. 2004; Yang et al. 2014).
However, correlation functions between monthly means of climatic variables and annual tree-ring records do not consider the radial increment dynamics on an intra-annual scale. For a given tree, growth rates may vary considerably from year-to-year due to climate fluctuations. However, contrasting climatic conditions can yield similar amounts of annual growth depending on the timing of the climatic events during the growing season. On a yearly basis, the same amount of growth can be achieved by a faster growth rate or by a longer growing period. Moreover, the climate/growth relationships on an inter-annual time scale can sometimes be inconsistent with the climate/growth association on an intra-annual scale (Seo et al. 2011). Therefore, more species-specific and site-specific highly resolved data on radial increment dynamics are required both to better understand the association between tree growth processes and their responses to climate variability.
On the other hand, intra-annual cambium dynamics is one of the four emerging new topics in dendroclimatology and dendroecology (Eckstein and Schweingruber 2009). Detailed investigations of tree radial growth throughout the growing season are becoming more frequent in tree-ring studies in different climatic zones, especially in the Europe (Deslauriers et al. 2008; Gruber et al. 2010; King et al. 2013) and North America (Lupi et al. 2012; Rossi et al. 2011; Turcotte et al. 2009). Herein, high-resolution dendrometer measurements have been used to detect the seasonal growth patterns with a high level of precision (Biondi and Hartsough 2010; Deslauriers et al. 2007; Vitas 2011). Obviously, dendrometers have the advantage of providing continuous time series of high-resolution stem diameter variations, indicating water-related stem size fluctuations, cambial cell division, and enlargement of newly formed xylem and phloem. However, since dendrometers measure stem radius or circumference changes rather than wood formation, it is difficult to distinguish between true wood formation and hydrological swelling and shrinking (Mäkinen et al. 2003). Alternatively, wood formation can also be monitored by directly extracting wood samples at short time intervals (Rossi et al. 2008a, b, c; Saderi et al. 2013; Lenz et al. 2013). With the micro-coring technique (Bäucker et al. 1998), small wood cores are removed from a stem using an injection needle or a cutting tube. The state of wood formation at the time of coring can be determined microscopically from the cores without causing severe physiological impact on the tree. Currently, micro-coring represents one of the most reliable techniques for detailed monitoring of wood formation (Mäkinen et al. 2008; Oladi et al. 2011; Rossi et al. 2011). Despite that small increment cores are proved useful in assessing actual xylem formation, the method is laborious (Mäkinen et al. 2003) and requires sampling at short intervals during the main growing season, restricting its applicability in remote study areas. Consequently, dendrometers and micro-coring methods were used in combination to detect the cambium dynamics and wood formation (Bräuning et al. 2009; Camarero et al. 2010; Krepkowski et al. 2013). Studies like this have provided broad knowledge about which processes in the growing tree control cell production and which determine the characteristics of the produced cells, and thus are important for realizing tree growth–climate relationships on a finer time scale.
In China, however, limited researches using either the micro-coring method (Li et al. 2013; Liang et al. 2009; Ren et al. 2015) or dendrometer measurements (Jiang et al. 2014; Wang et al. 2012, 2014) to analyze xylem formation on the intra-annual scale have been conducted. Considering the vast diversity in tree species and forest ecosystems in China’s mountain areas stretching over different climatic zones, there exists large demand for further research to provide a comprehensive picture of tree growth dynamics in high altitude regions under different climate regimes.
The aim of this study is to monitor juniper tree radial formation by combining high-resolution dendrometer measurements and wood anatomy studies and to detect relationships with cambium activity-controlling climate factors in the semi-arid northeastern TP region.
Materials and methods
Study area
The study site is located at the Sidalong Forestry Station of the Qilian Mountains National Natural Reserve, on the northern slope of the middle Qilian Mountains in the northeastern Tibetan Plateau (38°26′N, 99°55′E, ~3200 m a.s.l.). According to the nearest meteorological station in Qilian (38°11′N, 100°15′E, 2787 m a.s.l.), the mean annual precipitation during the period 1951–2013 is 406 mm, about 90 % of which falls during the rainy season from May to September. The winter (December–February) mean temperature, mean minimum temperature and mean maximum temperature are −11.40, −18.89 and −1.15 °C, respectively, while those for the summer season (June–August) are 12.15, 5.70 and 20.18 °C, accordingly.
