
Abstract
This study investigated the influence of temporal and seasonal (i.e., rainy and dry seasons) in communities and infracommunities of metazoan parasites in Pimelodus ornatus, a Siluriformes from the Amazon River in the state of Amapá, in northern Brazil. A total of 144 fish were collected during 2020 and 2021. In 2020, five species of parasites were found (1 Monogenea, 3 Nematoda, and 1 Cestoda), and in 2021, seven species of parasites were found (1 Monogenea, 3 Nematoda, 1 Cestoda, 1 Digenea, and 1 Crustacea). In 2020, Spirocamallanus inopinatus was dominant, while in 2021, Contracaecum sp. was dominant. Brillouin diversity, species richness, and evenness were higher in 2021. There were differences for some parasite infracommunities between years and between seasonal periods. Brillouin diversity and species richness were higher in the rainy season, but there were no differences in evenness and Berger-Parker dominance between the seasonal periods. Temporal and seasonal variations were mainly influenced by rainfall levels, oxygen levels, and environmental temperature and, later, by the availability of infective stages of parasites in the environment and the size of the hosts. Lastly, the influence of the temporal and seasonal variations on the structure of communities and infracommunities of parasites was weaker than expected due to the low infection rates of the majority of parasites found.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Amazon River basin, which is about 8.7 to 11.8 million years old (Gorini et al. 2014; Hoorn et al. 2017; van Soelen et al. 2017), is one of the most complex ecosystems in the world. It has a great biodiversity of fish that feeds riverside populations that live from fishing (Salo et al. 2013). This fish biodiversity is due to the particular characteristics of this large hydrographic basin and its tributaries, which have complex food webs and large vegetation cover, both influenced by the seasonal cycles (rainy/dry) of the Amazon region (Sousa et al. 2017; Arantes et al. 2019). In the Amazon River system, parasites also represent a numerically and functionally important component of this biodiversity. Understanding which factors drive variations in the diversity of parasites in the region has been of interest in fish parasitology. Thus, efforts have been made to understand annual and seasonal effects on diversity, richness, and parasite communities in wild Amazonian fish populations (Neves et al. 2013; Tavares-Dias et al. 2014; Gonçalves et al. 2016; Negreiros et al. 2019a,b; Hoshino and Tavares-Dias 2019, 2020; Cavalcante et al. 2020).
Variation in the structure of parasite communities can be expected over time. Information about these dynamics in Amazonian fish has been little addressed (Negreiros et al. 2019a; Hoshino and Tavares-Dias 2019, 2020). Variations in the diversity and abundance of parasites with complex life cycles can be strongly altered by the population density of intermediate and definitive hosts, which can vary in space, time, and seasonal period, thus playing an important role in the structure of the parasite communities of wild fish populations (Blasco-Costa et al. 2015). These variations can occur mainly in the Amazon basin, where seasonality is characterized by the occurrence of two distinct periods: rainy and dry seasons. The rainy season includes the months from December to May, while the dry season includes the months from June to November (Souza and Cunha 2010). Thus, seasonal variation is an abiotic factor that can influence the life of many Amazonian fish species, including Pimelodus ornatus (Kner, 1857), a Siluriformes fish whose parasite diversity has recently been studied by Lima et al. (2021). Pimelodus ornatus is a catfish found in the Amazon, Madeira, Parnaiba, Negro, upper Parana, Orinoco rivers, as well as large rivers from Guyanas, Paraguay, Bolivia, Peru, Venezuela, and Ecuador (Nomura 1984; Torrente et al. 2013; Lima et al. 2021; Froese and Pauly 2022). This omnivorous fish is active at night and can be captured in main river beds, on rocky bottoms with dead tree trunks, upstream and downstream of rapids and backwaters (Lima et al. 2021; Froese and Pauly 2022). As it is a small size fish, its first sexual maturation occurs from 15.4 cm (Vazzoler 1996).
