
Abtract
Structure of the helminth community of Megaleporinus obtusidens collected in Lake Guaíba was evaluated, and the results indicated that the diversity of helminth species was probably determined by fish behavior and eating habits. The influence of sex, weight, and standard length of hosts for parasitic indices was also analyzed. Sixteen helminth species were found parasitizing M. obtusidens, including the following: platyhelminths, with the highest richness, represented by one species of Aspidobothrea; four species of Digenea; and eight species of Monogenea; the latter, presented the highest prevalence. Rhinoxenus arietinus, found in nasal cavities, had the greater abundance, and was the only species classified as core. The prevalence of Urocleidoides paradoxus was significantly influenced by the sex of the host; females had the highest values. Abundance was weakly influenced by fish weight and the body length of the hosts. Urocleidoides sp. had its abundance weakly influenced by the host weight. The other helminths were not influenced by biometric characteristics of the hosts. The total species richness was similar between male and female fish, and both had 14 helminth species of parasites.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Leporinus obtusidens, known as “piava,” was recently included in the genus Megaleporinus proposed by Ramirez et al. (2017) after phylogenetic study of Anastomidae. Megaleporinus obtusidens Ramirez, Birindelli and Galetti, 2017 is the only species of Megaleporinus occurring in the State of Rio Grande do Sul, southern Brazil (Reis et al. 2003). It is found from north to south in Brazil, as well as in Argentina, Uruguay, and Paraguay (Britski et al. 2012). Three genetically distinct populations can be separated: (a) basin of the Paraná, Uruguay, and Jacui rivers; (b) São Francisco River; and (c) Paraguay River (Ramirez et al. 2016, 2017). Zaniboni-Filho and Schultz (2003) observed a reduction in the M. obtusidens population and related it to agriculture and industrial practices. Thus, species survey studies in this area become crucial in evaluating threats to diversity.
Megaleporinus obtusidens was intensely studied in the 1980s, and aspects of its morphology (Sidlauskas and Vari 2008; Britski et al. 2012), biology (Mello et al. 1999; Hartz et al. 2000; Penchaszadeh et al. 2000; Santos 2000; Piana et al. 2003; Moraes et al. 2009), and ecology (Oldani et al. 1992; Araya 1999; Araya et al. 2005) have been elucidated.
Studies dealing with the helminth fauna of this species, however, are scarce and are concentrated mainly in the States of Paraná and Minas Gerais (Feltran et al. 2004; Takemoto et al. 2009; Kohn et al. 2011). Knowledge about the parasitic community can contribute to better understanding host habits and behaviors and whether they may influence the population dynamics of its host. It is known that some factors can influence the community composition of parasites, such as dietary habits, sex, and host size. Barber and Poulin (2002) stated that males and females have physiological, morphological, and behavioral differences, and these factors may influence parasitism. Studies have shown that the opposite also occurs, that is, parasites can mediate sexual selection (Barber and Poulin 2002). The influence of sex, standard length, and host weight on the helminth community has been poorly studied in Leporinus species and especially in the genus Megaleporinus.
Scarce information addressing the parasitic fauna of M. obtusidens in the State of Rio Grande do Sul, as well as the few studies about community component, motivated the present study, which is composed of description and analysis of the helminth community structure.
Material and methods
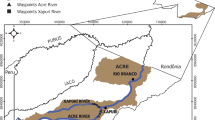
Sixty specimens of M. obtusidens (20 males and 40 females) were captured by professional fishermen from Lake Guaíba (30°01′S, 51°16′W), State of Rio Grande do Sul, southern Brazil, from August to October 2013 (Fig. 1). The fish were bought directly from the fishermen as they were being taken to the local market immediately after the catch. The study was approved by the Universidade Federal do Rio Grande do Sul (UFRGS) Ethics Committee (Process Number 27531).

Collection site; (a) South America, Brazil in light gray; (b) Rio Grande do Sul; (c) Delta do Jacuí, highlighted the Ilha da Pintada
The fish were weighed, measured (total and standard length), and necropsied following Amato et al. (1991). The helminths were collected and placed in 0.65% saline solution, fixed, and further stained and/or clarified according to Amato and Amato (2010). Ecological descriptors used—prevalence, mean intensity of infestation or infection, abundance, range, and site of infection—followed Bush et al. (1997). Species with prevalence greater than or equal to 10% were used in the analysis of helminth component community following Bush et al. (1990) recommendations.
Mann–Whitney’s “U” test was calculated to evaluate if the sex of the host had influence on the parasite abundance and intensity. Fisher’s exact test was used to evaluate if the sex of the host had influence on the parasite prevalence. To analyze if the weight and length of the hosts had influenced abundance and intensity of parasites, the Spearman’s correlation test “rs” was used, and to analyze the influence of this parameter over the prevalence of the parasite species, the Pearson’s correlation test “r” was used in pre-determined classes of fish size and weight (Zar 1996). The weight and length classes were determined according to the Sturges’ rule, using mean values between standard deviations. Hosts were classified in classes according to the ranges presented.
