
Abstract
Main conclusion
The drought-stimulated gene expression of NCED, SUS, and KS - DHN and ABA signal cross-talk with other phytohormones maintains barley root growth under drought stress at pH 4.0 plus polyethylene glycol plus aluminum.
Aluminum (Al) toxicity and drought are two major factors that limit barley production. In this work, the individual and combined effects of Al/acid and polyethylene glycol (PEG 6000) induced drought stress that suppressed root growth and caused oxidative damage as characterized by increased H2O2 and \(\text{O}_{2}^{\cdot -}\) accumulation. The wild-barley genotypes, XZ5 and XZ29, exhibited a higher tolerance than the two cultivars Dayton (Al tolerant) and Tadmor (drought tolerant) under combined stress (pH 4.0 + PEG + Al). The oxidative damage induced by PEG was more severe at pH 4.0 than at pH 6.0. In XZ29, the highest root secretion of malate and citrate was recorded, and the least Al uptake in the four genotypes. In XZ5, a peak accumulation of ABA and minor synthesis of zeatin riboside and ethylene were found being essential in maintaining primary root elongation and root hair development. PEG-induced drought stress repressed Al uptake in root tips, with a lower increase in callose formation and HvMATE (Hordeum vulgare multidrug and toxic compound exudation) expression compared to Al-induced callose production. Stress by pH 4.0 + PEG + Al up-regulated 9-cis-epoxycarotenoid dioxygenase (NCED) which is involved in ABA biosynthesis. Such treatment stimulated the regulation of ABA-dependent genes sucrose synthase (SUS) and KS-type dehydrin (KS-DHN) in root tips. Our results suggest that the tolerance ranking to pH 4.0 + PEG + Al stress in Tibetan wild barley by gene expression is closely correlated to physiological indices. The results show that acclimatisation to pH 4.0 + PEG + Al stress involves specific responses in XZ5 and XZ29. The present study provides insights into the effects of Al/acid and drought combined stress on the abundance of physiological indices in the roots of barley varieties.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Abiotic stresses such as drought and soil acidity cause extensive losses of global agricultural productivity and thereby affect food security. Aluminum (Al) toxicity is a well-known major factor that limits crop productivity on acid soils: it affects up to 50 % of world’s potential arable land (Arroyave et al. 2013). Approximately 1/3 of world’s arable land currently faces yield reduction because of cyclical or unpredictable drought, which is a great threat to agricultural production (Chaves and Oliveira 2004). In many regions, drought and Al toxicity are two frequently co-occurring abiotic stresses (Beebe et al. 2011). To satisfy requirements of growing world population, it is essential to effectively use dehydrated and acid soils, which include lime application to raise soil pH and use of highly drought- and Al-resistant plants. However, the progress toward developing drought- and Al-tolerant crops is significantly hampered by the physiological and genetic complexity of these traits. Therefore, it is important to understand mechanisms of drought/Al tolerance in plants and identify genetic resources that could result in high levels of tolerance for multiple stresses.
The root apex is the most Al-sensitive root zone, and transition zone (1–2 mm) and elongation zone are targets of Al injury (Delhaize and Ryan 1995). Excess Al results in a rapid inhibition of root elongation (Delhaize and Ryan 1995; Staß and Horst 2009). Root elongation is also inhibited under water stress, but there are two main differences between Al stress and drought stress: (1) under drought stress, shoot growth is much more affected than root growth (Yamaguchi and Sharp 2010), whereas short- and medium-term Al excess may strongly reduce root growth without affecting shoot growth (Kochian et al. 2004). (2) Aluminum toxicity reduces cell elongation along entire elongation zone (Kollmeier et al. 2013), whereas under water deficit or osmotic stress, cell elongation is only inhibited in basal and central elongation zones (definition of zones according to Ishikawa and Evans 1993) and maintained toward root apex in the distal and apical elongation zones (Shimazaki et al. 2005). The maintenance of root growth during water deficit is a prerequisite for water uptake from subsoil (Serraj and Sinclair 2002). In maize, three mechanisms in primary root growth maintenance under water deficit have been proposed: osmotic adjustment, modification of cell wall (CW) extension properties, and the role of ABA accumulation (Sharp et al. 2004; Yamaguchi and Sharp 2010). It was expected and hypothesized that low soil moisture (drought) in an acid Al-toxic soil would aggravate Al toxicity, which further impedes root growth, which may strongly restrict water acquisition from subsoil and ability of plants to withstand drought stress (Goldman et al. 1989). It was shown that the response of plants to a combination of two different abiotic stresses was unique and could not be directly extrapolated from the response of plants to each individually applied stress (Mittler 2006). However, little information is available regarding combined stresses of drought and Al. Therefore, studies on the combination of these stresses are of considerable ecological significance and are needed to improve abiotic stress tolerance in plants.
Barley (Hordeum vulgare L.) is one of the most Al-sensitive species among small-grain cereals (Zhao et al. 2003). To breed barley cultivars that are tolerant or resistant to Al toxicity, it is important to identify genetic resources with significant potential for Al tolerance. Wild-barley germplasm is a treasure trove of useful genes and offers rich sources of genetic variation for crop improvement. In our previous work, we identified two Tibetan wild-barley genotypes (H. vulgare L. ssp. spontaneum), XZ5 and XZ29, with high tolerance to drought and Al stress, respectively (Zhao et al. 2010; Cai et al. 2013). However, the underlying physiological, biochemical and molecular mechanisms in combined drought and Al stresses in Tibetan wild-type barley have not been investigated and compared with Al-tolerant cv. Dayton (Wang et al. 2007) and drought-tolerant cv. Tadmor (Forster et al. 2004), thus preventing identification and exploiting of candidate genes into commercial barley cultivars. The knowledge of the underlying mechanisms of tolerance to combined stresses of drought and Al in wild barley is needed for successful breeding. The main objective of the present study was to compare the physiological and molecular levels of the short-term effects of combined acid/Al toxicity and drought stress in hydroponics using PEG as a substitute for drought. This study particularly emphasized the root growth in wild and cultivated barley.