In Amazonian systems, invertebrates that are intermediate hosts for parasite species with a complex life cycle can also respond to environmental variations in different ways, from the individual level to the component community level (Gonçalves et al. 2016; Negreiros et al. 2019b; Hoshino and Tavares-Dias 2019, 2020). Parasites play an intrinsic role, however, hidden in the Amazonian ecosystems. Thus, understanding many aspects of the ecology of parasites in wild fish from this important Neotropical region has been hampered by the scarcity of annual and seasonal datasets and studies on almost all aspects of environment-host-parasite interactions. As the effects of temporal and seasonal variation on the communities and infra communities of P. ornatus metazoan parasites are not known, we hypothesized that (1) annual and seasonal variations are factors that act directly on the diversity, structure of communities and infracommunities of metazoan parasites in P. ornatus of the Amazon River, and (2) the diversity, communities and infracommunities of metazoan parasites are greater in the rainy season, when there is an increase in the availability and variety of food, and consequently greater contact of fish in the infective stages of the parasites in the environment.
Material and methods
Study area and fish collection
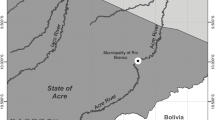
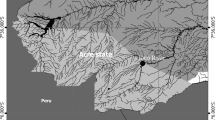
In bimonthly collections, a total of 144 P. ornatus were collected (18.8 ± 2.5 cm and 19.5 ± 8.8 g) from January 2020 to November 2021 in the Amazon River, near the Island of Santana, in the municipality of Santana, Amapá, Brazil (Fig. 1). Fish were collected with gill nets with different sizes and meshes (15 mm, 20 mm, 25 mm, 30 mm, and 35 mm between knots), cast nets (mesh of 20 mm between knots) and hand lines. Then, the fish were euthanized by the spinal cord transection method, preserved in 10% formaldehyde, and transported to the Aquaculture and Fisheries Laboratory of Embrapa Amapá, Macapá, state of Amapá (Brazil), for analysis of the parasites.

Collection area of Pimelodus ornatus in the Amazon River, the state of Amapá, northern Brazil
At the fish sampling sites, water quality parameters such as electrical conductivity, pH, and total dissolved solids were measured using a multiparameter (Akso, model Combo5-02–1016). Dissolved oxygen levels and water temperature were measured using an oximeter (Instrutherm, Model MO-900). Rainfall data from the fish collection region were obtained from the Hydrometeorology and Renewable Energy Center (NHMET) of The Institute of Scientific and Technological Research of the State of Amapá (IEPA) (Table 1).
Parasite analysis procedures
In the laboratory, fish were weighed (g) and measured in total length (cm) and were necropsied to collect the metazoan parasites. The mouth, opercular cavity, gills, and fins were analyzed for ectoparasites, and internal organs such as gastrointestinal tract and viscera were examined for endoparasites with the aid of a stereomicroscope. The parasites found were preserved in 70% ethyl alcohol for the preparation of permanent slides following previous recommendations (Eiras et al. 2006).
Data analysis
Parasite indices such as prevalence and mean abundance of infracommunities were calculated (Bush et al. 1997). To describe the parasite community, species richness, Brillouin diversity index (HB), evenness (E), and Berger-Parker dominance index (d) data were calculated using PAST software (Hammer et al. 2001). The Spearman correlation coefficient (rs) was used to evaluate possible correlations of body length and weight of host with parasite abundance (Zar 2010).
To determine whether total length (cm), weight (g), prevalence, abundance, species richness, evenness, Brillouin diversity index, and Berger-Parker dominance index data followed a normal distribution pattern, Shapiro–Wilk and Bartlett tests were used, respectively. To verify if there were differences in the prevalence of parasites between the years 2020 and 2021 and the seasonal periods (rainy and dry), the chi-square test (χ2) with Yates’s correction was used, and to verify differences in the abundance of parasites, the Mann–Whitney test (U) was used (Zar 2010). To estimate differences in diversity indices (species richness, evenness, Brillouin diversity index, and Berger-Parker dominance index) of metazoan parasites between 2020 and 2021 and seasonal periods, the Kruskal–Wallis followed by Dunn test was employed. These analyses were performed in R software (R CoreTeam 2021).
Variance analysis of permutation (PERMANOVA) was used to detect differences in the parasite community between the years 2020 and 2021 and the rainy and dry seasons. Temporal and seasonal variables were fitted in the ordering of principal coordinate analysis (PCoA) based on Bray–Curtis distance, using the envfit function of the vegan package (Oksanen et al. 2020), and p-values were calculated by the permutation test (number of permutations = 999) in R software. To analyze how differences in sampling effort can influence the results, we plotted a species accumulation curve (observed and expected) for the years 2020 and 2021 and for the rainy and dry seasons. These analyses were performed using R software (R CoreTeam 2021).
