
Abstract
Equimolar amounts of lactobionic acid and sorbitol may be obtained in a reaction catalyzed by the enzymes glucose–fructose oxidoreductase and glucono-δ-lactonase, which are found in the periplasm of Zymomonas mobilis. These reactions are generally conducted using immobilized bacterial cells, and the cell treatment and immobilization steps are costly and time-consuming. This study evaluated alternatives to simplify the preparation of calcium alginate-immobilized biocatalyst and its application in different operation modes and types of reactors. It was possible to eliminate cell permeabilization with cetyltrimethylammonium bromide, and the reticulation of Z. mobilis cells with glutaraldehyde sufficed to inhibit the fermentative metabolism of carbohydrates by the bacterium, with accumulation of bioconversion products. When the process was carried out in a mechanically stirred reactor in batch mode, 530 mmol L− 1 of products were obtained in 24 h. The process was also tested in fed-batch mode so as to use of a larger amount of lactose, since it could not be used in the batch because of its low solubility in water. Under this condition, final products concentration reached 745 mmol L− 1 within 42 h. Similar results were obtained for reactions conducted in a pneumatically stirred reactor in batch and fed-batch modes, proving the potential use of this process in several industrial settings.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Lactobionic acid is an aldonic acid produced when lactose is oxidized [1, 2]. The compound has several important applications, as in anti-ageing skin care [3], preservation of human organs for transplants [4], vectorization of antitumor drugs [5], and improvement of solubility of macrolide antibiotics [6]. As a potent sweetener, sorbitol (C6H14O6) is used in the food industry, especially in the formulation of diabetes products [7], toothpaste [7, 8], and vitamin C tablets [7].
Equimolar amounts of lactobionic acid and sorbitol may be obtained in a reaction catalyzed by the enzymes glucose–fructose oxidoreductase (GFOR) and glucono-δ-lactonase (GL), which are found in the periplasm of Zymomonas mobilis [9, 10]. This anaerobic, Gram-negative bacterium metabolizes sucrose, glucose, and fructose as carbon sources through the same biochemical route, the Entner–Doudoroff pathway [11]. The process typically includes a first step in which Z. mobilis cells, containing the periplasmatic enzymes (GFOR/GL), are grown in a cultivation medium with glucose as carbon source [11]. In the sequence, the cells obtained are concentrated by centrifugation [12,13,14] and permeabilized using toluene [15] or cationic detergents like cetyltrimethylammonium bromide (CTAB) [16]. The aim of this permeabilization step is to inactivate the fermentative metabolism of Z. mobilis (since the catalytic action of GFOR/GL does not depend on cell viability [15]) and, therefore, to produce lactobionic acid and sorbitol in the reaction medium. The subsequent stages of the process include cell immobilization in calcium alginate [13,14,15] and the customary final treatments to recover and purify these compounds [17].
Although good yields are obtained with cell permeabilization in laboratory [14,15,16, 18,19,20], this process may face technical obstacles at industrial scale, increasing production costs [21, 22]. In the effort to address this issue, Silveira et al. [22] demonstrated that high initial concentrations of substrates (glucose and fructose) and a subsequent rise in concentrations of products inhibit the production of ethanol, which makes the permeabilization of the cellular mass unnecessary.
The permeabilized cellular mass is reticulated with glutaraldehyde before and after immobilization in alginate gels [12,13,14, 20]. Glutaraldehyde is used in the cross-linking of proteins [23], since it reacts with several chemical groups, like amino, thiol, phenol, and imidazole, for instance [24, 25]. The high reactivity of glutaraldehyde in the reticulation of amino groups present in the outer layers of the microorganisms (lipoprotein components) is the factor responsible for the high biocide efficiency against Gram-negative bacteria [24].
As long as enzyme activity is not inhibited, the high initial concentrations of substrates in bioconversion assays improve reaction rates, since the kinetic constant (K M) of GFOR is 1.2 mol L− 1 at 30 °C and pH 6.2 [10]. Nevertheless, solubility of lactose in water at 30 °C is only 0.6 mol L− 1 [10]. Even though lactose-saturated substrate solutions prepared in boiling water are used in bioconversion, the highest product yields and maximal products formation rates in reactions based on calcium alginate-immobilized Z. mobilis were achieved employing 700 mmol L− 1 lactose and 600 mmol L− 1 fructose [12, 14].