Materials and methods
Plant materials and hydroponic culture
The hydroponic experiment was performed on Zijingang Campus of Zhejiang University in Hangzhou (China). Barley seeds were obtained from Zhejiang Key Laboratory of Crop Germplasm, Zhejiang University (China). Seeds of the Tibetan wild-barley genotypes XZ29 and XZ5 (H. vulgare L. ssp. spontaneum) and cv. Dayton and Tadmor were disinfected with 3 % H2O2 for 30 min and rinsed with distilled water. They were then soaked for 4 h at room temperature. Seeds were transferred onto moist filter papers in germination boxes in a growth chamber (22/18 °C, day/night) in darkness for 3 days and incubated for further 4 days in light. Seven-day-old uniform healthy seedlings were transplanted into 5-L containers, which were filled with 4.5 L basal nutrient solution (BNS). The composition of BNS was as described by Wu et al. (2003). Each container was covered with a polystyrene plate with 6 evenly spaced holes (2 plants per hole) and placed in a greenhouse. The plants were grown in a greenhouse with natural light and a temperature of 20 ± 2 °C/day and 15 ± 2 °C/night.
Treatments and sampling
Twelve days after transplanting, 100 μM Al (as AlCl3·6H2O) and/or 20 % (w/v) polyethylene glycol (PEG 6000) for drought stress (Liu et al. 2013) were added to the culture solution to form the following six treatments: pH 6.0 (control, BNS at pH 6.0); pH 4.0 (BNS at pH 4.0); pH 6.0 + PEG (BNS + 20 % PEG, pH 6.0); pH 4.0 + PEG (BNS + 20 % PEG, pH 4.0); pH 4.0 + Al (BNS + 100 μM Al, pH 4.0) and pH 4.0 + PEG + Al (BNS + 20 % PEG +100 μM Al, pH 4.0). The experiments were laid in a split-plot design with treatment as the main plot and genotype as sub-plot, and there were seven replicates for each treatment. The solution pH in each container was adjusted once a day with 1 M HCl or NaOH as required. The solution was continuously aerated with an air pump and renewed every 3 days. Root samples for physiological, biochemical and molecular studies were collected after 7 days of Al and PEG exposure. Fresh samples were immediately frozen in liquid nitrogen and stored frozen at −80 °C for further analyses or directly used for biochemical assays. All reagents were of analytical grade, and all stock solutions were prepared with deionized water.
Measurements of root length, dry weight, Al content and root hair observation
After 7 days treatment, plants were up-rooted, and root hairs were observed using a stereomicroscope (SZX12 Olympus, Tokyo, Japan). After root length was measured, the other plant samples were dried at 80 °C and weighed. The dried root samples were ground and digested in an acid mixture (HNO3:HClO4 = 4:1, v/v) at 150 °C for 6 h. Al contents were determined using inductively coupled plasma atomic emission spectrometry (ICP/AES) (IRIS/AP optical emission spectrometer, Thermo Jarrell Ash, San Jose, CA, USA).
Determination of hydrogen peroxide (H2O2), superoxide radical (\(\text{O}_{2}^{\cdot -}\)), and their histochemical detection
H2O2 and \(\text{O}_{2}^{\cdot -}\) in roots were extracted, and the content was determined spectro-photometrically as previously described by Willekens et al. (1997) and Jiang and Zhang (2002), respectively. H2O2 was detected using 3, 3-diaminobenzidine (DAB) staining according to Zeng et al. (2014) with some modification. Fresh root tips (~0.5 cm) were incubated in 1 mg/mL DAB–HCl solution for 5 h and washed once with a 2-N-morpholino-ethanesulfonic acid/potassium chloride (MES/KCl) buffer (10−3 M, pH 6.15). Superoxide anion (\(\text{O}_{2}^{\cdot -}\)) was detected using nitro blue tetrazolium (NBT) staining (Zeng et al. 2014). Root tip segments were dyed for 2 h with 0.1 mg/mL NBT (in 0.2 M phosphate buffer, pH 7.6) in darkness and subsequently washed once with phosphate buffer. After staining, roots were washed with distilled water for 10 min. All stained segments were observed using a Leica Fluorescence Stereomicroscope (Model MZ16 FA, Leica Microsystems, Heerbrugg, Switzerland) under visible light and photographed with a charge-coupled device (CCD) imaging system, which was fitted to microscope.