Results
Component community and parasite infracommunities in 2020 and 2021
Fish collected in 2020 measured 15.5 ± 2.4 cm, while those collected in 2021 measured 16.0 ± 2.5 cm, showing no significant differences (t = − 1.3, p = 0.18) between years. Fish collected in 2020 weighed 18.9 ± 8.1 g, and those collected in 2021 weighed 20.0 ± 9.4 g, showing no significant differences (t = − 7.5, p = 0.4) between the years.
Host length did not present correlation with the abundance of S. inopinatus (rs = 0.16, p = 0.052), Contracaecum sp. (rs = 0.04, p = 0.57), and Proteocephalidae gen. sp. (rs = 0.01, p = 0.83), nor did the weight with the abundance of S. inopinatus (rs = 0.06, p = 0.43), Contracaecum sp. (rs = − 0.01, p = 0.84), and Proteocephalidae gen. sp. (rs = 0.03, p = 0.70).
Of the fish examined, 412 parasites were collected, those being 146 parasites in 2020 and 266 parasites in 2021. In 2020, 70.4% of fish were parasitized by Demidospermus Suriano, 1983; Cucullanus pinnai (Travassos, Artigas & Pereira, 1928); Spirocamallanus inopinatus (Travassos, Artigas & Pereira, 1928); larvae of Contracaecum Railliet & Henry, 1912; and plerocercoids of Proteocephalidae La Rue, 1911. In 2021, 86.3% of fish were parasitized by Demidospermus sp.; C. pinnai; S. inopinatus; larvae of Contracaecum sp.; Proteocephalidae gen. sp., Genarchella genarchella (Travassos, Artigas & Pereira, 1928); and Thelotha henselli (Von Martens, 1869).
There was a dominance of S. inopinatus in 2020 and Contracaecum sp. in 2021. Prevalence and abundance of Contracaecum sp. was higher in 2021, while the other parasite species did not differ between the years (Table 2).
Axes 1 and 2 of PCoA of were responsible for 63% of the total data variation in the composition of parasite abundance in 2020 and 2021. Despite the figure showing an overlap and sharing of species between the years, there was a significant difference (PERMANOVA: F = 0.060, p = 0.001) in parasite infracommunity, influenced mainly by the variation in the abundance of S. inopinatus (R2 = 0.356, p = 0.001), Contracaecum sp. (R2 = 0.719, p = 0.001), and Proteocephalidae gen. sp. (R2 = 0.339, p = 0.001) (Fig. 2).

Principal coordinate analysis (PCoA) using a Bray–Curtis distance matrix for communities of metazoan parasites of Pimelodus ornatus from Amazon River, in the state of Amapá (Brazil), collected in 2020 and 2021. The percentage of variation explained by the plotted principal coordinates is indicated on the axes
The Brillouin diversity index, species richness of parasites, and evenness were higher in 2021, while Berger-Parker dominance showed no differences between years (Fig. 3). The parasite species accumulation curve showed that the species richness was different between years. It also showed that the parasite richness was not affected by the sampling effort between years (Fig. 4).

Diversity parameters of metazoan parasites in Pimelodus ornatus from Amazon River, in the state of Amapá (Brazil), collected in 2020 and 2021 (Box plots represent medians, interquartile ranges, minimum–maximum ranges, and outliers). Different letters indicate differences between the medians according to Dunn’s test (p < 0.001)

Species accumulation curve of metazoan parasites in Pimelodus ornatus from Amazon River, in the state of Amapá (Brazil), collected in 2020 and 2021
Component community and infracommunity of parasites in the rainy and dry seasons
The fish collected in the rainy season measured 15.0 ± 2.6 cm, while those collected in the dry period measured 16.9 ± 1.8 cm, showing significant differences (t = − 4.3, p = 0.0001) between the seasonal periods. The fish collected in the rainy season weighed 16.8 ± 8.5 g, and those collected in the dry season weighed 16.9 ± 1.8 g, showing significant differences (t = − 4.6, p = 0.0001) between the seasonal periods.
Of the hosts examined in the rainy season, 74.1% were parasitized by Demidospermus sp., C. pinnai, S. inopinatus, Contracaecum sp., Proteocephalidae gen. sp., G. genarchella, and T. henselli. Of the fish examined in the dry period, 84.7% were parasitized by Demidospermus sp., C. pinnai, S. inopinatus, Contracaecum sp., and Proteocephalidae gen. sp. Genarchella genarchella and T. henselli were found only in the rainy season. In the rainy and dry seasons, S. inopinatus was the parasite with the highest prevalence and abundance. The prevalence of Demidospermus sp. and C. pinnai was higher in the dry season, while the other species did not differ between the seasonal periods (Table 3).