Generalized linear models (GLM) were used for the analysis. First, a model with first-order interactions was fitted: G(abund) = sex + Weight + parasite species + interation; where G is a link function and abund represents abundance of species. Abundance was chosen as a dependent variable because it was the index that showed the greatest variability among parasite species; so, we want to explain which other variables influence this first. Weight was considered as covariate and sex and parasite species as factor. General linear models (McCullagh 1984) followed by analysis of variance (ANOVA; F test) was performed to verify which factors influence the abundance of the parasite species (Gaussian model). The significance of the model was tested from the null model test, “rdiagnostic” and observing the values Residual deviance/degree of freedom. Further information on test values and significance is found in the supplemental material.
The co-occurrence of parasite species was evaluated by the Spearman correlation “rs” (Zar 1996) and was plotted on a graph using the Corrplot (package in R program). All statistical tests followed the significance level of 0.05 and were calculated with R® program, Version 3.4.1. The relationship between species was also tested using the index of affinity proposed by Fager (1957). The index of affinity between pair of parasite species occurring in the same breeding site was calculated, and we determined if the affinity index obtained was statistically significant using the t test (with p ≤ 0.05). Rhinoxenus arietinus was not included in this analysis because it did not share an occurrence site with any other species. The results of the Fager index and the t test are found in Table I of the Supplementary Material.
The helminth species were classified according to their frequency in the component community as follows: core (prevalence ≥ 66.6%), secondary (prevalence between 33.3% and 66.6%), or satellite (prevalence ≤ 33.3%), following Bush and Holmes (1986). The diversity indices followed Magurran (2004) and Nehring and Von Zuben (2010) to describe the component community with respect to its heterogeneity. The species richness (S), Brillouin Diversity (HD), Equitability Shannon-Winner (J), modified Hill Equitability (E), and Dominance Simpson (D) were estimated with the aid of DIVES version 3.0 program (Rodrigues 2015). Helminth representative specimens were deposited in the “Coleção Helmintológica do Laboratório de Helmintologia” (CHDZ) of the Departamento de Zoologia, Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, RS, Brazil. Voucher specimens of Tereancistrum paranaensis Karling, Lopes, Takemoto and Pavanelli, 2014 (CHIOC 38085; CHIOC 38086) and Tereancistrum parvus Kritsky, Thatcher and Kayton, 1980 (CHIOC 38087; CHIOC 38088) were deposited at the “Coleção Helmintológica do Instituto Oswaldo Cruz” (CHIOC), Rio de Janeiro, RJ, Brazil.
Results
Component community structure
Fifty nine of 60 (98.33%) M. obtusidens were parasitized by at least one species of helminth. A total of 2376 helminths were collected; of these, 1720 were classified as endoparasites and the remaining 656 as ectoparasites. The mean abundance and mean intensity of infection were 39.6 and 40.27 specimens/host, respectively. The ecological indices, prevalence, mean intensity, mean abundance, range of infection/infestation, and site infection/infestation are presented in Table 1. Sixteen species of helminths were found, 13 belonging to the phylum Platyhelminthes; one species of Aspidobothrea (Aspidogastridae), four species of Digenea, and eight species of Monogenea. The digeneans were represented by Saccocoelioides godoyi Kohn and Fróes, 1986 (31.66%), Creptotrema lynchi Brooks, 1976 (21.66%), Saccocoelioides nanii Szidat, 1954 (15%), and Genarchella genarchella Travassos, Artigas and Pereira, 1928 (3.3%). The most representative group was Monogenea, with eight species, among these, Kritsky eirasi Guidelli, Takemoto, and Pavanelli, 2003, the only endoparasite of the group, was found in the ureters of 58.33% of the analyzed hosts. Other monogeneans were ectoparasites, Rhinoxenus arietinus Kritsky, Boeger, and Thatcher, 1988, Urocleidoides paradoxus Kritsky, Thatcher, and Boeger, 1986, Urocleidoides sp., Tereancistrum parvus Kritsky, Thatcher, and Kayton, 1980, Tereancistrum paranaensis Karling, Lopes, Takemoto, and Pavanelli, 2014, Jainus piava Karling, Bellay, Takemoto, and Pavanelli, 2011, and an undetermined species of Dactylogyridae were found. The other species are represented by Acanthocephala, with one species, Echinorhynchus sp., while Nematoda was represented by two species, Cucullanus sp., (larvae and adults) and Contracaecum sp. (larvae), where all these species presenting low ecological indices.
Of all species found in this study, seven were registered for the first time in M. obtusidens (Aspidogastridae gen. sp., S. nanii, J. piava, T. paranaensis, K. eirasi, Urocleidoides sp., and Echinorhynchus sp.). Additionally, eight other species, G. genarchella, C. lynchi, Dactylogyridae gen. sp., R. arietinus, T. parvus, U. paradoxus, Contracaecum sp., and Cucullanus sp., were recorded for the first time in Lake Guaíba.
