
Abstract
The engagement of the cerebellum during reading tasks is not unprecedented. However, it is still unclear which regions in the cerebellum are specifically involved in reading and how the cerebellum processes different languages. With functional magnetic resonance imaging, we compared the cerebellar neural activity in Chinese child learners of English between reading and non-reading tasks to identify functionally specialized areas for reading, and between Chinese characters and English words in a passive viewing paradigm to detect regions sensitive to different scripts. Two posterior subregions of right lobule VI, as well as right lobule VIIIA, demonstrated greater activation to viewing Chinese characters and English words compared to the non-reading tasks. However, we did not find any cerebellar regions that were differentially responsive to Chinese versus English print. Instead, we observed that functional connectivity between the two above-mentioned cerebellar regions (lobules VI and VIIIA) and the left inferior parietal lobule was significantly greater in English reading compared to Chinese reading. Overall, these results indicate that the posterior parts of right lobule VI and the right lobule VIIIA could be reading-specific regions, and deepen our understanding of how the cerebellum contributes to reading.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Learning to read changes the mind and brain, resulting in a complex cerebral network connecting frontal, parietal, and occipitotemporal regions (Dehaene et al. 2010). Decades of research have supported our understanding of this universal reading network in the cerebrum (Rueckl et al. 2015). Less well understood, however, is the engagement of the cerebellum during word reading. This study extends a burgeoning body of work about the functional specificity of cerebellum. In particular, we ask: are there specific cerebellar subregions that demonstrate specificity for reading, and are these regions sensitive to different languages or orthographies?
In recent years, accumulating studies have observed remarkable activation in the cerebellum during reading or reading-related tasks (Moore et al. 2017; Stoodley and Stein 2013; Mariën et al. 2014). In particular, the right lobule VI and its neighboring region, Crus I, have gained attention due to their association with core reading-related processes (Turkeltaub et al. 2002; Cattinelli et al. 2013; Martin et al. 2015). For instance, the right lobule VI has been consistently associated with phonological processing during English tasks (Tan et al. 2005). Other studies suggest that the right cerebellar lobules, including Crus I, may be involved in semantic integration or prediction (Gatti et al. 2020; D'Mello et al. 2017). Moreover, these regions are functionally connected with the cerebral regions associated with phonological and semantic processing (Gao et al. 2015; Booth et al. 2007; Alvarez and Fiez 2018), and structural or functional deficits in the right lobule VI /Crus I have been associated with reading impairments (Stoodley 2014; Eckert et al. 2016; Feng et al. 2017; Hancock et al. 2017). These findings jointly suggest that the cerebellum, and the right lobule VI/Crus I in particular, may warrant greater attention in our theories of literacy.
Functional connectivity analyses have also revealed cerebro-cerebellar connections that are associated with literacy-related skills. For instance, resting-state functional connectivity between the right lobule VI and the left supramarginal gyrus is related to rapid naming skill (Ang et al. 2020), a critical factor in early reading success. Moreover, a recent study identified six functional subregions in the right lobule VI by examining functional connectivity between each region and the cerebrum in Chinese child readers (Li et al. 2020). Some subregions, located at the anterior part of the right lobule VI, were preferentially connected with the cerebral sensory-motor network, while the posterior parts of the right lobule VI were preferentially connected with regions within the cerebral language network. However, it remains unclear whether functional activity in these subregions in the right lobule VI is specific to literacy, or whether these regions might be equally engaged during other non-linguistic tasks.
Beyond the involvement of the right lobule VI/Crus I, several other cerebellar regions, such as the left lobule VI and right lobule VIII (consisting of VIIIA or VIIIB), have also been reported in reading research. For example, Wu et al. (2012) reported consistent activation of the left posterior part of lobule VI in tasks involving orthographic, phonological, and semantic processing. Moore et al. (2017) observed a significant correlation between gray matter volume in the right lobules VIIIA and VIIIB and reading performance. Yet while these regions were engaged in literacy tasks, it remains unknown whether they are incidental or active participants in reading. The present study investigates these gaps in our understanding, asking if any other cerebellar regions outside the right lobule VI, may be selectively involved in reading.
Previous studies have mainly investigated the relationship between the right lobule VI and reading in a single language (Ang et al. 2020; Feng et al. 2017; Li et al. 2020). However, an increasing number of people begin to learn a second language at a very young age. Furthermore, research focusing on the cerebrum has revealed language-specific differences in bilingual reading networks, particularly for bilingual speakers of Chinese and English (e.g., Cao et al. 2014; Ip et al. 2019; Xu et al. 2017). For example, when comparing the brain bases of first and second language reading across bilingual language pairings, studies suggest the left inferior parietal lobe may be particularly sensitive to second-language reading (Barbeau et al. 2017; Li et al. 2021b). As the cerebellum has rarely been included in neurobiological models of literacy, it remains unknown whether the cerebellum may adapt to different language experiences like the cerebrum.
Therefore, in the current study, we aimed to address three questions. First, are there functional subregions in the right lobule VI that are functionally specific to reading? Second, beyond the right lobule VI, are there any other reading-specific regions? And third, if so, how might these regions contribute to reading in different languages?
To address these issues, we examined brain activation during visual word processing in a sample of Chinese-speaking children (aged 8–12) who were learning English as a second language in school. We exploited a passive reading paradigm adapted from Dehaene et al. (2010), during which the participants were required to view linguistic (Chinese characters and English words) and non-linguistic stimuli (i.e., faces, houses, and tools). These first two conditions served as an implicit reading task, whereas the latter conditions were characterized as a non-reading control. We first compared the neural activity of each functional subregion in the right lobule VI between the reading and non-reading tasks with regions of interest analyses (ROI-based analysis). We then conducted a whole cerebellum analysis to detect reading-selective regions beyond the right lobule VI (voxel-based analysis on the whole cerebellum). Finally, we examined the possible effect of language in the cerebellum during reading. To this end, we compared cerebellar activation, cerebro-cerebellar connection patterns, and brain-behavior correlations associated with different languages (i.e., Chinese and English). These complementary methods aimed to comprehensively reveal the specific engagement of the cerebellum in reading across languages.
Materials and methods
Participants
Fifty-one children, aged 8.7–12.5 years (31 girls), participated in this study. Subjects from third to sixth grade were recruited from several primary schools in Beijing, China. Participants were from a large cohort with a sample size of 107, in which 55 subjects received fMRI scanning with the cerebellum fully covered. Among these children, three were excluded because of their excessive head movements (via Artifact Repair toolbox, see section “Data processing”, step 2), and one was excluded due to missing demographic information.
All participants were native Chinese speakers who were learning English as a second language in school. All children had received formal reading instruction in Chinese since 1st grade (around age six) and English since 3rd grade (around age eight). The majority of the sample had received formal reading instruction in English for at least one year, while eight participants were in the 3rd grade and had received less than half a year of English instruction. All participants were right-handed, according to self-report (Edinburgh Handedness Inventory, Oldfield 1971). None of them had hearing impairments or any psychiatric disease history. All reported normal or corrected-to-normal vision. This study was approved by the Ethical Review Board of Beijing Normal University. Before the experiment, written consent was obtained from each participant and his/her guardians.
Behavioral measures
The Raven's Standard Progressive Matrices was used to tap into children's non-verbal IQ (Raven and Court 1998) with standard scores obtained based on Chinese norm (Zhang and Wang 1985). Children with typical intelligence, defined as above or equal to the 50th percentile on this test, were recruited in the current study (Feng et al. 2017; Li et al. 2020; Ang et al. 2020). Children also completed a battery of Chinese and English literacy assessments.
The tests of Chinese literacy consisted of a Chinese word reading test (Feng et al. 2017; Li et al. 2018; Xia et al. 2016) and the standardized Chinese Character Recognition Test (Wang and Tao 1993). The Chinese word reading test contains 150 characters arranged in order of difficulty. Children were asked to read each of them aloud until 15 consecutive errors were made. The standardized Chinese Character Recognition Test includes 210 Chinese characters. The participants were asked to make a word or phrase based on the given characters/morphemes. For example, '大/ da4/' (meaning ‘big’) can be used in the word '伟大/ wei2da2 /' (meaning ‘greatness’). This test was time limited to 40 min, and children were asked to write as many words as possible.