Axes 1 and 2 of PCoA were responsible for 63% of the total variation in the composition of parasite abundance in the rainy and dry seasons (Fig. 5). This figure shows overlap and sharing of species, showing that there were no significant differences (PERMANOVA: F = 0.009, p = 0.34) in the parasite infracommunities between the seasonal periods. However, there was a significant variation in the abundance of S. inopinatus (R2 = 0.356, p = 0.001), Contracaecum sp. (R2 = 0.405, p = 0.001), and Proteocephalidae gen. sp. (R2 = 0.339, p = 0.001).

Principal coordinate analysis (PCoA) using a Bray–Curtis distance matrix for communities of metazoan parasites of Pimelodus ornatus from the Amazon River, in the state of Amapá (Brazil), during rainy and dry seasons. The percentage of variation explained by the plotted principal coordinates is indicated on the axes
The Brillouin diversity index and parasite species richness were higher in the dry season, while evenness and Berger-Parker dominance did not show significant differences between the seasons (Fig. 6). The species accumulation curve showed that the species richness of parasites diverged between the rainy and dry seasons, and that the richness of parasites was not affected by the sampling effort. The accumulation curve of parasite species in the rainy and dry seasons reached an asymptote (Fig. 7).

Diversity parameters of metazoan parasites in Pimelodus ornatus from Amazon River, in the state of Amapá (Brazil), during rainy and dry seasons (Box plots represent medians, interquartile ranges, minimum–maximum ranges, and outliers). Different letters indicate differences between the medians according to Dunn’s test (p < 0.001)

Species accumulation curve of metazoan parasites in Pimelodus ornatus from Amazon River, in the state of Amapá (Brazil), during rainy and dry seasons
Discussion
Temporal variation of metazoan parasites
Surprisingly, no relationship between P. ornatus size and abundance of parasites between the years was observed in the present study; however, we also examined within the pooled parasite species. This indicated that despite host size being a significant determining factor in the diversity, species richness, and abundance of parasites (Poulin and Leung 2011; Marcogliese et al. 2016; Baia et al. 2018; Lehun et al. 2022), it did not influence the temporal differences observed here, because other factors are related. Furthermore, many parasite communities experience temporal variations due to seasonal changes in biotic and abiotic environmental factors such as parasite life cycle, environmental dynamics, host-specific immune responses, and reproduction period (Cavalcante et al. 2020; Lehun et al. 2022).
As the Amazon River basin is one complex ecosystem, the temporal variations in the fish communities that inhabit this region remain poorly understood. The community and infracommunities of metazoan parasites in P. ornatus from the Amazon River did not show a similar pattern between the years studied due to seasonal variations. In Pimelodus blochi (Valenciennes, 1840) from the Iaco River, in western Amazon (Brazil), the long-term temporal variations in the community and infracommunity have been attributed to factors linked to the body size of the host and variations in the availability of infective stages in the environment. In Hemibrycon surinamensis (Gery, 1962) (Hoshino and Tavares-Dias 2019) and Metynnis lippincottianus (Hoshino and Tavares-Dias 2020) both host fishes from the eastern Amazon region (Brazil), short-period, temporal variation was associated with seasonal cycle (rainy/dry); availability of infective stages; changes in the recruitment process of parasite species; urban eutrophication in the environment; and host body size.
The results showed that there was an alternation in the dominance of parasite species in P. ornatus, with the dominance of S. inopinatus in 2020 and larvae of Contracaecum sp. in 2021. These nematodes in adult and larval stages, respectively, are related to possible changes in the diet of P. ornatus or volume of food consumed or changes in availability and efficiency of infection by intermediate hosts in the environment (Sabas and Brasil-Sato 2014; Blasco-Costa et al. 2015; Tavares-Dias et al. 2017), since this host fish has an omnivorous feeding habit (Lima et al. 2021; Froese and Pauly 2022) and acts as an intermediate (Contracaecum sp.) and definitive host (S. inopinatus). In P. blochii, from the Iaco River in the western Amazon (Brazil), a long-term study has shown a dominance of larvae and adults of 11 species of nematodes (Negreiros et al. 2019a). However, in H. surinamensis, from the eastern Amazon, the dominance of metazoan parasites was of the monogenean species during the two years of study (Hoshino and Tavares-Dias 2019). In M. lippincottianus, from eastern Amazon (Brazil), the dominance of metazoan parasites was of monogenean species in a year and digenean Dadayus pacupeva (Lacerda, Takemoto & Pavanelli, 2003) in the other year (Hoshino and Tavares-Dias 2020). Therefore, these results show a different pattern among these species of Siluriformes and Characiformes.