Regarding the contribution of these species to the composition of the parasite community, it was observed that of the 16 species found, nine were endoparasites (Aspidogastridae gen. sp., G. genarchella, S. godoyi, S. nanii, C. lynchi, K. eirasi, Echinorhynchus sp., Contracaecum sp., and Cucullanus sp.) and seven were ectoparasites (Dactylogyridae gen. sp., J. piava, R. arietinus, T. paranaensis, T. parvus, U. paradoxus, and Urocleidoides sp.). Digenean trematodes with 1472 specimens represented the clear majority of the helminths found (61.95% of specimens), followed by monogeneans with 861 specimens (36.23%), nematodes with 24 specimens (01.01%), Acanthocephala with 18 specimens (0.75%), and Aspidogastrid represented by a single specimen (0.04%). With respect to species richness, Monogenea was the most representative group, with eight species, thus contributing to half of parasitic richness estimated to this host species for this geographical location. Digenea was the second most representative group with four species (25%), followed by Nematoda with two species (12.5%), Acanthocephala and Aspidobothrea each represented by a single species (6.25%).
The species classification, following the value of importance, is indicated in Table 1. Only one species, R. arietinus, was considered core; K. eirasi, T. parvus, U. paradoxus, and Urocleidoides sp. were classified as secondary and the others were classified as satellite.
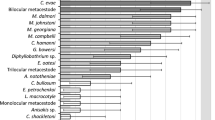
The analysis of co-occurrences of helminth species, using the Spearman correlation test (Fig. 2), showed that 20 pairs of species were formed, five formed by endoparasites, and 15 by ectoparasites. Just a pair of species (J. piava–R. arietinus) showed a significant negative correlation, when J. piava was found in fish gills, R. arietinus was absent in the nose. However, although this result is significant, the value of this correlation is low. On the other hand, the affinity test was performed with 28 pairs of species of the gastrointestinal tract and 15 pairs of species parasitizing host gills. Only three pairs of affinities obtained significance (p ≤ 0.05), all occurring in the gills. The relationships between T. parvus with U. paradoxus (IAB = 0.677; t = 2.596), T. parvus with Urocleidoides sp. (IAB = 0.750; t = 3.498), and U. paradoxus with Urocleidoides sp. (IAB = 0.666; t = 2.397) presented high affinity values.

Co-occurrence of the helminth species of Megaleporinus obtusidens of Lake Guaíba, Rio Grande do Sul, Brazil
The influence of sex, standard length, and weight of the host on the ecological index of the component helminth population
Sixty fish were analyzed, 20 (40%) were males and 40 (60%) were females, and the biometric data are indicated in Table 2. Females were larger (t = − 3.458, p = 0.001) and heavier (t = − 4.941, p < 0.001) than males. The prevalence of U. paradoxus (rr = 0.444, p = 0.048) was significantly higher in females than in males, basing on Fisher’s exact test (Table 3). Regarding the mean intensity, only one species (K. eirasi) presented a significant difference between this index and the size of the host (rs = 0.35, p = 0.03). Other helminths showed no significant influence of host sex on ecological values, and no correlation between the intensity of infection/infestation of any helminth with the weight and/or standard length of the hosts, based on correlation analyses (Tables 3 and 4).
However, abundance had correlation analyses significant in some cases. There was correlation between the abundance and weight of the hosts for U. paradoxus (rs = 0.312, p = 0.015) and Urocleidoides sp. (rs = 0.262, p = 0.044) (Table 4). We also observed correlation between the standard length of the fish and the abundance of U. paradoxus (rs = 0.315, p = 0.014); however, this correlation value was low (< 0.5) (Table 4).
Significant correlation between body weight of the host and the prevalence of four species of helminths was found (Table 5). Creptotrema lynchi was the only species which presented a significant negative correlation (r = − 0.827, p = 0.042). Urocleidoides paradoxus (r = 0.947, p = 0.004) and Urocleidoides sp. (r = 0.841, p = 0.036) presented a positive significant correlation between body of the host and prevalence of helminths as well as the nematode Cucullanus sp. (r = 0.818, p = 0.047). According to the correlation analyses, the standard length of hosts and prevalence of helminths were not significant (Table 5).
The GLM analysis showed a significant difference between the abundance and the parasite species (F = 3.1769; p < 0.001), where two species had higher infection rates, being C. lynchi and R. arietinus (Table 6). In addition, there was a significant influence on the abundance of S. godoyi when the sex and species factors were analyzed (F = 2.3609; p < 0.01), as well as when the weight, sex, and species factors were analyzed (F = 3.7428; p < 0.001) (Table 6).
Diversity index component community
The richness of parasite communities in males and females was similar for the 14 species of helminths (Table 7). Genarchella genarchella and Echinorhynchus sp. were recorded only in females. Aspidogastridae gen. sp. and Dactylogyridae gen. sp. were recorded only in male hosts. When the same analysis was performed, but separately for endo- and ectoparasites, seven species of endoparasites were observed in males, while in females, eight species were found (Table 7). Regarding ectoparasites in male hosts, seven species were found, while in female hosts six species were found (Table 7); this difference was not significant.
The helminth component community was analyzed with the aid of the diversity index, dominance, and evenness (Table 7). Differences between the ecological indices were small and were not significant for most of them. For the total parasites in male and female, the Brillouin diversity index showed a significant difference, where females of M. obtusidens showed a greater diversity (t = − 2.379, p = 0.021) than males (Table 7).
