
Abstract
To accelerate extensive application of biological manganese removal technology, a pilot-scale biofilter for ammonia, iron and manganese removal was constructed to investigate the removal performance and microbial community profiles at different manganese concentrations. When manganese in influent increased from 1 to 10 mg/L, the pollutants were completely removed. Ammonia and iron was slightly changed along the filter depth, while manganese obviously increased. In 0 m of the filter depth, the abundance of Gallionella (iron oxidizing bacteria, IOB) increased, while Crenothrix (IOB) decreased. The abundance of Gallionella (manganese oxidizing bacteria, MnOB) in 0.4 and 0.8 m increased to 16.82% and 12.37%, respectively; and Crenothrix (MnOB) in 0.8 m increased to 19.95%, but decreased to 25.08% in 0.4 m. The abundance of ammonia oxidizing bacteria (AOB, Nitrosococcus) decreased in 0.4 and 0.8 m. The biofilter presented a high ability to remove manganese, and had a broad application prospect.
Graphical abstract

Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
When groundwater flows through rocks, soils and minerals, soluble ferrous iron and bivalent manganese are often observed in groundwater [1], since groundwater is usually devoid of dissolved oxygen (DO) and mildly acidic [2]. Iron and manganese existed in drinking water with high concentrations could result in esthetic and operational problems [3]. For example, (1) iron and manganese oxides stain on laundry and sanitary facilities [1, 4]; (2) iron and manganese lead to an unpleasant metallic taste of water [2]; (3) iron and manganese deposits build up in pipelines, and then reduce pipe diameter [5]. Ammonia associated with ferrous iron and bivalent manganese is widely existed in groundwater with concentrations above the permitted limits [6]. Furthermore, the concentration of ammonia continuously increased in the past years [7]. Ammonia existed in drinking water could have an adverse effect on the chlorination process due to the formation of disinfection by-products with chlorine [8,9,10], which would reduce disinfection efficiency [8], deteriorate taste and odor of water and damage human nervous system [11]. Moreover, ammonia would consume excessive oxygen during nitrification in the manganese biofiltration process, resulted in earthy and mouldy tasting water [11]. Therefore, iron, manganese and ammonia presented in drinking water should be limited and the maximum contaminant levels (MCLs) for total iron of 0.3 mg/L, manganese of 0.1 mg/L and ammonia of 0.5 mg/L have been established in China (GB 5749-2006).
Biological removal of ammonia, iron and manganese is preferable compared with chemical methods, since it is not necessary to add extra chemicals which may introduce other pollutants into the produced water and produce potential hazardous by-products [12], and the volume of generated sludge is appreciably smaller and easier to handle [13]. As demonstrated, the pollutants could be removed simultaneously in a single biological system [14, 15]. In the biological system, ammonia was oxidized by ammonia oxidizing bacteria (AOB) and nitrite oxidizing bacteria (NOB). Several types of bacteria have been confirmed as AOB or NOB, such as Nitrosomonas mobilis, Comamonas, Nitrosomonas europaea, Nitrospira, Nitrosomonas halophila and Acinetobacter [3, 15]. Iron and manganese were oxidized by iron oxidizing bacteria (IOB) and manganese oxidizing bacteria (MnOB), respectively. Up to now, several types of bacteria have been confirmed as IOB (Crenothrix, Leptothrix, Bacillus and Gallionella [3, 16]), and MnOB (Leptothrix, Metallenium, Pseudomonas, Crenothrix, Siderocapsa, Hyphomicrobium and Gallionella [15, 17,18,19]).
The concentration of manganese in the groundwaters is greatly varied among different regions, and the presence of manganese often accompanies with iron and ammonia. When the pollutants were biologically oxidized by AOB, IOB and MnOB in biological systems, there must be a link between the removal efficiencies and the microbial community profiles. Therefore, it is necessary to investigate the relationship between the microbial community structure and the removal efficiencies at different manganese concentration to optimize the biological manganese removal technology. Although several researchers investigated the influence of manganese on ammonia, iron and manganese removal, few studies focus on the relationship between the microbial community structure and the removal efficiencies when manganese varied. Bacteria contamination should be prevented when biological treatment was applied. Therefore, biological treatment in this experiment was used as pre-treatment, and membrane treatment was advised followed.
In this study, a pilot-scale biofilter system was constructed for ammonia, iron and manganese removal from real groundwater. The biofilter was operated approximately 180 day to evaluate the removal efficiency of ammonia, iron and manganese, and the microbial community structures at different manganese concentrations were analyzed and compared using 454 HTP. The work herein aimed to gain a deep insight into the relationship between the removal efficiency of ammonia, iron and manganese and the microbial community structure to generalize biological manganese removal technology and provide guidance for the optimization of biofilter.