The PCoA indicated differences between 2020 and 2021, mainly due to differences in the abundance of S. inopinatus, Contracaecum sp., and Proteoceplalidae gen. sp. These differences may be related to variations in rainfall levels between the years studied, thus influencing the availability of intermediate hosts (e.g., crustaceans) and variations in environmental characteristics, favoring the recruitment of species with a complex life cycle, in addition to the position of P. ornatus in the food web, as has also been reported for other wild fish species (Villalba-Vasquez et al. 2018; Hoshino and Tavares-Dias 2019, 2020; Lehun et al. 2022).
Wild fish populations act as hosts for various taxa of parasites with varied life cycle strategies. Hence, the diversity and richness of parasites depends on the host species and other factors related to the host and environment such as parasite life cycle, feeding habits, and the reproductive period of host fish (Tavares-Dias et al. 2014, 2017; Baia et al. 2018; Cavalcante et al. 2020). For P. ornatus, Brillouin diversity, parasite species richness, and evenness were higher in 2021, caused by the presence of G. genarchella and T. henselii, which occurred only in 2021. Similar studies have also reported annual differences in the diversity and species richness of parasites in P. blochii from the Iaco River (Negreiros et al. 2019a) and in H. surinamensis from the Igarapé Fortaleza basin (Hoshino and Tavares-Dias 2019). These differences have been influenced by the precipitation levels between the years, thus altering the flow patterns of the rivers and the water quality of these Amazonian hydrographic basins.
Seasonal variation of metazoan parasites
The specimens of P. ornatus collected in the dry season were larger and heavier than in the rainy season, possibly indicating a greater feeding in this seasonal period. The Amazon River systems present periods of flooding and droughts, dynamics that also influence the communities of invertebrates and fish that serve as food for many omnivorous fish such as P. ornatus. As the diversity of zooplankton and other invertebrates is higher during the rainy season, this indicates that Amazonian fish have better feeding conditions and food diversity in this seasonal period (Tavares-Dias et al. 2014; Gonçalves et al. 2016), preparing for reproduction. As in wild fish populations, host size is an important factor influencing parasite communities and infracommunities and may be associated with their trophic level, diet, and vulnerability (Poulin and Leung 2011) – This factor usually accounts, in part, for the variation in infracommunities and diversity of parasites (Neves et al. 2013; Tavares-Dias et al. 2014; Baia et al. 2018).
The Amazonian region has a well-defined seasonality, with periods of intense rainfall (rainy season) and less rainy periods (dry season). Hence, in the Amazon River system, variations in communities and infracommunities of parasites in wild fish populations have been related to changes generated by the rainy/dry cycle, affecting the recruitment of parasite species, increased availability of food for the host fishes, and availability of infective stages in the environment (Neves et al. 2013; Gonçalves et al. 2018; Negreiros et al. 2019a; Hoshino and Tavares-Dias 2019, 2020), as well as by the season of reproduction of fishes (Cavalcante et al. 2020). Therefore, levels of rainfall can cause effects on aquatic ecosystems such as changes in fish habitat, water quality, and velocity, consequently increasing the stress on host fishes and susceptibility to parasite infections, affecting thus the structure of parasite communities and infracommunities and of intermediate hosts (Gonçalves et al. 2016).
The PCoA showed no seasonal difference in the infracommunity of S. inopinatus, Contraceacum sp., and Proteocephalidae gen. sp. However, parasite species richness and Brillouin diversity were higher in the dry season, while evenness and Berger-Parker dominance had no seasonal influence, because the dominant species did not vary between seasonal periods. In contrast, in P. blochii from the rivers Acre and Xapuri, these diversity parameters did not have a seasonal influence (Cavalcante et al. 2020). Environmental factors such as the decrease in dissolved oxygen levels and increase in water temperature during the dry period increased the species richness and parasite diversity in P. ornatus and, therefore, the recruitment processes of parasite species found here.