The tests of English literacy consisted of a word reading test and an English spelling test. The Word Identification subtest of the Woodcock Diagnostic Reading Battery (Woodcock 1987) was used to measure English reading ability. In this test, the participants were asked to read each word (or letter) aloud until they made six consecutive errors. The English spelling test was a dictation test, comprised of 40 words. Each word was read twice by a recording, during which children were asked to write down these words on an answer sheet (Feng et al. 2017; Li et al. 2018; Liu et al. 2016; You et al. 2011).
Experiment design and tasks
Participants performed an incidental target-detection task in the scanner. This task was adapted from a passive viewing paradigm designed by Dehaene et al. (2010), which has been widely used to investigate neural correlates of reading (Dehaene et al. 2010; Dehaene-Lambertz et al. 2018; Monzalvo et al. 2012; Feng et al. 2019). The original paradigm was a block design consisting of seven types of stimuli, including letter strings, false fonts, faces, houses, tools, stars, and checkerboards.
The current study contains five main visual categories: Chinese characters, English words, faces, houses, and tools. The Chinese characters and English words were from children’s textbooks, to ensure that they were all familiar to our participants. The meanings of English words and Chinese characters were not the same, and no words or characters presented were similar to concepts presented in the other visual stimuli. House and tool stimuli were the same as Dehaene et al. (2010), and faces were changed into those of Chinese people (Zhang et al. 2015). All words/characters were black on a white background, while faces, houses, and tools were all highly contrasted gray-level photographs matched for size and overall luminance. In addition, a ring-shaped checkerboard condition was also included to reduce visual fatigue. To maintain children’s attention, participants were asked to focus on the visual stimuli in the center of the screen and to press a button whenever they saw a star, which appeared at random intervals on either the left or right side of the screen.
Stimuli were presented via E-prime 2.0, which was similar to Dehaene et al. (2010). Children completed two task runs of approximately 4 min and 40 s. Each run consisted of 11 blocks: two blocks for each of the five main visual categories, plus one checkerboard block. Each block consisted of 17 trials, including five fixation cross trials, ten image/word trials, and two star trials. For each trial, a pair of stimuli from the same category was presented sequentially. After 200ms fixation, the first element was presented for 200 ms, followed by a 200 ms fixation point, which was followed by the second element for 500 ms, and finally a 400 ms fixation point.
In this paradigm, passive viewing of Chinese characters and English words was regarded as the implicit reading task, whereas passive viewing of other stimuli served as the non-reading visual task. The current implicit task is well suited to examine cerebellar activity, because it does not induce explicit articulation or hand motion. In addition, this task is quite simple, requiring relatively comparable processing demands for Chinese characters and English words. This advantage allows us to investigate regions sensitive to different scripts without the influence of task difficulties associated with processing different languages.
Scan acquisition
Whole-brain images were acquired with a 3 T Siemens Trio Scanner at Beijing Normal University, using a T2*-weighted echo planar imaging (EPI) sequence. The parameters were as follows: flip angle (FA) = 81°; echo time (TE) = 30 ms; repetition time (TR) = 2400 ms; field of view (FOV) = 200 mm; voxel size = 3 × 3 × 4 mm, 40 slices. T1-weighted images were also acquired using the following parameters: FA = 7°; TE = 4.18 ms; TR = 2300 ms; FOV = 256 mm; voxel size = 1.3 × 1.0 × 1.3 mm, 128 slices.
Data processing
fMRI data pre-processing and first-level analyses of the cerebellum
The data were pre-processed with SUIT (Diedrichsen et al. 2009) implemented in Statistical Parametric Mapping, version 12 (SPM12; Welcome Department of Cognitive Neurology, London), which offers a spatially unbiased atlas template of the cerebellum. This toolbox has been highly recommended in the study of the cerebellum (Diedrichsen 2006; Diedrichsen et al. 2009). Detailed pre-processing steps are as follows:
(1) Functional images were slice-time corrected and realigned. (2) Outlier volumes were repaired using Artifact Repair toolbox software (Mazaika et al. 2009; Younger et al. 2017), which has been widely used in fMRI studies with children (Dehaene-Lambertz et al. 2018; Younger et al. 2017; Weiss et al. 2018; Ashburn et al. 2020; Battista et al. 2018; Karipidis et al. 2018; Langer et al. 2015; Cherodath and Singh 2015). Outlier volumes were defined as volumes whose global intensity was larger than 1.5% and volumes with scan-to-scan motion exceeding 0.5 mm (Karipidis et al. 2018), and were replaced using linear interpolation of the nearest two non-outlier volumes (Weiss et al. 2018; Paz-Alonso et al. 2018; Mazaika et al. 2009; Dehaene-Lambertz et al. 2018). (3) Functional images and structural images were co-registered; (4) First-level analyses were conducted in native space. The onset times and durations across runs were extracted to build a general linear model for each category (Chinese characters, English words, faces, houses, tools, checkboards, and star trials) with six motion parameters and outliers as covariates (outliers information associated with each category was presented in Table S1 and Fig. S2). Notably, three kinds of contrasts were defined during this stage. The first was based on the comparisons between each condition of interest (i.e., Chinese characters, English words, faces, houses, or tools) and fixation. The second was between implicit reading tasks (Chinese characters and English words) and non-reading conditions (faces, houses, and tools) to detect regions specific to reading, and the third was between Chinese characters and English words to investigate language effects.
(5) After first-level analysis, structural images were segmented into gray matter, white matter, and cerebrospinal fluid. (6) Images were co-registered to the cerebellum template implemented in SUIT. (7) Functional images obtained from the first-level analysis were resliced to the standard space. Finally, (8) data for activation analyses were smoothed with a 3 mm full width at half maximum kernel.
Three participants were excluded from analyses due to a high number of outliers (> 25% of all volumes). The mean number of repaired volumes was 8.6 (SD = 9.3) in the first run and 13 (SD = 12.3) in the second run. We conducted a two-way repeated ANOVA analysis to compare the outliers across conditions and runs. The results showed that the main effect of condition was significant (F(4, 368) = 9.17, p < 0.001). Specifically, outliers in passive viewing of houses were significantly lower than the other four conditions, whereas there were no significant differences between the other four conditions. In addition, a significant main effect of runs (F(1, 92) = 16.3, p < 0.001) was observed, with outliers in run 2 significantly higher than run 1 (Meanrun1 = 0.7, SDrun1 = 0.093; Meanrun2 = 1.1, SDrun2 = 0.15), which could be attributed to visual fatigue. The interaction between runs and conditions was not significant.
fMRI data pre-processing and first-level analyses of the cerebrum
As a comparison, we also pre-processed imaging data in the cerebrum. Data pre-processing and first-level analyses were almost the same as in the cerebellum, except for the order of the first-level analyses. For the cerebrum, detailed information about data pre-processing was as follows:
(1) Functional images were slice-time corrected and realigned. (2) Outlier volumes were repaired using Artifact Repair toolbox software. (3) Functional images and structural images were co-registered. (4) Structural images were segmented into gray matter, white matter, and cerebrospinal fluid. (5) Functional images were spatially normalized to Montreal Neurological Institute (MNI) space. (6) Imaging data for activation analyses were smoothed with a 3 mm full width at half maximum kernel. (7) First-level analyses were conducted. The onset times and durations across runs were extracted to build a general linear model for each category (Chinese characters, English words, faces, houses, tools, checkboards, and star trials) with six motion parameters and outliers as covariates. The same three contrasts (above) were defined during this stage.
Second-level analyses and statistical analyses
Reading-specific activation: We used voxel-based and ROI-based one-sample t tests to estimate task-specific effects in the cerebellum.
Definition of ROIs: Six functional subregions of the right cerebellar lobule VI (Li et al. 2020) were defined as regions of interest.
We then conducted one-sample t tests on each of the contrast images generated during first-level processing. First, we conducted one-sample t tests on the contrast images between each category (Chinese characters, English words, faces, houses, and tools) versus fixation. This analysis was to investigate whether the cerebellum was engaged in processing these visual stimuli. Next, we examined the contrast images between the implicit reading (passive viewing of characters and words) and non-reading tasks (passive viewing of faces, houses, tools), to identify regions specifically engaged in reading. Finally, we examined the contrast images of Chinese characters versus English words to locate regions that were sensitive to different scripts. For the voxel-based analysis, the threshold was set at a voxel level of p < 0.001 uncorrected, cluster level corrected with FWE p < 0.05 (Dehaene-Lambertz et al. 2018; Weiss et al. 2018; Karipidis et al. 2017; Boros et al. 2016). For ROI-based analysis, the threshold was corrected for multiple comparisons with FDR p < 0.05. Adjusted p values are presented in the current study.