Micro-core sampling
During the year 2013, four isolated, about 100 years old, healthy Qilian juniper (Sabina przewalskii Kom.) trees were selected for analysis. The selected trees have 38.7 cm (standard deviation = 7.8 cm) of diameter and 4.21 m (standard deviation = 1.1 m) height. Trees with dead crowns, tilted stems probably containing reaction wood or evident biological deterioration or mechanical damages were avoided. Micro-core samples were taken at breast height (about 1.3 m) with a Trephor corer (Costruzioni Meccaniche Carabin C., Belluno, Italy; Rossi et al. 2006) in weekly intervals from late April to early October. All samples contained the preceding 3–4 tree rings and the developing annual wood layer including the cambial zone and adjacent phloem. Immediately after sampling, the micro-cores were placed in Eppendorf microtubes filled with absolute ethanol and stored as soon as possible at 5 °C to avoid tissue deterioration.
In the laboratory, each sample was oriented by marking the transversal side with a pencil under a stereo-microscope, and then embedded in paraffin. Cross sections of ~8 to 12 µm thickness were cut with a Leica RM 2245 rotary microtome (Leica, Wetzlar, Germany). Sections were stained with safranin (0.5 % in 95 % ethanol) and astra blue (0.5 % in 95 % ethanol), and photographed with a Leica microscope system. Main xylem differentiation phases were distinguished as enlargement, secondary wall thickening and lignifications. The number of cells in the growing tree ring of each tree was counted along three radial lines, then averaged and used to assess the overall timing of xylem growth. In spring, after at least one cell row can be observed in the enlarging phase in the post-cambial zone, xylem formation is considered to have begun. In the late summer, when cambium divisional activity has ceased and no cell is observed in the phase of cell enlargement, secondary wall thickening or lignification, xylem formation can be considered complete.
To minimize the differing cambial activity around the stem circumference and to eliminate the effect of the sampling position at the same tree (Van der Werf et al. 2007; Rossi et al. 2003; Swidrak et al. 2014), standardization was applied for correcting the cell number of each date. For each tree and each sample, we counted the cell numbers of the previous ring. The number of cells in each j-sample and by each i-phase was corrected as follows:
where
nc ij is the corrected number of cells, n ij is the counted number of cells, r is the number of j-samples, a j is the mean cell number of the previous ring for each j-sample and am is the mean cell number of the previous ring of all j-samples
Finally, the total number of cells for each sampling tree was modeled with the Gompertz equation using the nonlinear regression procedure included in the Origin software package (OriginLab Corporation, Northampton, MA, USA):
where y is the weekly cumulative sum of cells, t is the time computed in Julian days, A is the upper asymptote (maximum growth expressed as cell number), β is the x-axis placement parameter and k is the rate of change of the shape.
In addition, the related radial increment for each sampling date was measured with the ImageJ software (http://rsbweb.nih.gov/ij/). The same standardization method and Gompertz function model as mentioned above were used for the radial increment and cumulated radial increment.
Dendrometer data
To avoid influence of frequent hammering and wounding on dendrometer traces, different sample populations for dendrometer measurements and wood anatomical studies were selected within the same study site. Electronic point dendrometers (Ecomatik, Germany; type DR, accuracy ±2 μm) were installed at three trees (ML1, ML2 and ML3) before the initiation of the 2013 growing period. The dendrometers were attached to the stems at 1.3–1.5 m height in a perpendicular orientation to the slope. To reduce the influence of expansion and contraction processes of the bark, some outer parts of the bark were removed accordingly. Daily stem radial changes (mm) were registered automatically in 30-min intervals and saved in data loggers. According to the stem circadian cycle approach (Downes et al. 1999; Deslauriers et al. 2003), the typical diurnal cycle can be divided into three phases (Fig. 1): (a) contraction phase (CP), period between the daily maximum and daily minimum; (b) expansion phase (EP), from the daily minimum to the position observed at that morning’s maximum; (c) increment phase (IP), from when the stem radius exceeds the morning maximum until the subsequent maximum. Herein, the corresponding differences are determined as contraction amount, expansion amount and stem radial increment. According to these definitions, some days did not experience any stem radial increment phase, and those data were treated as missing values in the subsequent analyses (Downes et al. 1999). Daily stem radial variation was determined by calculating the difference between mean values of two consecutive days (Deslauriers et al. 2007), which represents a combination of water- and growth-induced radius expansion. All the amounts were compared among the three trees using analysis of variance (ANOVA).