When ectoparasites of hosts of different sexes were analyzed, there was a significant difference in the Equitability Index Shannon–Wiener (t = − 2.535, p = 0.014), which was higher for female hosts. Likewise, the modified Hill’s evenness index showed differences between males and females (t = − 2.224, p = 0.029), in which males had the higher evenness. For the other indices compared, there were no significant differences.
Discussion
Component community structure
A narrow 3-month time window was used in this study on parasite diversity in M. obtusides, and continuation of the work should involve more time and all the seasons of the year. Despite this, our data show an important preliminary view of the component community of the parasites in M. obtusidens.
The large number of helminth species recorded for the first time for Lake Guaíba is probably due to the scarcity of studies carried out with M. obtusidens in this region, and most previous work was focused on gastrointestinal parasites, leaving out the monogeneans, which in the present study contributed with most of the richness.
The structure of helminth populations showed that the ectoparasites, especially monogeneans, were more prevalent than the endoparasites, especially digeneans. This is directly associated with the food habit of “piava,” because this fish feeds mainly on plant material (Hartz et al. 2000), decreasing contamination with endoparasites, which are usually associated with intermediate hosts. However, as previously seen, infection rates were always higher in endoparasites. Shaw and Dobson (1995) justify this result stating that parasitism tends to be a supra-dispersal phenomenon, in which few hosts will have most of the parasite species. The factors that lead to aggregation distribution are not well understood but it is believed that the intrinsic characteristics of each individual, such as agility in predation, age, length, and host weight, are the most likely variables (Anderson and Gordon 1982; Shaw and Dobson 1995; Bandilla et al. 2005; Poulin 2013). In addition, heterogeneity in parasite exposure and susceptibility to them has divided opinions on the causes of aggregation (Poulin 2013). Additionally, the life cycle of the digenean trematodes where there is larval multiplication within the mollusk host explains the high intensity of some species of Digenea (Gibson et al. 2002; Guidelli et al. 2011).
According to Yamaguti (1975), redia and cercariae of G. genarchella develop in snails Littorina australis Gray, 1826, which are ingested by fish where the cercariae will develop to the adult stage. But, Yamaguti also mentioned that fish of the genus Leporinus were incapable of hosting Genarchella, being only an erratic host for this digenean trematode. So, this observation could explain the low prevalence found in the present study, as well as the immaturity of two out of three specimens collected. Other authors mention mollusks and Cypriniform fish as intermediate hosts for this species of parasite (Martorelli 1989; Scholtz et al. 1995; Lefebvre and Poulin 2005), being these, food items little or never consumed by M. obtusidens.
Digeneans of the genus Saccocoelioides have gastropods as intermediate hosts; the metacercariae encyst on aquatic vegetation, and the adults are found in freshwater fish (Díaz and González 1990).
The low prevalence of Echinorhynchus sp. is probably related to its life cycle, to be found by the intermediate host where it will develop to cystacanth to infect the definitive, or the paratenic host (Schmidt and Nickol 1985). Yamaguti (1963) mentioned that species of Echinorhynchus have Ampithoe rubricate Montagu, 1818, Calliopius rathkei (Zaddach, 1844), Pontoporeia jemorata and Cyphocaris challengeri Thorsteinson, 1941 as intermediate hosts while Marcogliese (1995) and Measure and Bossé (1993) found the amphipods Aeginina longicornis (Krøyer, 1843) and Gammarus lawrencianus Bousfield, 1956 infected with Echinorhynchus. The low prevalence observed in our sample may mean that microcrustaceans and/or paratenic hosts of Echinorhynchus sp. are rare in the environment or in the diet of the studied host.
Species of Contracaecum also have microcrustaceans as intermediate hosts, which might explain the low prevalence of this nematode in M. obtusidens. Anderson (2000) reported several intermediate hosts for species of Contracaecum; namely, cnidarians, gastropods, polychaetes, copepods (microcrustaceans), amphipods, decapods, and echinoderms, but copepods are probably the most important hosts for this helminth. Low levels of Contracaecum sp. indicate that the above-mentioned food items are uncommon in the diet of “piava” of Lake Guaíba. These results corroborate those reported by Hartz et al. (2000) when they analyzed the diet of “piava” of Lake Guaíba and reported plants and sediment as the most consumed items, followed by mollusks (Corbicula spp., Ampullaria spp., and Heleobia spp.) and lastly microcrustaceans.
The low prevalence of endohelminths was also observed in previous studies. Guidelli et al. (2011) reported that most digeneans present in M. obtusidens showed no prevalence above 10%, but the intensity range of 1–150 specimens/hosts probably reflects the presence of larvae grouped in intermediate and/or paratenic hosts. The acanthocephalans are among the least reported helminth species in Leporinus, a genus similar to Megaleporinus, when present always showed low prevalence (Guidelli et al. 2011). Octospiniferoides incognita Schmidt and Hugghins, 1973 and Quadrigyrus torquatus Van Cleave, 1920 were recorded in Leporinus lacustris Amaral Campos, 1945 in upper Rio Paraná, with prevalences of 0.66 and 1.33%, respectively.