Nematode S. inopinatus was dominant in both seasonal periods studied. The prevalence of Demidospermus sp. and C. pinnai were higher in the dry season, when the water quality levels were lower, favoring the reproduction of monogeneans and nematodes, in contrast to what was detected in the rainy season, when there was a high oxygen level and an average flow of water of 1.0 × 105m3/s, factors that can reduce the accumulation of pollutants and eutrophication of the studied area (Abreu et al. 2020). Similar results have been reported for P. blochii from Acre River, in western Amazon, infected by monogeneans Demidospermus spp. and Ameloblastella sp. in the dry season (Negreiros et al. 2019a). Metynnis lippincottianus from the Igarapé Fortaleza basin, in eastern Amazon, also presented a higher prevalence of Contracaecum sp. larvae in the dry season (Hoshino and Tavares-Dias 2020). In contrast, H. surinamensis from the Igarapé Fortaleza basin showed no seasonal variation in the levels of infection by Contracaecum sp. (Hoshino and Tavares-Dias 2019), therefore indicating a differentiated pattern between host fish species.
Digenea is part of the parasite community of wild Amazonian fish populations (Neves et al. 2013; Negreiros et al. 2019a,b; Hoshino and Tavares-Dias 2019, 2020). The transmission of these endoparasites occurs when metacercariae are ingested by host fish such as P. ornatus, which has an omnivorous diet, which facilitates the infection by digeneans. Infection by G. genarchella and T. henselii occurred only during the rainy season when the increase in the water level of the Amazon River facilitated the chances of contact of these parasites with P. ornatus. However, in H. surinamensis (Hoshino and Tavares-Dias 2019) and M. lippincottianus (Hoshino and Tavares-Dias 2020), both hosts of tributaries of the Amazon River system, infection by G. genarchella was not influenced by the seasonality, and Ergasilus xinguensis (Taborba, Paschoal & Luque, 2016) also occurred only in the rainy season.
In conclusion, about 43% of parasites were larvae, indicating that P. ornatus is the host intermediate. These results, based on a two-year sampling timescale, indicated that behaviors differed between years and between seasons regarding diversity and some infracommunities of parasites. The effects of temporal and seasonal variations in the diversity and levels of infection were related to variations in rainfall levels and, consequently, the availability of infective stages of parasites with a direct and indirect life cycle and changes in the recruitment of parasite species in the environment caused by seasonality. Furthermore, this first study on the temporal and seasonal variations recorded the first occurrence of G. genachella and T. henselii for P. ornatus. Lastly, the studied region from the Amazon River has average water flow which, to a certain degree, prevents urban eutrophication; however, it has environmental factors that influence the infection rates of some metazoan parasites communities and infracommunities, as well as diversity and species richness over the years and seasonal periods, thus corroborating the hypotheses suggested.
Data availability
Not applicable for that section.
Code availability
Not applicable for that section.
References
Abreu CHM, de Barros LCM, Brito DC et al (2020) Hydrodynamic modeling and simulation of water residence time in the estuary of the Lower Amazon River. Water 12:660. https://doi.org/10.3390/w12030660
Arantes CC, Winemiller KO, Petrere M, Freitas CE (2019) Spatial variation in aquatic food webs in the Amazon River floodplain. Freshw Sci 38:213–228. https://doi.org/10.1086/701841
Baia RRJ, Florentino AC, Silva LMA, Tavares-Dias M (2018) Patterns of the parasite communities in a fish assemblage of a river in the Brazilian Amazon region. Acta Parasitol 63:304–316. https://doi.org/10.1515/ap-2018-0035
Blasco-Costa I, Rouco C, Poulin R (2015) Biogeography of parasitism in freshwater fish: spatial patterns in hot spots of infection. Ecography 38:301–310. https://doi.org/10.1111/ecog.01020