Typical daily patterns of recorded dendrometer data during the main growing season. Max the daily maximum stem diameter, Min the daily minimum stem diameter, CP contraction phase, EP expansion phase, IP increment phase, SRI stem radial increment
Relationships with climate data
High-resolution (30-min interval) climate data were extracted from an automatic meteorological station (HOBO U30) installed within 150 m of the monitored dendrometer and micro-coring trees. Climate parameters measured include relative humidity (%), summed precipitation (mm), mean temperature (°C), mean maximum temperature (°C), and mean minimum temperature (°C). Herein, the mean maximum and minimum temperatures were extracted from the daily 48 temperature data measured during one diurnal circle. Since the precipitation values are not normally distributed, nonparametric Kendall’s Tau correlation coefficients, instead of Pearson correlations were used to assess the relationships between daily radial increments and water losses severity obtained from dendrometer data and climate factors.
Results and discussions
Microclimate during the growing season
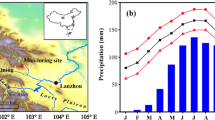
In the year 2013, monthly mean temperature (T mean), mean maximum temperature (T max) and mean minimum temperature (T min) from January to October were all higher than the mean climate conditions over the period 1957–2012 (calculated from the nearest meteorological station Qilian). Especially in March, monthly T max was around 5 °C above the long-term mean, resulting in a warmer pre-spring in the region (Fig. 2a–c). From May to August, the total precipitation in 2013 was 96 mm above the long-term mean, with July accounting for 32 % of this difference (Fig. 2d). Therefore, the pre-growing and growing seasons in the conducted experimental year can be identified as warmer and wetter compared with the long-term mean. Extracted from our installed automatic meteorological station, daily mean minimum, mean and mean maximum temperatures are 3.5, 8.8 and 15.6 °C, respectively, and the corresponding relative humidity and summed precipitation are 73 % and 440 mm from May to August at an elevation of 3200 m a.s.l. in the study region.

Local climate conditions at the study site in the year 2013 compared to the long-term mean of 1957–2012 at the nearby climate station Qilian. T mean monthly mean temperature, T max monthly mean maximum temperature, T min monthly mean minimum temperature, and Pre monthly precipitation
Xylogenesis
Applying the micro-coring method revealed the main differentiation periods of Qilian juniper radial growth (Fig. 3). The cambial zone (Fig. 3a) was characterized by thin cell walls and small radial diameters with typically 5–6 rectangular cell layers during the inactive period. After the initiation of cambial activity, the number of cambium cell layers increased and reached its peak with 7–10 cells, thereafter, the activity of the cambial zone declined gradually until the end of the growing season. When cell radial diameter was at least twice the size of cambial cells, cell enlargement (Fig. 3b) was considered to begin (Rossi et al. 2006). According to this definition, the onset of cambial activity in spring was first observed in one juniper tree on 3 May and in the other three trees on 12 May. During the enlargement period, the tracheids contained protoplast and had thin primary walls, whereas the number of cell layers was 1–5, showing an initial increase until a culmination around mid June, then a decreasing trend until the end of July. The beginning of secondary wall thickening (Fig. 3c) can be determined by means of polarized light microscopy. Polarized light does not pass through the irregularly arranged fibrils of primary cell walls, so that tracheids containing only primary wall appear dark whereas cells with secondary walls shine under the polarized light (Rossi et al. 2006). Accordingly, secondary wall thickening was first observed on 20 May and ended until early August. The beginning of cell wall lignification (Fig. 3d) can be determined when red staining by safranin is gradually replacing the blue staining of unlignified cell walls. Lignification (around 1 week after secondary wall thickening) was first observed on 26 May and lasted nearly 3 months until the middle of August. This phase of cell development normally started from the corners and outer parts of the cell walls and preceded towards the tracheid lumens. When the process of differentiation is completed, the walls of mature cells are completely red stained and the cell lumina are empty. Overall, the average number of cells during each differentiation phase is produced as bell-shaped patterns, whereas the accumulated cell numbers are associated with s-shaped curves across the period of tree-ring formation. Other softwood (e.g. Larix decidua, Pinus cembra, Picea abies and Pinus pinaster) and hardwood (e.g. Fagus orientalis, Quercus robur, Populus sp.) species also show similar trends of xylem production and differentiation during the growing season at various study sites (Rossi et al. 2006; Van der Werf et al. 2007; Deslauriers et al. 2009; Oladi et al. 2011; Vieira et al. 2014a,b).