Materials and methods
Description of biofilter system
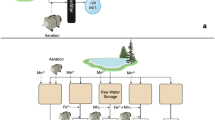
A pilot-scale biofilter system employed to simultaneously remove ammonia, iron and manganese was developed in a groundwater treatment plant (GWTP) located in Harbin city, P. R. China (Fig. 1). The biofilter consisted of a plexiglass column with a height of 3000 mm and an inner diameter of 150 mm. A mixing chamber used to mix the incoming waters was set at the top of the biofilter, and a perforated plate used to support the media and collect the treated water was located at the bottom. Three media sampling ports at 0, 400 and 800 mm and seventeen water sampling ports at 100 mm intervals were distributed along the height of the column from media top to bottom. The aerated raw groundwater obtained from the GWTP was stored in Tank 1 (volume 1000 L), and the DO concentration was about 8 mg/L. Tank 2 (volume 50 L) was used to collect stock solution of 10 g/L Mn(II) prepared by diluting MnSO4·H2O.

Schematic drawing of the pilot-scale biofilter system
Two kinds of support materials were packed in the biofilter, and the upper part of the media (300 mm) was columnar anthracite with a height of 5 mm and a mean diameter of 1 mm, while the lower part (1200 mm) was manganese sand with a mean diameter of 0.8–1 mm. Down-flow pattern was adopted, and the flow rate was controlled at the entry point [6]. The biofilter was operated about 1 year before this experiment, and the system was backwashed about every 2 days by water according to the water head loss. The backwashing time was 5 min and backwashing intensity was 8–10 L/(s·m2). In this experiment, real groundwater extracted from the wells with a depth of 40–50 m, in Harbin city, P. R. China, was used. The concentration of ammonia, total iron and manganese in the aerated raw groundwater was about 0.9–1.4, 7–10 and 0.9–1.4 mg/L, respectively, and the temperature was about 8 °C (Tab. S1).
Removal efficiency of ammonia, iron and manganese with increased manganese concentration
The water in tank 1 was aerated and the concentration of DO increased to about 11 mg/L. The aerated water in tank 1 was pumped into the biofilter to investigate the removal efficiency of ammonia, iron and manganese (phase I). About 1 month later, the aerated water in tank 1 and the stock solution of Mn(II) in tank 2 were pumped into the biofilter with a suitable proportion to ensure the manganese concentration in influent was about 5 mg/L (phase II). After 2 months, the concentration of manganese in influent increased to about 10 mg/L in the same way (phase III). In addition, the flow rate of the biofilter was fixed at 4 m/h in this experiment because of the high iron concentration in influent.
Sampling and chemical analysis
Analysis of DO, oxidation–reduction potential (ORP) and pH were conducted using a DO meter (Oxi 315i-WTW), an ORP meter (pH 315i-WTW) and a pH meter (pH 315i-WTW), respectively. Concentration of ammonia, iron and manganese was measured by photometric method according to Standard Methods for the Examination of Water and Wastewater [20]. The filter media (manganese sand or anthracite) were collected at 0, 400 and 800 mm of the filter bed, and stored in 50-mL sterile plastic test tubes at − 80 °C for further analysis. Furthermore, the microorganisms in mature media from the biofilter (day 158) were examined by scanning electron microscopy (SEM, JSM-6480LV) according to previous studies [18].
Microbial diversity analysis
DNA extraction and polymerase chain reaction (PCR) amplification
Media samples from 0, 400 and 800 mm of the filter layer were explored to deeply understand the microbial community structure evolution at different phases. These samples were analyzed by 454 HTP at days 31 (A1, A2 and A3), 93 (A4, A5 and A6) and 158 (A7, A8 and A9). DNA was extracted from the media samples by Powersoil DNA Isolation Kit (MoBio Laboratories Inc, USA). The quality of the DNA was examined by 1% (w/v) agarose gel electrophoresis and the concentration was measured with a UV–Vis spectrophotometer (NanoDrop 2000, USA) [3]. The V3–V4 region of the 16S rRNA gene was amplified using bacterial primers 515F (5′-GTG CCA GCM GCC GCG GTA A-3′) and 806R (5′-GGA CTA CHV GGG TWT CTA AT-3′). PCR amplifications were carried out in a GeneAmp 9700 thermocycler (ABI, USA) according to previous research [21].
454 high-throughput 16S rRNA gene pyrosequencing
Sequencing of the DNA samples was carried out using the same method as previous research [21]. Raw sequence data of this study were deposited to the NCBI Sequence Read Archive with accession No. SRP108227.