Several studies have described the reactions to produce organic acids like gluconic or lactobionic acids and sorbitol by Z. mobilis immobilized in calcium alginate in reactors with mechanical and magnetic agitation [12,13,14,15, 18, 26, 27]. Most of these works were carried out using batch mode [12,13,14,15, 27], although the study published by Jung et al. [18] investigated a continuous operation. These reactions have not been described in fed-batch, except for the study carried out by Satory et al. [10], who used purified GFOR/GL. More specifically in the production of lactobionic acid and sorbitol, Malvessi et al. [28] evaluated different reactor settings (mechanically stirred, fixed-bed and fluidized-bed reactors). The authors underscored the high efficiency of mechanically stirred reactors due to the improved contact between enzyme and substrate and simple process control, as observed for pH. Typically, the reactions conducted in magnetically stirred reactors afforded to reach concentrations of up to 500 mmol L− 1 and yields of about 70% [12,13,14, 27].
In this context, this study assessed the production of lactobionic acid and sorbitol by GFOR/GL by Z. mobilis cells immobilized in calcium alginate under different operational conditions. The aim was to increase the potential of this technology to be transferred to the industrial sector. The experiments were conceived to evaluate the use of glutaraldehyde as cell metabolism inhibitor instead of permeabilization with CTAB, and the application of the immobilized Z. mobilis in different operation modes and types of bioreactors to improve the final results of the process.
Materials and methods
Strain and cultivation conditions
Zymomonas mobilis strain ATCC 29191 was used in this work. Cultures were maintained in liquid medium suspension, at 4 °C, according to the procedures described by Malvessi et al. [29], and monthly replicated in order to keep cell viability. Liquid medium [29] used for maintenance, inoculum preparation, cell growth, and enzyme production had the following composition (in g L− 1): glucose, 20 (maintenance), 100 (inoculum), 150 (biomass and enzyme production); (NH4)2SO4, 2.0; MgSO4·7H2O, 1.0; KH2PO4, 3.5; FeSO4.7H2O, 0,01; Prodex Lac® yeast extract (Prodesa S.A., Brazil), 7.5.
For inoculum preparation, initial pH was adjusted to 5.5 and the excessive pH drop during the cultivation was avoided by adding 5 g L− 1 of CaCO3 to the medium. Concentrated glucose solutions and CaCO3 were separately sterilized and added to the medium before inoculation. Sterilization of nutrients and glucose solutions was accomplished by autoclaving at 1 atm for 20 min.
Inocula were prepared in 500 mL anaerobic bottles, with CO2 release filters, filled with 450 mL of medium and kept under orbital agitation of 200 rpm (Certomat U, Sartorius Stedim Biotech, Germany), at 30 °C, for approximately 10 h. Batch cultivations of Z. mobilis cells were carried out in a 5.5 L bioreactor (model Tecbio, Tecnal, Brazil). The temperature was kept at 30 °C, the impeller speed was 450 rpm, and the pH was maintained at 5.5 with 5.0 mol L− 1 NaOH. Cell mass was harvested from the medium by centrifugation at 5836g, for 10 min, using a centrifuge (Sigma 4K-15, Sartorius Stedim Biotech, Germany). Cells were re-suspended in distilled water to a concentration of 25 g L− 1 on dry basis.
Cell permeabilization
When required, Z. mobilis cells were permeabilized with CTAB as described by Rehr et al. [16]. Equal volumes of concentrated biomass suspension and 0.2% (w/v) CTAB solution were mixed, and the mixture was gently stirred for 15 min at 4 °C. The permeabilized cells were then centrifuged at 5836g and re-suspended to a concentration of 50 g L− 1 on dry basis.
Glutaraldehyde cross-linking of cells
In some tests, viable Z. mobilis cells were cross-linked with 0.5% (v/v) glutaraldehyde under magnetic stirring for 10 min, as described by Jang et al. [18], centrifuged and re-suspended to the convenient concentrations to be used in free-cell bioconversion runs.
Immobilization method
The cells were immobilized in calcium alginate beads by entrapment as described in a previous work [20]. To immobilize Z. mobilis cell in calcium alginate, equal volumes of concentrated cell suspension (70 g L− 1 on dry basis), with no previous treatment, and 4% (w/v) of sodium alginate solution (Algogel 5540, Cargill S.A.) were mixed. In this heterogeneous system, 64% of dry cell mass per total mixture mass (% w/w), respectively, was obtained. The mixture was kept under agitation for 1 h to achieve complete homogenization and then dropwise poured through hypodermic needles onto a CaCl2 0.3 mol L− 1 solution under magnetic stirring in order to form cell-containing calcium alginate beads. The beads thus formed were cross-linked with 0.5% (w/v) glutaraldehyde for 15 min under magnetic stirring at room temperature. The calcium alginate beads were kept at 4 °C in distilled water and were subsequently used in bioconversion experiments. The average size of calcium alginate beads was approximately 2.35 × 10− 3 m.