Qilian juniper tree radial growth process on different sampling dates in the conducted year 2013. a Cell pattern in the cambial zone (CZ) on 27 April while cambium is still dormant; b enlarging (EL) cells on the date of 12 May; c mature cells (MC, red colored) and lignifying (LG) cells (gradual change from blue color to red) on 17 July under the polarized light. The upper dark cells consist of cambial and enlarging cells which do not have secondary wall while the birefringence of the remaining tracheid walls is the indicator of secondary wall thickening; d mature cells (MC, red colored) and lignifying (LG) cells (gradual change from blue color to red) on 17 July. PR previous ring
From initiation of cell enlargement to completion of lignification, variation among individual organisms is expected, not only because of differences of micro-environment, but also because of endogenous genetic factors. However, common growth patterns prevail. Xylem ring formation of all investigated ~100-year-old juniper trees started before the middle of May and continued to the middle of August, resulting in growth duration of 3 months in the observed year 2013. Herein, it is pointless to compare the whole growth process with the few currently available other studies conducted in North China in the year 2006 (Liang et al. 2009) or in the southeastern Tibetan Plateau (TP) in the year 2008 (Li et al. 2013), because of the differences in geographic location, tree species, site elevation, tree age and year-to-year variations of climatic conditions. We expect more thorough comparisons of the cambial phenology if the monitoring at our study site continues for several more years.
Although the amounts of cumulated and daily growth obtained from two different measuring processes, i.e. counting the cell number and measuring radial increment are very similar, they are not identical. The cell number and radial growth modeled with the Gompertz function are shown in Fig. 4. The averaged adjusted R 2 values of xylem growth fitted 93.1 % to the Gompertz function (p < 0.0001). The detailed percentages of cell numbers and radial increment formed during each month for the four sampled trees are presented in Table 1. Accordingly, about 79 % of the total cell production and 72 % of the annual radial growth were formed during June and July, constituting the main portion of the annual ring width in the study region. Consistently, Ren et al. (2015) also observed that >65 % tracheids of Qilian juniper were produced in June and July in the years 2009, 2010 and 2011 in the northeastern TP. Such related results provide useful information for the dendroclimatological reconstruction work conducted in the study region (e.g. Yang et al. 2014).

Qilian juniper growth modeled by Gompertz function from the micro-core sections of the four monitored trees. a Cumulated cell number, b daily produced cell number, c cumulated radial growth and d daily radial growth during the 2013 growth season
Dendrometer measurement and relationships with climate factors
Despite small individual variations recorded by the dendrometer data logger, as shown in Fig. 5, the daily stem radial increments (ANOVA, p = 0.48, F = 0.73) and daily stem radial variations (ANOVA, p = 0.54, F = 0.62) do not show significant differences among the three monitored trees, all present similar growth patterns throughout the measurement period. Typical diurnal patterns of stem shrinking and swelling were detected: the minimum stem radii occurred around the late afternoon or early evening, and the maximum stem radii were mostly found in the early morning. The stem circadian cycle generally lasts around 24 h, but changing soil moisture conditions caused by rain events may result in longer or shorter cycles. According to the dendrometer data, the main stem radius increment period can be identified from middle of May to the end of July or early August, coincident with the results obtained from the micro-coring method. However, the determination of important parameters, such as onset and cessation of the growing season and cambial growth rate, is difficult to derive from dendrometer measurements alone, since they provide time series related to stem water status and radial growth (Deslauriers et al. 2007; Chan et al. 2015). Nevertheless, dendrometers are the most direct way to obtain continuous time series that can be used for the detection of intra-annual tree stem radial growth variations and their relationships with climate (Belien et al. 2014; Oberhuber et al. 2014; Urrutia-Jalabert et al. 2015).