Biodiversity analysis and phylogenetic classification
Low quality reads (ambiguous nucleotides and quality value < 20) were removed from the raw sequence data as described in [22]. The paired-end reads from the samples were overlapped to assembly V3–V4 tags of 16S rRNA gene using SeqPrep (https://github.com/jstjohn/SeqPrep), and chimera sequences from the tags were removed by usearch6. Eventually, the numbers of high quality sequences were 11, 640 (A1), 11, 961 (A2), 11, 023 (A3), 5, 737 (A4), 10, 208 (A5), 8, 853 (A6), 10, 990 (A7), 16, 015 (A8) and 4, 110 (A9) with an average length of 291 bp. The sequences were processed using the same method as previous research [21].
Results
Overall performance of the biofilter and SEM images
When the concentration of manganese in influent was about 1 (Phase I), 5 (Phase II) and 10 mg/L (Phase III), nearly all of manganese, ammonia and iron were removed in steady phase, and the concentration of ammonia, total iron and manganese in effluent was below 0.1, 0.1 and 0.05 mg/L, respectively (Fig. 2). When ammonia, total iron and manganese in influent were 1.21, 8.33, and 1.12 mg/L in the 31st day (Table 1), respectively, ammonia quickly decreased to 0.39 and 0.10 mg/L in 0.2 and 0.8 m of the filter depth, respectively; and total iron sharply decreased to 0.16 mg/L in 0.2 m, and then decreased to 0.058 mg/L in 0.4 m. While manganese hardly removed in 0–0.2 m, quickly decreased in 0.2–0.4 m, and decreased to 0.023 mg/L in 1.2 m. When manganese in influent increased to about 5 mg/L in the 37th day, ammonia and iron removal were hardly affected, surprisingly, manganese in effluent was also below 0.05 mg/L. In the 93rd day, the concentration of ammonia and total iron in 0.2 m was 0.37 and 0.18 mg/L, respectively, which was slightly changed compared with phase I. However, manganese obviously increased to 1.89 mg/L in 0.4 m, and decreased to 0.026 mg/L in 1.5 m. Ammonia and iron removal were hardly affected too, as manganese increased to about 10 mg/L in the 105th day. However, manganese in effluent increased to 0.26 mg/L in the 110th day, and then increased to 1.02 mg/L in the 112th day, finally decreased to 0.0098 mg/L in the 120th day. In the 158th day, ammonia and total iron concentration along the filter depth was still hardly varied; while manganese concentration obviously increased in 0.2–1.2 m, and decreased to 0.046 mg/L in 1.5 m. In the biofilter, ammonia was biologically oxidized to nitrite with a concentration of about 0.8 mg/L. Ferrous iron was oxidized to iron oxy-hydroxides, and oxidation form was amorphous ferrihydrite according to the previous reference [23]. While manganese was biologically oxidized to MnOx (x is between 3 and 4) [23]. Furthermore, pH and ORP in influent was 7.17–7.34 and 16–62 mV, and pH in effluent decreased to 6.98–7.13, while ORP increased to 204–287 mV.

Removal efficiency of ammonia (a), iron (b) and manganese (c) in different phases
The removal kinetics of ammonia and manganese was investigated to obtain the required time to efficiently remove ammonia and manganese. Iron was mainly removed in 0–0.2 m of the filter depth, and ferrous iron was removed through chemical and biological oxidation; hence the removal kinetics of iron was not investigated in this experiment. Kinetics of biological ammonia and manganese oxidation in days 31, 93 and 158 were investigated, respectively. The results have expressed as ln[(NH4+–N)t/(NH4+–N)o] and ln[(Mn2+)t/(Mn2+)o] versus the empty bed contacted time (EBCT) (Figs. 3, 4). For a determined DO constant and a constant pH value, the ammonia and manganese depletion rate would be first order, i.e.,

Ammonia concentration profiles along the filter depth in different phases (a), linear regression analysis of ammonia depletion in relation with the empty bed contact time (b)

Manganese concentration profiles along the filter depth in different phases (a), linear regression analysis of manganese depletion in relation with the empty bed contact time (b)
The kinetic constant of biological ammonia oxidation was found equal to 0.0738, 0.0781 and 0.0770 min−1 in days 31, 93 and 158, respectively, and the half-life constant was calculated to 9.394, 8.876 and 9.001 min, respectively. Figure 4 indicated that the ln[(Mn2+)t/(Mn2+)o] versus time (EBCT) was linear in days 31 and 93. The value of k was 0.115 and 0.119 min−1, and the half-life time for the depletion of manganese was 6.042 and 5.852 min, respectively. But the ln[(Mn2+)t/(Mn2+)o] versus time (EBCT) was not linear in day 158, meaning the biological manganese oxidation did not followed a first order kinetic rate in that condition.