Whole-cerebrum functional connectivity analysis: We further investigated functional connectivity (FC) patterns between the cerebrum and the cerebellum using a generalized psycho-physiological interaction (gPPI) toolbox (https://www.nitrc.org/projects/gppi; McLaren et al. 2012). gPPI can accommodate more than two task conditions in the same PPI model, is well suited to estimating FC in block design experiments (Cisler et al. 2014; Michels et al. 2018), and is reported to be more powerful than the conventional PPI analysis. In the current study, regions identified as reading-specific served as seeds (i.e., the posterior part of right lobules VI and VIIIA, see Result Sect.), and the cerebrum was characterized as the target region. For each seed, PPIs targeting all voxels in the cerebrum were calculated with general linear models that included: (1) regressors for each experimental task in the design matrix convolved with the hemodynamic response function (HRF); (2) the time course of the seed region; (3) six movement and volume outlier regressors; (4) PPI interaction terms for each condition; and (5) a constant. Beta values of the PPI interaction term were saved in brain images with each index in each voxel representing FC strength.
To explore the FC differences associated with the two cerebellar areas, and the possibility of connectivity differences for Chinese and English word reading, we conducted a repeated two-way ANOVA, in which seeds and languages were the independent variables and FC was the dependent variable. We focused on the main effect of seeds and languages, as well as the interaction effect between the two. The threshold was set at a voxel level of p < 0.001 and cluster level corrected with FWE p < 0.05.
Brain and behavioral correlation analysis: We were also interested in the extent to which brain activity in the cerebellum was modulated by reading abilities. An index for reading-specific brain activation was estimated using the contrast between reading and non-reading tasks. The threshold was set as voxel level of p < 0.001 and cluster level corrected with FWE p < 0.05.
Bayesian estimation: Finally, we used a Bayesian approach to determine the evidence for the null hypothesis (H0) via the JASP toolbox (https://jasp-stats.org/). This toolbox estimated Bayes’ factors (BF01), which are the ratio between the marginal likelihoods of the null model (corresponding to the null hypothesis, H0) and the alternative model (corresponding to the alternative hypothesis, H1). BF01 less than 3 suggests data insensitivity, indicating anecdotal evidence for the null hypothesis (Leppink et al. 2017; Quintana and Williams 2018). BF01 between 3 and 10 indicates moderate evidence for the null hypothesis, and BF01 larger than 10 indicates strong evidence for the null hypothesis (Quintana and Williams 2018). This approach was applied for six ROIs and two clusters (right lobules VI and VIII in Result Sect.) showing reading specificity driven by voxel-based analysis on the whole cerebellum.
Results
Demographic information and behavioral results
Table 1 shows children's demographic information and behavioral performance.
The validity of the implicit reading task
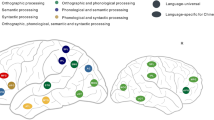
Because passive viewing of words could potentially serve only as a perceptual task rather than a reading task, we compared the neural activity between reading and non-reading tasks in the cerebrum. If this was a purely visual perceptual task, we should only observe significant task-related activity in the cerebral visual areas, such as the bilateral occipital gyri. However, we found significantly greater activation in the passive viewing of linguistic stimuli compared to non-linguistic stimuli in canonical reading-related regions, including the inferior frontal gyrus, temporoparietal junction, and occipitotemporal cortex (Table S2, Fig. 1). Therefore, it is reasonable to believe that the passive viewing of linguistic stimuli in the current study functioned as an implicit reading task rather than a perceptual task, and that the comparison between linguistic and non-linguistic stimuli is valid for detecting reading-related areas.

Reading specificity in the cerebrum. The left panel represents examples of different categories used in the current study, consisting of two linguistic stimuli (Chinese characters and English words) and three other visual stimuli (faces, houses, and tools). The right panel represents regions showing greater activation during passive viewing of linguistic stimuli as compared to the other visual stimuli
Cerebellar activation specific to reading (linguistic stimuli vs. non-linguistic stimuli)
ROI-based univariate analysis
Our first research question asks whether any functional subregions of the right lobule VI were specialized for reading. To address this issue, we conducted ROI-based analyses using the six subregions of right lobule VI (named R1–R6, Fig. 2A) defined by Li et al. (2020). We extracted the mean beta value of each subregion from the contrast between each category and fixation. For reading categories, one-sample t tests showed that passive viewing of both Chinese characters and English words elicited significant activation in R4 (Chinese, t = 2.9, p-adjusted = 0.03; English, t = 3.4, p-adjusted = 0.01). Chinese characters also induced significant activation in R3 (t = 3.0, p-adjusted = 0. 02) and marginally significant activation in R6 (t = 2.3, p-adjusted = 0. 06). For non-linguistic categories, passive viewing of faces and tools induced a similar pattern in these six subregions, with R1, R2, R5 showing significant or marginally significant deactivation (Table S3, Fig. 2B). The activation pattern in the house condition was similar to reading tasks, which showed significant activation in R3 (t = 2.9, p-adjusted = 0. 03) and R4 (t = 2.9, p-adjusted = 0. 02; Table S3, Fig. 2B).
We then extracted the mean beta value of each subregion based on the contrast between the reading and non-reading tasks, and conducted one-sample t tests. The results revealed significant differences in activation in R4 (t = 2.00, p-adjusted = 0. 02) and R6 (t = 3.01, p-adjusted = 0.02, Table S3, Fig. 2C), with greater activation associated with visual word processing as compared to face, house, or tool processing. In contrast, we did not observe any subregions showing greater activation in non-reading tasks compared to reading tasks. These results suggest that the subregions of the right lobule VI are differently engaged in reading. Specifically, the posterior parts of the right lobule VI (i.e., R4 and R6) showed preferences for linguistic stimuli and could be reading-preferring areas. Other regions, such as the anterior part of the right lobule VI (e.g., R1) were not biased towards any of these tasks.
Voxel-based analyses of the whole cerebellum
We further conducted voxel-based analyses on the whole cerebellum. Voxel-wise one-sample t tests on the contrasts between each category and fixation showed that the bilateral cerebellar lobule VI/VII and the right lobule VIIIA were significantly activated during passive viewing of characters, words, faces, tools, and houses (Fig. S3), suggesting that the cerebellum was actively engaged in a variety of visual tasks.
We then conducted voxel-wise one-sample t tests based on the contrast between the reading and non-reading tasks. The results showed significant activation differences in the right cerebellar lobule VIIIA (peak coordinates, x = 26, y = − 64, z = − 51; Z value = 5.02, voxel level, p < 0.001; cluster level, FWE corrected p = 0.006; cluster size = 98; Fig. 2D) and the cerebellar right lobule VI extending to Crus I (peak coordinates, x = 32, y = − 58, z = − 29; Z value = 4.3, voxel level, p < 0.001; cluster level, FWE corrected p = 0.02; cluster size = 76; Fig. 2D), with greater activation in the reading tasks compared to the non-reading tasks. Notably, the right lobule VI overlapped with R4. In contrast, the left cerebellar lobule VI (peak coordinates, x = − 30, y = − 56, z = − 21; Z value = 5.7, voxel level, p < 0.001; cluster level, FWE corrected p < 0.001; cluster size = 139; Fig. 2D) showed significantly greater activation in non-reading tasks compared to reading tasks. These results remained unchanged when age and sex were used as covariates, and when we regressed out the number of outliers for each participant.