Cumulated daily stem radial change monitored by the three dendrometer trees and climate condition during the observed year 2013. T mean daily mean temperature, T max daily mean maximum temperature and T min daily mean minimum temperature. ML1, ML2 and ML3 the three monitored dendrometer trees
The calculated nonparametric Kendall’s Tau correlation coefficients (Table 2) between the averaged stem radial increments and climate factors indicate that May relative humidity and precipitation significantly affect daily radial increment. After the initiation of cambial activity, the increasing mean (5.1 °C) temperatures are sufficient for tree growth. However, the main rainy season has not yet began in May and precipitation is limited (59.6 mm), so the significant influence of relative humidity or precipitation and the unapparent effect of temperatures are rational. However, no significant correlations can be detected in June, which may imply that after the onset of the rainy season, both mean temperature (8.8 °C) and precipitation (127 mm) are appropriate for tree growth, without any clear limitation on growth rate. In July, the influence of relative humidity on radial increment reached the highest level, accompanied by a significant negative effect of maximum temperature. Herein, the monthly mean and mean maximum temperature of 10 and 16 °C probably lead to heat-induced drought stress on tree growth. On the one hand, more precipitation would directly promote tree ring formation, and on the other hand, more precipitation can also mediate heat-induced drought, both beneficial to tree growth. Furthermore, the nonparametric Kendall’s Tau correlations in Table 2 also evidenced that both relative humidity and maximum temperature significantly influence tree stem contraction or expansion amount. That is, the higher maximum temperature combined with less precipitation or lower relative humidity will result in more variability of water in the contraction and expansion periods. In total, all correlations indicate that local available water status is a major controlling factor for tree stem radial variations during the growing season in the study region.
Precipitation and relative humidity have been found to positively influence stem increment in different environments (Deslauriers et al. 2007; Duchesne and Houle 2011; Krepkowski et al. 2011; Köcher et al. 2012). In response to decreased precipitation or heat-induced drought effects during the vegetation period, trees may trigger an earlier stop of cambial activity (Vieira et al. 2014), declined cell division rate (Chaves et al. 2002), reduced cell size (Abe et al. 2003) or lumen area (Belien et al. 2012), or a bigger proportion of latewood tracheids (Pasho et al. 2012). Still, cambial activity and xylem cell development are considerable sinks of sucrose from photosynthesis (Oribe et al. 2003), the effects of drought thus can exacerbate the reduced wood production (Arend and Fromm 2007). Related studies indicated that under limited water availability, the non-structural carbohydrates in both cambium and xylem were significantly reduced during the growing season (Pantin et al. 2013; Deslauriers et al. 2014). It is generally considered that under slight water deficit, only cell expansion is physically inhibited by a decrease of cell turgor (Steppe et al. 2006; Köcher et al. 2012). If water deficit gets more severe, stress will affect the whole plant physiology, inhibiting cell metabolism and finally limiting growth (Rossi et al. 2008c). These relationships should be explored at shorter time scales by high-resolution monitoring, thus allowing higher levels of precision and more complete understanding of the roles of environmental factors on the mechanisms and components of wood formation.
Conclusions
This study is a systematical effort to combine wood anatomical analyses of thin sections obtained with regular micro-coring of the cambial zone, dendrometer measurements and automatic meteorological station data to analyze wood formation and stem radial variation, and their relationships with climate factors on the northeastern Tibetan Plateau. Results showed that in the experiment year 2013, the monitored Qilian juniper (Sabina przewalskii Kom.) cambial activity initiated before the middle of May. June and July were the main growth period, during which three quarters of the whole ring width were formed. At the end of July or early August, cell division stopped to leave enough time for the completion of cell formation before winter. All cell differentiation periods completed at the middle of August. Dendrometer data indicate that precipitation or related relative humidity are the main factors influencing the recorded stem radial changes on the intra-annual scale in the study region. Our current results provide detailed knowledge of Qilian juniper growth processes and contribute to a better understanding of the relationships between stem radial variation and climate factors across the full vegetation period. Qilian juniper growing in the study region has the potential to provide millennial, continuous and annually resolved paleoclimate records (Gou et al. 2014; Yang et al. 2014). Thus, to obtain a comprehensive understanding of growth reactions in the high-elevation forest ecosystems of the region, continued monitoring of wood formation during climatologically varying years is still needed.
Author contribution statement
Conceived and designed the experiments: BY, MHH. Performed the experiments: MHH, BY, ZYW, AB, KP and RO. Analyzed the data: MHH, BY, ZYW, AB, KP and RO. Contributed reagents/materials/analysis tools: MHH, BY, ZYW, AB, KP and RO. Wrote the paper: MHH, BY, ZYW, AB, KP and RO.
