
Abstract
This study differentiated the contributions of physical and observational practice to the learning of a single-limb multi-joint coordination pattern. Three groups (physical-practice, observation-practice, observation-physical) practiced for 2 days and were given two performance tests 24 h after the second practice session. The performance tests revealed that physical and observational practice contributed similarly to identifying and using kinematic information related to the relative motion direction between joints (lead/lag relationship) and to the to-be-learned relative phase pattern (ϕ = 90°). Physical practice resulted in more stable coordination during performance tests and in the ability to produce different joint amplitudes with less variability. A serendipitous finding was that maximum elbow flexion (point of movement reversal) emerged as a kinematic event around which elbow and wrist coordination were organized. Movement reversals often serve to anchor the movement dynamics, and this anchoring effect was evident following both physical and observational practice, yet physical practice resulted in an advantage with regard to this anchor point on several kinematic measures. The results are discussed within the context of contemporary behavioral theories (coordination dynamics, visual perception perspective) of observational learning.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Observing the actions of others serves many purposes. Regardless of whether the purpose is socially based or sports based, a key feature of observation is to facilitate the development of the appropriate motor responses that meet the demands of a given context. In sports contexts, executing efficient and precise movements is paramount to success. In rehabilitation contexts, finding novel ways to move the limbs and joints to achieve every day task goals is paramount to success. The use of demonstration and observation as training protocols is a key aspect of sport and dance training, and shows relevance in rehabilitation contexts (Celnik, Webster, Glasser, & Cohen, 2008; Franceschini et al., 2012). A key research issue is then identifying what movement information is extracted or picked-up through observation of another’s actions, and identifying how that information is used by the observer in generating the demonstrated action. Many studies have shown that observation or demonstration facilitates motor skill learning of sequential actions, discrete actions, and rhythmic actions. However, the extent that observation contributes differently than physical practice to the pick-up and use of specific kinematic information about an action has not been examined extensively (Hamilton & Grafton, 2007). The current study was designed to reveal the different contributions of observation and physical practice to the extraction and use of kinematic information quantified by the relative motion direction between joints (lead/lag relationship), the specific relative phase between joints, and the specific joint amplitudes.
The visual perception theory was developed to provide a framework to understand how observation facilitates the control and coordination of limbs in motor learning contexts and drew heavily on Gibson’s theory of direct perception (Gibson, 1979) and research on point-light displays (Cutting & Proffitt, 1982; Johansson, 1973). From the visual perception perspective, observation provides for the identification of topological invariants (kinematic information) that represent the global movement strategies or coordination patterns a model is producing, such as the lead/lag relationship between limbs and the sequence of limb motions (Al-Abood, Davids, Bennett, Ashford, & Marin, 2001; Buchanan & Dean, 2010; Buchanan, Ryu, Zihlman, & Wright, 2008; Scully, 1986). Physical practice then allows for the scaling of limb and joint motions to fine tune the coordination pattern through more precise control (Newell, 1991; Scully & Carnegie, 1998).
Research has clearly shown that demonstration facilitates the identification of specific movement strategies in observers. For example, observers evaluate a model’s movement strategies with respect to goal attainment (Martens, Burwitz, & Zuckerman, 1976), and observers can mimic the kinematics (velocity profiles) of a model’s strategy when required (Al-Abood, Davids, & Bennett, 2001; Al-Abood et al., 2001). Observers without explicit instructions to do so will mimic the movement strategy employed by an expert model or novice learning model if the strategy is repeatedly used and achieves the goal (Buchanan & Dean, 2014; Buchanan, Ryu, Zihlman, & Wright, 2008). Thus, observers through visual analysis can evaluate goal attainment probabilities, consistency in modeled performance (expert), and performance improvements (learning model). Thus, the linking of a movement strategy to an action goal through observation requires the estimation of control processes that will produce the coordinated kinematic patterns of the strategy without overt physical practice.
A variety of studies using full-body actions have examined the accuracy with which observers can reproduce a model’s kinematic patterns (Al-Abood, Davids, & Bennett, 2001; Al-Abood et al., 2001; Hayes, Hodges, Huys, & Williams, 2007; Hodges, Williams, Hayes, & Breslin, 2007). Observing a model results in better approximation of the model’s relative motion pattern between the upper-arm and forearm in a throwing task compared to receiving verbal instructions (Al-Abood, Davids, & Bennett, 2001). Comparisons between a model’s joint kinematics (hip-knee and knee-ankle coordination) and a group of observers’ joint kinematics have also been examined in a soccer kicking task (Hodges, Hayes, Breslin, & Williams, 2005) and a cricket bowling task (Breslin, Hodges, Williams, Kremer, & Curran, 2006; Hayes et al., 2007). The results revealed a change in the kinematic pattern of the observers in the direction of the model and show that both relative motion direction and end-point trajectory information extracted from the model’s action can benefit an observer’s control of their kinematic patterns. Breslin et al. (2006) concluded that observational learning may be more of an emergent feature rather than a direct approximation of the modeled kinematic patterns in these types of tasks. This statement implies that while the model is important, the observer must organize and develop coordination strategies based on their experience and knowledge. In the above reviewed studies, the observers practiced at least as much as they observed and this leads to a confound in isolating the impact of physical practice and observation on the development of the control processes that make coordination more precise and accurate. Even when the amount of physical practice is controlled in observational studies, emphasis is most often placed on identifying the contribution of observation to the action outcome, and not specifically to the kinematics of the limbs which provide the means to the outcome (Andrieux & Proteau, 2014; Badets & Blandin, 2010; Bird, Osman, Saggerson, & Heyes, 2005; Osman, Bird, & Heyes, 2005).
Theoretically, research shows that observational practice supports the development of visual–spatial codes whereas physical practice supports the development of codes based on motor coordinates (Gruetzmacher, Panzer, Blandin, & Shea, 2011). A study that manipulated feedback schedules (KR frequency and KR bandwidth) came to the conclusion that observation benefits the development of motor planning processes more than motor execution processes (Badets & Blandin, 2010). The visual perception theory is in agreement with the above conclusions by predicting that observation supports the extraction of relative motion features (lead/lag relationships, topological invariants or spatial codes). Extensive research using point-light displays has revealed that the visual system rapidly detects relative motion patterns representing full-body actions (Johansson, 1973) and is sensitive to perturbations of the relative motion between limbs (Jacobs & Shiffrar, 2005; Pinto & Shiffrar, 1999; Shiffrar, Lichtey, & Chatterjee, 1997). Studies have also shown that the relative motion between limbs is extracted by observers and used in both action–imitation contexts and observational learning contexts to organize novel coordination patterns in rhythmic arm tasks (Buchanan, 2015; Buchanan, Ramos, & Robson, 2015; Buchanan et al., 2008). Intimately linked to the relative motion between components is the relative phase (ϕ) between components. As stated previously, relative motion direction characterizes the general lead or lag between limbs or joints during a rhythmic motion. The measure relative phase describes the specific lead or lag based on a mean relative phase (e.g., a lead of ϕ = + 30° or a lag of ϕ = −60°) while also providing an estimate of the stability of the specific lead or lag based on the variability in the mean relative phase. With regard to the link between perception and action, research has shown that visual perception processes and coordination processes are constrained by both relative motion and relative phase information (Bingham, Schmidt, & Zaal, 1999; Bingham, Zaal, Shull, & Collins, 2001; Buchanan, 2015; Kelso & Pandya, 1991; Wilson, Collins, & Bingham, 2005; Zaal, Bingham, & Schmidt, 2000).
