
Abstract
Cuticle melanism in insects is linked to a number of life history traits: a positive relationship is hypothesized between melanism, immune function, fecundity and lifespan. However, it is not clear how activation of the immune system affects trade-offs between life history traits in female mealworm beetles (Tenebrio molitor) differing in cuticle melanization. The females with tan, brown and black cuticles examined in the present study did not differ in the intensity of encapsulation response, fecundity and longevity when their immune system was not activated. However, we found that immune activation and cuticle melanization have a significant effect on life history traits. Offspring number and lifespan decreased in females with tan and brown cuticles, while the fecundity and lifespan of black females were not affected. Importantly, we inserted the implants again and found a significant decrease in the strength of encapsulation response in females with tan and brown cuticles. In contrast, black females increased melanotic reactions against the nylon implant, suggesting immunological priming. The results show that cuticle melanization plays an important adaptive role under the risk of being infected, while the lack of these benefits before the insertion of nylon monofilaments suggests that there are costs associated with an activated immunity system.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Melanin is responsible for a major part of the variation in coloration in both vertebrates and invertebrates (Fox 1976; McGraw 2006; Hsiung et al. 2015). This substance has been shown to be involved in a wide range of vital adaptive functions as diverse as camouflage (Kettlewell 1973; Hoekstra et al. 2006), photoprotection (Ortonne 2002), sexual signaling (Jawor and Breitwisch 2003; Svensson and Waller 2013), thermoregulation (Vences et al. 2002), protection against reactive oxygen species (Galván and Solano 2015) and strengthening insect cuticles (Riley 1997). The role of melanin pigmentation has also been demonstrated in immune defense (Wilson et al. 2001; Männiste and Hõrak 2014, but see Contreras-Garduño et al. 2007). For example, in invertebrates, a major aspect of the innate immune defense system against invading pathogens involves melanin. Within a few hours of infection, the invader is encapsulated in melanin layers (Nappi et al. 1995; Gillespie et al. 1997) and then dies by suffocation, or as a result of toxic compounds released by phenoloxidase (PO) activity (Siva-Jothy et al. 2005; Sugumaran 2002). Melanin itself has antimicrobial activity, which is likely to contribute to its effectiveness in defense (Montefiori and Zhou 1991; Montefiori et al. 1990; Sidibe et al. 1996) and wound healing (Sugumaran 2002).
The heritability of cuticle melanization in arthropods reaches 0.77 (Yurtsever 2000; Lee and Wilson 2006; Cotter et al. 2008; Singh et al. 2009; Roff and Fairbairn 2013). This suggests that the heritability of cuticle melanization is high enough to allow rapid evolutionary change in a phenotype. The relationship between cuticular melanization and several life history traits such as immune function, developmental time and fecundity has been studied extensively in insects. Immune defense is vital for an organism, as it reduces the deleterious impact of other infectious organisms. Many studies report a positive relationship between cuticular darkness and immune function. This includes PO activity (Wilson et al. 2001; Reeson et al. 1998; Cotter et al. 2004; Armitage and Siva-Jothy 2005; Bailey 2011), encapsulation response (Mikkola and Rantala 2010; Bailey 2011; Kivleniece et al. 2010, but see Dubovskiy et al. 2013a, b), hemocyte density (Cotter et al. 2004; Armitage and Siva-Jothy 2005), immune activation with an artificial parasite (Freitak et al. 2005), susceptibility to parasitoids (Wilson et al. 2001), fungal disease (Wilson et al. 2001; Barnes and Siva-Jothy 2000; Krams et al. 2013a) and viral disease (Reeson et al. 1998). However, there is also evidence for a negative relationship between cuticular melanization and immune traits such as antibacterial (lysozyme-like) activity (Cotter et al. 2004), hemocyte density (Rolff et al. 2005), PO activity (Rolff et al. 2005) and susceptibility to a viral disease (Goulson and Cory 1995).
A positive relationship between cuticular melanization and the strength of immune response suggests the concentration of cuticular melanin as an indicator of immune function, while the negative relationships indicate trade-offs between the efficiency of the immune system and cuticular melanin as predicted by life history theory (Stearns 1989). A number of studies have revealed the costs of immunity (Schmid-Hempel 2003; Sheldon and Verhulst 1996; Roff and Fairbairn 2013; see González-Santoyo and Córdoba-Aguilar 2012 for a review). Evidence shows that there are trade-offs between cuticle melanization and some fitness-related traits (Roff and Fairbairn 2013; Talloen et al. 2004). For example, darker individuals had a slower growth rate and exhibited larger wing asymmetry in a satyrine butterfly Pararge aegeria (Talloen et al. 2004). Melanic forms have been found to develop longer and weaker resistance against pathogens (Wilson et al. 2001; True 2003; Cotter et al. 2008; Wittkopp and Beldade 2009; Dubovskiy et al. 2013a, b). A recent report notably suggests that the condition-dependent component of melanin-based coloration is much stronger in invertebrates than vertebrates (Roulin 2015).
On the one hand, trade-offs between cuticle melanization, immune function and other traits may arise because melanin-based cuticular darkening and melanotic encapsulation response share the same melanin production pathway (González-Santoyo and Córdoba-Aguilar 2012). On the other hand, organisms exhibiting different intensities of cuticular melanization may be adapted to different environmental conditions with differing pathogen pressures and thus represent equally fit survival strategies (Galeotti et al. 2003). For example, if there is a linkage disequilibrium between genes coding for dark and pale cuticle coloration, these genes would code not only for a certain concentration of melanin, but also for some physiological features that allow a color morph to outcompete other morphs under specific ecological conditions (Ducrest et al. 2008). However, if the expression of genes coding for the alternate phenotypes is sensitive to environmental factors, individuals expressing different cuticle melanization would achieve a higher fitness only under specific environments. Such local adaptations may show up as a covariation between cuticle melanization and other life history traits only under highly specific conditions (Gonzales et al. 1999). Some studies found that environmental factors affect both the magnitude and the sign of covariations between coloration and life history traits in vertebrates (Gonzales et al. 1999; Fargallo et al. 2007; Piault et al. 2008; Roulin et al. 2008; Moore et al. 2014). Identifying the factors that mediate covariation between cuticle melanization, environmental factors and life history traits is important in order to determine the functional role of cuticle coloration.