Visualization and determination of callose content
To quantitatively measure callose content, root samples were extracted with 1 N NaOH at 80 °C. The supernatants were mixed with an aniline blue solution (40 vol of 0.1 % aniline blue in water, 21 vol of 1 N HCl, and 59 vol of 1 M glycine/NaOH buffer, pH 9.5), and fluorescence was measured using a fluorospectrometer (Jobin–Yvon/SPEX) as previously described in Köhle et al. (1985). Callose was stained according to Zeng et al. (2014). Fresh root tips (~0.5 cm) were immediately immersed into 85 % (v/v) ethanol and incubated for over 12 h at room temperature (RT, approximately 25 °C) with gentle shaking. They were then transferred into 1 % (w/v) aniline blue solution in 1 M glycine (pH 9.5) for 5 h at RT and thoroughly washed with double distilled water. The cross-sections of root segments, which were stained with aniline blue, were observed under a fluorescence microscope (BX61, Olympus) with UV illumination. Micrographs for both lignification and callose were taken with a megapixel cooled CCD camera (DP71, Olympus) and Image-Pro Plus 6.0 software (Media cybernetics Inc., Rockville, MD, USA).
Determination of root organic-acid (OA) secretion
Root OA (citrate and malate) secretion was determined using HPLC (Waters 2695, Waters Corp, Milford, MA, USA), which was equipped with a Symmetry C18 column (4.6 mm i.d × 200 mm, 5 mm). The method was as described in Zheng et al. (2005) with some modifications. Briefly, mobile phase was a 10 mM (NH4)2HPO4 solution at pH 2.5 with a flow rate of 0.8 mL min−1 at 25 °C, and injection volume was 20 μL. The detection wavelength was set at 214 nm.
Analysis of abscisic acid (ABA), zeatin riboside (ZR) and 1-aminocyclopropane-1-carboxylic acid (ACC) oxidase
Extraction and analysis of endogenous plant hormones were measured according to Müller and Munné-Bosch (2011) method with some modifications. Briefly, around 0.3 g fresh root sample was ground in liquid nitrogen, and 1 mL of 80 % methanol was added to the sample, and homogenized using a vortex for 10 s every 15 min for a total of 2 h. Samples were centrifuged at 9600g for 15 min, and the supernatant was used for injection into liquid chromatography mass spectroscopy (LC–MS). Four replicates for each treatment were maintained and injected independently into the LC–MS.
Standard compounds, i.e. abscissic acid (ABA), zeatin riboside (ZR) and 1-aminocyclopropane-1-carboxylic acid (ACC) oxidase (ethylene), were purchased from Sangon (Shanghai, China). The standards were dissolved in 80 % methanol and five replicates for each standard were run in the LC–MS-MS (Agilent Technologies 6460 Triple Quad LC/MS, Agilent Technologies, Santa Clara, CA, USA) under the same conditions as the extracted samples.
Total RNA extraction, cDNA synthesis, and quantitative real-time PCR (qRT-PCR) analyses
Following manufacturer’s protocol, total RNA was extracted from ~100 mg of root tissue using Total RNA Miniprep Kit (Axygen Biosciences, Union City, CA, USA). Genomic DNA was removed using RNeasy Mini Kit (Qiagen, Hilden, Germany). The cDNA samples were assayed using quantitative real-time PCR (qRT-PCR) in iCycler iQTM Real-time PCR Detection System (Bio-Rad, Hercules, CA, USA) with SYBR Green PCR Master Mix (Applied Biosystems). PCR conditions consisted of denaturation at 95 °C for 3 min, 40 cycles of denaturation at 95 °C for 30 s, annealing at 58 °C for 45 s and extension at 72 °C for 45 s. Gene-specific primers (Table 1) for qRT-PCR were designed based on mRNA or expressed sequence tag (EST) using Primer Express software (Applied Biosystems). Software that was provided with PCR system was used to calculate threshold cycle values and mRNA levels was quantified according to the method of Livak and Schmittgen (2001). Threshold cycle (Ct) value of actin was subtracted from that of gene of interest to obtain a ΔCt value. The Ct value of untreated control sample was subtracted from ΔCt value to obtain a ΔΔCt value. The fold changes in expression level relative to control were expressed as 2−ΔΔCt. Barley actin gene (AY145451, Fw-5′-GACTCTGGTGATGGTGTCAGC-3′, Rv-5′-GGCTGGAA-GAGGACCTCA-3′) was used as an internal control.
Statistical analysis
All data are presented as the mean values for each treatment. Each physiological or histochemical analysis included six replicate samples. An analysis of variance (ANOVA) was conducted to determine differences among treatments. Significance of differences between Tibetan wild and cultivated barley was evaluated using LSD multiple range tests (P < 0.05) in SAS 9.2 statistical software (SAS Institute Inc., Cary, NC, USA). Origin Pro version 8.0 (Origin lab corporation, Wellesley Hills, Wellesley, MA, USA) was used to prepare graphs.
Results
Root hair morphology of four barley genotypes
Effects of pH, PEG-induced drought and Al stresses showed a significant variation of root hair morphology (Fig. 1). Root hair length and number (data not shown) significantly increased in XZ5 under low pH and PEG stress with a higher increase at pH 6.0 + PEG, although root hairs of XZ29 and Tadmor had a degree of growth which was smaller and shorter than that of controls, particularly of cv. Dayton. In comparison with controls, root hair length and number were less affected in ZX29 and XZ5 than in other genotypes under Al and combined stresses (pH 4.0 + PEG + Al).