The current study utilized an animation training protocol to differentiate contributions of physical and observational practice to the learning of a single-limb multi-joint action. The animated arm had three segments, an upper-arm, forearm, and hand (Fig. 1a). The animation served as the model for all groups. The animated behavior was rhythmic flexion–extdoes not facilitate the ability toension motions of the forearm and hand segments in the sagittal motion plane with each segment characterized by different amplitudes. The multi-jointed animation portrayed the elbow leading the wrist throughout every motion cycle (general relative motion organization) at a specific relative phase pattern of ϕ = + 90°. The relative motion direction and relative phase pattern may be viewed as the most global level of the action goal in this task. The amplitude values are not the most global level in that many different joint amplitudes can be used to achieve a specific relative phase pattern (Buchanan & Wright, 2011). To differentiate distinct contributions, if any, of physical and observational practice in this motor learning task three groups were trained: (1) a physical-practice group viewed the animation and physically attempted to produce the animated pattern displayed on every practice trial over 2 days; (2) an observation-practice group viewed as many animated trials as the physical group, yet they were not allowed to attempt the observed pattern until a series of day 3 performance tests; and (3) an observation-physical group viewed the animation as many times as the other two groups, and was allowed to physically practice the pattern on 1/5 of the observation trials. A key feature of the current task is that all groups are trained with the same model (a model without variability), yet participate in different training contexts. Previous work has shown that when physical and observation practice are combined; the observer may gain an advantage through the identification of strategies performed by the model, a so-called social facilitation effect (Shea, Wright, Wulf, & Whitacre, 2000; Buchanan & Dean, 2010). The animated arm will eliminate the social facilitation effect and strategy identification possibility. This will allow for a more direct comparison of the impact of physical practice versus observation on the extraction of kinematic information.

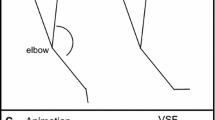
a The stick figure animation is shown on the left and a representation of the virtual stick figure (VSF) of the participant’s arm is shown on the right. The VSF was used to provide concurrent and terminal feedback. Five IREDS were attached to the arm and four (gray circles) were used to construct the VSF. b Experimental setup. c This illustration represents computation of the harmonicity (H) measure. The time series represent normalized joint angular displacement (dashed line) and normalized angular acceleration (solid line). The value of H for each panel is as follows: left panel the reversals are flexion, extension, flexion, and each reversal has a different value of H; middle panel H = 1 for all reversals; and right panel H = 0 for all reversals
Previous research has examined the combined impact of physical and observational practice on motor skill learning (Ellenbuerger, Boutin, Blandin, Shea, & Panzer, 2012; Shea et al., 2000). In the above two studies, the observers watched a physical participant practice and physically practiced the task themselves, receiving equal amounts of physical-practice and observational-practice trials. Shea et al.’s (2000) study was designed to determine if combined practice created an advantage on retention or transfer tests, whereas Ellenbuerger et al.’s (2012) study was designed to see if the order of practice, observation following physical or preceding physical, produced an advantage. The goals of these studies probably required that equal amounts of physical and observational practice be given. In the current task, the goal was not to determine if a certain combination of observation and physical practice is better than either practice type alone. The goal was to determine what the different contributions of the two practice contexts are to extracting and utilizing the same visually based movement information. Therefore, the observation-physical group received less physical practice trials than the physical-practice group, yet received the same number of observations of the animation as the observer-practice group. This creates a bias towards observation for the observation-physical group. Thus, this group should be more similar to the observation-practice group on those kinematic variables that benefit the most from observation while limiting the impact on those variables that benefit the most from physical practice.
To determine the different contributions of physical and observation practice, two separate performance tests were used after the training sessions. The first performance test allowed the participant to move with the animation, yet provided no augmented feedback. This test mimicked part of the training procedures of the physical-practice group. The second performance test had the participants watch the animation and then move once the animation finished. This test mimicked part of the training procedure of the physical-practice group and the observation-physical group. The performance tests provide the physical-practice and observation-physical groups an advantage because they are consistent with parts of their training contexts. Thus, any differences between the groups may be linked back to the amount of physical practice, since all three groups were trained with the same visual stimulus.
Based on the visual perception perspective of observational learning, three primary predictions were developed. First, it was predicted that all three groups would consistently produce a relative motion relationship with the elbow leading the wrist throughout a cycle in both performance tests. This is consistent with the idea that topological invariants are available for pick-up through visual processes and the conclusions that observation facilitates the development of spatial codes and planning processes (Badets & Blandin, 2010; Gruetzmacher et al., 2011). Second, with regard to the relative phase goal, it was predicted that the physical-practice group’s performance would be more accurate and stable than both observation groups for both performance tests with no difference between the observation groups. Three relative phase measures were examined, a continuous measure and two point estimates. Research has shown that synchronizing reversal points (e.g., max pronation or supination) with a metronome signal can stabilize in-phase and anti-phase coordination patterns (Byblow, Carson, & Goodman, 1994; Fink, Foo, Jirsa, & Kelso, 2000). Moreover, without instructions, participants in a bimanual finger task used a reversal point (max flexion or extension) to help them learn a 90° relative phase pattern (Zanone & Kelso, 1997). An interesting finding in the bimanual task of Zanone and Kelso (1997) was that performance was closer to the target relative phase around the selected reversal point than anywhere else in the cycle. The point estimate measures of relative phase were used to determine if the three training contexts impacted the use of reversal points differently. It was predicted that the physical-practice group would have an advantage in utilizing reversal points compared to the observation groups, with the observation-practice group not benefiting from the limited physical practice. However, if an advantage exists for the observation-physical group it would emerge in the second performance test since this test was similar to their physical training.
Third, it was predicted that the physical-practice group would be able to scale elbow and wrist amplitudes as required and produce smoother reversals in joint motion suggesting more harmonic motion of the elbow and wrist in both performance tests. This prediction is consistent with the visual perception thoery that states that observation as a practice context does not facilitate the ability to scale absolute motion features such as required joint amplitudes or absolute movement time. Here again, the observation-physical group is important because if they are more similar to the observation-practice group, this would show the importance of more physical practice in scaling certain kinematic variables, an issue that has not been examined in the studies that have compared the impact of physical and observational practice on motor skill learning.
Methods
Participants
A total of 24 individuals (6 males, 18 females, mean age = 21.8) were recruited from Texas A&M University. The experiment and consent form were approved by the Texas A&M University IRB committee in accordance with the Helsinki Declaration. Each participant provided written consent and received class credit for participation. Participants were assigned to one of the three groups: (1) physical-practice, (2) observation-practice, and (3) observation-physical practice.
Task and training procedures
The task was to learn to produce an elbow–wrist rhythmic coordination pattern with the right-arm that was modeled by a stick figure animation of the human arm presented on a computer monitor (Fig. 1a). The animation produced 16 cycles of motion and only the forearm and hand segments moved. The frequency of motion was .4 Hz. The task was defined by three goals: (1) a relative motion goal—the elbow leads the wrist; (2) a relative phase (ϕ) goal of ϕ = 90° between the elbow and wrist, and (3) an amplitude goal—wrist amplitude = 62°, elbow amplitude = 92°. Participants were instructed to match their forearm and hand motions to the animation and they were not told of the numerical values of goals 2 and 3.