Several studies on invertebrates have recently demonstrated the role of ecological conditions on trade-offs between immune function and fecundity (Zerofsky et al. 2005), availability of food (Moret and Schmid-Hempel 2000; Krams et al. 2014a, 2015), especially nitrogen-rich proteins, lifespan (Ye et al. 2009; Krams et al. 2013b) and larval competitive ability (Kraaijeveld and Godfray 1997). Selection for increased cuticular melanization in mealworm beetles Tenebrio molitor resulted in an increase in immune response (Armitage and Siva-Jothy 2005). Barnes and Siva-Jothy (2000) found that pathogen resistance is phenotypically plastic in T. molitor, where lower mortality, higher degree of cuticular melanization and stronger immune function were found in beetles reared at high larval densities. These findings support a crucial role of environmental conditions in general and pathogens and parasites in particular in life history trade-offs, showing that if there are costs involved with the maintenance of pathogen resistance, then higher investment in this trait is expected when the risk of pathogenesis is high (Wilson and Reeson 1998). Life history theory predicts different strategies in different sexes, where males are considered to be selected for mating rates, while females are thought to invest more in their immunity or longevity (Stearns 1992). Thus, higher investment in immunity and other fitness-related traits might be expected in females, especially if their coloration is dark, their environment contains pathogens and/or they have been affected by pathogens earlier. However, consistent evidence on positive relationships between cuticular melanization, the strength of immune response and female fitness is still missing.
In the current study, we examined the type of response that T. molitor females mount against the insertion of a nylon monofilament in their hemocoel. We also tested whether T. molitor females with dark cuticles invest in individual immune priming (Moret 2006; Kivleniece et al. 2010; Krams et al. 2011a; Mikonranta et al. 2014), lifespan and fecundity more than females with brown and tan cuticles. This is important because covariations between cuticle melanization and life history traits can sometimes be detected only under specific conditions, for instance, where one phenotype enjoys the existing ecological benefits while other ones are selected against (Gonzales et al. 1999; Piault et al. 2009; Roulin 2009; Moore et al. 2014).
Materials and methods
Insects
To avoid inbreeding effects (Polkki et al. 2012), we mixed beetles taken from a long-term, over 10-year laboratory population maintained at the University of Tennessee, Knoxville (60 %), with beetles obtained from Big Apple Pet Supply (30 %) (Boca Raton, FL, USA) and those obtained from a natural population (10 %). We used the next generation of beetles for this study, which was maintained on a diet of chick starter mash supplemented with occasional vegetables and fruit, such as carrots, apples and potatoes. The beetles were kept at 24 ± 2 °C. We removed pupae from the culture on the day of pupation. They were weighed and their sex was determined by examining genitalia on the eighth abdominal segment (Bhattacharya et al. 1970). The pupae and newly emerged adult females were kept individually in 200 ml plastic containers filled with food ad libitum. Only individuals with no visible abnormalities were used in the experiments. All of the experimental trials were performed in winter 2014/2015.
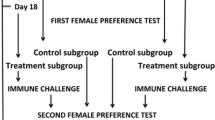
Study design
We weighed 12 days old virgin females to the nearest 0.1 mg (mean body weight ± SD = 109.46 ± 7.02 mg). The beetles were randomly assigned to experimental and control groups (Fig. 1). In the treatment group, each female was placed on ice and received one sterile nylon monofilament implant (2 mm length, 0.18 mm diameter, knotted at one end) through their pleural membrane between the third and fourth abdominal sternite (Rantala et al. 2002; Krams et al. 2011a, b; Daukšte et al. 2012) for 6 h at 24 ± 0.5 °C. Females of the control group were handled similarly, but their cuticle was not punctured and these animals were not implanted.

The experimental protocol used to study immune response, fecundity and lifespan of female mealworm beetles
The treatment females were further divided into three subgroups: breeding, repeated implantation and survival subgroups (Fig. 1). The control group was divided into two subgroups: breeding and survival subgroups. Each of these subgroups (3 subgroups in the treatment group and 2 subgroups in the control group) consisted of females of three different types of cuticle (elytra) coloration: ‘black’, ‘brown’ or ‘tan’ (Figs. 1, 2).

The elytra melanization classes (tan, brown, black) of female mealworm beetles (color figure online)
Treatment group, breeding subgroup: on day 17 after imaginal eclosion (5 days after the insertion and removal of the implants), we placed two males to each of 30 females of each elytra color class (90 individuals in total) for 24 h. These females were left alone in their boxes to lay eggs for 7 days. We counted the number of larvae on day 55 after imaginal eclosion (Fig. 1).
Treatment group, survival subgroup: after the removal of the implants, 30 females of each color class (90 individuals in total) were provided with bran and fresh apple. Their survival was verified on a daily basis. These females were not allowed to reproduce (Fig. 1).
Treatment group, repeated implantation subgroup: 30 females of each of elytra color class (90 individuals in total) received the second nylon implant for 6 h 5 days after the removal of the first implant (day 17 after imaginal eclosion). In this way, we checked whether the first activation of the immune system resulted in an increased immune response 5 days later and whether this possible individual immune priming was related to cuticle melanization (Fig. 1).
Control group, breeding subgroup: 30 females of each elytra color group (90 individuals in total) were paired with two males on day 17 after imaginal eclosion for 24 h, and we counted offspring number on day 55 after imaginal eclosion (Fig. 1).