Activation difference in the cerebellum between reading and non-reading tasks. A represents six subregions in the right lobule VI defined by Li et al. (2020), which served as ROIs in the current study. B depicts the activation level associated with each category in each ROI (Results Sect.). CH = Chinese characters (red color), EN = English words (blue color), F = faces (brown color), T = tools (orange color), H = houses (green color). C represents the activation difference between two reading tasks (passive viewing of Chinese characters and English words) and three non-reading tasks (passive viewing of faces, houses, or tools). D represents the contrast between two reading conditions (i.e., Chinese characters and English words) and three non-reading conditions (i.e., faces, houses, and tools) based on voxel-based analyses on the whole cerebellum
In the current study, two conditions (passive viewing of Chinese characters and English words) served as implicit reading tasks, whereas three conditions (passive viewing of faces, houses, and tools) were considered as non-reading tasks. This might cause an imbalanced number of trials and different signal-to-noise ratios. Given volume outliers in the house condition differed significantly from the other four conditions, as a complementary analysis, we only used face and tool conditions as the baseline. The above reported significant results held (see Supplementary Materials, Fig.S4). In other words, different baselines did not significantly influence the preference of the right lobules VI and VIII for reading, even at a more lenient threshold with cluster-level uncorrected and voxel-level p < 0.001.
In short, voxel-based univariate analysis showed that the left cerebellar lobule VI demonstrated a bias towards non-linguistic stimuli, whereas the right cerebellar lobules VI and VIIIA demonstrated a bias towards linguistic stimuli. Supplementary multivariate analyses using Representational Similarity Analysis (RSA, see Supplementary Materials, Fig.S5) were consistent with the univariate analysis, again suggesting that the posterior part of right lobules VI and VIIIA could be potential reading-selective regions.
Cerebellar activation for Chinese (L1) vs. English (L2) reading
ROI- and voxel-based univariate analysis
Our second research question asks whether reading-selective regions may respond differently to different languages/orthographies. We addressed this issue by comparing the neural activation patterns induced by Chinese characters and English words and performed both ROI-based analysis and voxel-based analysis on the whole cerebellum.
First, we examined the contrast between Chinese characters and English words in the six functional regions of the right cerebellar lobule VI. One-sample t tests failed to reveal significantly greater activation for Chinese or English reading in any of these ROIs (Fig. 2B). We then conducted voxel-based one-sample t tests in each voxel on the whole cerebellum based on the same contrast. Similarly, no significant differences in activation across languages were observed, even at a more lenient cluster-level uncorrected threshold. Supplementary RSA analyses also failed to reveal any remarkably differences in cerebellar activation for Chinese character reading as opposed to English word reading (see Supplementary Materials).
Finally, we used a Bayesian approach to assess support for the null hypothesis, that the cerebellum is not sensitive to reading in Chinese versus English. We focused on six ROIs in the right lobule VI, and two clusters (right lobule VI and right lobule VIIIA) showing greater activation in reading tasks compared to non-reading tasks, as estimated by the voxel-based analysis on the whole cerebellum level. The Bayes’ factors, presented in Table 2, suggest that our data provide moderate support for the null hypothesis in the right lobules VI, VIIIA, and in regions R1–R5, all with BF01 > 3. Whether or not R6 might be responsive to different languages/orthographies remains an open question. Nevertheless, these findings together suggest that reading-preferring regions (the right lobules VI and VIIIA) might not be sensitive to different scripts.
Cerebro-cerebellar functional connectivity in Chinese vs. English
To further examine how the cerebellum might contribute to different languages, we estimated the FC between the cerebrum and the cerebellum, with the right lobules VI and VIIIA as the seeds and the cerebrum as target regions. A two-way repeated ANOVA analysis was conducted, with two seeds and two languages as the independent variables, and FC as the dependent variable.
We observed a significant main effect of languages and seeds, but no significant interaction effect between the two. Specifically, compared to Chinese characters, passive viewing of English words induced greater FC between the two right cerebellar clusters and the left inferior parietal lobule (L.IPL, peak coordinates, x = − 58, y = − 20, z = 46; Z value = 5.43; voxel level, p < 0.001; cluster level, FWE corrected to p < 0.001; cluster size = 211; Fig. 3, blue color). When age, sex, and outlier numbers were used as covariates, all results reported above remained significant. In contrast, passively reading Chinese did not show any notable greater FC with the cortical areas as compared to English. Notably, in our sample, eight children in the third grade had received less than a year of English reading instruction. Language differences in terms of FC could, thus, be due to the greater effort in reading English words compared to reading Chinese characters in these participants. To address this issue, we conducted the same analysis within the remaining 43 participants. As expected, main results held that FCs between the right cerebellar seeds and L.IPL remained significantly greater in English word processing compared to Chinese word processing.

Contrast of FC (functional connectivity) across languages and seeds. The upper panel indicates regions showing significantly greater FC with two cerebellar clusters in reading English words (EN) compared to reading Chinese characters (CH). The lower panel indicates regions showing different FC with different seeds. L.IPL = left inferior parietal lobule; R.STG = right superior temporal gyrus; R.LG = right lingual gyrus
In terms of the main effect of seeds, we found that the right lobule VI showed significantly greater FC with the right superior temporal gyrus (peak coordinates, x = 66, y = − 38, z = 22; Z value = 4.40; voxel level, p < 0.001; cluster level, FWE corrected to p = 0.030; cluster size = 87; Fig. 3, orange color) and the right lingual gyrus (peak coordinates, x = 6, y = − 72, z = − 2; Z value = 4.44; voxel level, p < 0.001; cluster level, FWE corrected to p = 0.061; cluster size = 74; Fig. 3, orange color). In contrast, the right lobule VIIIA showed greater FC with two subcortical areas, overlapping with the bilateral caudate and left thalamus (Fig. 3, yellow color).
Correlations with reading performance in Chinese vs. English
Finally, we investigated the correlations between the cerebellar activation (estimated by the contrast between reading and non-reading tasks) and reading performance in the different languages, i.e., behavioral performance on the Chinese Character Recognition Test, the Chinese word reading test, the English word spelling test, and the Letter-Word Identification test. However, we did not observe activation in any cerebellar regions showing a significant correlation with Chinese or English reading abilities either with ROI-based analysis or voxel-based analysis on the whole cerebellum.
Discussion
Specificity of the right cerebellar lobules VI and VIIIA to linguistic processing
Our first question asked which subregion(s) in the right lobule VI demonstrated specificity for reading. To answer this question, we compared cerebellar activation during implicit reading (i.e., viewing English words and Chinese characters) and non-reading tasks (i.e., viewing faces, houses, and tools). Only regions in the posterior parts (R4 and R6) of the right lobule VI demonstrated significant preferences for linguistic stimuli, whereas the anterior parts of the right lobule VI did not differentiate between linguistic stimuli and non-linguistic stimuli. We then expanded our inquiry to cerebellar regions outside of the right lobule VI. Similar analyses on the whole cerebellum further showed that in addition to the posterior part of the right lobule VI, the right lobule VIIIA was also more active during passive viewing of linguistic stimuli. These results suggest that the posterior subregion of the right lobule VI and the right lobule VIIIA could be functionally specialized areas for reading.
The result that right cerebellar lobules showed specificity for reading rather than left cerebellar lobules is aligned with prior findings of right dominance for language processing in the cerebellum (Stoodley and Schmahmann 2009). Studies have shown that individuals with tumors in the right cerebellum showed deficits in literacy skills, whereas those with tumors in the left cerebellum presented disturbance or delayed non-verbal skills, such as spatial and visual sequential memory (Riva and Giorgi 2000; Scott et al. 2001). Atypical lateralization for reading in the cerebellum has also been associated with reading impairments (Eckert et al. 2003; Kibby et al. 2008). Rae et al. (2002) found that cerebellar structural asymmetry was only observed in typical readers, with greater gray matter volume in the right cerebellum compared to that in the left cerebellum. In contrast, dyslexic readers showed less cerebellar asymmetry, and the extent of this difference was correlated with the severity of their phonological decoding impairments. This result has been supported by other studies, which also observed a decreased rightward asymmetry pattern in the cerebellum in dyslexic readers (Kibby et al. 2008). The contralateral dominance pattern between the cerebellum and cerebrum could be a by-product of contralateral functional and structural connections (Schmahmann 1996; Sokolov Arseny et al. 2017). Neuroimaging studies showed that reading-related regions in the right cerebellum functionally connected with the left cerebral language areas (Alvarez and Fiez, 2018; Booth et al., 2007). This is consistent with the anatomical observation that signal projection between the cerebellum and cerebrum follows a closed-loop architecture and is contralateral (Buckner 2013). Similarly, our findings suggest right dominant asymmetry in the cerebellum’s specificity for linguistic processing.