There were two consecutive practice days and two performance tests were administered in a 3rd session 24 h after the 2nd practice day. In the first session, all participants performed nine pre-practice familiarization trials (3 per pattern) defined by the animation, in-phase ϕ = 0°, anti-phase ϕ = 180°, and ϕ = 90°. In-phase required flexing-extending the elbow and wrist simultaneously and anti-phase required extending-flexing the elbow as the wrist flexed–extended. The 90° pattern required elbow maximum flexion/extension to lead wrist maximum flexion/extension by one quarter of a cycle. Participants moved with the animation; however, they did not receive any augmented feedback of their performance in these trials.
The training for all groups consisted of two 40 min sessions (separated by 24 h.) with 30 trials of the to-be-learned 90° relative phase pattern per session. Each session consisted of three blocks with ten trials per block. Training and feedback conditions were different for each group. For the physical-practice group, training and feedback within a block were as follows: Concurrent feedback was presented with trials 1, 2, 5 and 6. For these trials, a virtual stick figure (VSF) representing the participant’s arm was constructed online using infra-red LEDs attached to the participant (Fig. 1a). The VSF was presented next to the animated arm. During trials 3, 4, 7 and 8 the physical-practice participants moved with the animation and the VSF was not presented. During trials 9 and 10, participants watched the animation, and after a 5 s delay attempted to reproduce the animated motion. The observation-physical group watched the animation for trials 1 to 8, and performed trials 9 and 10 in the same manner as the physical-practice group. The observation-practice group watched the stick figure for all 30 trials. It was emphasized to all three groups that they would be tested on their ability to generate the pattern of limb motion produced by the animation in session 3, 24 h after the second practice session.
In session three, each performance test consisted of five trials. In the first test, participants performed by moving with the animation. No feedback was provided during this test and this test is referred to as the performance-animation test. This test was consistent with the training of the physical group on trials 3, 4, 7, and 8 of each block. In the second test, participants watched the 16 cycles of animated motion. When the animation finished and after a 5 s delay the participants tried to produce the demonstrated pattern. No feedback was provided during this test and this test is referred to as the performance-retention test. This test was consistent with the training of the physical-practice and observation-physical groups on trials 9 and 10 of each block.
Data collection and analysis
An OPTOTRAK Certus camera (Northern Digital Ontario, Canada) recorded the xyz position of five infra-red light emitting diodes (IREDs) mounted on the arm (Fig. 1a). The camera has three pre-calibrated lenses (resolution is .1 mm in x and y and .15 mm in z). The camera was mounted horizontally 76.2 cm above the ground and the participant sat 2.5 m from the camera’s center lens (Fig. 1b). The xyz trajectories from each IRED were filtered (dual pass Butterworth, 10 Hz) before computing wrist and elbow angular time series. The wrist angle was derived from IREDs 1, 2, and 3. IRED 1 was attached to a dowel grasped in a participant’s hand, IRED 2 was positioned over the radius styloid process with the forearm supine, and IRED 3 (not shown) was attached to the forearm proximal to IRED 2. The elbow angle was derived from IREDs 3, 4, and 5, with IRED 4 attached to the lateral side of elbow joint and IRED 5 placed on the shoulder’s acromion process. All dependent measures were computed using MATLAB (MathWorks, Inc.)
Coordination measures
Coordination was evaluated by deriving a continuous relative phase measure (ϕ C) and two point estimate measures, one for flexion (ϕ F) and one for extension (ϕ E). All relative phase measures were derived from the elbow and wrist joint angle time series. The continuous relative phase was computed by determining the individual phase angles (θ) for the elbow (θ e) and wrist (θ w) (Scholz & Kelso, 1989). From the individual phase angles, a continuous relative phase was calculated as ϕ C = θ e − θ w. The continuous relative phase values for each trial were subjected to a circular transformation. The circular transformation returned a mean resultant vector representing the average continuous relative phase over a trial (ϕ CMn) and a standard deviation (ϕ CSD) which represents elbow–wrist coordination stability.
Point estimates of relative phase were computed from the elbow and wrist angular time series and reflect coordination at the reversal points (Zanone & Kelso, 1992), whereas the continuous relative phase measure reflects coordination throughout an entire cycle. In computing the point estimates, the elbow reversals were always the reference event and defined the cycle time. The wrist reversals were always the target events. The point estimate measure converts the relative time of a target event within a cycle of a reference event into a relative phase value ranging from 0° to ±180°. A mean and standard deviation were computed for flexion (ϕ FMn, ϕ FSD) and extension (ϕ EMn, ϕ ESD) reversal events.
Individual joint measures
Elbow and wrist amplitudes and elbow and wrist harmonicity measures were computed from the elbow and wrist angular time series. The angle values associated with flexion and extension peaks were used to compute half-cycle amplitudes for the wrist and elbow for every cycle in a trial. A mean wrist and elbow joint amplitude for each trial was computed from the half-cycle amplitudes as was a standard deviation.
A measure known as movement harmonicity (H) was computed from the wrist and elbow angular acceleration time series. Harmonicity provides an estimate to the extent that terminal braking and movement re-acceleration fuse to save mechanical energy during a movement reversal (Guiard, 1993). This measure has also been used to provide an estimate of the extent that a repetitive action is harmonic or inharmonic in nature (Buchanan, 2013; Buchanan, Park, Ryu, & Shea, 2003; Buchanan, Park, & Shea, 2006; Guiard, 1993, 1997). The first step in computing the value of H is to locate zero crossings in a displacement time series measured with respect to the spatial midpoint of an oscillation (Fig. 1c). Location of zero crossings in the normalized displacement trace segments the time series into a set of non-overlapping time windows, with each time window containing a movement reversal. In the current task, time windows were identified for both elbow and wrist flexion and extension reversals independently. The second step identifies local extrema (minima and maxima) in the normalized acceleration time series for each window in which a movement reversal occurs. The third step examines the ratio between the maximum and minimum extrema in each time window. The value of H varies from 0 to 1. For time windows with positive displacement and negative acceleration, H was computed as follows, H = maximum/minimum. For time windows with negative displacement and positive acceleration, H was computed as H = minimum/maximum (Fig. 1c left). If the acceleration time series is sinusoidal and only one local extrema occurs in a time window the value of H = 1 (because the ratio is of the local extrema to itself) and this indicates harmonic motion (Fig. 1c middle). If motion comes to a full stop for a prolonged period (such that an acceleration extrema = 0), then the value of H = 0 representing in-harmonic motion (Fig. 1c right). In the current task, the value of H will be used to determine if physical practice results in joint angular motion that is more harmonic in nature compared to observational practice. For each trial, a mean value of H was computed for the flexion and extension reversal events of the elbow and wrist angular time series.
Statistics
The analyses presented focused on isolating differences in kinematics between the three groups across the two performance tests. The continuous relative phase data were analyzed in a 3 × 2 ANOVA with Training context (physical-practice, observation-physical, observation-practice) and Performance test (performance-animation, performance-retention) as factors. The point estimate measures of relative phase were analyzed in a 3 × 2 × 2 ANOVA with Training context, Performance test, and Reversal point (flexion, extension) as factors. The joint amplitude means and standard deviations were analyzed in a 3 × 2 × 2 ANOVA with Training context, Performance Test, and Joint (elbow and wrist) as factors. The harmonicity data were analyzed in a 3 × 2 × 2 × 2 ANOVA with Training context, Performance test, Joint, and Reversal point as factors.