Evaluation of the inactivation of fermentative metabolism of Z. mobilis
The evaluation of inactivation of cell metabolism in Z. mobilis was initially carried out submitting the biomass treated with CTAB or glutaraldehyde to normal cultivation conditions (glucose 100, 0.5 g L− 1 cell biomass, 30 °C, pH 5.5) in anaerobic bottles kept under orbital agitation of 200 rpm (Certomat U/H, B. Braun Biotech, Germany) for a 20 h of incubation. Untreated biomass was used in control experiments. Samples were collected at the beginning of the experiment and after 20 h to determine sugars and thus establish substrate consumption. Subsequently, the influence of these treatments on enzyme activity was evaluated based on the determination of enzyme activity of the GFOR/GL complex.
Bioconversion experiments
Bioconversion experiments were initially carried out in a glass reactor with 63 mm of diameter and 115 mm of height, with a working volume of 200 mL, the agitation being provided by magnetic stirring. Furthermore, mechanical and pneumatic stirred reactors were used. The mechanically stirred reactor consisted of a glass vessel with 110 mm of diameter and 160 mm of height that was operated with 400 mL of reaction medium. In this reactor, mixing was provided by an anchor type impeller with 10 mm of wide, 61 mm of diameter and 79 mm of height, which was coupled to a motor running at 100 rpm. The pneumatically stirred reactor consisted of a glass tube with 54 mm of diameter and 300 mm of height in which the medium volume was also 400 mL. The agitation of this reactor was promoted by a pre-heated air flow of 1.0 L min−1. All reactors were water jacketed in order to control the reaction temperature.
Batch bioconversion assays were conducted in the 200-mL reactor to evaluate the effect of the single treatment with glutaraldehyde on the catalytic activity of the enzyme complex for long periods (24 h) to detect the generation of the products of interest in the reaction medium. A first group of experiments was carried out using free cells in a suspension that was treated with glutaraldehyde. Untreated cells were used as control. Subsequently, the tests were carried out using the calcium alginate-immobilized biocatalyst prepared as previously described. These reactions were carried out using two concentrations (400 and 700 mmol L− 1) of the substrate pairs glucose/fructose and lactose/fructose each.
In both 400-mL bioreactors, batch and fed-batch bioconversion experiments with immobilized cells were done to assess alternatives for an eventual industrial application of this process. In batch and fed-batch modes, the initial concentrations of lactose and fructose used were 700 and 600 mmol L− 1, respectively. In fed-batch runs, substrates were fed into the reactors as products were formed. The concentration of substrates was kept at the initial level for up to 9 h of process. As such, with every 18 mmol L− 1 of products formed (lactobionic acid and sorbitol), the required amounts of substrates were added so as to provide an 18 mmol L− 1 increase in lactose and an 17 mmol L− 1 increase in fructose levels in the reaction medium.
All bioconversion tests were performed with 20 g L− 1 of Z. mobilis cells, at 39 °C, and at pH 6.4 by controlling with NaOH 7 mol L− 1.
Analytical methods
Cell concentration was determined by measuring the optical density of Z. mobilis suspensions at 560 nm. These measurements gave a linear relationship with dry cell mass concentration.
The concentration of reducing sugars in the culture media was determined according to the DNS (3,5-dinitro-salicylic acid) method [30].
In the tests for evaluation of the inactivation of Z. mobilis fermentative metabolism, the concentration of substrates (glucose or lactose and fructose) and products (gluconic or lactobionic acids and sorbitol) was preferentially quantified by high-performance liquid chromatography (HPLC) as described by Pedruzzi et al. [31] with modifications. The system used was an HPLC Agilent Technology model 9100 equipped with a refractive index detector (IR). The chromatographic column used was Aminex HPX-87H (300 mm × 7.8 mm × 9 µm), with 0.0005 mol L− 1 sulfuric acid being used as mobile phase at a flow rate of 0.4 mL min− 1. The system was operated at 60 °C and the volume of sample injection was 5 µL. In bioconversion experiments with inactivated Z. mobilis cells, the concentration of substrates and products was estimated based on the consumption of NaOH used to control the pH (1 mol of aldonic acid formed per mol of NaOH consumed). As shown by Pedruzzi et al. [13], the responses of both methods (HPLC and NaOH automatic titration) in this process are practically identical. In these cases, once that organic acid and sorbitol are formed in equimolar basis, and also assuming no other product is formed in the bioconversion experiments, the molar concentration of sorbitol and of the acid can be considered the same.
The activity of GFOR/GL was determined as described by Malvessi et al. [20], using glucose/fructose 700 mmol L− 1, cell concentration 4.0 g L− 1, temperature 39 °C and pH 6.4. One GFOR/GL (U) is defined as the amount of enzyme that produced 1 mmol h− 1 gluconic acid. Activity was expressed as units per gram of cells, dry basis (U g− 1).