Control group, survival subgroup: 30 females of each elytra color group (90 individuals in total) were kept separately with food ad libitum and we checked for their survival until the last individual died (Fig. 1).
Cuticle coloration
To assess the cuticle melanization, we followed the recommendations by Barnes and Siva-Jothy (2000). In brief, we assessed the beetle coloration under a Nikon stereo microscope with LED illumination. ‘Tan’ females were easy to tell from the rest of the groups, because their elytra was light brown (Fig. 2). The elytra of ‘brown’ beetles was dark brown and easily distinguishable from ‘tan’ beetles (Fig. 2). Discrimination of ‘brown’ and ‘black’ beetles was done by including into the ‘black’ group only females with no traces of brown coloration to their elytra even under higher zoom positions (Fig. 2). We also took digital images of the beetles and analyzed them using image analysis software (Image J, Abramoff et al. 2004). The elytra melanization was expressed as a grayscale value between 0 (white) and 240 (black). We found a significant effect of coloration on the darkness grayscale values obtained (ANOVA: df = 2, F = 3628.10, P < 0.0001, Fig. 2). The grayscale values were significantly different between each elytra color category (‘tan’ = 135.40 ± 11.45, ‘brown’ = 188.52 ± 6.98, ‘black’ = 223.25 ± 7.93) (Tukey tests: all P < 0.001, Fig. 3).

Median cuticle melanization in three elytra color classes of mealworm beetles; the box indicates the 25–75 % percentiles, the whiskers indicate the min–max range, the numbers indicate the sample size
Immune assays
To quantify the strength of encapsulation response to a foreign body, we analyzed the lightness of each nylon insert. It is widely acknowledged that insect immune systems respond to the insert by attempting to encapsulate the foreign body as though it were a parasitoid or fungal invasion (e.g., Rantala et al. 2000; Dubovskiy et al. 2013a, b). The resulting melanization correlates with the level of immune system response (e.g., Sadd et al. 2006; Krams et al. 2011a, 2013a, b; Rantala et al. 2000, 2002). The ability to encapsulate a synthetic substrate is also positively related to the encapsulation of parasites (Paskewitz and Riehle 1994; Gorman et al. 1998) and to the ability to resist an entomopathogenic fungal disease (Rantala and Roff 2007; Dubovskiy et al. 2013a, b; Krams et al. 2013a). Overall, high levels of melanization or darkening of the inserts indicates increased levels of immune system activity and response (Yourth et al. 2001, 2002; Krams et al. 2013a, b). However, it is important to note that a number of studies failed to find a positive correlation between melanization of artificial inserts and the ability to encapsulate real parasites (e.g., Schwartz and Koella 2002; Mallon et al. 2003; Honkavaara et al. 2009; Nagel et al. 2014).
To quantify lightness, we photographed each of the removed inserts from two directions under constant light conditions using a Nikon stereo microscope. We then analyzed the digital images using image analysis software (Image J, Abramoff et al. 2004). We marked the area of that portion of the insert that had been within the beetle’s body and the program calculated the lightness value. Since increasing melanization indicated a stronger immune response in this study, we calibrated reflectance of an implant before the insertion to zero level.
Statistics
The strength of the encapsulation response was distributed normally in all groups (Kolmogorov–Smirnov tests: all P > 0.20). We used ANOVA to test differences in offspring numbers across control and treatment groups and elytra melanization classes. Lifespan was analyzed using a Cox proportional hazards model, while we used a linear mixed model with female identity as a random factor to find possible treatment and elytra color effects on the activation of the immune system. All statistical tests used in this study were two-tailed.
Results
Offspring number
Cuticle darkness had no effect on offspring number in the control group (F2,87 = 0.29, P > 0.05, Fig. 4). In contrast, we found a significant effect of cuticle melanization on offspring number after the activation of the immune system in beetles of the treatment group, revealed as cuticle melanization group and treatment interaction (F2,174 = 20.22, P < 0.001, Fig. 4). All three cuticle melanization groups differed significantly from each other in the number of offspring produced after the insertion of the nylon monofilament. The offspring number of tan females was smaller than that of brown females (Tukey HSD: P = 0.004), and brown females produced less offspring than black females (P < 0.001, Fig. 4). The decrease in the number of offspring was found to be greatest among tan females, while the fecundity of black females was not impaired by the activation of their immunity system: the offspring number of the black females of the treatment group did not differ from that of the black females in the control group (Tukey HSD: P > 0.05, Fig. 4).

Average offspring number (±95 % CIs) of three elytra color classes in the control (open circles) and treatment (black circles) groups of mealworm beetles
Lifespan
Cuticle color was not related to the longevity of females in the control group (χ 2 = 0.90, P > 0.05, Fig. 5). In the treatment group, the insertion of implants significantly decreased lifespan, as revealed by a significant interaction between elytra color and treatment (χ 2 = 48.92, P < 0.001, Fig. 5). While the activation of the immunity system via the insertion of the nylon monofilament did not affect the lifespan of the females with black elytra (z = −0.03, P > 0.05), the implantation significantly decreased the lifespan of the females with tan (z = 5.29, P < 0.001) and brown cuticles (z = 6.85, P < 0.001, Fig. 5).

Average lifespan (±95 % CIs) of three elytra color classes in the control (open circles) and treatment (black circles) groups of mealworm beetles
Encapsulation response
The first activation of the immune system via the insertion of the nylon monofilament did not show any difference in the intensity of implant melanization among the female groups (ANOVA: F2,87 = 1.19, P > 0.05, Fig. 6). The second implantation reflects the investment into encapsulation ability done by females between the first and second implantations. We found the encapsulation response to be significantly different among cuticle color groups (F2,122 = 63, P < 0.001, Fig. 6). Females with black elytra increased the strength of their encapsulation response (t122 = 2.34, P = 0.021), suggesting immune priming of melanotic reactions. In contrast, the strength of the encapsulation response decreased in females with brown (t122 = −9.92, P < 0.001) and tan cuticles (t122 = −9.53, P < 0.001).