Results
Pre-training performance
Distributions of the continuous (ϕ CMn) mean relative phase values from the familiarization trials show that all three groups distinguished the initial three different elbow–wrist coordination patterns (Fig. 2a–c). When viewing the training pattern of 90°, the elbow leading feature in the animation was consistently identified as evidenced by the positive values in the distribution for the attempts at the 90° pattern (solid lines), even though the target pattern of 90° was not consistently produced.

Distributions of mean relative phase based on the continuous relative phase measure (ϕ CMn): a physical-practice group, b observation-physical group, and c observation-practice group
Post-training performance: relative phase distributions
Performance-animation test
Overall, 92 % of the continuous mean relative phase values were within a range of 45° < ϕ CMn < 135°, and this shows that the participants were successful at learning the target training pattern of 90° (Fig. 3a). Ten trials fell outside the above range and were produced by two participants (5 trials each), a physical-practice participant and an observation-practice participant.

Distributions of mean relative phase based on the continuous relative phase measure for the performance-animation (a) and performance-retention (b) tests: Phy physical-practice group, Obs-phy observation-physical group, and Obs observation-practice group
Performance-retention test
Across the three groups, 85 % of the continuous relative phase means fell within the range of 45° < ϕ CMn < 135° (Fig. 3b), and this also shows success at learning the target relative phase of 90° across the groups. For the 15 % of trials outside this range, 10 were produced by two observation-physical participants (with one the same participant with poor performance in the performance-animation test), 4 were produced by one physical-practice participant (the same participant with poor performance in the performance-animation test), and 3 were produced by three different observation-practice participants.
The relative phase data for the trials that fell outside the range of ±45° around the 90° target value were not statistically analyzed.Footnote 1
Post-training performance: relative phase measures
Continuous relative phase
The analysis of the mean continuous relative phase (ϕ CMn) data revealed a significant main effect for Training context, F(2, 199) = 5.91, p < .01, η 2 = .04. A post hoc test (Tukey–Kramer p < .05) revealed that the mean performance of the physical-practice group was significantly different from the two observer groups and was closer to the target of 90° (Fig. 4a, c). The Performance test main effect was found to be significant, F(1, 199) = 16.15, p < .01, η 2 = .06, with mean continuous relative phase farther from 90° in the performance-animation test (Fig. 4a) compared to the performance-retention test (Fig. 4c).

Mean relative phase and variability in the relative phase are plotted for the performance-animation test (a, b) and the performance-retention test (c, d). The dashed lines in a and c represent the target relative phase of 90°. The mean relative phase and variability in relative phase for the flexion reversal, continuous relative phase, and extension reversal are plotted by group in each plot. The asterisks represent significant differences between the physical-practice group and the other two groups. The error bars represent 1 standard deviation around the mean
The analysis of the continuous relative phase variability data (ϕ CSD) found a significant main effect for Training context, F(2, 199) = 35.25, p < .01, η 2 = .23. A post hoc test (Tukey–Kramer p < .05) found that variability in performance was significantly less in the physical-practice group compared to the two observer groups (Fig. 4b, d). The Performance test main effect was also found to be significant, F(1, 199) = 7.72, p < .01, η 2 = .03, with variability smaller in the performance-animation test (Fig. 4b) compared to the performance-retention test (Fig. 4d).
Point estimate relative phase
The analysis of the point estimate relative phase means (ϕ FMn and ϕ EMn) revealed significant main effects of Training context, F(2, 405) = 11.03, p < .01, η 2 = .05, and Reversal, F(1, 405) = 81.4, p < .01, η 2 = .16. The interaction of Training context × Reversal was significant, F(2, 405) = 3.67, p < .05, η 2 = .02. Post hoc tests (p < .05) revealed that the relative phase means for the flexion reversal were different from the extension reversal across all three groups (Fig. 4a, c). Also, the relative phase means at the flexion reversal for the physical-practice group were significantly different from the two observer groups, with no difference found between groups for the extension reversal. The interaction of Performance test × Reversal was also significant, F(1, 405) = 6.3, p < .01, η 2 = .02. Post hoc tests of this interaction found that the relative phase means for the flexion reversal were different from the extension reversal across both tests, with a significant difference in relative phase found for the flexion reversal between the performance-animation (Fig. 4a) and performance-retention (Fig. 4c) tests.
The analysis of the point estimate relative phase variability data (ϕ FSD and ϕ ESD) found a significant main effect of Training context, F(2, 405) = 35.5, p < .01, η 2 = .15. Post hoc tests (p < .05) found that the physical-practice group was characterized by less variability compared to the two observation groups across both performance tests (Fig. 4b, d). The main effect of Reversal was also significant, F(1, 405) = 18.28, p < .01, η 2 = .04, and on average variability was slightly larger for the flexion reversal compared to the extension reversal across both performance tests (Fig. 4b, d).
In the performance-animation test, the physical-practice group on average was the only group to overshoot the target of 90° at the flexion reversal (ϕ FMn), while for the extension reversal (ϕ EMn) all three groups undershoot the target of 90° (Fig. 4a). In the performance-retention test, all three groups undershoot the target of 90° for both reversal points, a different pattern compared to the retention test (Fig. 4c). This suggests a performance difference between groups as a function of the reversal point in the two tests that was not revealed in the previous ANOVA. To further examine the difference around the reversal points, separate ANOVAs with Training context as the only factor were performed on the data for each reversal point for each performance test.
In the performance-animation test, the Training context effect was found to have a significant effect on performance at the flexion reversal, F(2, 107) = 8.47, p < .01, η 2 = .16. Post hoc tests of the flexion reversal found that performance of the physical-practice group was different from the observation-practice and observation-physical groups in this test (Fig. 4a). The analysis performed on the flexion data from the performance-retention test found that the Training context effect approached standard levels of significance (p = .054) (Fig. 4b). For the extension event in the performance-animation test, Training context was also found to be significant, F(2, 107) = 5.07, p < .01, η 2 = .09. The post hoc tests of the extension reversal found a significant difference between the physical-practice group and observation-physical group (Figure 4a). The ANOVA performed on the extension reversal data from the performance-retention test did not find a significant Training context effect (p = .32) (Fig. 4c).
Post-training performance: joint amplitudes
The analysis of the amplitude means revealed a significant main effect of Joint, F(1, 461) = 16.12, p < .01, η 2 = .03. The Performance test main effect was also found to be significant, F(1, 461) = 17.31, p < .01, η 2 = .03, with joint amplitudes on average smaller in the performance-animation test (Mn = 74°, SD = 16°) compared to the performance-retention test (Mn = 80°, SD = 17°). The Training context × Joint interaction effect, F(2, 461) = 12.31, p < .01, η 2 = .04, was significant. Post hoc tests (p < .05) found that only the physical-practice group had a significantly larger elbow compared to wrist amplitude across both performance tests (Fig. 5a, c). The wrist amplitude of the physical-practice group was significantly smaller than the observation-physical and observation-practice groups across both performance tests.