SEM images of the mature media were examined to verify the existence and growth of bacteria in the biofilter. Plenty of microorganisms covered by large amounts of iron and manganese oxides were observed in the micrograph, which demonstrated the catalytic role of bacteria in removal of iron and manganese in the biofilter (Fig. S1). Two species of microorganisms, which had a very typical structure of twisted stalk and a rod-shaped, were found in the biofliter.
Richness and diversity of microbial community
The parameters related to the alpha diversity of microbial community for the nine samples at distance cutoff level of 0.03 were shown in Table 2. 1, 091 (A1), 1, 188 (A2), 1, 105 (A3), 818 (A4), 1, 040 (A5), 1, 091 (A6), 1, 119 (A7), 1, 367 (A8) and 706 (A9) OTUs were clustered by performing the alignment at a uniform length of 291 bp. The total OTUs number estimated by Chao1 estimator were 1, 946 (A1), 2, 572 (A2), 2, 034 (A3), 1, 526 (A4), 2, 002 (A5), 1, 824 (A6), 2, 224 (A7), 2, 329 (A8) and 1, 272 (A9) with infinite sampling, indicating A9 had the lowest richness while A2 had the greatest richness. The OTUs number of the nine samples revealed that the microbial communities were distinctly different in different filter beds at different phases. The Shannon diversity index provides the simply species richness and how the abundance of each species is distributed among all the species in the community [24]. A1 presented the lowest diversity (Shannon = 7.06) in the nine communities, while A6 had the highest diversity (Shannon = 8.04).
Taxonomic complexity of the bacterial community
The phylogenetic diversity of the bacterial communities from the samples at phylum level is shown in Fig. 5. An extremely high diversity was showed in the nine samples, reflected in the fact that 29 (A1), 28 (A2), 27 (A3), 19 (A4), 32 (A5), 31 (A6), 29 (A7), 34 (A8) and 25 (A9) identified bacterial phyla were detected, and 38 identified bacterial phyla and 2 identified archaea phyla were observed in total. Even so, 1.31% (A1), 1.18% (A2), 2.91% (A3), 1.95% (A4), 3.12% (A5), 3.70% (A6), 1.65% (A7), 4.52% (A8) and 4.95% (A9) of the total reads in each sample were not classified, indicating that those bacteria were unknown. Proteobacteria which existed widely in biofilters for iron and manganese removal was the dominant bacterial phyla [23], accounted for 76.32% (A1), 78.20% (A2), 67.99% (A3), 81.54% (A4), 77.31% (A5), 64.77% (A6), 79.31% (A7), 74.66% (A8) and 51.48% (A9) of the total reads. Nitrospirae involved in nitrification was existed in all nine communities [25], and the abundance of Nitrospirae was highest in A3 (8.61%).

Taxonomic classification of pyrosequences from bacterial communities of the nine samples at the phylum levels. Phyla with relative low abundance in all nine libraries were classified as “others”
At the class level, pyrosequencing in the nine samples detected 92 bacterial classes and 4 archaea classes, and 62 (A1), 58 (A2), 63(A3), 37 (A4), 61 (A5), 65 (A6), 69 (A7), 72 (A8) and 57 (A9) bacterial classes were detected in each sample (Fig. 6). However, 1.78% (A1), 1.59% (A2), 3.84% (A3), 2.35% (A4), 3.96% (A5), 7.18% (A6), 2.18% (A7), 5.53% (A8) and 9.28% (A9) of the total reads in each sample were not classified. Betaproteobacteria and Gammaproteobacteria were the primary bacterial community in the nine communities. The abundance of Alphaproteobacteria, Deltaproteobacteria and Flavobacteriia was relatively high in the nine communities.

Taxonomic classification of pyrosequences from bacterial communities of the nine samples at the class levels. Classes with relative low abundance in all nine libraries were classified as “others”
At the genus level, the dominant population in the nine communities except A3 was Crenothrix (Fig. 7), which could oxidize both iron and manganese. While the dominant population in A3 was Achromobacter, which could oxidize organic matter [26]. AOB (Nitrosomonas, Nitrosococcus and Nitrosovibrio) and NOB (Nitrospira) which presented in the biofilter demonstrated that nitritification and nitratification occurred to remove ammonia and nitrite, respectively [25, 27]. IOB (Crenothrix, Bacillus, Pseudomonas, Leptospirillum and Gallionella) and MnOB (Crenothrix, Bacillus, Gallionella, Arthrobacter, Pseudomonas and Hyphomicrobium) were also found in the biofilter. Besides, kinds of other genera were also found in the biofilter because of the complex composition of the real groundwater, such as Burkholderia, Paludibacter, Novosphingobium and Flavobacterium may be organic matter oxidizing bacteria, Methylotenera and Methylomonas may be methane oxidizers, and Sulfuritalea may oxidize sulfur.