Strength of encapsulation response against the first and second implantation in tan, brown and black elytra color classes of female mealworm beetles
Discussion
Life history theory asserts that the schedule and duration of key events in an organism’s lifetime are shaped by natural selection to produce the largest possible number of surviving offspring (Stearns 1992). Males of many species tend to increase the number of copulations (Bateman 1948), and in case of terminal investment in reproduction male individuals may increase their sexual attractiveness at the expense of their longevity (Krams et al. 2014b, 2015). In females, fitness is often positively linked to lifespan (Trivers 1972), while longevity largely depends on investment into immune function (Lin et al. 1998; Krams et al. 2014a).
It has been recently shown that melanin pigmentation is linked with the ability to cope with infections: darker melanic individuals usually have a lower infection intensity and a greater immune response than paler individuals (Jacquin et al. 2011; Prokkola et al. 2013). Darker melanic individuals can even reduce the fecundity of parasites (Roulin et al. 2001). Two hypotheses have been suggested to explain the covariation between the intensity of melanin pigmentation and the expression of life history traits. The genetic link hypothesis states that positive links between melanin-based coloration and the strength of immune function could be explained by the pleiotropic effects of genes coding for melanin pigmentation on the immune system (Ducrest et al. 2008; Gasparini et al. 2009). The exposure hypothesis posits that melanin-based pigmentation and immunity are linked, because melanin pigmentation develops and may be exploited in habitats that differ in parasite exposure (Galeotti and Rubolini 2004; Roulin 2004). In T. molitor, beetles reared at high larval densities showed a higher degree of cuticular melanization and lower mortality against an entomopathogenic fungus than those reared solitarily (Barnes and Siva-Jothy 2000). Larvae and pupae of T. molitor can assess conspecific densities using both mechanical and chemical cues (Tschinkel and Willson 1971; Kotaki and Fujii 1995). This indicates that perceived pathogen exposure did not differ among T. molitor individuals used in this study, because all the beetles grew under similar larval densities. Thus, our results are likely to support the genetic link hypothesis, because females with black elytra showed signs of immune priming, while tan and brown females mounted a weaker encapsulation upon second challenge. Furthermore, these effects cannot be attributed to a density-dependent prophylactic response to rearing density.
The genetic link and the exposure hypotheses are considered to be competing explanations of melanism. Disentangling their mechanisms is important for understanding the adaptive function of melanin-based pigmentation. However, our results cannot provide any decisive evidence on the distinction of the two hypotheses. First of all, the strength of the immune response, the number of offspring and lifespan did not differ among females with different cuticle melanization before the activation of the immune system, suggesting costs of cuticle coloration. The benefits of the dark cuticle appeared only when the immune responses against artificial infection were induced. This demonstrates that the innate immunity of adult insects may be adjusted to changes in the risk of infection by individual priming, which increased the lifespan of the females with black cuticles. This provides support for the parasite exposure hypothesis, since T. molitor females needed an environmental cue to prime their innate immunity. Thus, the exposure and the genetic link hypotheses might not be mutually exclusive explanations of the adaptive role of cuticle melanization in T. molitor. To provide further tests for these hypotheses, different levels of larval densities and parasite exposures should be manipulated in future work. This is important in order to test whether darker or paler cuticles are associated with adaptations to one highly specific environment and represent the lack of adaptations in other circumstances (Kawecki and Ebert 2004). This is a key aspect when considering the role of environmental heterogeneity in the maintenance of polymorphism (Piault et al. 2009; Roulin 2009).
It is known that melanogenesis involves the formation of melanin pigments and toxic by-products from the action of PO on quinone precursors, and occurs primarily in the cuticular structures, midgut epithelium and hemolymph (Cerenius and Söderhäll 2004). However, the strength of the encapsulation response did not differ between black and paler females before their immune system was activated via the insertion of the nylon monofilament. The absence of excess activation in females with black elytra can be explained by energetic costs and oxidative stress of permanently activated immune system (Freitak et al. 2003; Krams et al. 2014b). The damaging action of chemical radicals produced during the activation of the immune response against intruders can also harm the host cells and tissues (von Schantz et al. 1999; Finkel and Holbrook 2000; Metcalfe and Alonso-Alvarez 2010). Melanic insect morphs exhibit an unusually high concentration of cuticular melanin (Barnes and Siva-Jothy 2000; Wilson et al. 2001, 2002), and the immune system of black females potentially generates too harmful a response. It is probably more adaptive for the immune system to avoid the permanent harm to self and respond just when the host is under attack by infection, even if the response is delayed.
In insects, melanin pigments and their precursors are important as structural and protective components of the cuticle. In the greater wax moth (Galleria mellonella), the cuticle of melanic larvae is shown to be substantially thicker than in a non-melanic morph. The cuticle of melanic larvae of the greater wax moth can generate a short burst of enhanced cuticular PO activity during the early stages of fungal penetration (Dubovskiy et al. 2013a, b). A thicker cuticle and slower penetration of the intruder allows sufficient time for the insect to activate its defenses, such as encapsulation of the intruder (Sweeney et al. 1983; Butt et al. 1988). It is important to note that a high concentration of cuticle melanin plays a significant role in wound healing after damage by intruders, as seen in mosquito midguts following penetration by the malaria parasite (Shiao et al. 2006). This might be especially important in explaining the greater offspring number and the longer lifespan of black T. molitor females after the activation of the immune system. However, future research is needed to test whether dark and pale T. molitor differs with respect to wound healing ability, the energetic costs of wound healing and whether any possible differences in wound healing are responsible for the higher fitness of the darker female T. molitor beetles.