Parameters of evaluation
The conversion yield in relation to the initial substrate (Y P/S0, mmol of product per mmol of initial substrate) was determined by dividing the number of mmol of formed product by the number of mmol of substrate in the initial solution.
The volumetric productivity (p, mmol of product per liter of solution per h) was calculated by dividing the final concentration of products by the process time.
The specific productivity (q, mmol of product per gram of cell per h) was determined by dividing the volumetric productivity by the cell concentration used in the bioconversion run.
Statistical analysis
The results were evaluated by analysis of variance (ANOVA) followed by Tukey’s post hoc test, at a probability level of less than 5% (p < 0.05) with the SPSS Statistics 20 program, and were presented as the means and standard deviation calculated from duplicate for each condition tested.
Results and discussion
Evaluation of the inactivation of Z. mobilis fermentative metabolism
To investigate the inactivation of Z. mobilis metabolism, bacterial growth assays after cells were submitted to different treatments: only with CTAB 0.2% or only glutaraldehyde 0.5%. The results were compared with untreated cells. Substrate consumption was determined 20 h after incubation and enzyme activity was established for each case. The results are presented in Table 1.
Twenty hours after the beginning of cultivation, untreated cells consumed essentially all substrate. However, substrate consumption by cells treated with CTAB was significantly lower at the same period, indicating metabolism inhibition. These results confirm the efficiency of this cell permeabilization method to inhibit Z. mobilis, as previously described by Rehr et al. [16]. Similar results were observed for the treatment with glutaraldehyde, pointing to the potential use to inhibit the fermentative metabolism of this bacterium. The comparison between untreated cells and those treated with CTAB and glutaraldehyde shows significant differences in substrate consumption. On the other hand, no such differences were observed between the two treatments, leading to the conclusion that treatment of cells with glutaraldehyde induced the inhibition of bacterial growth to a level that was statistically similar to what was observed for treatment with CTAB. The catalytic activity of enzymes (Table 1) was statistically identical, independently of the treatment cells were submitted to, meaning that the activity of the enzymes GFOR/GL was not affected by treatment with glutaraldehyde to inhibit the fermentative metabolism of Z. mobilis. This results indicate that it is possible to use this simple treatment to replace permeabilization of cell wall with toluene or CTAB, as previously described by Chun and Rogers [15] and Rehr et al. [16].
Cell inactivation with glutaraldehyde significantly reduced substrate consumption and did not induce significant changes in enzymatic activity. However, in the technique of determination of GFOR/GL activity only the first 30 min of reaction are considered, when the high initial concentration of substrates may inhibit the fermentative pathway, preventing substrate consumption. For this reason, the bioconversion experiments were conducted for 24 h using free cells of Z. mobilis treated with glutaraldehyde and without any treatment to evaluate the effect of this treatment on the accumulation of products under industrial production condition (long reaction times). As mentioned above, the substrates evaluated were glucose/fructose and lactose/fructose at 400 and 700 mmol L− 1. Glucose was used because it is preferred by Z. mobilis fermentative metabolism. The tests conducted using reduced substrates concentrations aimed to prevent inhibition of fermentative metabolism of the bacterium by substrates, as described by Silveira et al. [22] and to confirm whether the accumulation of bioconversion products resulted from the treatment with glutaraldehyde. Although lactose is not a carbon source used by Z. mobilis [11], the bacterium metabolizes fructose through the Entner–Doudoroff pathway to produce ethanol, which means less product concentrations (lactobionic acid and sorbitol) in the reaction medium. The results of these assays are shown in Table 2.
When 400 mmol L− 1 glucose and fructose were used with untreated cells, the amount of gluconic acid produced was completely consumed by the fermentative metabolism of the bacterium 4.5 h after the beginning of the reaction (Fig. 1a). These results confirm the findings reported by Silveira et al. [22], who conducted reactions with an initial substrate concentration of 280 mmol L− 1 and untreated biomass and observed total consumption of gluconic acid after 1 h of reaction. In the present study, the final concentration of gluconic acid was null, indicating that all gluconate produced entered the Entner–Doudoroff pathway after being converted to 6-P-gluconate by gluconate kinase. During bioconversion, the production of gas (probably CO2) and foam was observed, which is a typical behavior of fermentation processes. On the other hand, under the same condition, but using glutaraldehyde-treated cells, the suppression of the fermentative metabolism of Z. mobilis was confirmed, promoting the accumulation of products in the reaction medium (356 mmol L− 1) (Fig. 1b). In Fig. 1b, it is interesting to observe that gluconic acid concentrations measured by HPLC and estimated from NaOH consumption were very similar. The stoichiometric relationship observed between alkali consumption and gluconic acid formation is a further evidence of the effectiveness of glutaraldehyde as Z. mobilis inactivating agent. When the initial substrate concentration was 700 mmol L− 1, a small accumulation of gluconic acid was observed with untreated cells (0.29 mmol L− 1), confirming the results obtained by Silveira et al. [22]. The authors reported partial inhibition of Z. mobilis fermentative metabolism when substrate concentration was 800 mmol L− 1, preventing the total consumption of this product. The treatment with glutaraldehyde when the highest substrate concentration was used also promoted the accumulation of considerable concentration of products in the reaction medium (620 mmol L− 1).