Taxonomic classification of pyrosequences from bacterial communities of the nine samples at the genus levels. Genera with relative low abundance in all nine libraries were classified as “others”
In phase I (day 31), Crenothrix and Gallionella which could oxidize iron and manganese were found in 0, 0.4 and 0.8 m of the biofilter, and manganese removal was occurred after ferrous iron was oxidized completely due to the fact that ferrous iron could react with manganese oxides [6]. Therefore, iron was removed in 0 m by Crenothrix and Gallionella, and manganese was removed in 0.4 and 0.8 m since ferrous iron in 0.4 m was not detected. The high abundance of Crenothrix and Gallionella in 0 m reflected in the fact that an excellent performance for iron removal in 0–0.2 m was achieved. The abundance of Crenothrix and Gallionella decreased from 30.77% and 8.16% (0.4 m) to 14.78% and 3.28% (0.8 m), respectively, which was consistent with the concentration of manganese that decreased from 0.39 mg/L (0.4 m) to 0.11 mg/L (0.8 m). The abundance of Nitrosococcus increased from 1.51% (A1) to 2.46% (A2) and 8.61% (A3) along the filter depth, while ammonia decreased from 1.21 mg/L (0 m) to 0.28 (0.4 m) and 0.10 mg/L (0.8 m).
In phase II (day 93), the concentration of manganese in influent increased to 4.71 mg/L. The removal efficiency of iron was slightly changed, but the abundance of IOB-Crenothrix and Gallionella was affected. The abundance of Crenothrix in 0 m of the filter depth decreased from 31.75% (A1) to 29.33% (A4), while Gallionella increased from 9.77% (A1) to 10.51% (A4). The concentration of manganese in 0.4 and 0.8 m increased from 0.39 to 0.11 mg/L to 1.89 and 0.68 mg/L, respectively; thus the abundance of MnOB-Gallionella in 0.4 and 0.8 m obviously increased from 8.16% (A2) and 3.28% (A3) to 12.13% (A5) and 5.42% (A6), respectively. And the abundance of Crenothrix in 0.8 m increased from 14.73% (A3) to 16.61% (A6), surprisingly Crenothrix in 0.4 m decreased from 30.77% (A2) to 28.37% (A5). The concentration of ammonia in 0–0.8 m in phase II was slightly varied compared with phase I, but the abundance of AOB-Nitrosococcus in 0.4 and 0.8 m obviously decreased from 2.46% (A2) and 8.61% (A3) to 2.10% (A5) and 3.22% (A6).
In phase III (day 158), the concentration of manganese in influent increased to 10.21 mg/L. The removal efficiency of iron was hardly affected; but the abundance of Crenothrix in 0 m of the filter depth continuously decreased from 31.75% (A1) and 29.33% (A4) to 27.99% (A7), while the abundance of Gallionella continuously increased from 9.77% (A1) and 10.51% (A4) to 12.63% (A7). The concentration of manganese in 0.4 and 0.8 m obviously increased to 6.90 and 4.31 mg/L in phase III, respectively. The growth rate of MnOB-Gallionella was faster than that of Crenothrix in the biofilter, therefore, the abundance of Gallionella in 0.4 and 0.8 m obviously increased from 12.13% and 5.42% (phase II) to 16.82% and 12.37% (phase III), respectively. The abundance of Crenothrix in 0.8 m increased from 16.61% (phase II) to 19.95% (phase III), but Crenothrix in 0.4 m decreased from 28.37% (phase II) to 25.08% (phase III). The abundance of AOB-Nitrosococcus in 0.8 m decreased from 3.22% (A6) to 2.14% (A9), but increased from 2.10 to 2.49% (A8) in 0.4 m.
Composition of the archaea community
The numbers of reads of the archaea 16S rRNA gene accounted for 0.32% (A1), 0.33% (A2), 0.29% (A3), 0.40% (A4), 0.42% (A5), 0.39% (A6), 0.45% (A7), 0.46% (A8) and 0.40% (A9) of the total 16S rRNA genes. Pyrosequencing detected two archaea phyla-Euryarchaeota and [Parvarchaeota], and four archaea classes-Thaumarchaeota, Methanobacteria, Methanomicrobia and [Parvarchaea] in all nine communities (Fig. 8), and the dominant phylum and class were Euryarchaeota and Thermoplasmata, respectively. The dominant genera in the nine communities were Ferroplasma (40.00%, 46.88%, 32.14%, 50.00%, 38.89%, 13.79%, 36.59%, 37.10%, and 28.57%) and Thermogymnomonas (30.00%, 31.25%, 25.00%, 33.33%, 36.11%, 31.03%, 31.71%, 38.71%, and 42.86%) (Fig. 9). Additionally, Ferroplasma was iron oxidizing archaea (IOA) [28], and Methanosarcina, Methanoculleus, Methanospirillum, Candidatus Methanoregula, Methanosaeta and Methanobacterium were possible methane oxidizers [29, 30].