Root hair morphology of cultivated and Tibetan wild-barley genotypes. Roots were affected by pH, PEG, only Al and combined (pH 4.0 + PEG + Al) stresses at 7 days after treatment. For details of experimental procedures, see “Materials and methods”. Scale bars 1 mm
Root length, biomass accumulation and Al content
Effects of single and combined stresses of PEG and Al on root length, plant dry weight and Al concentration are shown in Fig. 2 and Supplementary Table S1. Exposure of plants to low pH (4.0) inhibited plant growth, which led to a significant decrease in dry weight compared to pH 6.0 case (control); i.e. decrease by 85.57 % in XZ29, 86.07 % in XZ5, 90.75 % in Dayton and 75.37 % in Tadmor. PEG stress significantly decreased dry weight and root length compared to controls. Moreover, the reduction was more pronounced for pH 4.0 + PEG than for pH 6.0 + PEG, which indicates that the PEG effect is more severe at lower pH. Addition of Al reduced dry weight of four genotypes relative to those at pH 4.0 without Al. When plants were exposed to a combined stress of PEG and Al, they displayed a further reduction in dry weight. Among the four genotypes, the highest dry weight and root length were recorded for XZ29 and XZ5 with the pH 4.0 + PEG + Al treatment. For example, under combined stress (pH 4.0 + PEG + Al), dry weight/root length decreased by 28.56/50.41 % in XZ29, 27.12/48.33 % in XZ5, 38.93/56.33 % in Dayton and 36.57/58.72 % in Tadmor compared to controls (Supplementary Table S1).

Root length, dry weight and root Al content of Tibetan wild and cultivated barley genotypes. Barley plants were affected by pH, PEG, only Al and combined (pH 4.0 + PEG + Al) stresses at 7 days after treatment. Error bars represent SD values (n = 6). Different letters indicate significant differences among the genotypes according to Duncan’s multiple range tests with P < 0.05
Compared with controls, no differences were found in the Al content in roots of the four barley genotypes at lower pH (4.0) and PEG stress. Aluminum stress resulted in a strong increase in Al content in roots of all genotypes, and the increase was highest in Tadmor. On the other hand, reduced Al uptake and accumulation in barley roots were observed when plants were treated with combined stress (pH 4.0 + PEG + Al) compared with only Al-treated plant. There was a significant genotypic difference: Tibetan wild-barley genotypes XZ5 and XZ29 had a lower Al content than cv. Dayton and Tadmor. Remarkably, both XZ5 and XZ29 exhibited 148.9/168.67 % and 92.34/112.14 % lower root Al content than the roots of Dayton/Tadmor, respectively, under combined stress (pH 4.0 + PEG + Al), and a similar trend was observed under Al stress alone (Supplementary Table S1).
Hydrogen peroxide (H2O2), superoxide radical (\(\text{O}_{2}^{\cdot -}\)) and callose content
The H2O2, \(\text{O}_{2}^{\cdot -}\) and callose contents in barley roots are shown in Fig. 3 and Supplementary Table S1. Overall, the low pH (4.0) stress increased H2O2, \(\text{O}_{2}^{\cdot -}\) and callose contents in all genotypes compared to pH 6.0 case (control). After addition of PEG to the growth medium, H2O2, \(\text{O}_{2}^{\cdot -}\) and callose contents in roots of all barley genotypes increased significantly. The H2O2, \(\text{O}_{2}^{\cdot -}\) and callose contents in roots of barley plants also significantly increased when plants were exposed to Al stress. However, when plants were exposed to combined stresses of Al and PEG, the H2O2, \(\text{O}_{2}^{\cdot -}\) and callose contents further increased relative to the low pH and PEG single stress but significantly decreased relative to Al stress. Under all stress treatments, the average percent increases in activities of H2O2, \(\text{O}_{2}^{\cdot -}\) contents among the four genotypes in descending order were as follows: XZ5 > XZ29 > Tadmor > Dayton (Fig. 3 and Supplementary Table S1). For instance, the average \(\text{O}_{2}^{\cdot -} /\text{H}_{2} \text{O}_{2}\) contents increased by 279.22/201.54 % in XZ29, 250.06/168.0 % in XZ5, 338.94/267.94 % in Dayton and 329.57/223.12 % in Tadmor compared to controls (Supplementary Table S1). Both wild genotypes (XZ29 and XZ5) exhibited a higher callose content than those of cultivated genotypes (Dayton and Tadmor) under Al alone and combined stress (pH 4.0 + PEG + Al).

H2O2, \(\text{O}_{2}^{\cdot -}\) and callose content of Tibetan wild and cultivated barley genotypes. Barley seedlings were exposed to pH, PEG and Al alone and combined (pH 4.0 + PEG + Al) stresses for 7 days. Error bars represent SD values (n = 6). Different letters indicate significant differences among the genotypes according to Duncan’s multiple range tests with P < 0.05
In addition, DAB and NBT staining showed an intensive production of reactive oxygen species (ROS) in roots under PEG and Al stress. Low pH did not result in obvious damage of root tips of all genotypes. Al stress alone caused increases in H2O2 (Supplemental Fig. S1a) and \(\text{O}_{2}^{\cdot -}\) (Supplemental Fig. S1b) in roots in the elongation zones, and XZ5 accumulated H2O2 in a less and smaller part of the elongation zone than the other 3 genotypes. The combined treatment of PEG and Al resulted in no visible increase of H2O2 and \(\text{O}_{2}^{\cdot -}\) staining relative to each single stress. Among the four genotypes, the two Tibetan wild barleys (XZ29 and XZ5) accumulated less H2O2 and \(\text{O}_{2}^{\cdot -}\) than cv. Tadmor and Dayton under PEG and Al stress.