In this study, we did not find any costs related to melanin pigmentation of the females with black elytra. However, a number of studies revealed many trade-offs between immunity and other life history traits in T. molitor. Also in the greater wax moth, the heavy defense investments made by melanic insects result in a lower body mass, decreased longevity and lower fecundity in comparison with the non-melanic morph. One possible explanation for the similar fecundity of darker and paler females before the activation of the immune system is that the egg-laying period of brown and tan females was longer than that of black females, and exceeded the period of 7 days that was allowed to oviposit.
To conclude, the present study shows some benefits that the melanized cuticle brings to T. molitor females. While fecundity, the strength of encapsulation response and longevity do not differ between darker and paler individuals, highly melanized females can prime their immune response, increase their lifespan and not decrease their fecundity upon parasitoid-like attacks (here, the insertion of the nylon implant). Although cuticular darkening and encapsulation response may compete for the same limiting resources necessary for melanin synthesis, such as tyrosine, we did not find any costs associated with immune response, fecundity and longevity in females with black elytra. The availability of food is an important predictor of survival and reproductive strategies in T. molitor (Krams et al. 2015). However, the access to tyrosine, a food-derived melanin precursor, was likely the same for dark and pale females in this study. It is possible that female beetles differed in their ability and efficiency to transform tyrosine and properly invest it into cuticular and immunity-related melanin. We also do not know whether melanin deposited in cuticle could be re-invested into immediate immune response. Importantly, relationships between cuticle darkening, fecundity, immune response and longevity of T. molitor females cannot be estimated without finding possible trade-offs between these life history traits. Cuticle melanization provides a widespread source of pigmentation in insects, yet the relationship between cuticle melanin and its use in immune response appears to be more complex than previously thought.
Our results emphasize the fact that the benefit of being melanic is accrued only under specific environmental conditions, a phenomenon observed in several vertebrates (e.g., Gonzales et al. 1999; Piault et al. 2009). This raises the possibility that the covariation between the degree of melanism and other phenotypes can be detected only under specific conditions (Roulin 2009). Finally, our study shows that males and females may differ in trade-offs between parameters involved in immune response and reproductive strategies, which suggests a more important role of sex- and hormone-related regulation of immune function and senescence in T. molitor.
References
Abramoff MD, Magalhaes PJ, Ram SJ (2004) Image processing with ImageJ. Biophotonics Int 11:36–42
Armitage SAO, Siva-Jothy MT (2005) Immune function responds to selection for cuticular colour in Tenebrio molitor. Heredity 94:650–656
Bailey NW (2011) A test of the relationship between cuticular melanism and immune function in wild-caught Mormon crickets. Physiol Entomol 36:155–164
Barnes AI, Siva-Jothy MT (2000) Density-dependent prophylaxis in the mealworm beetle Tenebrio molitor L. (Coleoptera: Tenebrionidae): cuticular melanization is an indicator of investment in immunity. Proc R Soc B 267:177–182
Bateman AJ (1948) Intra-sexual selection in Drosophila. Heredity 2:349–368
Bhattacharya AK, Ameel JJ, Waldbauer GP (1970) Method for sexing living pupal and adult yellow mealworms. Ann Entomol Soc Am 63:1783
Butt TM, Wraight SP, Galaini-Wraight S, Humber RA, Roberts DW, Soper RS (1988) Humoral encapsulation of the fungus Erynia radicans (Entomophthorales) by the potato leafhopper, Empoasca fabae (Homoptera: Cicadellidae). J Invertebr Pathol 52:49–56
Cerenius L, Söderhäll K (2004) The prophenoloxidase-activating system in invertebrates. Immunol Rev 198:116–126
Contreras-Garduño J, Buzatto BA, Abundis L, Najera-Cordero K, Córdoba-Aguilar A (2007) Wing colour properties do not reflect male condition in the American rubyspot (Hetaerina americana). Ethology 113:944–952
Cotter SC, Hails RS, Cory JS, Wilson K (2004) Density-dependent prophylaxis and condition-dependent immune function in Lepidopteran larvae: a multivariate approach. J Anim Ecol 73:283–293
Cotter SC, Myatt JP, Benskin CMH, Wilson K (2008) Selection for cuticular melanism reveals immune function and life-history trade-offs in Spodoptera littoralis. J Evol Biol 21:1744–1754
Daukšte J, Kivleniece I, Krama T, Rantala MJ, Krams I (2012) Senescence in immune priming and attractiveness in a beetle. J Evol Biol 25:1298–1304
Dubovskiy IM, Whitten MMA, Yaroslavtseva ON, Kryukov VY, Grizanova EV, Mukherjee K, Vilcinskas A, Glupov VV, Butt TM (2013a) Can insects develop resistance to insect pathogenic fungi? PLoS One 8:e60248
Dubovskiy IM, Whitten MMA, Kryukov VY, Yaroslavtseva ON, Grizanova EV, Greig C, Mukherjee K, Vilcinskas A, Mitkovets PV, Glupov VV, Butt TM (2013b) More than a colour change: insect melanism, disease resistance and fecundity. Proc R Soc B 280:20130584
Ducrest AL, Keller L, Roulin A (2008) Pleiotropy in the melanocortin system, coloration and behavioural syndromes. Trends Ecol Evol 23:502–510