References
Abe H, Nakai T, Utsumi Y, Kagawa A (2003) Temporal water deficit and wood formation in Cryptomeria japonica. Tree Physiol 23:859–863
Arend M, Fromm J (2007) Seasonal change in the drought response of wood cell development in poplar. Tree Physiol 27:985–992
Bäucker E, Bues C, Vogel M (1998) Radial growth dynamics of spruce (Picea abies) measured by micro-cores. IAWA J 19:301–309
Belien E, Rossi S, Morin H, Deslauriers A (2012) Xylogenesis in black spruce subjected to rain exclusion in the field. Can J For Res 42:1306–1315
Belien E, Rossi S, Morin H, Deslauriers A (2014) High-resolution analysis of stem radius variations in black spruce [Picea mariana (Mill.) BSP] subjected to rain exclusion for three summers. Trees 28:1257–1265
Biondi F, Hartsough P (2010) Using automated point dendrometers to analyze tropical treeline stem growth at Nevado de Colima, Mexico. Sensors (Basel) 10:5827–5844
Bräuning A, Volland-Voigt F, Burchardt I, Ganzhi O, Nauss T et al (2009) Climatic control of radial growth of Cedrela montana in a humid mountain rain forest in southern Ecuador. Erdkunde 63:337–345
Briffa KR, Melvin TM, Osborn TJ, Hantemirov RM, Kirdyanov AV, Mazepa VS, Shiyatov SG, Esper J (2013) Reassessing the evidence for tree-growth and inferred temperature change during the Common Era in Yamalia, northwest Siberia. Quat Sci Rev 72:83–107
Büntgen U, Tegel W, Nicolussi K, McCormick M, Frank D, Trouet V, Kaplan JO, Herzig F, Heussner KU, Wanner H, Luterbacher J, Esper J (2011) 2500 years of European climate variability and human susceptibility. Science 331:578–582
Camarero JJ, Olano JM, Parras A (2010) Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol 185:471–480
Chan T, Holtta T, Berninger F, Makinen H, Nöjd P, Mencuccini M, Nikinmaa E (2015) Separating water-potential induced swelling and shrinking from measured radial stem variations reveals a cambial growth and osmotic concentration signal. Plant Cell Environ. doi:10.1111/pce.12541
Chaves MM, Pereira JS, Maroco J, Rodrigues ML, Ricardo CPP, Osorio ML, Carvalho I, Faria T, Pinheiro C (2002) How plants cope with water stress in the field? Photosynthesis and growth. Ann Bot 89:907–916
Cook ER, Anchukaitis KJ, Buckley BM, D’Arrigo RD, Jacoby GC, Wright WE (2010) Asian monsoon failure and megadrought during the last millennium. Science 328:486–489
Deslauriers A, Morin H, Urbinati C, Carrer M (2003) Daily weather response of balsam fir (Abies balsamea (L.) Mill.) stem radius increment from dendrometer analysis in the boreal forests of Québec (Canada). Trees 17:477–484
Deslauriers A, Rossi S, Anfodillo T (2007) Dendrometer and intra-annual tree growth: what kind of information can be inferred? Dendrochronologia 25:113–124
Deslauriers A, Rossi S, Anfodillo T, Saracino A (2008) Cambial phenology, wood formation and temperature thresholds in two contrasting years at high altitude in southern Italy. Tree Physiol 28:863–871
Deslauriers A, Giovannelli A, Rossi S, Castro G, Fragnelli G, Traversi L (2009) Intra-annual cambial activity and carbon availability in stem of poplar. Tree Physiol 29:1223–1235
Deslauriers A, Beaulieu M, Balducci L, Giovannelli A, Gagnon MJ, Rossi S (2014) Impact of warming and drought on carbon balance related to wood formation in black spruce. Ann Bot 114:335–345
Downes G, Beadle C, Worledge D (1999) Daily stem growth patterns in irrigated Eucalyptus globulus and E. nitens in relation to climate. Trees 14:102–111
Duchesne L, Houle D (2011) Modeling day-to-day stem diameter variation and annual growth of balsam fir (Abies balsamea (L.) Mill.) from daily climate. For Ecol and Manag 262:863–872
Eckstein D, Schweingruber F (2009) Dendrochronologia—a mirror for 25 years of tree-ring research and a sensor for promising topics. Dendrochronologia 27:7–13
Fritts HC (1976) Tree rings and climate. Academic Press, London
Gou XH, Deng Y, Gao LL, Chen FH, Cook ER, Yang MX, Zhang F (2014) Millennium tree-ring reconstruction of drought variability in the eastern Qilian Mountains, northwest China. Clim Dyn. doi:10.1007/s00382-014-2431-y