Taxonomic classification of pyrosequences from archaeal communities of the nine samples at the class levels. Relative abundance was defined as the number of sequences affiliated with that taxon divided by the total number of sequences per sample

Taxonomic classification of pyrosequences from archaeal communities of the nine samples at the genus levels. Relative abundance was defined as the number of sequences affiliated with that taxon divided by the total number of sequences per sample
Discussion
In this study, the biofilter was operated about 1 year before this experiment, therefore, a plenty of AOB, NOB, IOB and MnOB were attached on the media of the biofilter, which could quickly oxidize ammonia, iron and manganese, respectively. Experimental results also showed that an excellent performance for ammonia, iron and manganese removal was achieved. Iron and ammonia were simultaneously removed from the top of filter bed, while manganese was removed from 0.2 m of the filter depth. Because removal of manganese occurred after ferrous iron was oxidized completely due to ferrous iron could react with manganese oxides [21]. When manganese in influent increased to about 5 mg/L, iron and ammonia in effluent were both below 0.1 mg/L, indicating that iron and ammonia removal were hardly affected. Surprisingly, manganese in effluent was lower than 0.05 mg/L; this is because manganese sand presented a high ability to adsorb manganese. When manganese increased to about 10 mg/L, iron and ammonia in effluent were both below 0.1 mg/L, too. In the biofilter, manganese was removed by manganese sand adsorption and biological oxidation [6]. First, manganese in effluent was below 0.05 mg/L due to the high adsorption ability of manganese sand to manganese; and then gradually increased to 1.02 mg/L due to the fact that the adsorption capacity of manganese sand was getting saturated; finally decreased to lower than 0.05 mg/L several days latter, since the biological manganese removal ability was enhanced with the growth of MnOB. When manganese in influent increased from about 1 to 10 mg/L, ammonia and iron removal were not affected, while the concentration of manganese along the filter depth obviously increased. Ammonia oxidation followed a first order kinetic rate in different phases, and the half-life time of ammonia oxidation was less varied. The kinetics of manganese oxidation were indeed first order when manganese in influent was about 1 and 5 mg/L, which is in agreement with previous studies of biological manganese removal [23, 31]. But it did not follow first order when manganese increased to about 10 mg/L.
Large amounts of iron and manganese oxides, which covered on the surfaces of the bacteria observed in SEM images, evidenced that microorganisms acted as catalytic role in iron and manganese removal from groundwater [16]. The type of microorganisms presented typical structure of twisted stalk may be recognized as the most common IOB and MnOB (Gallionella), which was commonly found in groundwater treatment plants treated iron and manganese [15, 22, 30]. The other type of microorganisms presented rod-shaped characteristic was also found in the biofilter, and was most likely Bacillus, which is an endospore-forming bacterium commonly found in water sources, soil and in association with plants [32]. This bacterium can oxidize manganese [16].
The distribution and genetic diversity of the microorganisms in the biofilter in different phases was also investigated. In phase I (day 31), iron was oxidized by IOB-Crenothrix and Gallionella in 0 m of the filter depth, while MnOB-Crenothrix and Gallionella oxidized manganese in 0.4–0.8 m since manganese removal was occurred after ferrous iron was oxidized completely [6]. And the abundance of MnOB-Crenothrix and Gallionella decreased along the filter depth with the decreasing of manganese. The abundance of Nitrosococcus (AOB) increased along the filter depth, but removal amount of ammonia decreased. The reason may be that the quantity of microbe in A1 was higher than that in A2 and much higher than that in A3, and although the quantity of Nitrosococcus obviously decreased from A1 to A2 and A3, the abundance of Nitrosococcus increased.
In phase II (day 93), although the concentration of iron along the filter depth was hardly changed, the abundance of IOB-Crenothrix in 0 m of the filter depth decreased with the increasing of manganese concentration, while the abundance of IOB-Gallionella increased. When manganese in influent increased to about 5 mg/L, the abundance of MnOB-Gallionella obviously increased in 0.4 and 0.8 m, and the abundance of Crenothrix increased in 0.8 m, but Crenothrix decreased in 0.4 m. The reasons may be as follows: (1) Gallionella had a higher activity than Crenothrix in high manganese condition. (2) The quantity of the MnOB in 0.4 m increased with the increasing of manganese concentration, and the quantity of the microorganism increased, too; the growth rate of microorganism was higher than Crenothrix in 0.4 m. In phase II, the quantity of the microorganism in 0.4–0.8 m increased, because the quantity of the MnOB increased with increasing of manganese concentration, therefore the abundance of AOB-Nitrosococcus decreased in 0.4–0.8 m, although the quantity of the Nitrosococcus may be hardly varied compared with phase I.