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on its own terms: Margolis et al. revisited. J Parasitol 575–583. https://doi.org/10.2307/3284227
Cavalcante PHO, Silva MT, Pereira ANS et al (2020) Helminth diversity in Pimelodus blochii Valenciennes, 1840 (Osteichthyes: Pimelodidae) in two Amazon Rivers. Parasitol Res 119:4005–4015. https://doi.org/10.1007/s00436-020-06906-x
Eiras JC, Takemoto RM, Pavanelli GC (2006) Métodos de estudo e técnicas laboratoriais em parasitologia de peixes, 2nd edn. Editora da Universidade Estadual de Maringá, Maringá
Froese R, Pauly D (2022) FishBase. World wide web electronic publication. www.fishbase.org, version (02/2022)
Gonçalves RA, Oliveira MSB, Neves LR, Tavares-Dias M (2016) Seasonal pattern in parasite infracommunities of Hoplerythrinus unitaeniatus and Hoplias malabaricus (Actinopterygii: Erythrinidae) from the Brazilian Amazon. Acta Parasitol 61:119–129. https://doi.org/10.1515/ap-2016-0016
Gonçalves BB, Oliveira MSB, Borges WF et al (2018) Diversity of metazoan parasites in Colossoma macropomum (Serrasalmidae) from the lower Jari River, a tributary of the Amazonas River in Brazil. Acta Amaz 48:211–216. https://doi.org/10.1590/1809-4392201704371
Gorini C, Haq BU, Reis AT, et al (2014) Late Neogene sequence stratigraphic evolution of the Foz do Amazonas basin Brazil. Terra Nov 26:179–185. https://doi.org/10.1111/ter.12083
Hammer O, Harper DAT, Ryan PD (2001) PAST: paleontological statistics software package for education and data analysis. Palaeont Elect 4:9
Hoorn C, Bogotá-A GR, Romero-Baez M et al (2017) The Amazon at sea: onset and stages of the Amazon River from a marine record, with special reference to Neogene plant turnover in the drainage basin. Global Planet Change 153:51–65. https://doi.org/10.1016/j.gloplacha.2017.02.005
Hoshino EM, Tavares-Dias M (2019) Interannual and seasonal variation in protozoan and metazoan parasite communities of Hemibrycon surinamensis, a Characid fish inhabiting the Brazilian Amazon. Acta Parasitol 64:479–488. https://doi.org/10.2478/s11686-019-00057-5
Hoshino EM, Tavares-Dias M (2020) Temporal and seasonal variations in parasites of Metynnis lippincottianus (Characiformes: Characidae), a host from the eastern Amazon (Brazil). J Nat Hist 53:2723–2736. https://doi.org/10.1080/00222933.2020.1733121
Lehun AL, Cavalcanti LD, Lizama M de los AP, et al (2022) Temporal effects and changes in the parasitic community of Prochilodus lineatus (Valenciennes, 1837) (Characiformes: Prochilodontidae) in a floodplain. J Helminthol 96https://doi.org/10.1017/S0022149X21000742
Lima ES, Oliveira MSB, Tavares-Dias M (2021) Diversity and community ecology of metazoan parasites in Pimelodus ornatus (Siluriformes: Pimelodidae) from the Amazonas River in Brazil. Rev Bras Parasitol Vet 30(3):e006021. https://doi.org/10.1590/S1984-29612021065
Marcogliese DJ, Locke SA, Gélinas M, Gendron AD (2016) Variation in parasite communities in spottail shiners (Notropis hudsonius) linked with precipitation. Int J Parasitol 102:27–36. https://doi.org/10.1645/12-31
Negreiros LP, Florentino AC, Pereira FB, Tavares-Dias M (2019a) Long-term temporal variation in the parasite community structure of metazoans of Pimelodus blochii (Pimelodidae), a catfish from the Brazilian Amazon. Parasitol Res 118:3337–3347. https://doi.org/10.1007/s00436-019-06480-x
Negreiros LP, Pereira FB, Tavares-Dias M (2019b) Metazoan parasites of Calophysus macropterus (Siluriformes: Pimelodidae) in the Acre and Iaco rivers in the western Amazon region of Brazil: diversity, similarity and seasonal variation. J Nat Hist 53:1465–1479. https://doi.org/10.1080/00222933.2019.1657195
Neves LR, Pereira FB, Tavares-Dias M, Luque JL (2013) Seasonal influence on the parasite fauna of a wild population of Astronotus ocellatus (Perciformes: Cichlidae) from the Brazilian Amazon. Int J Parasitol 99:718–721. https://doi.org/10.1645/12-84.1
Nomura H (1984) Dicionário dos peixes do Brasil. Technical Books Livraria, Rio de Janeiro