Gruber A, Strobl S, Veit B, Oberhuber W (2010) Impact of drought on the temporal dynamics of wood formation in Pinus sylvestris. Tree Physiol 30:490–501
Jiang Y, Wang BQ, Dong MY, Huang YM, Wang MC, Wang B (2014) Response of daily stem radial growth of Platycladus orientalis to environmental factors in a semi-arid area of North China. Trees 29(1):87–96
King G, Fonti P, Nievergelt D, Büntgen U, Frank D (2013) Climatic drivers of hourly to yearly tree radius variations along a 6°C natural warming gradient. Agr Forest Meteorol 168:36–46
Köcher P, Horna V, Leuschner C (2012) Environmental control of daily stem growth patterns in five temperate broad-leaved tree species. Tree Physiol 32:1021–1032
Krepkowski J, Bräuning A, Gebrekirstos A, Strobl S (2011) Cambial growth dynamics and climatic control of different tree life forms in tropical mountain forest in Ethiopia. Trees 25:59–70
Krepkowski J, Gebrekirstos A, Shibistova O, Brauning A (2013) Stable carbon isotope labeling reveals different carry-over effects between functional types of tropical trees in an Ethiopian mountain forest. New Phytol 199:441–451
Lenz A, Hoch G, Körner C (2013) Early season temperature controls cambial activity and total tree ring width at the alpine treeline. Plant Ecol Divers 6:365–375
Li X, Liang E, Gricar J, Prislan P, Rossi S, Čufar K (2013) Age dependence of xylogenesis and its climatic sensitivity in Smith fir on the south-eastern Tibetan Plateau. Tree Physiol 33:48–56
Liang EY, Eckstein D, Shao XM (2009) Seasonal cambial activity of relict Chinese pine at the northern limit of its natural distribution in north China-exploratory results. IAWA J 30:371–378
Lupi C, Morin H, Deslauriers A, Rossi S (2012) Xylogenesis in black spruce: does soil temperature matter? Tree Physiol 32:74–82
Mäkinen H, Nöjd P, Saranpää P (2003) Seasonal changes in stem radius and production of new tracheids in Norway spruce. Tree Physiol 23:959–968
Mäkinen H, Seo J-W, Nöjd P, Schmitt U, Jalkanen R (2008) Seasonal dynamics of wood formation: a comparison between pinning, microcoring and dendrometer measurements. Eur J Forest Res 127:235–245
Oberhuber W, Gruber A, Kofler W, Swidrak I (2014) Radial stem growth in response to microclimate and soil moisture in a drought-prone mixed coniferous forest at an inner Alpine site. Eur J For Res 133:467–479
Oladi R, Pourtahmasi K, Eckstein D, Bräuning A (2011) Seasonal dynamics of wood formation in Oriental beech (Fagus orientalis Lipsky) along an altitudinal gradient in the Hyrcanian forest. Iran. Trees 25:425–433
Oribe Y, Funada R, Kubo T (2003) Relationships between cambial activity, cell differentiation and the localization of starch in storage tissues around the cambium in locally heated stems of Abies sachalinensis (Schmidt) Masters. Trees 17:185–192
Pantin F, Fanciullino AL, Massonnet C, Dauzat M, Simonneau T, Muller B (2013) Buffering growth variations against water deficits through timely carbon usage. Front Plant Sci 4:483
Pasho E, Julio Camarero J, Vicente-Serrano SM (2012) Climatic impacts and drought control of radial growth and seasonal wood formation in Pinus halepensis. Trees 26:1875–1886
Qin C, Yang B, Melvin TM, Fan ZX, Zhao Y, Briffa KR (2013) Radial growth of Qilian Juniper on the northeast Tibetan Plateau and potential climate associations. PLoS One 8(11):e79362
Ren P, Rossi S, Gričar J, Liang EY, Čufar K (2015) Is precipitation a trigger of the onset of xylogenesis in Juniperus przewalskii on the northeastern Tibetan Plateau? Ann Bot 115(4):629–639
Rossi S, Deslauriers A, Morin H (2003) Application of the Gompertz equation for the study of xylem cell development. Dendrochronologia 21:33–39
Rossi S, Deslauriers A, Anfodillo T (2006) Assessment of cambial activity and xylogenesis by microsampling tree species: an example at the alpine timberline. IAWA J 27:383–394
Rossi S, Deslauriers A, Gričar J, Seo J-W, Rathgeber CBK, Anfodillo T, Morin H, Levanic T, Oven P, Jalkanen R (2008a) Critical temperatures for xylogenesis in conifers of cold climates. Global Ecol Biogeogr 17:696–707