The effect of Al and PEG stresses on barley roots was further examined by visualizing callose, which is a component of cell walls. Exposure of plants to pH 6.0, pH 4.0, pH 6.0 + PEG and pH 4.0 + PEG did not result in a distinct callose fluorescence in the root epidermis, yet the whole cross section exhibited an overall minor fluorescence. With PEG the signal was a bit stronger. However, callose accumulation in the vascular bundles of XZ29 and XZ5 was markedly higher compared to that in epidermal cells under Al stress alone (Fig. 4, pH 4.0 + Al). An increased fluorescence in the central vascular cylinder and vascular bundles in XZ5 and Dayton was observed under pH 4.0 + PEG + Al. However, in XZ29 and Tadmor, the whole cross section exhibited fluorescence, which could not be considered as a specific callose fluorescence of the vascular cylinder.

Callose deposition in barley roots. Barley seedlings were exposed to pH, PEG and Al alone and combined (pH 4.0 + PEG + Al) stresses for 7 days. Root tips (~0.5 cm) were fixed with 85 % ethanol, stained with 1 % aniline blue in 1 M glycine (pH 9.5), then cross-sectioned and examined under a fluorescence microscopy (Olympus BX61). An overall minor (unspecific) fluorescence is observed, with a slightly higher signal in the epidermal area after PEG treatment. Under Al treatment, the central cylinder and vascular bundles of XZ29 and XZ5 deliver fluorescence signals, XZ5 and Dayton after pH 4.0 + PEG + Al. Insets higher magnification of outer root tissue. Scale bar 1 mm
Organic-acid (OA) content
Citrate and malate were measured in root exudates of the plants at 7 days after PEG and Al treatments (Fig. 5 and Supplementary Table S2). With regard to exposure of plants to low pH (4.0), there was no significant difference among treatments and genotypes. However, addition of PEG to the solution increased both citrate and malate contents compared to controls. The Al stresses strongly increased contents of citrate and malate in all genotypes compared to controls. Yet, XZ29 showed the highest increase, and the increase value was in the order XZ29 > XZ5 > Dayton > Tadmor under Al stress relative to controls. Combined stresses of pH 4.0 + PEG + Al significantly decreased citrate and malate contents compared to Al stress alone while showing an increase relative to the case of only PEG stress. Both citrate and malate were less repressed in XZ5 (c.f. repressed by 57.17 and 57.44 % in XZ29, 40.99 and 49.34 % in XZ5, 101.48 and 74.29 % in Dayton, 211.5 and 66.78 % in Tadmor, respectively) than in the other three genotypes, as expressed by reduced proportion between combined stresses and only Al stress.

Citrate and malate secretions from roots of four barley genotypes. Barley seedlings were exposed to pH, PEG and Al alone and combined (pH 4.0 + PEG + Al) stresses for 7 days. Error bars represent SD values (n = 6). Different letters indicate significant differences among the genotypes according to Duncan’s multiple range tests with P < 0.05
Abscisic acid (ABA) and zeatin riboside (ZR) content and ACC oxidase from barley root
Effects of different treatments on abscisic acid (ABA), zeatin riboside (ZR) content and ACC oxidase of barley roots are presented in Fig. 6 and Supplementary Table S2. ABA content significantly increased when plants were exposed to low pH compared to control. Under PEG stress, significant increases in ABA were observed in all genotypes. Overall, a prominent effect of pH was found, which indicates that the PEG effect is more severe at lower pH of the cultivation solution. The Al stress alone significantly decreased ABA in all genotypes relative to pH 4.0 treatment without Al and PEG. Combined stresses of pH 4.0 + PEG + Al significantly reduced ABA compared to only PEG stress but increased relative to Al stress alone. Compared to the other three genotypes, XZ5 recorded the highest increase in ABA under PEG alone and pH 4.0 + PEG + Al stress. In context, ZR content increased significantly under only Al stress and combined stresses in all genotypes, when compared to controls, and the increase value was in the order Dayton > Tadmor > XZ29 > XZ5 (Fig. 6 and Supplementary Table S2). ACC oxidase significantly increased when plants were treated with Al alone or combined stresses of pH 4.0 + PEG + Al and this increase was highest under Al stress alone and with the order of Dayton > Tadmor > XZ29 > XZ5. However, under PEG stress treatments, the average decreases in ZR content and ACC oxidase among the four genotypes in the order was as follows: XZ5 > XZ29 > Tadmor > Dayton (Supplementary Table S2). For example, the average ZR content/ACC oxidase decreased by 59.92/52.34 % in XZ29, 54.39/46.51 % in XZ5, 83.20/86.42 % in Dayton and 65.40/58.75 % in Tadmor compared to controls (Supplementary Table S2).

ABA and ZR content, and ACC oxidase in roots of four barley genotypes. Barley seedlings were exposed to pH, PEG and Al alone and combined (pH 4.0 + PEG + Al) stresses for 7 days. Error bars represent SD values (n = 6). Different letters indicate significant differences among the genotypes according to Duncan’s multiple range tests with P < 0.05
The transcript levels of certain genes related to cell wall- and osmotic stress
As shown in Fig. 7, compared with pH 6.0 case (control), the lower pH (4.0) and PEG levels decreased the transcript levels of HvMATE in all genotypes. However, Al and pH 4.0 + PEG + Al treatments up-regulated HvMATE expression in all genotypes, with the highest expression in XZ29. Lower pH 4.0 and PEG significantly increased expression of the NCED gene, which is involved in ABA biosynthesis, in all genotypes (with a higher increase in XZ5) compared with pH 6.0 case (control). The PEG effect is more severe at lower pH because the reduction is more pronounced under pH 4.0 + PEG treatment than under pH 6.0 + PEG treatment. The Al treatment affected NCED, which was down-regulated in Tadmor and up-regulated in XZ29, but there was no effect in the other genotypes, which reversed pH 4.0 + PEG + Al enhanced expression of this gene in all genotypes except Dayton compared to the control (pH 6.0). Interestingly, transcript levels of SUS and KS-DHN significantly increased under PEG treatments in XZ5 compared to that of XZ29, Dayton and Tadmor. However, SUS and KS-DHN transcripts significantly decreased in all genotypes under Al conditions and increased under pH 4.0 + PEG + Al stress compared to the control (with a larger increase in XZ5).