Notably, bilateral cerebellar activation has also been reported in previous studies. However, this effect might be mainly associated with a low baseline. When fixation was used as the baseline, the cerebellar activation related to reading tends to be bilateral (Turkeltaub et al. 2002; D'Mello et al. 2020; Rueckl et al. 2015; Ashburn et al. 2020; Chan et al. 2008; Das et al. 2011; Kumar et al. 2010; Li et al. 2021a), whereas when other visual tasks (e.g., font size decision or line pattern match) were used as the baseline, the cerebellar activation was only reported in the right hemisphere (Tan et al. 2005; Xu et al. 2015; Gao et al. 2015). Consistent with these results, when we compared linguistic conditions with the fixation condition, we observed significant reading-related activity in bilateral lobules VI and VIIIA. However, when we used the non-linguistic stimuli as baselines, only significant activation in the right lobules VI and VIIIA was observed. Even though a lenient threshold was applied, no significant reading-related activity was observed in the left cerebellum. Similarly, we also did not observe any significant activation in the cerebellar nuclei, consistent with our other studies (Li et al. 2020; Feng et al. 2017). This could suggest that the cerebellar nuclei were not related to reading. This could also be due to the spatial location of cerebellar nuclei, which might be too deep to be scanned or the associated signal-to-noise ratios are too small.
This result contributes to clarifying the specific relationship between the right cerebellar lobules and reading. Previous studies have mainly reported the consistent involvement of the right lobule VI/Crus I in reading or reading-related tasks (Stoodley and Stein 2013; Moore et al. 2017; Tan et al. 2005; Martin et al. 2015). However, anatomically defined cerebellar lobules include functional subdivisions, which may vary in their relation to reading. In the current study, we extend our understanding by comparing activation during the reading tasks and non-reading control tasks (as opposed to a lower-level fixation) in each functional subregion of the cerebellum. The results showed that only areas in the posterior parts of right lobule VI showed preferences for linguistic stimuli, whereas subdivisions in the anterior parts of right lobule VI, connected with the cerebral domain general networks were not biased towards any of the five visual categories. We further revealed a novel region of the cerebellum that is specific to linguistic stimuli, i.e., the right lobule VIIIA. Engagement of the right lobule VIIIA has been less frequently reported in literacy research, which could be related to methodological approach. As previous studies mainly focused on the cerebrum, the inferior parts of the cerebellum (e.g., lobules VIIIA, VIIIB, and lobule IX) have often been excluded during scanning, or neglected during data analyses (Ashburn et al. 2020). In the current study, we only included participants whose imaging data covered the whole cerebellum, which contributes to detecting effects in the inferior parts of the cerebellum. Interestingly, we observed that the cerebellar activity during passively viewing house was similar to English words and Chinese characters. On the one hand, this result could indicate that the specificity of the right cerebellar clusters to scripts was limited, which may not be able to differentiate word stimuli from house stimuli. On the other hand, the observed similarity could be due to some shared cognitive processes; more research is needed to explore this issue further.
To disentangle the contribution of these two cerebellar clusters to linguistic processing, we also compared the FC to the cerebral cortex seeded by these two clusters. We found that the right lobule VI cluster had greater FC with the cerebral cortical regions, whereas the right lobule VIIIA cluster had greater FC with the subcortical regions, suggesting functional segregations of these two regions. Future studies are required to further dig into the specific role of these two regions in reading.
Previous studies proposed that the cerebellum’s contribution to reading could be due to motor or motor-related processes, such as articulation (Nicolson et al. 2001). In the current study, to control hand movement or articulatory process, we used a passive viewing task, during which participants were only required to view visual stimuli presented on the screen. Therefore, greater activation during reading in the cerebellum was less likely to be associated with motor-related processes. However, it might be argued that passive viewing of scripts could implicitly induce articulatory processes, resulting in a difference between reading and non-reading tasks. Prior work suggests that the posteromedial part of the right lobule VI may be associated with articulation (Frings et al. 2006; Thürling et al. 2011), whereas the posterolateral part may be responsible for higher-level linguistic processing (e.g., semantic processing; Frings et al. 2006; Thürling et al. 2011; Stoodley et al. 2012). The central coordinates of our right lobule VI are closer to regions responsible for higher-level linguistic processing compared to that responsible for articulation (e.g., compared with Frings et al. 2006). Furthermore, the right lobule VI observed in the current study did not overlap with regions that connected with the cerebral articulatory apparatus, e.g., bilateral insula (R2, Fig. 4 in Li et al. 2020). Instead, our observed activation largely overlapped with the subregion in the posterior lateral part of the right lobule VI connected with the left fusiform gyrus (R4, Fig. 4 in Li et al. 2020), an important reading-related cerebral area. For the right lobule VIIIA, studies also reported greater activation in this region during verb generation tasks compared to the reading-only tasks (Stoodley et al. 2012), or during Chinese semantic tasks (Wu et al. 2012). These studies suggest that both the posterior parts of right lobule VI and the right lobule VIIIA were more likely to be involved in high-level linguistic processing during reading (e.g., semantic processing) rather than articulation.
Language differences associated with the cerebellum
To investigate the possibility of a language effect, we compared the cerebellar activation between passive viewing of Chinese characters and English words. However, we did not observe any regions that could significantly differentiate two scripts via either ROI-based analysis or voxel-based analysis of the whole cerebellum. Bayesian analyses further offered moderate evidence for null significant difference in terms of the cerebellar activation across scripts.
There are a few possible interpretations of this null result. On one hand, the lack of script sensitivity may suggest that while cerebellar lobules VI and VIIIA are specifically responsive to visual linguistic stimuli, they may only be involved in lower-level reading processes. In support of this interpretation, we failed to uncover any significant correlations between children’s cerebellar activity and their reading abilities in either Chinese or English. Notably, in the current sample, children have substantially less experience with English reading. If lobules VI and VIIIA were to be involved in language-general higher-level processes such as semantics or comprehension, we might expect to see a difference between children’s automatic Chinese reading, and their more effortful English decoding. Although current findings demonstrate that these cerebellar regions show sensitivity to linguistic vs. non-linguistic stimuli, it remains possible that this sensitivity is not specific to reading per se.
On the other hand, cerebellar lobules VI and VIIIA may well be engaged in higher-level reading processes that are not language or script specific. This interpretation consistent with results from adult bilingual readers. In reviewing previous studies on adult reading (see, Li et al. 2021b), we found that only 3 out of 33 bilingual studies reported language effects in the left cerebellum, with greater activation during reading in the second language compared to the first language (Park et al. 2012; Buchweitz et al. 2009; Xu et al. 2017). No significant effects were reported in the right cerebellum, which might suggest that activation in the right cerebellar lobules is not sensitive to different languages, independent of age. One possibility is that both the right lobule VI and VIIIA are associated with language-general aspects of reading. For instance, previous studies have reported that languages with different writing systems could share semantic networks (Rueckl et al. 2015; Van de Putte et al. 2017), and the active regions of lobules VI and VIIIA in the current study may be involved in higher-level linguistic processes such as semantics (Frings et al. 2006; Thürling et al. 2011; Stoodley et al. 2012; Wu et al. 2012).
Interestingly, and perhaps in support of this second interpretation, we observed significant language differences in the functional connectivity between the active cerebellar regions and the cerebrum. Specifically, we observed greater FC between the two right cerebellar clusters and the L.IPL in English reading compared to Chinese reading. In contrast, we did not observe any greater FC in Chinese reading compared to English reading. This result is in line with previous studies suggesting a critical role of the left IPL in second-language reading (Li et al. 2021b; Barbeau et al. 2017). The present study suggests that for English visual word processing, the right lobule VI might increase its FC with the cerebral regions that specifically support L2 reading. These results indicate that although the cerebellum itself may not be sensitive to different languages/orthographies, its connectivity with other cerebral regions may be modulated by other linguistic demands.
These results deepen our understanding of how the cerebellum contributes to reading across languages. Specifically, activation in the cerebellum was able to differentiate between linguistic and non-linguistic stimuli, but unable to differentiate between Chinese characters and English words. This result emphasizes the importance of considering cerebro-cerebellar connection in our theories of reading, and opens the door to many future studies that can further illuminate the specific role of the cerebellum across literacy tasks.