In phase III (day 158), although the removal efficiency of iron was not affected with the increasing of manganese concentration, the abundance of IOB-Crenothrix in 0 m of the filter depth continuously decreased, while the abundance of IOB-Gallionella continuously increased, indicating that IOB-Gallionella was more able to adapt the high manganese condition than IOB-Crenothrix. When manganese in influent increased to about 10 mg/L, the growth rate of MnOB-Gallionella was faster than that of Crenothrix, and the abundance of Gallionella obviously increased with the increasing of manganese concentration. The abundance of Crenothrix increased in 0.8 m, but Crenothrix decreased in 0.4 m, because the total number of the microorganism increased. The removal efficiency of ammonia along the filter depth and the half-life time of ammonia oxidation were hardly affected by manganese, but the abundance of AOB-Nitrosococcus in 0.8 m decreased because of the increasing of total number of the microorganism.
When manganese increased from 1 to 5 and 10 mg/L, iron and ammonia removal were not affected, while manganese increased in 0.4, 0.8 and 1.2 m of the filter depth. Biological ammonia oxidation followed a first order kinetic rate, while manganese oxidization was indeed first order except manganese in influent was about 10 mg/L. AOB, NOB, IOB, IOA, MnOB related to ammonia, iron and manganese removal were found in the biofilter. With the increasing of manganese concentration, IOB-Crenothrix was restrained, while IOB-Gallionella was promoted. The abundance of MnOB-Gallionella quickly increased, and MnOB-Crenothrix in 0.8 m increased, too, but decreased in 0.4 m. The abundance of Nitrosococcus in 0.4 and 0.8 m decreased because of the increasing of total number of the microorganism.
References
Jusoh AB, Cheng WH, Low WM, Nora’aini A, Megat MJ, Noor M (2005) Study on the removal of iron and manganese in groundwater by granular activated carbon. Desalination 182:347–353
Azher NE, Gourich B, Vial C, Soulami MB, Ziyad M (2008) Study of ferrous iron oxidation in Morocco drinking water in an airlift reactor. Chem Eng Process 47:1877–1886
Li XK, Chu ZR, Liu YJ, Zhu MT, Yang L, Zhang J (2013) Molecular characterization of microbial populations in full-scale biofilters treating iron, manganese and ammonia containing groundwater in Harbin, China. Biores Technol 147:234–249
Tekerlekopoulou AG, Vayenas DV (2008) Simultaneous biological removal of ammonia, iron and manganese from potable water using a trickling filter. Biochem Eng J 39:215–220
Zeng XP, Xia J, Wang ZZ, Li WH (2015) Removal of iron and manganese in steel industry drainage by biological activated carbon. Desalin Water Treat 56:2543–2550
Cheng QF (2016) Competitive mechanism of ammonia, iron and manganese for dissolved oxygen using pilot-scale biofilter at different dissolved oxygen concentrations. Water Sci Technol Water Supply 16:766–774
Mustika SA (2004) Potential optimisation of chlorination at Buaran WTP, Jakarta Water and Environmental Engineering. Degree of Master of Science, University of Surrey, Guildford
Cheng QF, Nengzi LC, Bao LL, Wang YJ, Yang JX, Zhang J (2017) Interactions between ammonia, iron and manganese removal using pilot-scale biofilters. J Water Supply Res Technol AQUA 66:157–165
Du X, Liu GY, Qu FS, Li K, Shao SL, Li GB, Liang H (2017) Removal of iron, manganese and ammonia from groundwater using a PAC-MBR system: the anti-pollution ability, microbial population and membrane fouling. Desalination 403:97–106
Richardson SD, Postigo C (2012) Drinking water disinfection by-products. In: The handbook of environmental chemistry emerging organic contaminants and human Health, vol 20. Springer, New York, pp 93–137 (Chapter 4)
Hasan HA, Abdullah SRS, Kamarudin SK, Kofli NT, Anuar N (2014) Kinetic evaluation of simultaneous COD, ammonia and manganese removal from drinking water using a biological aerated filter system. Sep Purif Technol 130:56–64
Cai YA, Li D, Liang YW, Luo YH, Zeng HP, Zhang J (2015) Effective start-up biofiltration method for Fe, Mn, and ammonia removal and bacterial community analysis. Biores Technol 176:149–155
Tekerlekopoulou AG, Vayenas DV (2007) Ammonia, iron and manganese removal from potable water using trickling filters. Desalination 210:225–235