Expression analysis on cell-wall- and osmotic-stress-associated genes of barley genotypes by qPCR. Effect of pH, only PEG, only Al, and combined (pH 4.0 + PEG + Al) stresses on HvMATE, NCED, SUS and KS-DHN expression in root tips of four barley genotypes at 7 days after treatment. Error bars represent SD values (n = 6). Different letters indicate significant differences among the genotypes according to Duncan’s multiple range tests with P < 0.05
Discussion
Drought can be exacerbated by subsoil Al toxicity, which reduces root elongation and restricts plant roots to explore acidic subsoil to absorb water, thus diminishing the ability to avoid drought stress. Normally, plants are exposed to combined stresses such as drought and aluminum (Yang et al. 2013); thus, understanding responses of plants to combined stresses is critical to provide a basis for developing strategies to maintain root growth and sustainable plant production. To study the influences of drought stress on Al uptake in root tips is difficult because we cannot discriminate whether Al or drought stress leads to changes in Al uptake and maintains or promotes the primary root elongation and root hair development. To overcome these drawbacks, Al stress alone was also applied in this study for comparison.
Higher root growth, dry weight and lower Al content
Wild barley has developed unique mechanisms to survive harsh environments; it has a wider genetic variation and much greater stress tolerance than cultivated barley. In the present study, Tibetan wild-barley genotypes XZ5 and XZ29 demonstrate high tolerance to drought and aluminum stresses, respectively, as our previous studies also confirmed (Zhao et al. 2010; Cai et al. 2013). Our results showed that, compared to cv. Dayton and Tadmor, the wild-barley genotypes XZ5 and XZ29 are comparatively tolerant to combined pH 4.0 + PEG + Al stresses and this tolerance is associated with higher root growth, dry weight and lower Al content in roots of these genotypes (Figs. 1, 2). Importantly, XZ5 shows a larger zone of root hair initiation and elongation than controls under PEG stress. These results indicate the potential of wild barley as a source of PEG/Al-tolerant alleles, and they can provide a wide genetic variation to cultivated barley.
The accumulation and content of callose
In higher plants, callose deposition can be induced by wounding, infection of pathogens, aluminum, abscisic acid and other physiological stresses (Chen and Kim 2004). It has been hypothesized that callose can be an induced defense response to biotic and abiotic stresses (Nishimura et al. 2003). In the present work, the treatments with different pH and different pH + PEG all showed a more general and weak callose fluorescence over the root tissue, in some cases epidermal cells seem to produce a higher signal, and in contrast to these observations the roots treated with Al or Al + PEG demonstrated a higher fluorescence signal of callose in the central cylinder and vascular bundles (except Tadmor) (Fig. 4). Such an enhancement of callose deposition in the central cylinder and vascular bundles would immobilize more Al ions to vascular bundles. Thus, it can be assumed that the capacity of immobilizing Al ions into the central cylinder and vascular bundles may be a common strategy for barley plants to cope with Al stress. Similarly, it has been observed in wheat that callose accumulation in root tissues was progressive with Al-exposure and the deposition occurred at inner cell layers (Silva et al. 2010, 2011). In contrast to Horst et al. (1997) we did not find an indication that Al sensitivity is related to callose accumulation in the root tips (Fig. 4). This might be due to the fact that the root parts analyzed (~5 cm) in the present study were much longer than the Al-sensitive part which is confined to the apical 3 mm (Ryan et al. 1993).
Lower xylem transport ability might be the reason of callose accumulation in the vascular bundle. Therefore, barley varieties with lower xylem transport capacity and feed-back regulating phloem transport in the developing roots might induce callose accumulation in the vascular bundle. However, the PEG treatment might also affect the accumulation of nutrients in roots. Studies are needed to resolve the contradiction on the interaction of nutrient accumulation and callose accumulation in both, epidermal cells and vascular bundle cells of barley genotypes with lower xylem transport capacity (e.g. XZ29 and XZ5). Our previous study showed that the individual and combined effects of drought and salinity increase callose content in XZ5 (Ahmed et al. 2015). One possible explanation is that callose could hinder the plants’ defense machinery against drought by increasing its water holding capacity, and it might be linked with higher water use efficiency in the wild genotype during the vegetative stage (Ahmed et al. 2013). Yet, this needs further verification and may shed new light on the biological function of callose in cell structure, water retention and cell protection in plants.