Time courses of bioconversion runs with free cells of Z. mobilis untreated (a) and treated with glutaraldehyde (b) with initial equimolar glucose and fructose concentrations of 400 mmol L− 1. Filled circle: glucose, filled triangle: fructose, open circle: gluconic acid determined by HPLC, open square: gluconic acid estimated from NaOH consumption, open triangle: sorbitol (pH, 6.4; temperature, 39 °C; 20 g L− 1 of free-cells)
Similar results were observed in the assays using lactose and fructose. Table 2 shows that the conversion yield in relation to the initial substrate for untreated cells was lower than that observed when glutaraldehyde was employed, in both substrate concentrations (400 and 700 mol L− 1). These results indicate that part of the fructose in solution was used by the fermentative metabolism of the bacterium. On the other hand, the conversion yield using glutaraldehyde-treated cells was higher than 0.75 mmol L− 1, which confirms the observations made by Malvessi et al. [14] and Carra [12] using free and permeabilized Z. mobilis cells.
Since the cell immobilization step includes cross-linking of the support beads with glutaraldehyde, similar bioconversion assays were carried out with biomass with no previous treatment immobilized in calcium alginate. The aim was to evaluate whether the subsequent treatment of beads with glutaraldehyde inhibits Z. mobilis fermentative metabolism and provides the accumulation of products at appreciable concentrations in the medium. The results are shown in Table 3.
The results obtained in these assays confirmed the accumulation of products of interest (gluconic or lactobionic acid and sorbitol) in the reaction medium regardless of the substrates concentration used (Table 3). When glucose and fructose were used as substrates, yields of around 95% in gluconic acid and sorbitol were obtained, which are similar to the values published by Chun and Rogers [15] in the bioconversion of toluene-permeabilized Z. mobilis cells immobilized in calcium alginate. When lactose and fructose were employed, lactobionic acid and sorbitol yields were approximately 75%, a value that is higher than that reported by Malvessi et al. [14] for a biomass permeabilized with CTAB and immobilized in calcium alginate. Figure 2 shows the curves of substrates consumption and products formation using 700 mmol L− 1 substrates (glucose/fructose and lactose/fructose). For the experiment with the pair glucose/fructose, the curve for the time course of gluconic acid concentration as estimated from the consumption is also shown. Again, as already discussed for glutaraldehyde-treated free cells, a very good stoichiometric relationship between gluconic acid and NaOH was observed, indicating the complete inactivation of the immobilized cells fermentative metabolism.

Time courses of bioconversion runs with Z. mobilis cells immobilized in glutaraldehyde-cross-linked calcium alginate beads with different initial concentrations of substrates. Glucose and fructose (a); lactose and fructose (b). Filled circle: aldoses (glucose or lactose), filled triangle: fructose, open circle: aldonic acid determined by HPLC (gluconic or lactobionic), open square: gluconic acid estimated from NaOH consumption, open triangle: sorbitol (pH, 6.4; temperature, 39 °C; 20 g L− 1 of immobilized-cells)
The preparation of immobilized biocatalyst with one single treatment with glutaraldehyde was technically possible, and did not affect the conversion yield and productivity of the process. Also, it afforded to eliminate a costly step of the production process. Even more interesting was the use of biomass that did not undergo any treatment prior to immobilization in calcium alginate, proving that the reticulation of the support containing whole cells with glutaraldehyde is enough to inhibit the fermentative metabolism of Z. mobilis. In this case, two stages of the process previously developed could be excluded, reducing production costs even more.
Evaluation of different systems to produce lactobionic acid and sorbitol
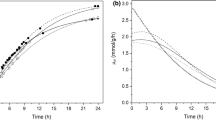
Despite the considerable amount of data in the literature about different aspects of the process evaluated in the present study, few publications have addressed the potential of this technology for use in industrial sector. Therefore, the present study evaluated alternative systems to produce lactobionic acids and sorbitol by Z. mobilis immobilized in calcium alginate with a view to applications in large-scale processing. The reactions were conducted in a mechanically stirred reactor and in a pneumatically stirred reactor in both batch and fed-batch modes. The results are shown in Table 4 and the kinetic profiles obtained are illustrated in Fig. 3.