Rossi S, Deslauriers A, Anfodillo T, Carrer M (2008b) Age-dependent xylogenesis in timberline conifers. New Phytol 177:199–208
Rossi S, Simard S, Rathgeber CBK, Deslauriers A, Zan C (2008c) Effects of a 20-day-long dry period on cambial and apical meristem growth in Abies balsamea seedlings. Trees 23:85–93
Rossi S, Morin H, Deslauriers A, Plourde P-Y (2011) Predicting xylem phenology in black spruce under climate warming. Global Change Biol 17:614–625
Saderi SM, Pourtahmasi K, Oladi R, Rathgeber CBK (2013) Wood formation in Juniperus excelsa ssp. polycarpos in the high mountains of north-east Iran. J Trop For Sci 25:421–428
Seo JW, Eckstein D, Jalkanen R, Schmitt U (2011) Climatic control of intra- and inter-annual wood-formation dynamics of Scots pine in northern Finland. Environ Exp Bot 72:422–431
Sheppard PR, Tarasov PE, Graumlich LJ, Heussner KU, Wagner M, Österle H, Thompson LG (2004) Annual precipitation since 515 BC reconstructed from living and fossil juniper growth of northeastern Qinghai Province, China. Clim Dyn 23(7–8):869–881
Steppe K, De Pauw DJ, Lemeur R, Vanrolleghem PA (2006) A mathematical model linking tree sap flow dynamics to daily stem diameter fluctuations and radial stem growth. Tree Physiol 26:257–273
Swidrak I, Gruber A, Oberhuber W (2014) Xylem and phloem phenology in co-occurring conifers exposed to drought. Trees 28(4):1161–1171
Turcotte A, Morin H, Krause C, Deslauriers A, Thibeault-Martel M (2009) The timing of spring rehydration and its relation with the onset of wood formation in black spruce. Agr Forest Meteorol 149:1403–1409
Urrutia-Jalabert R, Rossi S, Deslauriers A, Malhi Y, Lara A (2015) Environmental correlates of stem radius change in the endangered Fitzroya cupressoides forests of southern Chile. Agr Forest Meteorol 200:209–221
Van der Werf G, Sass-Klaassen UG, Mohren G (2007) The impact of the 2003 summer drought on the intra-annual growth pattern of beech (Fagus sylvatica L.) and oak (Quercus robur L.) on a dry site in the Netherlands. Dendrochronologia 25:103–112
Vieira J, Rossi S, Campelo F, Freitas H, Nabais C (2014) Xylogenesis of Pinus pinaster under a Mediterranean climate. Ann For Sci 71:71–80
Vitas A (2011) Seasonal growth variations of pine, spruce, and birch recorded by band dendrometers in NE Lithuania. Balt For 17:197–204
Wang ZY, Yang B, Deslauriers A, Qin C, He MH, Shi F, Liu JJ (2012) Two phases of seasonal stem radius variations of Sabina przewalskii Kom. in northwestern China inferred from sub-diurnal shrinkage and expansion patterns. Trees 26:1747–1757
Wang ZY, Yang B, Deslauriers A, Bräuning A (2014) Intra-annual stem radial increment response of Qilian juniper to temperature and precipitation along an altitudinal gradient in northwestern China. Trees 29(1):25–34
Yang B, Qin C, Wang JL, He MH, Melvin TM, Osborn TJ, Briffa KR (2014) A 3,500-year tree-ring record of annual precipitation on the northeastern Tibetan Plateau. PNAS 111:2903–2908
Acknowledgments
All necessary permits were obtained for the described field studies from the Administration of Gansu Qilian Shan National Nature Reserve. The study was jointly funded by the National Science Foundation of China (Grant No. 41325008, 31300412) and the Interdisciplinary Innovation Team project of the Chinese Academy of Sciences (29Y329B91). The research was also supported by Chinese Academy of Sciences visiting fellowship for researchers from developing countries (2014FFZA0006) and the CAS/SAFEA International Partnership Program for Creative Research Teams. The authors would like to thank Jie Wang, Jianqi Zhang, Hongyuan Ge, Wen Wang and Hongyi Wang for the maintenance of the field instruments.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
All the authors declare that they have no conflict of interest.
Additional information
Communicated by E. Priesack.
Rights and permissions
About this article

Cite this article
He, M., Yang, B., Wang, Z. et al. Climatic forcing of xylem formation in Qilian juniper on the northeastern Tibetan Plateau. Trees 30, 923–933 (2016). https://doi.org/10.1007/s00468-015-1333-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-015-1333-x