Fargallo JA, Laaksonen T, Korpimäki E, Wakamatsu K (2007) A melanin-based trait reflects environmental growth conditions of nestling male Eurasian kestrels. Evol Ecol 21:157–171
Finkel T, Holbrook NJ (2000) Oxidants, oxidative stress and the biology of ageing. Nature 408:239–247
Fox DL (1976) Animal biochromes and structural colours: physical, chemical, distributional & physiological features of coloured bodies in the animal world, 2nd edn. University of California Press, Berkeley
Freitak D, Ots I, Vanatoa A, Hõrak P (2003) Immune response is energetically costly in white cabbage butterfly pupae. Proc R Soc B 270:S220–S222
Freitak D, Vanatoa A, Ots I, Rantala MJ (2005) Formation of melanin-based wing patterns is influenced by condition and immune challenge in Pieris brassicae. Entomol Exp Appl 116:237–243
Galeotti P, Rubolini D (2004) The niche variation hypothesis and the evolution of colour polymorphism in birds: a comparative study of owls, nightjars and raptors. Biol J Linn Soc 82:237–248
Galeotti P, Rubolini D, Dunn PO, Fasola M (2003) Colour polymorphism in birds: causes and functions. J Evol Biol 16:635–646
Galván I, Solano F (2015) Melanin chemistry and the ecology of stress. Physiol Biochem Zool 88:352–355
Gasparini J, Bize P, Piault R, Wakamatsu K, Blount JD, Ducrest AL, Roulin A (2009) Strength and cost of an induced immune response are associated with a heritable melanin-based colour trait in female tawny owls. J Anim Ecol 78:608–616
Gillespie JP, Kanost MR, Trenczek T (1997) Biological mediators of insect immunity. Ann Rev Entomol 42:611–643
Gonzales G, Sorci G, de Lope F (1999) Seasonal variation in the relationship between cellular immune response and badge size in male house sparrows (Passer domesticus). Behav Ecol Sociobiol 46:117–122
González-Santoyo I, Córdoba-Aguilar A (2012) Phenoloxidase: a key component of the insect immune system. Entomol Exp Appl 142:1–16
Gorman MJ, Schwartz AM, Paskewitz SM (1998) The role of surface characteristics in eliciting humoral encapsulation of foreign bodies in Plasmodium-refractory and-susceptible strains of Anopheles gambiae. J Insect Physiol 44:947–954
Goulson D, Cory JS (1995) Responses of Mamestra brassicae (Lepidoptera, Noctuidae) to crowding—interactions with disease, color phase and growth. Oecologia 104:416–423
Hoekstra HE, Hirschmann RJ, Bundey RA, Insel PA, Crossland JP (2006) A single amino acid mutation contributes to adaptive beach mouse color pattern. Science 313:101–104
Honkavaara J, Rantala MJ, Suhonen J (2009) Mating status, immune defence, and multi-parasite burden in the damselfly Coenagrion armatum. Entomol Exp Appl 132:165–171
Hsiung B-K, Blackledge TA, Shawkey MD (2015) Spiders do have melanin after all. J Exp Biol 218:3632–3635
Jacquin L, Lenouvel P, Haussy C, Ducatez S, Gasparini J (2011) Melanin-based coloration is related to parasite intensity and cellular immune response in an urban free living bird: the feral pigeon Columba livia. J Avian Biol 42:11–15
Jawor JM, Breitwisch R (2003) Melanin ornaments, honesty, and sexual selection. Auk 120:249–265
Kawecki TJ, Ebert D (2004) Conceptual issues in local adaptation. Ecol Lett 7:1225–1241
Kettlewell HBD (1973) The evolution of melanism. Clarendon Press, Oxford
Kivleniece I, Krams I, Daukste J, Krama T, Rantala MJ (2010) Sexual attractiveness of immune-challenged male mealworm beetles suggests terminal investment in reproduction. Anim Behav 80:1015–1021
Kotaki T, Fujii H (1995) Crowding inhibits pupation in Tribolium freemani: contact chemical and mechanical stimuli are involved. Entomol Exp Appl 74:145–149
Kraaijeveld AR, Godfray HCJ (1997) Trade-off between parasitoid resistance and larval competitive ability in Drosophila melanogaster. Nature 389:278–280
Krams I, Daukšte J, Kivleniece I, Krama T, Rantala MJ, Ramey G, Šauša L (2011a) Female choice reveals terminal investment in male mealworm beetles, Tenebrio molitor, after a repeated activation of the immune system. J Insect Sci 11:56
Krams I, Daukšte J, Kivleniece I, Krama T, Rantala MJ (2011b) Overwinter survival depends on immune defence and body length in male Aquarius najas water striders. Entomol Exp Appl 140:45–51
Krams I, Daukšte J, Kivleniece I, Krama T, Rantala MJ (2013a) Previous encapsulation response enhances within individual protection against fungal parasite in the mealworm beetle Tenebrio molitor. Insect Sci 20:771–777
Krams I, Daukšte J, Kivleniece I, Kaasik A, Krama T, Freeberg TM, Rantala MJ (2013b) Trade-off between cellular immunity and life span in mealworm beetles Tenebrio molitor. Curr Zool 59:340–346
Krams I, Kecko S, Kangassalo K, Moore KF, Jankevics E, Inashkina I, Krama T, Lietuvietis V, Meija L, Rantala MJ (2014a) Effects of food quality on trade-offs among growth, immunity and survival in the greater wax moth (Galleria mellonella). Insect Sci 22:431–439
Krams I, Krama T, Moore FR, Kivleniece I, Kuusik A, Freeberg TM, Mänd R, Rantala MJ, Daukšte J, Mänd M (2014b) Male mealworm beetles increase resting metabolic rate under terminal investment. J Evol Biol 27:541–550
Krams I, Moore F, Krama T, Rantala MJ, Mänd R, Mierauskas P, Mänd M (2015) Resource availability as a proxy for terminal investment in a beetle. Oecologia 178:339–345
Lee KP, Wilson K (2006) Melanism in a larval Lepidoptera: repeatability and heritability of a dynamic trait. Ecol Entomol 31:196–205