Time courses of bioconversion runs with immobilized cells of Z. mobilis carried out in a batch (a) and fed-batch (b) modes. Filled circle: mechanically stirred reactor filled triangle: pneumatically stirred reactor
In fed-batch mode, substrates were added to the reaction medium as products were formed, the amount of NaOH consumed to control pH being used as the estimation parameter.
As shown in Table 4, the batch reactions conducted under mechanical stirring afforded to obtain approximately 530 mmol L− 1 of products, with conversion yield of 81% and volumetric productivity of 22 mol L− 1 h− 1. In most studies published in the literature, these reactions were carried out in a batch with magnetic stirring, which enabled a conversion yield of roughly 70%, maximum final concentration of 500 mmol L− 1, and productivity of 10 mmol L− 1 h− 1 [12,13,14, 27]. Although the final concentrations of products and conversion yield in the tubular reactor were lower than those observed for reactors with mechanical stirring in the present study, they were similar to those described in the literature for reactors with magnetic stirring [14, 27]. This enabled conducting reactions in mechanically stirred as well as in pneumatically stirred reactors.
Except for the work published by Malvessi et al. [28], no other data were available about the use of pneumatically stirred reactor to conduct the reactions investigated in the present study. Previous studies have found that air injection is an adequate approach to achieve good mixing and efficient mass transfer with much milder shear force on the immobilized support [32,33,34]. However, independently of the reactor type used, no differences in integrity of the support were noticed in the present study. This may have been due to the air flow (1 L min− 1) required to attain a good mixing and an efficient mass transfer. Another explanation is that the use of an anchor impeller would reduce the shear stress on the calcium-alginate beads, minimizing the differences in the integrity of the support. Despite that, a pneumatically stirred reactor is attractive in large-scale bioprocesses because of its simplicity, easy operation and low power consumption [34].
One of the alternatives to increase the reaction rate and, therefore, the final concentration of products is the use of high initial substrate concentrations, since K M of GFOR for lactose in combination with fructose is substantial [10, 14]. However, the use of high lactose levels may be problematic due to the low solubility of this carbohydrate, approximately 0.6 mol L− 1 [10]. This limitation may be controlled in fed-batch mode, when substrates are added to the medium as products are formed. But this method demands higher product solubility compared with that of substrates, under the reaction conditions (pH 6.4, 39 °C). The solubility of fructose in water at 20 °C is near 4.4 mol L− 1 [35, 36], while the value for sorbitol normally commercialized as 70% solutions (approximately 3.9 mmol L− 1) [37], showing that this half-reaction would not pose difficulties. Delagustin [38] observed that it is possible to use a solution of sodium lactobionate 1.3 mol L− 1, and noted that the compound is highly soluble. In the present study, the solubility of lactose and of sodium lactobionate was estimated in water at 39 °C and pH 6.4. Sodium lactobionate was found to be more soluble than lactose (> 1.3 and 0.94 mol L− 1, respectively), meeting the requirements for the reaction to be conducted in a fed-batch mode.
As shown in Table 4, the bioconversion conducted in fed-batch mode allowed to obtain significantly high final concentrations of products. These results were better than those obtained in batch in the present study and also in the literature, when maximum concentration was approximately 500 mmol L− 1 [14, 27]. The conversion yield in fed batch operation was 85%, slightly higher than in batch mode. Using calcium alginate-immobilized Z. mobilis, Pedruzzi et al. [13], Malvessi et al. [14] and Carra [27] obtained conversion rates of around 70%. Pedruzzi et al. [13] obtained lactobionic acid concentrations near 600 mmol L− 1 using a batch mode, but the reaction required 108 h using free-cells of Z. mobilis.