The influence of developmental stages
Notably, our results were based on children’s data, which raises the question of whether the reading-related activity in the right cerebellar clusters might be a characteristic of a given developmental stage. However, the involvement of the right lobule VI has been reported in readers of other ages, such as adult readers (Martin et al. 2015) and prereaders (Raschle et al. 2012). Direct comparison of child and adult readers found no significant group differences in cerebellar activity (Booth et al. 2004). In the present study, we found that greater engagement of the right lobule VI and VIII remained significant when age was regressed out. Put together, it is possible that age does not significantly influence the reading specificity of the right lobules VI and VIII. However, relations between brain activity and reading performance might change with age. One study of beginning kindergarten-aged readers suggests that activation in the bilateral lobule VII/VIIIA is significantly correlated with children’s reading abilities (Li et al. 2021a). In the current study with older children, we did not observe any significant correlations between cerebellar activation and reading performance. Thus, the relative involvement of the cerebellum in reading across different developmental stages, and the association between cerebellar activation and reading skills, remain important questions for continued research.
Limitations of the current study
The current study has several limitations related to the use of a passive reading paradigm. On one hand, our task successfully minimized motor or motor-related activities in the cerebellum, and was sufficient to elicit reading-related activity. However, compared to overt reading, implicit reading involves relatively lower cognitive involvement, which may make it more difficult to detect differences related to language or orthography. Future studies with overt reading tasks are needed to clarify the possibility of a language effect in the cerebellum. Second, the current study compared Chinese and English word processing to passive viewing of faces, houses, and tools. Using other visual stimuli as a baseline condition advances prior work, and helps to clarify the issue of reading specificity in the cerebellum. However, our task did not include other linguistic non-word stimuli, such as false fonts or letter strings. As such, we must be cautious in our conclusions about specificity for reading as opposed to specificity for linguistic vs. non-linguistic stimuli more generally. Future work should include conditions that modulate the level of language processing, which could investigate the reading specificity issue more clearly and deeply. Finally, in the present study, we can only indirectly infer the roles of right lobules VI and VIIIA in reading. More detailed contributions of these two regions to reading should be investigated in future studies.
Conclusion
The current study investigated which subregions of the cerebellum might show functionally specific activation for reading, and whether these regions are sensitive to different languages or orthographies. Using an implicit reading paradigm that involved passive viewing of Chinese characters and English words, we observed that the posterior part of right lobule VI and the right lobule VIIIA might be functionally specialized areas for reading. Neural activity in these two regions did not vary for Chinese vs. English implicit reading; however, further examination revealed differences in functional connectivity between these two cerebellar clusters and a cerebellar language-specific area across languages. These results inform our understanding of the specific contribution of the right lobule VI and right lobule VIIIA to reading, and emphasize the importance of cerebro-cerebellar connections in models of reading.
Data and code available statement
All data and code used during the study are available from the corresponding author by request.
References
Alvarez TA, Fiez JA (2018) Current perspectives on the cerebellum and reading development. Neurosci Biobehav Rev 92:55–66. https://doi.org/10.1016/j.neubiorev.2018.05.006
Ang C, Zhang J, Chu M, Li H, Tian M, Feng X, Zhang M, Liu L, Meng X, Ding G (2020) Intrinsic cerebro-cerebellar functional connectivity reveals the function of cerebellum VI in reading-related skills. Front Psychol 11:420. https://doi.org/10.3389/fpsyg.2020.00420
Ashburn SM, Flowers DL, Napoliello EM, Eden GF (2020) Cerebellar function in children with and without dyslexia during single word processing. Hum Brain Mapp 41(1):120–138. https://doi.org/10.1002/hbm.24792
Barbeau EB, Chai XJ, Chen J-K, Soles J, Berken J, Baum S, Watkins KE, Klein D (2017) The role of the left inferior parietal lobule in second language learning: an intensive language training fMRI study. Neuropsychologia 98:169–176. https://doi.org/10.1016/j.neuropsychologia.2016.10.003
Battista C, Evans TM, Ngoon TJ, Chen T, Chen L, Kochalka J, Menon V (2018) Mechanisms of interactive specialization and emergence of functional brain circuits supporting cognitive development in children. Npj Sci Learn 3(1):1. https://doi.org/10.1038/s41539-017-0017-2
Booth JR, Burman DD, Meyer JR, Gitelman DR, Parrish TB, Mesulam MM (2004) Development of brain mechanisms for processing orthographic and phonologic representations. J Cogn Neurosci 16(7):1234–1249. https://doi.org/10.1162/0898929041920496
Booth JR, Wood L, Lu D, Houk JC, Bitan T (2007) The role of the basal ganglia and cerebellum in language processing. Brain Res 1133(1):136–144. https://doi.org/10.1016/j.brainres.2006.11.074
Boros M, Anton J-L, Pech-Georgel C, Grainger J, Szwed M, Ziegler JC (2016) Orthographic processing deficits in developmental dyslexia: beyond the ventral visual stream. Neuroimage 128:316–327. https://doi.org/10.1016/j.neuroimage.2016.01.014
Buchweitz A, Mason RA, Hasegawa M, Just MA (2009) Japanese and English sentence reading comprehension and writing systems: An fMRI study of first and second language effects on brain activation. Biling Lang Cogn 12(2):141–151. https://doi.org/10.1017/S1366728908003970
Buckner RL (2013) The cerebellum and cognitive function: 25 years of insight from anatomy and neuroimaging. Neuron 80(3):807–815. https://doi.org/10.1016/j.neuron.2013.10.044
Cao F, Kim SY, Liu Y, Liu L (2014) Similarities and differences in brain activation and functional connectivity in first and second language reading: evidence from Chinese learners of English. Neuropsychologia 63:275–284. https://doi.org/10.1016/j.neuropsychologia.2014.09.001
Cattinelli I, Borghese NA, Gallucci M, Paulesu E (2013) Reading the reading brain: a new meta-analysis of functional imaging data on reading. J Neurolinguist 26(1):214–238. https://doi.org/10.1016/j.jneuroling.2012.08.001
Chan AH, Luke KK, Li P, Yip V, Li G, Weekes B, Tan LH (2008) Neural correlates of nouns and verbs in early bilinguals. Ann N Y Acad Sci 1145(1):30–40
Cherodath S, Singh NC (2015) The influence of orthographic depth on reading networks in simultaneous biliterate children. Brain Lang 143:42–51. https://doi.org/10.1016/j.bandl.2015.02.001
Cisler JM, Bush K, Steele JSJN (2014) A comparison of statistical methods for detecting context-modulated functional connectivity in fMRI. Neuroimage 84:1042–1052. https://doi.org/10.1016/j.neuroimage.2013.09.018
Das T, Bapi RS, Padakannaya P, Singh NC (2011) Cortical network for reading linear words in an alphasyllabary. Read Writ 24(6):697–707. https://doi.org/10.1007/s11145-010-9241-3
Dehaene S, Pegado F, Braga LW, Ventura P, Nunes Filho G, Jobert A, Dehaene-Lambertz G, Kolinsky R, Morais J, Cohen L (2010) How learning to read changes the cortical networks for vision and language. Science 330(6009):1359–1364. https://doi.org/10.1126/science.1194140