Amplitude means and variability in amplitude are plotted as a function of training context for the performance-animation (a, b) and performance-retention (c, d) tests. The dashed lines in a and c reflect the required joint amplitude for the elbow (92°) and wrist (62°) as represented in the animation. The asterisks represent a significant difference between the elbow and wrist within a group. The error bars represent 1 standard deviation around the mean
The analysis of the amplitude standard deviations revealed significant effects of Training context, F(2, 461) = 22.46, p < .01, η 2 = .07, and Joint F(1, 461) = 41.59, p < .01, η 2 = .07. The Training context × Joint interaction was found to be significant, F(2, 461) = 7.81, p < .01, η 2 = .03. Post hoc tests (p < .05) of the interaction revealed that joint variability was only different in the two observation groups, with both groups having more variability in wrist amplitudes compared to elbow amplitudes in both performance tests (Fig. 5b, d). The post hoc tests also revealed that only in the wrist was there a difference in variability between groups, with the physical-practice group less variable than the observation-practice group across both tests.
Post-training performance: joint harmonicity
The analysis of the harmonicity data found significant main effects of Joint, F(1, 929) = 684.73, p < .01, η 2 = .30, Reversal point, F(1, 929) = 172.75, p < .01, η 2 = .08, Training context, F(2, 929) = 7.70, p < .01, η 2 = .01, and Performance test, F(1, 929) = 4.15, p < .05, η 2 = .01. The two-way interactions of Training context × Joint, F(2, 929) = 13.23, p < .01, η 2 = .01, Training context × Reversal, F(2, 929) = 25.20, p < .01, η 2 = .02, and Joint × Reversal, F(1, 929) = 302.23, p < .01, η 2 = .13, were significant. The three-way interaction of Training context × Joint × Reversal was also significant, F(2, 929) = 22.92, p < .01, η 2 = .02. Simple main effect tests (p < .05) of the three-way interaction revealed the following three main findings. First, harmonicty on average for the elbow flexion reversal was larger than the elbow extension reversal in all three groups for both tests (Fig. 6a, c), with no difference in harmoncity between the wrist flexion and extension reversals in any of the groups (Fig. 6b, d). Second, elbow flexion harmonicity was significantly larger than wrist flexion harmoncity in each group, while elbow extension harmoncity was larger than wrist extension harmonicity only for the two observation groups. Third, the physical-practice group had significantly larger elbow flexion harmonicity values compared to the two observation groups in both tests (Fig. 6a, c), with no difference in harmonicity found between groups for elbow extension, wrist flexion or wrist extension.

The mean harmonicity values from the performance-animation test (a, b) and the performance retention test (c, d) are plotted as a function of training context. The asterisks represent a significant difference between flexion and extension events within a group. The error bars represent 1 standard deviation around the mean
Discussion
This experiment was designed to reveal if physical and observational practice contribute differently to the ability to acquire and retain three specific types of kinematic movement information, relative motion (lead/lag), relative phase, and joint amplitude. Three specific predictions were put forth regarding the contributions of physical and observational practice. The two performance tests produced data that supported predictions 1 and 3 and partially supported prediction 2. The findings are discussed within a framework drawing upon the visual perception perspective of observation learning and the coordination dynamic viewpoint of relative phase as an informational variable that links perception to action and vice versa (Bingham et al., 1999; Kelso, 1994; Scully & Newell, 1985).
Relative motion and relative phase
Overall, the performance-animation and performance-retention tests revealed that all three practice contexts supported the extraction and use of relative motion (lead/lag) information in this task and, thereby, lend support to the first prediction. What emerged with regard to relative motion were not distinct group differences, but individual participant differences with respect to performance failure or lack of learning in primarily two individuals. There was no conflict regarding the contribution of physical practice and observation in this task as seen in previous studies (Al-Abood et al., 2001; Breslin, Hodges, Williams, Curran, & Kremer, 2005; Hodges et al., 2005, 2007). Thus, the physical practice offered no benefit over observation with regard to extracting and using relative motion information. The results are consistent with the idea that relative motion direction is a topological invariant extracted through visual perception processes that can be used to initially organize timing aspects of outgoing muscle commands (Buchanan, 2015; Buchanan et al., 2015; Scully & Carnegie, 1998; Scully & Newell, 1985). Some research has shown that relative motion direction may be the information that supports the perceptual identification of relative phase (Wilson et al., 2005). In the current task, 27 trials did not fall within a range of ±45° of the training pattern of ϕ = 90°. Of these 27 trials, 24 were characterized by the elbow leading the wrist, the correct relative motion direction, yet the wrong relative phase. What did characterize these trials was a strong attraction to in-phase and anti-phase, which previous research has shown to be very stable patterns for this elbow–wrist rhythmic task (Buchanan & Kelso, 1993; Kelso, Buchanan, & Wallace, 1991). Overall, the findings show that relative motion direction and relative phase are independent sources of information in this task that can be extracted through visual perception processes and this finding is consistent with recent research (Buchanan, 2015; Buchanan et al., 2015). In other words, even the trials that showed an attraction to in-phase and anti-phase were most often characterized by the elbow leading the wrist as required.
The second prediction stated that the physical-practice group would be closer to the required relative phase in both performance tests based on all three relative phase measures. The continuous relative phase data and flexion point estimate relative phase data support this hypothesis. The physical-practice group in the overall analysis did not have an advantage at the extension reversal. For all three groups, performance was closer to the target in the perception–retention test based on the continuous and flexion relative phase measures, yet not for the extension measure. As predicted, the physical-practice group had an advantage because both tests were consistent with part of their training context. The small amount of physical practice for the observation-physical group did not allow them to separate from the observation-practice group. Even with the advantage associated with the training context, both observation groups were quite successful in acquiring and using relative phase information to coordinate their elbow and wrist motions.
Further tests of the point estimate measures revealed differences between groups that were most prominent in the performance-animation test. In the performance-animation test, the physical group had an advantage at the flexion reversal over both observation groups, with an advantage over the observation-physical group at the extension reversal. In the performance-retention test, no significant difference was found between the groups at either reversal, although the data for the flexion reversal approached standard levels of significance. Why the difference between the two tests? In the performance-animation test, the physical group was unique in that the relative phase for the flexion reversal on average was above the target of 90° while relative phase for the extension reversal on average was below the target of 90°. This condition matched the training for the physical-practice group, and most probably allowed the physical-practice group to better match visual and proprioceptive information when the stimulus was present. The limited physical practice for the observation-physical group did not help in this testing context because their practice was not associated with the concurrent matching of visual and proprioceptive information. Another aspect that may have contributed to the difference between tests is that the extensive observation for both observation groups allowed for a very rapid tuning of the relative phase pattern in the performance-animation test. Even though augmented feedback was not provided, the observation-practice group did receive for the first time proprioceptive information about the joint motions and this may have helped with the rapid tuning. Combined, the two tests show that observation contributes significantly to the extraction and use of relative phase information in this single-limb task when performance must rely only on an internal representation of the required action. Even though observation may allow for the ability to extract and use relative phase information with limited physical practice, the physical-practice group’s elbow–wrist coordination pattern was more stable overall in both performance tests. This shows that physical practice allows for a stabilization of coordination that observation cannot support to the same extent. This stabilization most probably comes from the use of proprioceptive information during training, and the limited physical training of the observation-physical group was not enough to increase stability in performance.