In the fed-batch mode, the high substrate-to-enzyme ratio was maintained for longer, favoring reaction rate and resulting in high final product concentrations in both reactors (mechanically stirred reactor and pneumatically-agitated tubular). The results obtained showed the important influence of lactose concentration in product formation rates, as described by Satory et al. [10], Pedruzzi et al. [13] and Malvessi et al. [14]. In these assays, the concentrations of lactobionic acid and sorbitol around 750 mmol L− 1 obtained represent a 50% increase in comparison with the best results described in the literature for Z. mobilis [14, 27]. In fact, since substrates were fed into the fed-batch mode, reaction times were longer than the 24 h needed in the batch mode, reducing volumetric (17.4 mmol L− 1 h− 1) and specific (0.97 mmol L− 1 h− 1) productivity values, compared with the results reported in the literature, 19 and 1.15 mmol L− 1 h− 1 [12, 14, 27]. Nevertheless, the results obtained after 24 of reaction in the fed-batch were also higher than those recorded for the batch mode, which reached 630 mmol L− 1 of products and productivity values of 24 and 1.50 mmol L− 1 h− 1. At this reaction time, residual lactose and fructose concentrations were 280 and 188 mmol L− 1, respectively. The longer reaction time afforded to reduce the residual lactose concentration to 139 mmol L− 1, with insignificant levels of fructose, which is an advantage considering the recovery and purification of the products. It should be emphasized that such reaction always ends with residual lactose levels due to the low affinity between GFOR and this aldose [10, 13, 14].
Conclusion
The results of the present study confirm the possibility to inhibit the fermentative metabolism of Z. mobilis with glutaraldehyde. It was possible to simplify this technique to prepare the calcium alginate-immobilized biocatalyst without loss of conversion yields and productivities of lactose and fructose conversion to lactobionic acid and sorbitol. In addition, a substantially high final products concentration was obtained in fed-batch mode both in mechanically and pneumatically stirred reactors. Taken together, these results represent a contribution to the technical and economic implementation of this process in industrial sector. Although these results have been demonstrated in this work for the production of gluconic and lactobionic acids, they certainly could be applied to the synthesis of other aldonic acids, such as xylonic, galactonic and maltobionic acids, that can be obtained with this enzymatic system.
References
Fischer E, Meyer J (1889) Oxydetion delr Milchsuckers. Berichte der Dtsch Chem Gesellschaft 22:361–364
Nath A, Verasztó B, Basak S et al (2016) Synthesis of lactose-derived nutraceuticals from dairy waste whey—a review. Food Bioprocess Technol 9:16–48
Grimes PE, Green BA, Wildnauer RH, Edison BL (2004) The use of polyhydroxy acids (PHAs) in photoaged skin. Cutis 73:3–13
Sumimoto R, Kamada N (1990) Lactobionate as the most important component in UW solution for liver preservation. Transplant Proc 22:2198–2199
Kim IS, Kim SH (2002) Development of a polymeric nanoparticulate drug delivery system: in vitro characterization of nanoparticles based on sugar-containing conjugates. Int J Pharm 245:67–73
Hoffhine CEJ (1956) Aqueous soluble salts of erythromycin. Pat Inven US2761859 A
Shwide-Slavin C, Swift C, Ross T (2012) Nonnutritive sweeteners: where are we today? Diabetes Spectr 25:104–110
Li QX, Yu ZY, Liao XS et al (2015) Preparation method and salification reactor of erythromycin lactobionate solution. Pat Inven CN104341474 A
Zachariou M, Scopes RK (1986) Glucose–fructose oxidoreductase, a new enzyme isolated from Zymomonas mobilis that is responsible for sorbitol production. J Bacteriol 167:863–869
Satory M, Fürlinger M, Haltrich D et al (1997) Continuous enzymatic production of lactobionic acid using glucose–fructose oxidoreductase in an ultrafiltration membrane reactor. Biotechnol Lett 19:1205–1208
Swings J, De Ley J (1977) The biology of Zymomonas. Bacteriol Rev 41:1–46
Carra S (2012) Estudo cinético da produção de ácido lactobiônico e sorbitol por enzimas periplasmáticas de Zymomonas mobilis. Diss Mestr 1–107
Pedruzzi I, Silva EAB, Rodrigues AE (2011) Production of lactobionic acid and sorbitol from lactose/fructose substrate using GFOR/GL enzymes from Zymomonas mobilis cells: a kinetic study. Enzyme Microb Technol 49:183–191
Malvessi E, Carra S, Pasquali FC et al (2013) Production of organic acids by periplasmic enzymes present in free and immobilized cells of Zymomonas mobilis. J Ind Microbiol Biotechnol 40:1–10