Dehaene-Lambertz G, Monzalvo K, Dehaene S (2018) The emergence of the visual word form: longitudinal evolution of category-specific ventral visual areas during reading acquisition. PLoS Biol 16(3):e2004103. https://doi.org/10.1371/journal.pbio.2004103
Diedrichsen J (2006) A spatially unbiased atlas template of the human cerebellum. Neuroimage 33(1):127–138. https://doi.org/10.1016/j.neuroimage.2006.05.056
Diedrichsen J, Balsters JH, Flavell J, Cussans E, Ramnani N (2009) A probabilistic MR atlas of the human cerebellum. Neuroimage 46(1):39–46. https://doi.org/10.1016/j.neuroimage.2009.01.045
D’Mello AM, Turkeltaub PE, Stoodley CJ (2017) Cerebellar tDCS modulates neural circuits during semantic prediction: a combined tDCS-fMRI study. J Neurosci 37(6):1604–1613. https://doi.org/10.1523/JNEUROSCI.2818-16.2017
D’Mello AM, Centanni TM, Gabrieli JD, Christodoulou JA (2020) Cerebellar contributions to rapid semantic processing in reading. Brain Lang 208:104828. https://doi.org/10.1016/j.bandl.2020.104828
Eckert MA, Leonard CM, Richards TL, Aylward EH, Thomson J, Berninger VW (2003) Anatomical correlates of dyslexia: frontal and cerebellar findings. Brain 126(2):482–494. https://doi.org/10.1093/brain/awg026
Eckert MA, Berninger VW, Vaden KI, Gebregziabher M, Tsu L, Consortium DD (2016) Gray matter features of reading disability: a combined meta-analytic and direct analysis approach. Eneuro. https://doi.org/10.1523/ENEURO.0103-15.2015
Feng X, Li L, Zhang ML, Yang XJ, Tian MY, Xie WY, Lu Y, Liu L, Belanger NN, Meng XZ, Ding GS (2017) Dyslexic children show atypical cerebellar activation and cerebro-cerebellar functional connectivity in orthographic and phonological processing. Cerebellum 16(2):496–507. https://doi.org/10.1007/s12311-016-0829-2
Feng X, Altarelli I, Monzalvo K, Ding G, Ramus F, Shu H, Dehaene S, Meng X, Dehaene-Lambertz G (2019) Shared anomalies in cortical reading networks in Chinese and French dyslexic children. bioRxiv 834945
Frings M, Dimitrova A, Schorn CF, Elles H-G, Hein-Kropp C, Gizewski ER, Diener HC, Timmann D (2006) Cerebellar involvement in verb generation: an fMRI study. Neurosci Lett 409(1):19–23. https://doi.org/10.1016/j.neulet.2006.08.058
Gao Q, Wang J, Yu C, Chen H (2015) Effect of handedness on brain activity patterns and effective connectivity network during the semantic task of Chinese characters. Sci Rep 5(1):1–11. https://doi.org/10.1038/srep18262
Gatti D, van Vugt F, Vecchi T (2020) A causal role for the cerebellum in semantic integration: a transcranial magnetic stimulation study. Sci Rep 10(1):1–12. https://doi.org/10.1038/s41598-020-75287-z
Hancock R, Richlan F, Hoeft F (2017) Possible roles for fronto-striatal circuits in reading disorder. Neurosci Biobehav Rev 72:243–260. https://doi.org/10.1016/j.neubiorev.2016.10.025
Ip KI, Marks RA, Hsu LS-J, Desai N, Kuan JL, Tardif TJB, Kovelman I (2019) Morphological processing in Chinese engages left temporal regions. Brain Lang 199:104696. https://doi.org/10.1016/j.bandl.2019.104696
Karipidis I, Pleisch G, Röthlisberger M, Hofstetter C, Dornbierer D, Stämpfli P, Brem S (2017) Neural initialization of audiovisual integration in prereaders at varying risk for developmental dyslexia. Hum Brain Mapp 38(2):1038–1055. https://doi.org/10.1002/hbm.23437 (Epub 2016 Oct 14)
Karipidis II, Pleisch G, Brandeis D, Roth A, Röthlisberger M, Schneebeli M, Walitza S, Brem S (2018) Simulating reading acquisition: the link between reading outcome and multimodal brain signatures of letter–speech sound learning in prereaders. Sci Rep 8(1):7121. https://doi.org/10.1038/s41598-018-24909-8
Kibby MY, Fancher JB, Markanen R, Hynd GW (2008) A quantitative magnetic resonance imaging analysis of the cerebellar deficit hypothesis of dyslexia. J Child Neurol 23(4):368–380. https://doi.org/10.1177/0883073807309235
Kumar U, Das T, Bapi RS, Padakannaya P, Joshi RM, Singh NC (2010) Reading different orthographies: an fMRI study of phrase reading in Hindi–English bilinguals. Read Writ 23(2):239–255. https://doi.org/10.1007/s11145-009-9176-8
Langer N, Benjamin C, Minas J, Gaab N (2015) The neural correlates of reading fluency deficits in children. Cereb Cortex 25(6):1441–1453. https://doi.org/10.1093/cercor/bht330
Leppink J, Osullivan P, Winston K, Pome J (2017) Evidence against vs in favour of a null hypothesis. Perspect Med Educ 6(2):115–118
Li H, Booth JR, Bélanger NN, Feng X, Tian M, Xie W, Zhang M, Gao Y, Ang C, Yang X, Liu L, Meng X, Ding G (2018) Structural correlates of literacy difficulties in the second language: evidence from Mandarin-speaking children learning English. Neuroimage 179:288–297. https://doi.org/10.1016/j.neuroimage.2018.06.037
Li H, Booth JR, Feng X, Wei N, Zhang M, Zhang J, Zhong H, Lu C, Liu L, Ding G, Meng X (2020) Functional parcellation of the right cerebellar lobule VI in children with normal or impaired reading. Neuropsychologia 148:107630. https://doi.org/10.1016/j.neuropsychologia.2020.107630
Li H, Kepinska O, Caballero JN, Zekelman L, Marks RA, Uchikoshi Y, Kovelman I, Hoeft F (2021a) Decoding the role of the cerebellum in the early stages of reading acquisition. Cortex 141:262–279. https://doi.org/10.1016/j.cortex.2021.02.033
Li H, Zhang J, Ding G (2021b) Reading across writing systems: a meta-analysis of the neural correlates for first and second language reading. Biling Lang Cogn 24(3):1–12. https://doi.org/10.1017/S136672892000070X
Liu L, Li H, Zhang M, Wang Z, Wei N, Liu L, Meng X, Ding G (2016) Aberrant topologies and reconfiguration pattern of functional brain network in children with second language reading impairment. Dev Sci 19(4):657–672. https://doi.org/10.1111/desc.12440
Mariën P, Ackermann H, Adamaszek M, Barwood CH, Beaton A, Desmond J, De Witte E, Fawcett AJ, Hertrich I, Küper M (2014) Consensus paper: language and the cerebellum: an ongoing enigma. The Cerebellum 13(3):386–410. https://doi.org/10.1007/s12311-013-0540-5
Martin A, Schurz M, Kronbichler M, Richlan F (2015) Reading in the brain of children and adults: a meta-analysis of 40 functional magnetic resonance imaging studies. Hum Brain Mapp 36(5):1963–1981. https://doi.org/10.1002/hbm.22749
Mazaika PK, Hoeft F, Glover GH, Reiss AL (2009) Methods and software for fMRI analysis of clinical subjects. Neuroimage 47(Suppl 1):S58. https://doi.org/10.1016/S1053-8119(09)70238-1
McLaren DG, Ries ML, Xu G, Johnson SC (2012) A generalized form of context-dependent psychophysiological interactions (gPPI): a comparison to standard approaches. NeuroImage 61(4):1277–1286
Michels L, O’Gorman R, Kucian KJD (2018) Functional hyperconnectivity vanishes in children with developmental dyscalculia after numerical intervention. Dev Cogn Neurosci 30:291–303. https://doi.org/10.1016/j.dcn.2017.03.005
Monzalvo K, Fluss J, Billard C, Dehaene S, Dehaene-Lambertz G (2012) Cortical networks for vision and language in dyslexic and normal children of variable socio-economic status. Neuroimage 61(1):258–274. https://doi.org/10.1016/j.neuroimage.2012.02.035
Moore DM, D’Mello AM, McGrath LM, Stoodley CJJD (2017) The developmental relationship between specific cognitive domains and grey matter in the cerebellum. Dev Cogn Neurosci 24:1–11. https://doi.org/10.1016/j.dcn.2016.12.001
Nicolson RI, Fawcett AJ, Dean P (2001) Developmental dyslexia: the cerebellar deficit hypothesis. Trends Neurosci 24(9):508–511. https://doi.org/10.1016/S0166-2236(00)01896-8
Oldfield (1971) The assessment and analysis of handedness: the Edinburgh inventory. Neuropsychologia 9:97–113. https://doi.org/10.1016/0028-3932(71)90067-4
Park HR, Badzakova-Trajkov G, Waldie KE (2012) Language lateralisation in late proficient bilinguals: a lexical decision fMRI study. Neuropsychologia 50(5):688–695. https://doi.org/10.1016/j.neuropsychologia.2012.01.005