The different practice contexts did have an impact on performance around the reversal points and even though a specific prediction was not made the data support the general prediction that the physical-practice group would have an advantage. Reversal points in limb motion often function as so-called anchor points that help stabilize in-phase and anti-phase rhythmic actions (Byblow et al., 1994; Fink et al., 2000). For example, instructing participants to synchronize maximum pronation or supination of the forearms or maximum flexion–extension of the wrists to a metronome signal will reduce variability at the reversal compared to other points in the limbs’ motion. Studies have shown that individuals will utilize reversal points without instruction to help learn a 90° relative phase pattern in a bimanual task (Zanone & Kelso, 1997). In the current task, the physical-practice group seemed to organize the movement more around elbow flexion to better achieve the global pattern. A distinct difference between the current findings and those of Zanone and Kelso (1997) was that the use of the reversal point did not impact global performance. In the Zanone and Kelso (1997) task, limb motion away from the reversal point was often attracted to 0° and 180°. The linkage between the elbow and wrist, however, may play a significant role in the different outcomes, as might the nature of the training stimulus. These are issues that can be explored in future research.
Previous research has argued that combined physical/observation practice may produce a social facilitation effect that leads to acquiring different knowledge about the task compared to physical or observational practice alone (Shea et al., 2000). Previous research has also argued that observational training supports the development of spatial codes more so than motor codes for action production (Ellenbuerger et al., 2012). In the current task, all participants were trained with the same visual stimulus and the observers were not yoked to a live model, thereby eliminating the possibility of a social facilitation effect. The similarity in performance based on relative motion direction across both tests in all three groups shows that physical, observation, and combined practice all support the development of spatial codes. The rapid gain based on relative phase performance between the two tests in the observation-practice group shows that observation alone most probably supports the development of motor codes to a certain extent in that the observation-practice group did not have an advantage in either test. The ability to produce the correct relative phase, regardless of relative motion direction, must require a developed motor code, since timed motor commands will be required to achieve specific relative phase patterns (Kelso et al., 1991). However, the efficiency of the motor codes clearly benefits from extensive physical-practice as evidenced by the greater stability in performance of the physical-practice group (Buchanan et al., 2008, 2015).
Joint amplitude scaling
The third prediction was directed at the amplitude components of the required task and stated that the physical-practice group would be able to produce different amplitudes between the joints as required. This hypothesis was supported in that the physical-practice group on average produced smaller wrist compared to elbow amplitudes in both performance tests. This finding is consistent with the Scully and Newell (1985) hypothesis that observation will not benefit the scaling of absolute motion features (Al-Abood et al., 2001; Buchanan et al., 2008, 2015). The analysis of the amplitude variability data revealed some similarities between the groups. For example, the physical-practice and observation-physical groups were characterized by less variability in joint amplitudes than the observation-practice group across both performance tests. This suggests that minimal physical practice did benefit the consistency of amplitude performance even if absolute scaling was not benefitted. In both performance tests, elbow amplitude variability was smaller than wrist variability overall. Differences in variability between groups were found only in the wrist joint with both observation groups characterized by larger wrist variability than elbow variability. Although neither observation group was good at producing different joint amplitudes on average, the limited amount of physical practice for the observation-physical group did result in less variability in wrist amplitudes. The big advantage for physical practice was linked more to reducing variability in wrist amplitudes and separating amplitudes, with both observation and physical practice contributing equally to the consistency of elbow motion. This suggests that observation, while not supporting the ability to scale amplitude, did not detract from possible organizational features of the action linked primarily to elbow motion.
Previous research using a live learning model producing the elbow–wrist pattern used in this task showed that observers have problems with producing joint amplitude differences (Buchanan et al., 2008). The use of a live learning model and the variability in their performance may have contributed significantly to the inability of observers to produce different joint amplitudes in the previous work. However, in this task, there was no model variability. Thus, the inability to produce the different joint amplitudes emerges from the observation context and associated processes. This is a key finding based on the idea of an action-observation network wherein similar neural areas are activated during both action-production and action-observation (Andrieux & Proteau, 2013; Calvo-Merino, Grezes, Glaser, Passingham, & Haggard, 2006; Cross, Hamilton, & Grafton, 2006; Cross, Kraemer, Hamilton, Kelley, & Grafton, 2009). The current results suggest that activation in the action-observation network may be primarily linked to producing more general kinematic features of the task, i.e., relative motion, relative phase, limb sequences, than to the ability to produce distinct joint amplitudes. An interesting issue is whether observation supports the perceptual identification of amplitude differences and just does not support the ability to tune the muscle commands (as does physical practice) to achieve the required differences. The use of a transfer paradigm with this elbow–wrist task suggested that observers do perceptually identify joint amplitudes differences through visual processes, yet have difficulty modulating the amplitudes (Buchanan & Wright, 2011). The issue of perceptually identifying amplitude differences through visual processes and the practice required to achieve them requires more extensive research.
Joint harmonicity
The harmonicity measure has been used to characterize the harmonic or inharmonic motion of the end-effector in reciprocal aiming tasks (Buchanan, 2013; Guiard, 1993; Kovacs, Buchanan, & Shea, 2008). Recently, it has been used to characterize the harmonic and inharmonic nature of the arms during bimanual coordination tasks with harmonicity values larger when relative phase variability and error are lower (Kovacs, Buchanan, & Shea, 2009, 2010). At both reversal points, harmonicity was significantly larger in the elbow for all groups in both tests, yet only the physical practice group was consistently more stable (relative phase variability) at both reversal points in both day three performance tests. With regard to elbow motion, harmonicity values were significantly larger for the flexion reversal compared to the extension reversal for all three groups. Moreover, the harmonicty values for the elbow flexion reversal were largest in the physical-practice group. Only for the extension reversals was either observation group characterized by larger harmonicty values for either joint, and the differences were inconsistent across the day three performance tests. Overall, the harmonicty data revealed two important features of the impact of practice context on performance: (1) regardless of practice context, elbow flexion emerged as an important kinematic event around which the learning process was organized; and (2) more physical practice produced a greater advantage with regard to organizing the learning process around elbow motion, yet had no consistent impact on wrist motion production.
Why did this distinct advantage emerge with respect to the elbow reversal points compared to the wrist reversal points? One idea regarding the control and coordination of the shoulder, elbow, and wrist is that one joint acts as the leading joint during arm movements (Dounskaia, 2005). Research has defined the relationship between the leading joint and secondary joints primarily through an analysis of muscle torque and interactive torque (Dounskaia & Wang, 2014). The leading joint hypothesis proposes that during arm movements one joint is rotated actively while the active control of other linked joints or trailing joints is minimized. Often the control of the trailing joint(s) motion is viewed as mostly passive to simplify the coordination between joints. Most of the research that has been used in support of the leading joint hypothesis has focused on overlearned coordination patterns between the shoulder and elbow joints. The issue emerges as to whether or not the learning observed in this task was the result of passive interactions between the elbow and wrist if the elbow acts as a leading joint. A previous study using elbow–wrist motions similar to those employed in this experiment addressed this issue (Kelso et al., 1991). In one phase of the Kelso et al. (1991) study, participants were asked to produce in-phase (ϕ = 0°) elbow–wrist coordination through active and passive motions of the wrist. In the active condition, participants were instructed to actively flex and extend the wrist and elbow, and in the passive condition, participants were told to actively flex and extend the elbow and to relax the wrist as much as possible, i.e., let it be passively moved by the elbow’s motion. The active instructions led to a relative phase relationship of ϕ = 6.4° while the passive instructions lead a relative phase relationship of ϕ = 13.8°. These results show that the ability to produce the observed relative phase patterns in the performance tests required participants to actively control wrist motion and that the observed performance did not arise from just passive forces associated with the mechanical link.