Chun UH, Rogers PL (1988) The simultaneous production of sorbitol from fructose and gluconic acid from glucose using an oxidoreductase of Zymomonas mobilis. Appl Microbiol Biotechnol 29:19–24
Rehr B, Wilhelm C, Sahm H (1991) Production of sorbitol and gluconic acid by permeabilized cells of Zymomonas mobilis. Appl Microbiol Biotechnol 35:144–148
Silveira MM, Malvessi E, Carra S et al (2007) Processo de produção e recuperação de sorbitol e ácidos orgânicos ou seus sais, preparação de elevada pureza isomérica de ácidos orgânicos ou seus sais. Pat Inven PI 0700421–0700424
Jang K-H, Jung S-J, Chang H-S, Chun U-H (1996) Improvement of the process for sorbitol production with Zymomonas mobilis immobilised in κ-carrageenan. Process Biochem 31:485–492
Wilberg KQ, Alves TLM, Nobrega R (1997) Enzymatic catalysis by permeabilized cells. Braz J Chem Eng 14:347–352
Malvessi E, Carra S, Silveira MM, Ayub MAZ (2010) Effect of substrate concentration, pH, and temperature on the activity of the complex glucose–fructose oxidoreductase/glucono-δ-lactonase present in calcium alginate-immobilized Zymomonas mobilis cells. Biochem Eng J 51:1–6
Wisbeck E, Ninow J, Jonas R, Silveira MM (1997) Evaluation of the flocculent strain Zymomonas mobilis Z1-81 for the production of sorbitol and gluconic acid. J Basic Microbiol 37:445–449
Silveira MM, Wisbeck E, Lemmel C et al (1999) Bioconversion of glucose and fructose to sorbitol and gluconic acid by untreated cells of Zymomonas mobilis. J Biotechnol 75:99–103
Ateş S, Mehmetoǧlu Ü (1997) A new method for immobilization of β-galactosidase and its utilization in a plug flow reactor. Process Biochem 32:433–436
Miner NA, McDowell JW, Willcockson GW et al (1977) Antimicrobial and other properties of a new stabilized alkaline glutaraldehyde disinfectant/sterilizer. Am J Hosp Pharm 34:376–382
Okuda K, Urabe I, Yamada Y, Okada H (1991) Reaction of glutaraldehyde with amino and thiol compounds. J Ferment Bioeng 71:100–105
Valle TA, Ruzza ÂA, Mastroeni Marco, Fabio Malvessi E et al (2013) Lactobionic acid produced by Zymomonas mobilis: alternative to prepare targeted nanoparticles. Pharm Anal Acta 4:3–7
Carra S, Cauzzi Rodrigues D, Moreno Conceição Beraldo N et al (2015) Reúso de células imobilizadas de Zymomonas mobilis para a produção de ácido lactobiônico. XX Simpósio Nacional de Bioprocessos
Malvessi E, Pasquali FC, Carra S et al (2006) Bioconversão de ácido lactobiônico por endoenzimas presentes em células imobilizadas de Zymomonas mobilis em diferentes configurações de biorreatores. VII Seminário Brasileiro de Tecnologia Enzimática
Malvessi E, Concatto K, Carra S, Silveira MM da (2006) Formulation of medium for growth and production of ethanol and intracellular enzymes by Zymomonas mobilis. Braz Arch Biol Technol 49:139–144
Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal Chem 31:426–428
Pedruzzi I, Malvessi E, Mata VG et al (2007) Quantification of lactobionic acid and sorbitol from enzymatic reaction of fructose and lactose by high-performance liquid chromatography. J Chromatogr A 1145:128–132
Obradovic B, Nedović VA, Bugarski B et al (2004) Immobilised cell bioreactors. Springer Netherlands, Dordrecht, pp 411–436
Margaritis A, Kilonzo PM (2005) Production of ethanol using immobilised cell bioreactor systems. In: Applications of cell immobilisation biotechnology. Springer, Berlin, pp 375–405
Nemati M, Webb C (2011) Immobilized cell bioreactors. In: Comprehensive biotechnology. Elsevier, Amsterdam, pp 331–346
Merck Ficha de Informações de Segurança de Produtos Químicos—FISPQ. http://www.merckmillipore.com/BR/pt/product/D(-)Fructose,MDA_CHEM-105323. Accessed 9 May 2017
European Pharmacopoeia Commission (2014) European directorate for the quality of medicines & healthcare, 8th edn
Silveira MM, Jonas R (2002) The biotechnological production of sorbitol. Appl Microbiol Biotechnol 59:400–408
Delagustin MG (2017) Caracterização e avaliação da estabilidade do ácido lactobiônico e de diferentes lactobionatos produzidos por Zymomonas mobilis visando à utilização na área farmacêutica. Diss Mestr 1–185
Acknowledgements
The authors are grateful to Coordenadoria de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), the Fundação de Amparo à Pesquisa do Estado do Rio Grande do Sul (FAPERGS) and the Universidade de Caxias do Sul (UCS) for the financial support to this work.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Folle, A.B., Baschera, V.M., Vivan, L.T. et al. Assessment of different systems for the production of aldonic acids and sorbitol by calcium alginate-immobilized Zymomonas mobilis cells. Bioprocess Biosyst Eng 41, 185–194 (2018). https://doi.org/10.1007/s00449-017-1856-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-017-1856-1