Paz-Alonso PM, Oliver M, Lerma-Usabiaga G, Caballero-Gaudes C, Quiñones I, Suárez-Coalla P, Duñabeitia JA, Cuetos F, Carreiras M (2018) Neural correlates of phonological, orthographic and semantic reading processing in dyslexia. NeuroImage Clin 20:433–447. https://doi.org/10.1016/j.nicl.2018.08.018
Quintana DS, Williams DRJB (2018) Bayesian alternatives for common null-hypothesis significance tests in psychiatry: a non-technical guide using JASP. BMC Psychiatry 18(1):1–8. https://doi.org/10.1186/s12888-018-1761-4
Rae C, Harasty JA, Dzendrowskyj TE, Talcott JB, Simpson JM, Blamire AM, Dixon RM, Lee MA, Thompson CH, Styles P (2002) Cerebellar morphology in developmental dyslexia. Neuropsychologia 40(8):1285–1292. https://doi.org/10.1016/S0028-3932(01)00216-0
Raschle NM, Zuk J, Gaab N (2012) Functional characteristics of developmental dyslexia in left-hemispheric posterior brain regions predate reading onset. Proc Natl Acad Sci 109(6):2156–2161. https://doi.org/10.1073/pnas.1107721109
Raven JC, Court JH (1998) Raven’s progressive matrices and vocabulary scales. Oxford Psychologists Press, Oxford, UK
Riva D, Giorgi C (2000) The cerebellum contributes to higher functions during development. Brain A J Neurol 5:5. https://doi.org/10.1093/brain/123.5.1051
Rueckl JG, Paz-Alonso PM, Molfese PJ, Kuo W-J, Bick A, Frost SJ, Hancock R, Wu DH, Mencl WE, Duñabeitia JA (2015) Universal brain signature of proficient reading: Evidence from four contrasting languages. Proc Natl Acad Sci 112(50):15510–15515. https://doi.org/10.1073/pnas.1509321112
Schmahmann JD (1996) From movement to thought: anatomic substrates of the cerebellar contribution to cognitive processing. Hum Brain Mapp 4(3):174–198. https://doi.org/10.1002/(SICI)1097-0193(1996)4:3%3c174::AID-HBM3%3e3.0.CO;2-0
Scott RB, Stoodley CJ, Anslow P, Paul C, Stein JF, Sugden EM, Mitchell CD (2001) Lateralized cognitive deficits in children following cerebellar lesions. Dev Med Child Neurol 43(10):685–691
Sokolov Arseny A, Miall RC, Ivry Richard B (2017) The cerebellum: adaptive prediction for movement and cognition. Trends Cogn Sci 21(5):313–332. https://doi.org/10.1016/j.tics.2017.02.005
Stoodley CJ (2014) Distinct regions of the cerebellum show gray matter decreases in autism, ADHD, and developmental dyslexia. Front Syst Neurosci. https://doi.org/10.3389/fnsys.2014.00092
Stoodley CJ, Schmahmann JD (2009) Functional topography in the human cerebellum: a meta-analysis of neuroimaging studies. Neuroimage 44(2):489–501. https://doi.org/10.1016/j.neuroimage.2008.08.039
Stoodley CJ, Stein JF (2013) Cerebellar function in developmental dyslexia. Cerebellum 12(2):267–276. https://doi.org/10.1007/s12311-012-0407-1
Stoodley CJ, Valera EM, Schmahmann JD (2012) Functional topography of the cerebellum for motor and cognitive tasks: an fMRI study. Neuroimage 59(2):1560–1570. https://doi.org/10.1016/j.neuroimage.2011.08.065
Tan LH, Laird AR, Li K, Fox PT (2005) Neuroanatomical correlates of phonological processing of Chinese characters and alphabetic words: a meta-analysis. Hum Brain Mapp 25(1):83–91. https://doi.org/10.1002/hbm.20134
Thürling M, Küper M, Stefanescu R, Maderwald S, Gizewski E, Ladd ME, Timmann DJN (2011) Activation of the dentate nucleus in a verb generation task: a 7 T MRI study. J NeuroIamage 57(3):1184–1191. https://doi.org/10.1016/j.neuroimage.2011.05.045
Turkeltaub PE, Eden GF, Jones KM, Zeffiro TA (2002) Meta-analysis of the functional neuroanatomy of single-word reading: method and validation. Neuroimage 16(3):765–780. https://doi.org/10.1006/nimg.2002.1131
Van de Putte E, De Baene W, Brass M, Duyck W (2017) Neural overlap of L1 and L2 semantic representations in speech: a decoding approach. Neuroimage 162:106–116. https://doi.org/10.1016/j.neuroimage.2017.08.082
Wang X, Tao B (1993) Chinese character recognition test battery and assessment scale for primary school children. Shanghai Education Press, Shanghai
Weiss Y, Cweigenberg HG, Booth JR (2018) Neural specialization of phonological and semantic processing in young children. Hum Brain Mapp. https://doi.org/10.1002/hbm.24274
Woodcock R (1987) Manual for the woodcock reading mastery test-revised. American Guidance Service, Circle Pines
Wu C-Y, Ho M-HR, Chen S-HA (2012) A meta-analysis of fMRI studies on Chinese orthographic, phonological, and semantic processing. Neuroimage 63(1):381–391. https://doi.org/10.1016/j.neuroimage.2012.06.047
Xia Z, Hoeft F, Zhang L, Shu H (2016) Neuroanatomical anomalies of dyslexia: disambiguating the effects of disorder, performance, and maturation. Neuropsychologia 81:68–78. https://doi.org/10.1016/j.neuropsychologia.2015.12.003
Xu M, Wang T, Chen S, Fox PT, Tan LH (2015) Effective connectivity of brain regions related to visual word recognition: an fMRI study of Chinese reading. Hum Brain Mapp 36(7):2580–2591. https://doi.org/10.1002/hbm.22792
Xu M, Baldauf D, Chang CQ, Desimone R, Tan LH (2017) Distinct distributed patterns of neural activity are associated with two languages in the bilingual brain. Sci Adv 3(7):e1603309. https://doi.org/10.1126/sciadv.1603309
You H, Gaab N, Wei N, Cheng-Lai A, Wang Z, Jian J, Song M, Meng X, Ding G (2011) Neural deficits in second language reading: fMRI evidence from Chinese children with English reading impairment. Neuroimage 57(3):760–770. https://doi.org/10.1016/j.neuroimage.2010.12.003
Younger JW, Tucker-Drob E, Booth JR (2017) Longitudinal changes in reading network connectivity related to skill improvement. Neuroimage 158:90–98. https://doi.org/10.1016/j.neuroimage.2017.06.044
Zhang H, Wang X (1985) Raven’s IQ reasoning standardized test. Beijing Normal University Press, Beijing
Zhang Y, Shao H, Gaspar CM, Chen W, Zhu J, Tang Y, He S, Weng XJCSB (2015) Face information processing in the left inferior temporal lobe: an ECoG case study. Chin Sci Bull 60(25):2438–2446. https://doi.org/10.1360/N972015-00366
Acknowledgements
This work was supported by Grants from the National Natural Science Foundation of China (NSFC: 31971039, 31971036, 32100846) and Shenzhen-Hong Kong institute of Brain Science-Shenzhen Fundamental Research Institutions (2021SHIBS0003). It was also supported by Project funded by China Postdoctoral Science Foundation (Grant No. 2020M682846) and the Open Research Fund of the State Key Laboratory of Cognitive Neuroscience and Learning (Grant No. CNLYB2005).
Author information
Authors and Affiliations
Contributions
HL: conceptualization, methodology, investigation, formal analysis, writing—original draft, funding acquisition. RAM: writing—original draft, writing—review and editing. LL: writing—original draft, writing—review and editing. XF: investigation, writing—review and editing. MZ: investigation, writing—review and editing. FA: investigation, writing—review and editing. YG: investigation, writing—review and editing. MT: investigation, writing—review and editing. XY: investigation, writing—review and editing. JZ: writing—review and editing. HZ: writing—review and editing. LL: investigation, writing—review and editing, project administration. XM: conceptualization, investigation, methodology, writing—review and editing, project administration, supervision, funding acquisition. GD: conceptualization, investigation, methodology, writing—original draft, writing—review and editing, project administration, supervision, funding acquisition.
Corresponding authors
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare.
Ethics approval
The use of human subjects for this study has been approved by the Ethical Review Board of Beijing Normal University.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Li, H., Marks, R.A., Liu, L. et al. The selective contributions of right cerebellar lobules to reading. Brain Struct Funct 227, 963–977 (2022). https://doi.org/10.1007/s00429-021-02434-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00429-021-02434-1