On the other hand, the ectoparasitic monogeneans showed high prevalence (above 25%) in this study, as in previous studies with species of Leporinus (Guidelli et al. 2006, 2011; Takemoto et al. 2009). These helminths, unlike digeneans, acanthocephalans and nematodes, do not rely on diet or intermediate hosts to complete their life cycle (Thatcher 2006); thus, the parasitic rates are usually higher, because they depend on their active search for the appropriate host. Contact or proximity among the fish facilitates the transference between hosts which corroborates to high indices of infestation. During or near to the reproductive periods, fish are more aggregate, thus facilitating new infestations.
Therefore, the data presented in this study demonstrated that the helminth fauna of Leporinus and Megaleporinus tend to be similar for the most part. This can be explained by the phenotypic plasticity of the parasite species, which may parasitize more than one host species, either because they are hosts with similar characteristics or simply because of the ability to use another resource (Agosta et al. 2010). This has already been explained by Agosta (2006), Agosta et al. (2010), and other authors when they mention the ecological fitting of the parasites, which have the capacity to switch between generalists and specialists throughout their evolution.
The influence of sex, standard length, and weight of the host on the ecological index of the component helminth population
The influence of sex, standard length, and weight of the host on the helminths was tested in a few studies with species Leporinus and Megaleporinus. For L. lacustris and L. friderici, Guidelli et al. (2006) tested these factors and found no significant influence of the parasitic indexes. Feltran et al. (2004) analyzed fish from the Ponte Nova Dam in the State of Minas Gerais, and found no significant correlation between the total intensity of infection of nematodes and the length of L. friderici. However, the same authors found a positive correlation between the length of M. obtusidens with the intensity of nematode infection. In this sense, the present work is configured as one of the few to examine the influence of sex, weight and standard length of hosts on the ecological indices for M. obtusidens. However, there are several other studies that have addressed the relationship of these factors to the parasitic load, where most showed a significant correlation of at least one of the factors on the ecological indices (Takemoto and Pavanelli 2000; Guidelli et al. 2003; Monteiro et al. 2011; Karling et al. 2013).
The significant correlation of U. paradoxus in female hosts could be linked to hormonal factors, or different sexual behavior between male and female hosts. The correlation between abundance of U. paradoxus and the size of the host (weight and length) is related to the quantity of energy resources where bigger fish can support larger parasite infrapopulations. Moreover, variation in body size probably reflects variation in susceptibility, leading to different ecological indices (Poulin 2013). The same seems to happen to larger infrapopulations of Urocleidoides sp. in heavier fish.
The significant relationship between the parasite load and the type of species, demonstrated in the GLM analysis, showed that it is the species itself that defines infection rates and may be linked mostly with the life cycle of the species or with their capacity in infecting the host. As discussed previously, some species have a supra-dispersed distribution as proposed by Shaw and Dobson (1995), where some hosts have a larger parasitic infection, such as C. lynchi. Moreover, the significance of infection rate with host sex and weight is justified by the fact that there is a significant difference in biometric characteristics between males and females.
Diversity index component community
The species richness of Monogenea found in this study for M. obtusidens was similar to that found in other studies using species of Leporinus, resembling them relative to ecological indices and species diversity, possibly due to the phylogenetic proximity between both genera. Previous studies on helminth fauna in Leporinus spp., found species of Tereancistrum, Urocleidoides, Jainus, Kritskyia, and Rhinoxenus, all are monogeneans (Pavanelli et al. 1997; 2008; Takemoto et al. 2009; Guidelli et al. 2011). Species of these genera also are included in the helminth component community of piava from Lake Guaíba. Such genera probably are more adapted to species of Leporinus and Megaleporinus and so are often found in these hosts, both in the basin of Guaíba as Paraná, corroborating several studies that affirm the existence of specific inter host–parasite relationships in Monogenea (Boeger and Kritsky 1997; Desdevises et al. 2002a, 2002b; Braga et al. 2014).
The number of digenean species reported in the present study was similar to other studies (Pavanelli et al. 1997; 2008; Takemoto et al. 2009; Guidelli et al. 2011). However, regarding the parasite diversity, surveys of M. obtusidens from the Paraná River showed the presence of different species, such as Herpetodiplostomum sp., Megacoelium sp., Neodiplostomum sp., and Paralecithobotrys brasiliensis Freitas, 1947, which were not found in Lake Guaíba. Nematodes in other studies had greater richness than in the present study, with seven species reported in M. obtusidens from the upper Paraná River (Takemoto et al. 2009; Guidelli et al. 2011). In this sense, the latitudinal gradient is probably one factor that could be interfering with the results found, such as differences in temperature and/or other abiotic conditions. Fish diet is another important factor that can account for the differences found related to the parasitic infrapopulations (Poulin 2001). In addition, the sampling effort becomes one factor of extreme importance to compose the species richness of a region, collections with a seasonal monitoring or repeated during several years can include rare helminth species and/or seasonal differences in helminths population densities.