Lin YJ, Seroude L, Benzer S (1998) Extended life-span and stress resistance in the Drosophila mutant methuselah. Science 282:943–946
Mallon EB, Loosli R, Schmid-Hempel P (2003) Specific versus nonspecific immune defense in the bumblebee, Bombus terrestris L. Evolution 57:1444–1447
Männiste M, Hõrak P (2014) Emerging infectious disease selects for darker plumage coloration in greenfinches. Front Ecol Evol 2:4
McGraw KJ (2006) The mechanics of melanin coloration in birds. In: Hill GE, McGraw KJ (eds) Bird coloration. I. Mechanisms and measurements. Harvard University Press, Cambridge, MA, pp 243–294
Metcalfe NB, Alonso-Alvarez C (2010) Oxidative stress as a life-history constraint: the role of reactive oxygen species in shaping phenotypes from conception to death. Funct Ecol 24:984–996
Mikkola K, Rantala MJ (2010) Immune defence, a possible nonvisual selective factor behind the industrial melanism of moths (Lepidoptera). Biol J Linn Soc 99:831–838
Mikonranta L, Mappes J, Kaukoniitty M, Freitak D (2014) Insect immunity: oral exposure to a bacterial pathogen elicits free radical response and protects from a recurring infection. Front Zool 11:23
Montefiori DC, Zhou JY (1991) Selective antiviral activity of synthetic soluble l-tyrosine and L-DOPA melanins against human-immunodeficiency-virus in vitro. Antiviral Res 15:11–26
Montefiori DC, Modliszewski A, Shaff DI, Zhou JY (1990) Inhibition of human-immunodeficiency-virus type-1 replication and cytopathecity by synthetic soluble catecholamine melanins in vitro. Biochem Biophys Res Commun 168:200–205
Moore FR, Cīrule D, Kivleniece I, Vrublevska J, Rantala MJ, Sild E, Sepp T, Hõrak P, Krama T, Krams I (2014) Investment in a sexual signal results in reduced survival under extreme conditions in the male great tit (Parus major). Behav Ecol Sociobiol 69:151–158
Moret Y (2006) Trans-generational immune priming: specific enhancement of the antimicrobial immune response in the mealworm beetle, Tenebrio molitor. Proc R Soc B 273:1399–1405
Moret Y, Schmid-Hempel P (2000) Survival for immunity: the price of immune system activation for bumblebee workers. Science 290:1166–1168
Nagel L, Mlynarek JJ, Forbes MR (2014) Comparing natural parasitism and resistance with proxies of host immune response in Lestid damselflies. Ecol Parasitol Immunol 3:art235884
Nappi AJ, Vass E, Frey F, Carton Y (1995) Superoxide anion generation in Drosophila during melanotic encapsulation of parasites. Eur J Cell Biol 68:450–456
Ortonne J-P (2002) Photoprotective properties of skin melanin. Br J Dermatol 146:7–10
Paskewitz S, Riehle MA (1994) Response of Plasmodium refractory and susceptible strains of Anopheles gambiae to inoculated Sephadex beads. Dev Comp Immunol 18:369–375
Piault R, Gasparini J, Bize P, Paulet M, McGraw KJ, Roulin A (2008) Experimental support for the makeup hypothesis in nestling tawny owls (Strix aluco). Behav Ecol 19:703–709
Piault R, Gasparini J, Bize P, Jenni-Eiermann S, Roulin A (2009) Pheomelanin-based coloration and the ability to cope with variation in food supply and parasitism. Am Nat 174:548–556
Polkki M, Krams I, Kangassalo K, Rantala MJ (2012) Inbreeding affects sexual signalling in males but not females of Tenebrio molitor. Biol Lett 8:423–425
Prokkola J, Roff D, Kärkkäinen T, Krams I, Rantala MJ (2013) Genetic and phenotypic relationships between immune defense, melanism and life-history traits at different temperatures and sexes in Tenebrio molitor. Heredity 111:89–96
Rantala MJ, Roff DA (2007) Inbreeding and extreme outbreeding cause sex differences in immune defence and life history traits in Epirrita autumnata. Heredity 98:329–336
Rantala MJ, Koskimaki J, Taskinen J, Tynkkynen K, Suhonen J (2000) Immunocompetence, developmental stability and wingspot size in the damselfly Calopteryx splendens L. Proc R Soc B 267:2453–2457
Rantala MJ, Jokinen I, Kortet R, Vainikka A, Suhonen J (2002) Do pheromones reveal male immunocompetence? Proc R Soc B 269:1681–1685
Reeson AF, Wilson K, Gunn A, Hails RS, Goulson D (1998) Baculovirus resistance in the noctuid Spodoptera exempta is phenotypically plastic and responds to population density. Proc R Soc B 265:1787–1791
Riley PA (1997) Melanin. Int J Biochem Cell Biol 29:1235–1239
Roff DA, Fairbairn DJ (2013) The costs of being dark: the genetic basis of melanism and its association with fitness-related traits in the sand cricket. J Evol Biol 26:1406–1416
Rolff J, Armitage SAO, Coltman DW (2005) Genetic constraints and sexual dimorphism in immune defense. Evolution 59:1844–1850
Roulin A (2004) The evolution, maintenance and adaptive function of genetic colour polymorphism in birds. Biol Rev 79:1–34
Roulin A (2009) Covariation between eumelanic pigmentation and body mass only under specific conditions. Naturwissenschaften 96:375–382
Roulin A (2015) Condition-dependence, pleiotropy and the handicap principle of sexual selection in melanin-based colouration. Biol Rev 91:328–348
Roulin A, Riols C, Dijkstra C, Ducrest A-L (2001) Female plumage spottiness signals parasite resistance in the barn owl (Tyto alba). Behav Ecol 12:103–110
Roulin A, Gasparini J, Bize P, Ritschard M, Richner H (2008) Melanin-based colorations signal strategies to cope with poor and rich environments. Behav Ecol Sociobiol 62:507–519
Sadd B, Holman H, Armitage F, Lock R, Marland R, Siva-Jothy MT (2006) Modulation of sexual signalling by immune challenged male mealworm beetles (Tenebrio molitor, L): evidence for terminal investment and dishonesty. J Evol Biol 19:321–325