Han M, Zhao ZW, Gao W, Cui FY (2013) Study on the factors affecting simultaneous removal of ammonia and manganese by pilot-scale biological aerated filter (BAF) for drinking water pre-treatment. Biores Technol 145:17–24
Hasan HA, Abdullah SRS, Kofli NT, Kamarudin SK (2012) Effective microbes for simultaneous bio-oxidation of ammonia and manganese in biological aerated filter system. Biores Technol 124:355–363
Yang L, Li XK, Chu ZR, Ren YH, Zhang J (2014) Distribution and genetic diversity of the microorganisms in the biofilter for the simultaneous removal of arsenic, iron and manganese from simulated groundwater. Biores Technol 156:384–388
Granger HC, Stoddart AK, Gagnon GA (2014) Direct biofiltration for manganese removal from surface water. J Environ Eng 140:223–224
Tang WW, Gong JM, Wu LJ, Li YF, Zhang MT, Zeng XP (2016) DGGE diversity of manganese mine samples and isolation of a lysinibacillus sp. efficient in removal of high Mn (II) concentrations. Chemosphere 165:277–283
Mckee KP, Vance CC, Karthikeyan R (2016) Biological manganese oxidation by pseudomonas putida in trickling filters. J Environ Sci Health Part A Toxic/Hazard Subst Environ Eng 51:523–535
Rittmann BE, Snoeyinck VL (1984) Achieving biologically stable drinking water. J AWWA 76:156–174
Cheng QF, Nengzi LC, Bao LL, Huang Y, Liu SY, Cheng XW, Li B, Zhang J (2017) Distribution and genetic diversity of microbial populations in the pilot-scale biofilter for simultaneous removal of ammonia, iron and manganese from real groundwater. Chemosphere 182:450–457
Caporaso JG, Lauber CL, Walters WA, Berg-Lyons D, Lozupone CA, Turnbaugh PJ, Fierer N, Knight R (2011) Global patterns of 16S rRNA diversity at a depth of millions of sequences per sample. Proc Natl Acad Sci USA 108:4516–4522
Katsoyiannis IA, Zouboulis AI (2004) Biological treatment of Mn(II) and Fe(II) containing groundwater: kinetic considerations and product characterization. Water Res 38:1922–1932
Chu ZR, Wang K, Li XK, Zhu MT, Yang L, Zhang J (2015) Microbial characterization of aggregates within a one-stage nitritation–anammox system using high-throughput amplicon sequencing. Chem Eng J 262:41–48
Lu L, Xing DF, Ren NQ (2012) Pyrosequencing reveals highly diverse microbial communities in microbial electrolysis cells involved in enhanced H2 production from waste activated sludge. Water Res 46:2425–2434
Lasík J, Koníček J, Roušalová O (1993) Production of an exocellular polymer in a mutant strain of Achromobacter delicatulus, on a new type of stirrer. Folia Microbiol 38:71–73
Li D, Stanford B, Dickenson E, Khunjar WO, Homme CL, Rosenfeldt EJ (2017) Effect of advanced oxidation on N-nitrosodimethylamine (NDMA) formation and microbial ecology during pilot-scale biological activated carbon filtration. Water Res 113:160–170
Rawlings DE (2005) Characteristics and adaptability of iron- and sulfur-oxidizing microorganisms used for the recovery of metals from minerals and their concentrates. Microb Cell Factor 4:13
Bharathi M, Chellapandi P (2017) Intergenomic evolution and metabolic cross-talk between rumen and thermophilic autotrophic methanogenic archaea. Mol Phylogenet Evol 107:293–304
Rocheleau S, Greer CW, Lawrence JR, Cantin C, Larame´e L, Guiot SR (1999) Differentiation of Methanosaeta concilii and Methanosarcina barkeri in anaerobic mesophilic granular sludge by fluorescent in situ hybridization and confocal scanning laser microscopy. Appl Environ Microbiol 65:2222–2229
Cheng QF, Nengzi LC, Xu DY, Guo JY, Yu J (2017) Influence of nitrite on the removal of Mn(II) using pilot-scale biofilters. J Water Reuse Desalin 7:264–271
Kunst F, Vassarotti A, Danchin A (1995) Organization of the European Bacillus subtilis genome sequencing project. Microbiology 389:84–87
Acknowledgements
This work was kindly supported by National Natural Science Foundation of China (51808062), and Department of Science and Technology of Sichuan Province (19ZDYF0150).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflict of interest to declare.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Cheng, Q., Huang, Y., Nengzi, L. et al. Performance and microbial community profiles in pilot-scale biofilter for the simultaneous removal of ammonia, iron and manganese at different manganese concentrations. Bioprocess Biosyst Eng 42, 741–752 (2019). https://doi.org/10.1007/s00449-019-02077-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-019-02077-x