Still, it is noteworthy that Aspidogastrea, Echinorhynchus sp., and Cucullanus sp. were reported in M. obtusidens and Lake Guaíba for the first time. The helminth fauna including both ecto- and endo species, found in M. obtusidens in the present study had the third largest richness (Feltran et al. 2004; Takemoto et al. 2009; Chemes and Takemoto 2011; Guidelli et al. 2011). Guidelli et al. (2011) found 28 helminth species in Leporinus friderici Bloch, 1794, followed by 27 species in L. lacustris, 21 in M. obtusidens and 21 in Leporinus elongatus (synonyms for Megaleporinus elongatus) Valenciennes, 1850. Takemoto et al. (2009) found 20 species of helminths in M. obtusidens in the Paraná river.
The present study showed a clear difference between the population organization of endo- and ectoparasites. Endoparasites showed lower prevalence than the ectoparasites, which may be associated with the eating habits of this fish host which feed mainly on vegetation. It was also observed that some species of helminths had their indexes influenced by biotic host factors (weight, standard length, and/or sex).
References
Agosta SJ (2006) On ecological fitting, plant–insect associations, herbivore host shifts, and host plant selection. Oikos 114:556–565
Agosta SJ, Janz N, Brooks D (2010) How specialists can be generalists: resolving the “parasite paradox” and implications for emerging infectious disease. Zoologia (Curitiba) 27(2):151–162
Amato JFR, Boeger WA, Amato SB (1991) Protocolos para laboratório: coleta e processamento de parasitos de pescado. Imprensa Universitária, UFRRJ, Seropédica, p 81
Amato JFR, Amato SB (2010) Técnicas gerais para coleta e preparação de helmintos endoparasitos de aves. In: Von Matter S et al (eds) Ornitologia e conservação: ciência aplicada, técnicas de pesquisa e levantamento. Technical Books, Rio de Janeiro, pp 369–393
Anderson RC (2000) Nematode parasites of vertebrates: their development and transmission. Wallingford, CABI Publishing, 2nd ed., XX+p. 650
Anderson RM, Gordon DM (1982) Processes influencing the distribution of parasite numbers within host populations with special emphasis on parasite-induced host mortalities. Parasitology 85:373–398
Araya PR (1999) Estudio preliminar de edad y crecimiento de La boga Leporinus obtusidens (Pisces, Anostominae), em um tramo Del rio Alto Paraná, Argentina. Revista de Ictiología 7: 59–65
Araya PR, Agostinho AA, Bechara JA (2005) The influence of dam construction on a population of Leporinus obtusidens (Valenciennes, 1847) (Pisces, Anostomidae) in the Yacyret’a reservoir (Argentina). Fish Res 74:198–209
Bandilla M, Hakalahti T, Hudson PJ, Valtonen ET (2005) Aggregation of Argulus coregoni (Crustacea: Branchiura) on rainbow trout (Oncorhynchus mykiss): a consequence of host susceptibility or exposure? Parasitology 130:169–176
Barber I, Poulin R (2002) Interactions between fish, parasites and disease. in P J B Hart & J D Reynolds (eds), The handbook of fish and fisheries. Wiley, pp. 359–389
Boeger WA, Kritsky DC (1997) Coevolution of the Monogenoidea (Platyhelminthes) based on a revised hypothesis of parasite phylogeny. Int J Parasitol 27(12):1495–1511
Braga MP, Araújo SBL, Boeger WA (2014) Patterns of interaction between Neotropical freshwater fishes and their gill Monogenoidea (Platyhelminthes). Parasitol Res 113:481–490
Britski HA, Birindelli JLO, Garavello JC (2012) A new species of Leporinus Agassiz, 1829 from the upper Rio Paraná basin (Characiformes, Anostomidae) with redescription of L. elongates Valenciennes, 1850 and L. obtusidens (Valenciennes, 1837). Papéis Avulsos de Zoologia 52(37):441–475
Bush AO, Holmes JC (1986) Intestinal helminths of lesser scaup ducks: patterns of association. Can J Zool 64(1):132–141
Bush AO, Aho JM, Kennedy CR (1990) Ecological versus phylogenetic determinants of helminth parasite community richness. Evol Ecol 4:1–20
Bush AO, Lafferty KD, Lotz JM, Shostak AW (1997) Parasitology meets ecology on terms: Margolis et al. revisited. J Parasitol 83(4):575–583
Chemes SB, Takemoto RM (2011) Diversity of parasites from Middle Paraná system freshwater fishes, Argentina. International Journal of Biodiversity and Conservation 3(7):249–266