The production of ROS in barley roots
Aerobic biological metabolism produces reactive oxygen species (ROS) such as hydrogen peroxide (H2O2), superoxide radicals (\(\text{O}_{2}^{\cdot -}\)), hydroxyl radicals (OH−) and singlet oxygen (\({}^{1}\text{O}_{2}\)) (Rama Devi and Prasad 1998). ROS subsequently attack various biomolecules, including lipids, amino acids, proteins, nucleic acids, leading to cell death. In the present study, increased levels of ROS were found under PEG and Al stresses in the four barley genotypes, as indicated by H2O2 and \(\text{O}_{2}^{\cdot -}\) contents and accumulation. Tadmor and Dayton had higher H2O2 and \(\text{O}_{2}^{\cdot -}\) contents than XZ29 and XZ5 in all stress treatments, which indicates less oxidative damage in Tibetan wild barley than in the cultivated barley. This result suggests that harmful effects of PEG and Al toxicity on plants are probably exerted through production of ROS. As expected, combined stresses of PEG and Al caused a further increase in H2O2 and \(\text{O}_{2}^{\cdot -}\) contents compared to PEG treatment but showed a significant reduction relative to only Al treatment (Fig. 3). More importantly, our results show that PEG-induced drought stress significantly reduces Al uptake in barley plants under pH 4.0 + PEG + Al stress. This PEG 6000-caused exclusion of Al from root tips of barley could not be explained by a decrease in cell-wall negativity, which is determined based on the concentration of cell-wall pectins and its degree of methylation (Yang et al. 2013). Thus, we assume that PEG 6000 might be beneficial to decrease Al accumulation of barley, which (1) consequently reduced ROS production, when plants were grown under pH 4.0 + PEG + Al stress, which (2) might reduce Al-induced inhibition of primary root elongation and root hair development. Yang et al. (2010) found that PEG 6000 strongly blocked Al accumulation in root tips and therefore reduced toxic Al-induced inhibition of root elongation in common bean. Thus, the effects of simultaneous exposure to PEG and Al on ROS system of plant roots were understandable.
Tibetan wild barley secretes more citrate and malate
The secretion of organic acids from roots is considered one of the most important mechanisms for Al tolerance in plants (Ma et al. 2001). Malate and citrate efflux from root apices is a mechanism of Al3+ tolerance in many plant species. Malate exudation mechanism by wheat has been investigated most thoroughly (Kochian 1995) while citrate seems to be the most common organic acid anion exudated by Al-tolerant maize and snap bean (Barceló and Poschenrieder 2002). However, citrate efflux is facilitated by members of the MATE (multidrug and toxic compound exudation) family, which are localized at the plasma membrane of root cells. In the present study, Al stress and pH 4.0 + PEG + Al induced a higher citrate and malate secretion in Al-resistant XZ29 than in Dayton, XZ5 and Tadmor (Fig. 5). By analyzing the combined effect of short-term water-deficit stress and Al toxicity on citrate exudation from soybean roots, Nian et al. (2005) showed that individual stress mainly impeded root growth, but the combined stresses reduced both root growth and shoot growth.
The expression of citrate transporter HvMATE in barley roots
In the present study, expression of HvMATE was rather specific for Al stress compared to PEG stress. The HvMATE genes showed a significant up-regulation under Al and pH 4.0 + PEG + Al treatments in XZ29 (Fig. 7). The PEG treatment alone had no effect on the expression of HvMATE gene (Fig. 7). In barley (Hordeum vulgare) HvMATE expression in root apices correlates with Al-activated citrate exudation and Al resistance in a set of barley cultivars (Furukawa et al. 2007).
ABA accumulation and the expression of NCED
Al suppressed drought-induced ABA accumulation (Fig. 6) and the expression of NCED (Fig. 7) in root tips of barley genotypes. However, with PEG alone, ABA accumulation and expression of NCED were significantly enhanced in all genotypes (with a higher increase in XZ5). It has been reported that overexpression of NCED in tobacco (Nicotiana plumbaginifolia) and Arabidopsis increases endogenous ABA levels and improves drought tolerance (Iuchi et al. 2001; Qin and Zeevaart 2002).
ABA, ZR and ethylene are integrated in maintaining root growth and the transcript levels of drought-stimulated genes
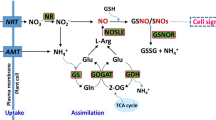
Drought stress induces the synthesis of ABA in roots, and ABA is transported to shoots through the transpiration stream in the xylem, which causes stomatal closure and reduces transpiration (Hartung et al. 2002). ABA may improve drought tolerance of barley by osmoregulation as a consequence of increasing the contents of proline and soluble sugar (Nayyar and Walia 2003; de Souza et al. 2014). Some previous studies have suggested that ABA accumulation restricts ethylene synthesis in drought-stressed roots, and ethylene can also interact with ZR [a biologically active form of cytokinin (CK) in root tips] production and transport in root tip (Sharp 2002; Souter et al. 2002; Růžička et al. 2007). Our results suggest that interactions among ABA, ZR and ethylene are integrated in maintaining root growth under drought and combined stresses. The accumulation of ABA in roots mainly toward the root apex is necessary to maintain the primary root elongation at low water potentials (Saab et al. 1992; Yamaguchi and Sharp 2010). However, Al treatment triggers a cross-talk among phytohormones, which reduces drought resistance of root apex and enhances the inhibition of root elongation of barley, as shown in Fig. 8.