Another factor that may account for the differences in elbow flexion and extension harmonicity is gravity. The flexion motions in this task require moving against gravity. More physical practice may have provided a greater opportunity for the physical-practice group to learn how to organize the action with respect to the impact of gravity, leading to larger harmonicity values at flexion and lower harmonicty values at extension. Future work needs to explore the impact of gravity on learning novel joint coordination patterns such as the ones used in the current task. For example, learning a 90° relative phase pattern with the forearm and wrist moving on the horizontal plane instead of the sagittal plane may alter performance regarding point estimates of relative phase and movement harmoncity.
Conclusions
The biggest difference between observation and physical practice did not emerge at the global level of performance defined by relative motion and continuous relative phase. Observation alone was beneficial in extracting and using relative motion and relative phase information to organize successful attempts of the coordination pattern without extensive physical practice. This is consistent with the idea that relative phase is an informational variable linking perception to action and vice versa (Bingham et al., 1999; Buchanan, 2015; Buchanan et al., 2015; Kelso, 1994; Kelso & Pandya, 1991; Zaal et al., 2000). The differences found in joint amplitudes as a result of physical and observational practice are consistent with the idea of scaling absolute motion features as defined by the visual perception theory of observation (Scully & Newell, 1985). The most unexpected difference emerged with regard to the flexion reversal point as evidenced in the point estimate relative phase data and harmonicity data. Overall, it may be concluded that in this task physical practice provided the opportunity to fine tune specific types of kinematic information that are important for amplitude control, generating harmonic motion, and stabilizing coordination.
Notes
The physical-practice participant that performed poorly during both performance tests showed a significant improvement in performance with practice. No explanation for poor performance during the performance tests is readily available. However, both the physical-practice participant and observer-practice participant that performed poorly based on the relative phase measures did perform very similar to the other participants in their groups based on joint amplitudes and harmonicity measures.
References
Al-Abood, S. A., Davids, K., & Bennett, S. J. (2001a). Specificity of task constraints and effects of visual demonstrations and verbal instructions in directing learners’ search during skill acquisition. Journal of Motor Behavior, 33(3), 295–305.
Al-Abood, S. A., Davids, K., Bennett, S. J., Ashford, D., & Marin, M. M. (2001b). Effects of manipulating relative and absolute motion information during observational learning of an aiming task. Journal of Sports Sciences, 19, 507–520.
Andrieux, M., & Proteau, L. (2013). Observation learning of a motor task: Who and when? Experimental Brain Research, 229(1), 125–137. doi:10.1007/s00221-013-3598-x.
Andrieux, M., & Proteau, L. (2014). Mixed observation favors motor learning through better estimation of the model’s performance. Experimental Brain Research, 232(10), 3121–3132. doi:10.1007/s00221-014-4000-3.
Badets, A., & Blandin, Y. (2010). Feedback schedules for motor-skill learning: The similarities and differences between physical and observational practice. Journal of Motor Behavior, 42(4), 257–268.
Bingham, G. P., Schmidt, R. C., & Zaal, F. T. J. M. (1999). Visual perception of the relative phasing of human limb movements. Perception and Psychophysics, 61(2), 246–258.
Bingham, G. P., Zaal, F., Shull, J. A., & Collins, D. R. (2001). The effect of frequency on the visual perception of relative phase and phase variability of two oscillating objects. Experimental Brain Research, 136(4), 543–552. doi:10.1007/s002210000610.
Bird, G., Osman, M., Saggerson, A., & Heyes, C. (2005). Sequence learning by action, observation and action observation. British Journal of Psychology, 96, 371–388. doi:10.1348/000712605x47440.
Breslin, G., Hodges, N. J., Williams, A. M., Curran, W., & Kremer, J. (2005). Modelling relative motion to facilitate intra-limb coordination. Human Movement Science, 24, 446–463.
Breslin, G., Hodges, N. J., Williams, A. M., Kremer, J., & Curran, W. (2006). A comparison of intra- and inter-limb relative motion information in modeling a novel motor skill. Human Movement Science, 25, 753–766.
Buchanan, J. J. (2013). Flexibility in the control of rapid aiming actions. Experimental Brain Research, 229(1), 47–60. doi:10.1007/s00221-013-3589-y.
Buchanan, J. J. (2015). Perceptual estimates of motor skill proficiency are constrained by the stability of coordination patterns. Journal of Motor Behavior. doi:10.1080/00222895.2015.1008687.
Buchanan, J. J., & Dean, N. (2010). Specificity in practice benefits learning in novice models and variability in demonstration benefits observational practice. Psychological Research, 74, 313–320.
Buchanan, J. J., & Dean, N. (2014). Consistently modeling the same movement strategy is more important than model skill level in observational learning contexts. Acta Psychologica, 146, 19–27.
Buchanan, J. J., & Kelso, J. A. S. (1993). Posturally induced transitions in rhythmic multijoint limb movements. Experimental Brain Research, 94, 131–142.
Buchanan, J. J., Park, J. H., Ryu, Y. U., & Shea, C. H. (2003). Discrete and cyclical units of action in a mixed target pair aiming task. Experimental Brain Research, 150, 473–489.
Buchanan, J. J., Park, J. H., & Shea, C. H. (2006). Target width scaling in a repetitive aiming task: Switching between cyclical and discrete units of action. Experimental Brain Research, 175, 710–725.
Buchanan, J. J., Ramos, J., & Robson, N. (2015). The perception–action dynamics of action competency are altered by both physical and observational training. Experimental Brain Research, 233, 1289–1305. doi:10.1007/s00221-015-4207-y.
Buchanan, J. J., Ryu, Y. U., Zihlman, K., & Wright, D. A. (2008). Observational practice of a relative phase pattern but not an amplitude ratio in a multijoint task. Experimental Brain Research, 191, 157–169.
Buchanan, J. J., & Wright, D. L. (2011). Generalization of action knowledge following observational learning. Acta Psychologica, 136, 167–178.
Byblow, W. D., Carson, R. G., & Goodman, D. (1994). Expressions of asymmetries and anchoring in bimanual coordination. Human Movement Science, 13, 3–28.
Calvo-Merino, B., Grezes, J., Glaser, D. E., Passingham, R. E., & Haggard, P. (2006). Seeing or doing? Influence of visual and motor familiarity in action observation. Current Biology, 16(19), 1905–1910. doi:10.1016/j.cub.2006.07.065.
Celnik, P., Webster, B., Glasser, D. M., & Cohen, L. G. (2008). Effects of action observation on physical training after stroke. Stroke, 39(6), 1814–1820. doi:10.1161/strokeaha.107.508184.
Cross, E. S., Hamilton, A., & Grafton, S. T. (2006). Building a motor simulation de novo: Observation of dance by dancers. Neuroimage, 31(3), 1257–1267. doi:10.1016/j.neuroimage.2006.01.033.
Cross, E. S., Kraemer, D. J. M., Hamilton, A. F. D., Kelley, W. M., & Grafton, S. T. (2009). Sensitivity of the action observation network to physical and observational learning. Cerebral Cortex, 19, 315–326.