Desdevises Y, Morand S, Jousson O, Legendre P (2002a) Coevolution between Lamellodiscus (Monogenea, Diplectanidae) and Sparidae (Teleostei) the study of a complex host–parasite system. Evolution 56(12):2459–2471
Desdevises Y, Morand S, Legendre P (2002b) Evolution and determinants of host specificity in the genus Lamellodiscus (Monogenea). Biol J Linn Soc London 77:431–443
Diaz MT, Gonzalez GT 1990 Ciclo de vida de Saccocoelioides tarpazensis n.sp. Trematoda: Haploporidae). Acta Científica Venezolana 41:327–336
Gibson DI, Jones A, Bray RA (2002) Keys to the Trematoda. Vol 1. 521p. London, CABI Publishing, Oxon Wallingford and The Natural History Museum
Guidelli GM, Isaac A, Takemoto RM, Pavanelli GC (2003) Endoparasite infracommunities of Hemisorubim platyrhynchos (Valenciennes, 1840) (Pisces: Pimelodidae) of the Baía river, upper Paraná river floodplain, Brazil: specific composition and ecological aspects. Braz J Biol 63(2):261–268
Guidelli G, Tavechio WLG, Takemoto RM, Pavanelli GC (2006) Fauna parasitária de Leporinus lacustris e Leporinus friderici (Characiformes, Anostomidae) da planície de inundação do alto rio Paraná, Brasil. Acta Scientiarum Biological Sciences 28(3):281–290
Guidelli G, Tavechio WLG, Takemoto RM, Pavanelli GC (2011) Relative condition factor and parasitism in anostomid fishes from the floodplain of the Upper Paraná River, Brazil. Vet Parasitol 177:145–151
Fager E (1957) Determination and analysis of recurrent groups. Ecology 38:586–595
Feltran RB, Júnior OM, Pinese JF, Takemoto RM (2004) Prevalência, abundância, intensidade e amplitude de infecção de nematoides intestinais em Leporinus friderici (Bloch,1794) e L. obtusidens (Valenciennes, 1836) (Pisces, Anostomidae), na represa de Nova Ponte (Perdizes, MG). Revista Brasileira Zoociências 6(2):169–179
Hartz SM, Silveira CM, Carvalho S, Villamil C (2000) Alimentação da piava, Leporinus obtusidens (Characiformes, Anostomidae), no Lago Guaíba, Porto Alegre, Rio Grande do Sul, Brasil. Pesq Agrop Gaúcha 6(1):145–150
Karling LC, Lacerda ACF, Alvarenga FMS, Takemoto RM, Pavanelli GC (2013) Endoparasites of Rhaphiodon vulpinus (Characiformes: Cynodontidae) and their relation to the standard length and sex of their hosts. Biosci J 29(5):1311–1316
Kohn A, Moravec F, Cohen SC, Canzi C, Takemoto RM, Fernandes BMM (2011) Helminths of freshwater fishes in the reservoir of the Hydroelectric Power Station of Itaipu, Paraná, Brazil. Check List 7(5):681–690
Lefebvre F, Poulin R (2005) Progenesis in digenean trematodes: a taxonomic and synthetic overview of species reproducing in their second intermediate hosts. Parasitology 130(6):587–605
Magurran AE (2004) Measuring biological diversity. VIII+256p. Malden, Blackwell Publishing
Marcogliese DJ (1995) Aeginina longicornis (Amphipoda: Caprellidea), New intermediate host for Echinorhynchus gadi (Acanthocephala: Echinorhynchidae). J Parasitol 80(6):1043–1044
Martorelli SR (1989) Estudios parasitológicos en biotopos lénticos de la República Argentina V: Desarrollo del ciclo biológico monoxeno de la metacercaria progenética de Genarchella genarchella Travassos, 1928 (Digenea, Hemiuridae) parásita de Littoridina parachappei (Mollusca, Hydrobiidae). Revista del Museo de la Plata 14:109–117
McCullagh P (1984) Generalized linear models. Eur J Oper Res 16:285–292
Mello RF, Moura MAM, Vieira I, Cyrino JEP (1999) Suplementação da dieta de alevinos de piauçu (Leporinus obtusidens) com vitamina C. Sci Agric 56(4):1223–1231
Measure LN, Bossé L (1993) Gammarus lawrencianus (Amphipoda) as intermediate host of Echinorhynchus salmonis (Acanthocephala) in an Estuarine environment. Can J Fish Aquat Sci 50(10):2182-2184. https://doi.org/10.1139/f93-244
Monteiro CM, Amato JF, Amato SB (2011) Helminth parasitism in the Neotropical cormorant, Phalacrocorax brasilianus, in southern Brazil: effect of host size, weight, sex, and maturity state. Parasitol Res 109(3):849–855
Moraes BS, Loro VL, Pretto A, Fonseca MB, Menezes C, Marchesan E, Reimche GB, Avila LA (2009) Toxicological and metabolic parameters of the teleost fish (Leporinus obtusidens) in response to commercial herbicides containing clomazone and propanil. Pestic Biochem Physiol 95(2):57–62
Nering MB, Von Zuben CJ 2010 Métodos quantitativos em parasitologia. Jaboticabal, Funep, 72p.