Oksanen J, Blanchet FG, Kindt R, et al (2020) Vegan: community ecology package. R package. https://CRAN.R-project.org/package=vegan
Poulin R, Leung T (2011) Body size, trophic level, and the use of fish as transmission routes by parasites. Oecologia 166:731–738. https://doi.org/10.1007/s00442-011-1906-3
R CoreTeam (2021) R: a language and environment for statistical computing. Vienna, Austria. Available at https://www.R-project.org/
Sabas CSS, Brasil-Sato MC (2014) Helminth fauna parasitizing Pimelodus pohli (Actinopterygii: Pimelodidae) from the upper São Francisco River, Brazil. Rev Bras Parasitol Vet 23:375–382. https://doi.org/10.1590/S1984-29612014067
Salo M, Sirén A, Kalliola R (2013) Diagnosing wild species harvest: resource use and conservation. Academic Press
Sousa R, Souza LA, Frutuoso ME, Freitas CEC (2017) Seasonal dynamic of amazonian small-scale fisheries is dictated by the hydrologic pulse. Bol Inst Pesca 3:207–221. https://doi.org/10.20950/1678-2305.2017v43n2p207
Souza EB, Cunha AC (2010) Climatologia de precipitação no Amapá e mecanismos climáticos de grande escala. In: Ecodesign T (ed) Tempo, Clima e Recursos Hídricos. 177–195
Tavares-Dias M, Oliveira MSB, Gonçalves RA, Silva LMA (2014) Ecology and seasonal variation of parasites in wild Aequidens tetramerus, a Cichlidae from the Amazon. Acta Parasitol 59:158–164. https://doi.org/10.2478/s11686-014-0225-3
Tavares-Dias M, Oliveira MSB, Gonçalves RA, Neves LR (2017) Parasitic diversity of a wild Satanoperca jurupari population, an ornamental cichlid in the Brazilian Amazon. Acta Amaz 47:155–162. https://doi.org/10.1590/1809-4392201602514
Torrente LJ de QG, Ohara VWM, Jansen TH da SP, da Costa Doria ZCR (2013) Peixes do Rio madeira. São Paulo INPA
van Soelen EE, Kim J-H, Santos RV et al (2017) A 30 Ma history of the Amazon River inferred from terrigenous sediments and organic matter on the Ceará Rise. Earth Planet Sci Lett 474:40–48. https://doi.org/10.1016/j.epsl.2017.06.025
Vazzoler A (1996) Biologia da reprodução de peixes teleósteos: teoria e prática. Maringá: Eduem 169:
VillalbaVasquez PJ, Violante-González J, Monks S et al (2018) Temporal and spatial variations in the metazoan parasite communities of the Panama spadefish, Parapsettus panamensis (Pisces: Ephippidae), from the Pacific coast of Mexico. Invertebr Biol 137:339–354. https://doi.org/10.1111/ivb.12232
Zar J (2010) Biostatistical analysis 5th ed Upper Saddle River, NJ: Prentice-Hall 557–558
Funding
This study received financial support from the National Council for Scientific and Technological Development (Conselho Nacional de Desenvolvimento Científico e Tecnológico, CNPq, Brazil) through a productivity grant to Tavares-Dias, M (#303013/2015–0).
Author information
Authors and Affiliations
Contributions
All authors contributed to the conception and design of the study. Material preparation, data collection, and analysis were performed by Elvis Silva Lima. The first version of the manuscript was written by Elvis Silva Lima, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Ethics approval
Ethics approval to obtain access to genetic heritage was authorized by the Brazilian Ministry of the Environment (SISBio n° 73550–1 and SisGen no AA4B6BA). This study was developed in accordance with the principles adopted by the Brazilian College of Animal Experimentation (COBEA) and was conducted under authorization from the Ethics Committee for Animal Use of Embrapa (protocol no 014/2018).
Consent to participate
Not applicable for that section.
Consent for publication
The authors declare that they are aware of the publication of this study.
Conflict of interest
The authors declare no competing interests.
Additional information
Section Editor: Shokoofeh Shamsi.
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Lima, E.S., Oliveira, M.S.B. & Tavares-Dias, M. Temporal and seasonal variation of metazoan parasites in Pimelodus ornatus (Siluriformes: Pimelodidae) from the Amazon River, Brazil. Parasitol Res 121, 3171–3181 (2022). https://doi.org/10.1007/s00436-022-07637-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-022-07637-x