Cutting, J. E., & Proffitt, D. R. (1982). The minimum principle and the perception of absolute, common, and relative motions. Cognitive Psychology, 14, 211–246.
Dounskaia, N. V. (2005). The internal model and the leading joint hypothesis: Implications for control of multi-joint movements. Experimental Brain Research, 166, 1–16.
Dounskaia, N. V., & Wang, W. (2014). A preferred pattern of joint coordination during arm movements with redundant degrees of freedom. Journal of Neurophysiology, 112(5), 1040–1053. doi:10.1152/jn.00082.2014.
Ellenbuerger, T., Boutin, A., Blandin, Y., Shea, C. H., & Panzer, S. (2012). Scheduling observational and physical practice: Influence on the coding of simple motor sequences. Quarterly Journal of Experimental Psychology, 65(7), 1260–1273. doi:10.1080/17470218.2011.654126.
Fink, P. W., Foo, P., Jirsa, V. K., & Kelso, J. A. S. (2000). Local and global stabilization of coordination by sensory information. Experimental Brain Research, 134, 12.
Franceschini, M., Ceravolo, M. G., Agosti, M., Cavallini, P., Bonassi, S., Dall’Armi, V., & Sale, P. (2012). Clinical relevance of action observation in upper-limb stroke rehabilitation: A possible role in recovery of functional dexterity. A randomized clinical trial. Neurorehabilitation and Neural Repair, 26(5), 456–462. doi:10.1177/1545968311427406.
Gibson, J. J. (1979). The ecological approach to visual perception. Boston, MA: Houghton Mifflin.
Gruetzmacher, N., Panzer, S., Blandin, Y., & Shea, C. H. (2011). Observation and physical practice: Coding of simple motor sequences. Quarterly Journal of Experimental Psychology, 64(6), 1111–1123. doi:10.1080/17470218.2010.543286.
Guiard, Y. (1993). On Fitts’s and Hooke’s laws: Simple harmonic movement in upper-limb cyclical aiming. Acta Psychologica, 82, 139–159.
Guiard, Y. (1997). Fitts’ law in the discrete vs. cyclical paradigm. Human Movement Science, 16(1), 97–131.
Hamilton, A., & Grafton, S. T. (2007). The motor hierarchy: From kinematics to goals and intentions. In P. Haggard, Y. Rossetti, & M. Kawato (Eds.), Sensorimotor foundations of higher cognition: Attention and performance XXll (pp. 381–408). Oxford: Oxford University Press.
Hayes, S. J., Hodges, N. J., Huys, R., & Williams, A. M. (2007). End-point focus manipulations to determine what information is used during observational learning. Acta Psychologica, 126, 120–137.
Hodges, N. J., Hayes, S. J., Breslin, G., & Williams, A. M. (2005). An evaluation of the minimal constraining information during observation for movement reproduction. Acta Psychologica, 119(3), 264–282. doi:10.1016/j.actpsy.2005.02.002.
Hodges, N. J., Williams, A. M., Hayes, S. J., & Breslin, G. (2007). What is modelled during observational learning? Journal of Sports Sciences, 25(5), 531–545.
Jacobs, A., & Shiffrar, M. (2005). Walking perception by walking observers. Journal of Experimental Psychology-Human Perception and Performance, 31(1), 157–169. doi:10.1037/0096-1523.31.1.157.
Johansson, G. (1973). Visual perception of biological motion and a model for its analysis. Perception and Psychophysics, 14, 201–211.
Kelso, J. A. S. (1994). The informational character of self-organized coordination dynamics. Human Movement Science, 13, 393–413.
Kelso, J. A. S., Buchanan, J. J., & Wallace, S. A. (1991). Order parameters for the neural organization of single limb, multijoint movement patterns. Experimental Brain Research, 85, 432–445.
Kelso, J. A. S., & Pandya, A. S. (1991). Dynamic pattern generation and recognition. In N. I. Badler, B. A. Barsky, & D. Zeltzer (Eds.), Making them move: Mechanics, control, and animation of articulated figures (pp. 171–190). San Mateo, CA: Morgan Kaufmann.
Kovacs, A. J., Buchanan, J. J., & Shea, C. H. (2008). Perceptual influences on Fitts’ law. Experimental Brain Research, 190, 99–103.
Kovacs, A. J., Buchanan, J. J., & Shea, C. H. (2009). Bimanual 1:1 with 90° degrees continuous relative phase: Difficult or easy! Experimental Brain Research, 193(1), 129–136. doi:10.1007/s00221-008-1676-2.
Kovacs, A. J., Buchanan, J. J., & Shea, C. H. (2010). Perceptual and attentional influences on continuous 2:1 and 3:2 multi-frequency bimanual coordination. Journal of Experimental Psychology: Human Perception and Performance, 36(4), 936–954.
Martens, R., Burwitz, L., & Zuckerman, J. (1976). Modeling effects on motor performance. The Research Quarterly, 47(2), 277–291.
Newell, K. M. (1991). Motor skill acquisition. Annual Review of Psychology, 42, 213–237.
Osman, M., Bird, G., & Heyes, C. (2005). Action observation supports effector-dependent learning of finger movement sequences. Experimental Brain Research, 165, 19–27.
Pinto, J., & Shiffrar, M. (1999). Subconfigurations of the human form in the perception of biological motion displays. Acta Psychologica, 102, 293–318.
Scholz, J. P., & Kelso, J. A. S. (1989). A quantitative approach to understanding the formation and change of coordinated movement patterns. Journal of Motor Behavior, 21(2), 122–144.
Scully, D. M. (1986). Visual perception of technical execution and aesthetic quality in biological motion. Human Movement Science, 5, 185–206.
Scully, D. M., & Carnegie, E. (1998). Observational learning in motor skill acquisition: A look at demonstrations. The Irish Journal of Psychology, 19(4), 472–485.
Scully, D. M., & Newell, K. M. (1985). Observational learning and the acquisition of motor skills: Toward a visual perception perspective. Journal of Human Movement Studies, 11, 169–186.
Shea, C. H., Wright, D. L., Wulf, G., & Whitacre, C. (2000). Physical and observational practice afford unique learning opportunities. Journal of Motor Behavior, 32(1), 10.
Shiffrar, M., Lichtey, L., & Chatterjee, S. H. (1997). The perception of biological motion across apertures. Perception and Psychophysics, 59(1), 51–59.
Wilson, A. D., Collins, D. R., & Bingham, G. P. (2005). Human movement coordination implicates relative direction as the information for relative phase. Experimental Brain Research, 165(3), 351–361.
Zaal, F. T. J. M., Bingham, G. P., & Schmidt, R. C. (2000). Visual perception of mean relative phase and phase variability. Journal of Experimental Psychology: Human Perception and Performance, 26(3), 1209–1220.
Zanone, P. G., & Kelso, J. A. S. (1992). The evolution of behavioral attractors with learning: Nonequilibrium phase transitions. Journal of Experimental Psychology: Human Perception and Performance, 18, 403–421.
Zanone, P. G., & Kelso, J. A. S. (1997). Coordination dynamics of learning and transfer: Collective and component levels. Journal of Experimental Psychology: Human Perception and Performance, 23(5), 1454–1480.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Buchanan, J.J., Park, I. Observation and physical practice: different practice contexts lead to similar outcomes for the acquisition of kinematic information. Psychological Research 81, 83–98 (2017). https://doi.org/10.1007/s00426-015-0723-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-015-0723-4