Oldani N, Iwaszkiw J, Padín O, Otaegui A (1992) Fluctuaciones de la abundancia de peces en el Alto Paraná (Corrientes, Argentina). Publicaciones de La Comisión Administradora del Río Uruguay. Série Técnico-Científica 1:43–55
Pavanelli GC, Machado MH, Takemoto RM (1997) Fauna helmíntica de peixes do rio Paraná, região de Porto Rico, Paraná. In: Vazzoler AEAM, Agostinho AA, Hahn NS (eds) A Planície de inundação do Alto rio Paraná: aspectos físicos, biológicos e sócioeconômicos. Eduem, Maringá, pp 307–329
Pavanelli GC, Eiras JC, Takemoto RM (2008) Doenças de peixes: profilaxia, diagnóstico e tratamento. 3ª Ed. Maringá, Editora da Universidade Estadual de Maringá, 311p
Penchaszadeh PE, Darrigran G, Angulo C, Averbuj A, Brogger M, Dogliotti A, Pirez N (2000) Predation of the invasive freshwater mussel Limnoperna fortunei (Dunker, 1857) (Mytilidae) by the fish Leporinus obtusidens Valenciennes, 1846 (Anostomidae) in the Rio de La Plata, Argentina. J Shellfish Res 19(1):229–231
Piana PA, Baumgartner G, Gomes LC (2003) Influência da temperatura sobre o desenvolvimento de juvenis de piapara (Leporinus cf. obtusidens). Maringá. Acta Scientiarum: Biological Sciences 25(1):87–94
Poulin R (2001) Another look at the richness of helminth communities in tropical freshwater fish. J Biogeogr 28(6):737–743
Poulin R (2013) Explaining variability in parasite aggregation levels among host samples. Parasitology 140:541–546
Ramirez JL, Carvalho-Costa LF, Venere PC, Carvalho DC, Troy WP, Galetti PM (2016) Testing monophyly of the freshwater fish Leporinus (Characiformes, Anostomidae) through molecular analysis. J Fish Biol 88:1204–1214
Ramirez JL, Birindelli JLO, Galetti PM (2017) A new genus of Anostomidae (Ostariophysi: Characiformes): diversity, phylogeny and biogeography based on cytogenetic, molecular and morphological data. Mol Phylogenet Evol 107:308–323
Reis RE, Kullander SO, Ferraris JC (2003) Check list of the freshwater fishes of South and Central America. EDIPUCRS, Porto Alegre, p 742
Rodrigues WC (2015) DivES—Diversidade de Espécies v3.0—Guia do Usuário. Entomologia do Brasil. p. 30. Disponível em: http://www.dives.ebras.bio.br
Santos GO (2000) Aspectos biológicos importantes para a piscicultura do gênero Leporinus Spix, 1829—Uma revisão. Pesq Agrop Gaúcha 6(1):151–156
Schmidt GD, Nickol BB (1985) Development and life cycles. In: Crompton DWT, Nickol BB (eds) Biology of the Acanthocephala. Cambridge University Press, Cambridge, p 519
Scholtz T, Vargas-Vasquez J, Salgado-Maldonado G (1995) Revision of Genarchella species (Digenea: Derogenidae) parasitizing freshwater fishes in Mexico and Central America. J Nat Hist 29:1403–1417
Shaw DJ, Dobson AP (1995) Patterns of macroparasite abundance and aggregation in wildlife populations: a quantitative review. Parasitology 111:111–113
Sidlauskas BL, Vari RP (2008) Phylogenetic relationships within the South American fish family Anostomidae (Teleostei, Ostariophysi, Characiformes). Zool J Linnean Soc 154:70–210
Takemoto RM, Pavanelli GC (2000) Aspects of the ecology of Proteocephalid Cestodes parasites of Sorubim lima (Pimelodidae) of the upper Paraná river, Brazil: I. Structure and influence of host’s size and sex. Rev Bras Biol 60(4):577–584
Takemoto RM, Pavanelli GC, Lizama MAP, Lacerda ACF, Yamada FH, Moreira LHA, Ceschini TL, Bellay S (2009) Diversity of parasites of fish from the Upper Paraná River floodplain, Brazil. Braz J Biol 69(2):691–705
Thatcher VE (2006) Amazon of fish parasites, 2nd edn. Pensoft, Sofia, p 496
Yamaguti S (1963) Systema Helminthum. Vol. V. Acanthocephala. Interscience Publishers, New York, p 423
Yamaguti S (1975) A synoptical review of life histories of digenetic trematodes of vertebrates with special references to the morphology of their larval forms, First edn. Keigaku Publishing Company, Tokyo, pp 590–2190
Zaniboni-Filho E, Schultz UH (2003) Migratory fishes of the Uruguay River. In: Carosfeld J, Harvey B, Ross C, Baer A (eds) 372Migratory fishes of South America. World Fisheries Trust, Canada, pp 157–194
Zar JH (1996) Biostatistical analysis. Prentice-Hall Inc., Upper Saddle River, New Jersey. 918.
Acknowledgments
Emília Wendt thanks the “Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq)” for the scholarship, Dr. José Felipe Amato for valuable considerations and corrections, and Dra. Carolina Santos Vieira for valuable help with statistical analysis.
Author information
Authors and Affiliations
Corresponding author
Additional information
Section Editor: Guillermo Salgado-Maldonado
Electronic supplementary material
ESM 1
(DOCX 21 kb)
Rights and permissions
About this article

Cite this article
Wendt, E.W., Monteiro, C.M. & Amato, S.B. Helminth fauna of Megaleporinus obtusidens (Characiformes: Anostomidae) from Lake Guaíba: analysis of the parasite community. Parasitol Res 117, 2445–2456 (2018). https://doi.org/10.1007/s00436-018-5933-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-018-5933-4