Schmid-Hempel P (2003) Variation in immune defence as a question of evolutionary ecology. Proc R Soc B 270:357–366
Schwartz A, Koella JC (2002) Melanization of Plasmodium falciparum and C-25 sephadex beads by field-caught Anopheles gambiae (Diptera: Culicidae) from southern Tanzania. J Med Entomol 39:84–88
Sheldon BC, Verhulst S (1996) Ecological immunology: costly parasite defences and trade-offs in evolutionary ecology. Trends Ecol Evol 11:317–321
Shiao S-H, Whitten MMA, Zachary D, Hoffmann JA, Levashina EL (2006) Fz2 and Cdc42 mediate melanization and actin polymerization but are dispensable for Plasmodium killing in the mosquito midgut. PLoS Pathog 2:e133
Sidibe S, Saal F, Rhodes-Feuillette A, Lagaye S, Pelicano L, Canivet M, Peries J, Dianoux L (1996) Effects of serotonin and melanin on in vitro HIV-1 infection. J Biol Regul Homeost Agents 10:19–24
Singh S, Ramniwas S, Parkash R (2009) Fitness consequences of body melanization in Drosophila immigrans from montane habitats. Entomol Res 39:182–191
Siva-Jothy MT, Moret Y, Rolff J (2005) Insect immunity: an evolutionary ecology perspective. In: Simpson SJ (ed) Advances in insect physiology, vol 32. Academic Press Inc, San Diego, pp 1–48
Stearns SC (1989) Trade-offs in life-history evolution. Funct Ecol 3:259–268
Stearns SC (1992) The evolution of Life histories. Oxford University Press, Oxford
Sugumaran H (2002) Comparative biochemistry of eumelanogenesis and the protective roles of phenoloxidase and melanin in insects. Pigment Cell Res 15:2–9
Svensson EI, Waller JT (2013) Ecology and sexual selection: evolution of wing pigmentation in Calopterygid damselflies in relation to latitude, sexual dimorphism, and speciation. Am Nat 182:174–195
Sweeney AW, Inman AO, Bland CE, Wright RG (1983) The fine structure of Culicinomyces clavisporus invading mosquito larvae. J Invertebr Pathol 42:224–243
Talloen W, Van Dyck H, Lens L (2004) The cost of melanization: butterfly wing coloration under environmental stress. Evolution 58:360–366
Trivers RL (1972) Parental investment and sexual selection. In: Campbell B (ed) Sexual selection and the descent of man, 1871–1971. Aldine, Chicago, pp 136–179
True JR (2003) Insect melanism: the molecules matter. Trends Ecol Evol 18:640–647
Tschinkel WR, Willson CD (1971) Inhibition of pupation due to crowding in some tenebrionid beetles. J Exp Biol 176:137–146
Vences M, Galán P, Vieites DR, Puente M, Oetter K, Wanke S (2002) Field body temperatures and heating rates in a montane frog population: the importance of black dorsal pattern for thermoregulation. Ann Zool Fenn 39:209–220
von Schantz T, Bensch S, Grahn M, Hasselquist D, Wittzell H (1999) Good genes, oxidative stress and condition-dependent sexual signals. Proc R Soc Lond B 266:1–12
Wilson K, Reeson AF (1998) Density-dependent prophylaxis: Evidence from Lepidoptera–baculovirus interactions? Ecol Entomol 23:100–101
Wilson K, Cotter SC, Reeson AF, Pell JK (2001) Melanism and disease resistance in insects. Ecol Lett 4:637–649
Wilson K, Thomas MB, Blanford S, Doggett M, Simpson SJ, Moore SL (2002) Coping with crowds: density-dependent disease resistance in desert locusts. Proc Natl Acad Sci USA 99:5471–5475
Wittkopp PJ, Beldade P (2009) Development and evolution of insect pigmentation: genetic mechanisms and the potential consequences of pleiotropy. Semin Cell Dev Biol 20:65–71
Ye YXH, Chenoweth SF, McGraw EA (2009) Effective but costly, evolved mechanisms of defense against a virulent opportunistic pathogen in Drosophila melanogaster. PLoS Pathog 5:9
Yourth CP, Forbes MR, Smith BP (2001) On understanding variation in immune expression of the damselflies Lestes spp. Can J Zool 79:815–821
Yourth CP, Forbes MR, Smith BP (2002) Immune expression in a damselfly is related to time of season, not to fluctuating asymmetry or host size. Ecol Entomol 27:123–128
Yurtsever S (2000) On the polymorphic meadow spittlebug, Philaenus spumarius (Homoptera: Cercopidae). Turk J Zool 24:447–459
Zerofsky M, Harel E, Silverman N, Tatar M (2005) Aging of the innate immune response in Drosophila melanogaster. Aging Cell 4:103–108
Acknowledgments
We thank Prof. Christine R.B. Boake and Prof. Todd M. Freeberg for their support during all phases of this study. Professors Jae H. Park, Mariano Labrador, Ranjan Ganguly and Joshua N. Bembenek kindly provided access to their laboratory facilities. We also thank Prof. Alexandre Roulin and one anonymous reviewer for valuable comments.
Author contribution statement
IK, T.K. and M.J.R. conceived and designed the study and participated in the drafting of the manuscript. I.K., T.K. R.K., S.L. and G.T. performed the study, and collected and extracted the data. A.K., G.T., G.M.B. and I.K. analyzed the data. M.J.R., T.K., S.L. and R.K. participated in data analysis, results interpretation and drafting the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Funding
This study was supported by Fulbright Program and Latvian Science Council (Grant #290/2012).
Additional information
Communicated by Thomas S. Hoffmeister.
Rights and permissions
About this article

Cite this article
Krams, I., Burghardt, G.M., Krams, R. et al. A dark cuticle allows higher investment in immunity, longevity and fecundity in a beetle upon a simulated parasite attack. Oecologia 182, 99–109 (2016). https://doi.org/10.1007/s00442-016-3654-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-016-3654-x