Graphic depiction based on the present findings. The potential regulatory mechanisms involved in acclimation to drought of root apices of Tibetan wild barley and how Al stress interferes with this acclimation is shown. Thick arrows indicate up- and down-regulated changes. Thick green arrows mark the effect of the sole drought treatment, and thick red arrows mark relative changes under combined drought and Al stress compared with only drought treatment. Thin dashed arrows indicate the potential connections
Drought and Al also inhibit both primary and root hair growths of barley in the following order: Dayton > Tadmor > XZ29 > XZ5 (Fig. 1). This observation is consistent with the increasing content of ZR (Fig. 6). Massot et al. (1994) were the first to report that excess Al increased endogenous levels of ZR in roots of common bean. Al further suppresses drought-stimulated gene expression of SUS and KS-DHN in root tips of barley (Fig. 7). However, these gene expression levels significantly increase in all genotypes, with the largest increase is found in XZ5 under PEG and pH 4.0 + PEG + Al. It has been reported that the expression of these genes plays crucial roles in plant cellular adaptation to drought (Colmenero-Flores et al. 1999; Wang et al. 2000; Bartels and Sunkar 2005; Yang et al. 2011), that they can be induced by ABA and may maintain root hair growth. Saftner and Wyse (1984) observed that ABA increased sucrose uptake in the roots of sugar beet (Beta vulgaris), and the ABA-insensitive (abi8) mutant showed a strong reduction of the expression of SUS in Arabidopsis (Brocard-Gifford et al. 2004). Therefore, it appears that the suppression of ABA-dependent drought-induced gene regulation by Al may lead to an aggravated inhibition of root elongation under drought, as schematically depicted in Fig. 8. We therefore propose that varieties with significant modulation of expression of these key genes (irrespective of direction of change) can be tentatively considered as tolerant. Yang et al. (2012) also proposed that Al interfered with regulatory pathways that were involved in ABA signal transduction and crosstalk with other hormones which were important, particularly during drought stress.
Conclusions
Collectively, our results showed that wild-barley genotypes are comparatively tolerant to pH 4.0 + PEG + Al treatment and this tolerance is linked to the higher increase in ABA and callose content, more citrate and malate secretion/efflux, and less Al, ZR and ethylene accumulation in the roots. The less accumulation of H2O2 and \(\text{O}_{2}^{\cdot -}\) adds additional evidence for the higher tolerance to pH 4.0 + PEG + Al stress in XZ29 and XZ5. Our results also provide circumstantial evidence that in Tibetan wild barley drought alleviates Al injury in root tips based on drought-stimulated gene expression of NCED, SUS, and KS-DHN and higher expression of an HvMATE gene and ABA signal cross-talk with other phytohormones which are necessary to maintain root growth under drought stress and pH 4.0 + PEG + Al. However, there is limited knowledge on the interaction of acid/Al and drought stresses on crop yield. Further studies using advanced molecular techniques and mutant analysis are required to better understand detailed mechanisms of the response of Tibetan wild barley to combined abiotic stresses such as drought and Al.
Author contribution statement
Conceived and designed the experiments: IMA FBW. Performed the experiments: IMA UAN FBC HXY. Analyzed the data: IMA UAN FBC HXY. Wrote the paper: IMA FBW GPZ. Revising: IMA UAN FBC HXY FBW GPZ.
Abbreviations
- ACCO:
-
1-Aminocyclopropane-1-carboxylic acid oxidase
- KS-DHN:
-
KS-type dehydrin
- MATE:
-
Multidrug and toxic compound exudation
- \(\text{O}_{2}^{\cdot -}\) :
-
Super oxide
- NCED:
-
9-cis-epoxycarotenoid dioxygenase
- PEG:
-
Polyethylene glycol
- SUS:
-
Sucrose synthase 2
- ZR:
-
Zeatin riboside
References
Ahmed IM, Dai H, Zheng W, Cao F, Zhang GP, Sun D, Wu FB (2013) Genotypic differences in physiological characteristics in the tolerance to drought and salinity combined stress between Tibetan wild and cultivated barley. Plant Physiol Biochem 63:49–60
Ahmed IM, Nadira UA, Bibi N, Cao F, He X, Zhang GP, Wu FB (2015) Secondary metabolism and antioxidants are involved in the tolerance to drought and salinity, separately and combined, in Tibetan wild barley. Environ Exp Bot 111:1–12
Arroyave C, Tolrà R, Thuy T, Barceló J, Poschenrieder C (2013) Differential aluminum resistance in Brachiaria species. Environ Exp Bot 89:11–18
Barceló J, Poschenrieder C (2002) Fast root growth responses, root exudates, and internal detoxification as clues to the mechanisms of aluminium toxicity and resistance: a review. Environ Exp Bot 48:75–92
Bartels D, Sunkar R (2005) Drought and salt tolerance in plants. Crit Rev Plant Sci 24:23–58
Beebe S, Ramirez J, Jarvis A, Rao IM, Mosquera G, Bueno JM, Blair MW (2011) Genetic improvement of common beans and the challenges of climate change. In: Yadav SS, Redden RJ, Hatfield JL, Lotze-Campen H, Hall AE (eds) Crop adaptation to climate change. Wiley-Blackwell, Oxford, pp 356–369
Brocard-Gifford I, Lynch TJ, Garcia ME, Malhotra B, Finkelstein RR (2004) The Arabidopsis thaliana abscisic acid-insensitive 8 encodes a novel protein mediating abscisic acid and sugar responses essential for growth. Plant Cell 16:406–421
Cai S, Wu D, Jabeen Z, Huang Y, Huang Y, Zhang GP (2013) Genome-wide association analysis of aluminum tolerance in cultivated and Tibetan wild barley. PLoS ONE 8:e69776
Chaves MM, Oliveira M (2004) Mechanisms underlying plant resilience to water deficits: prospects for water-saving agriculture. J Exp Bot 55:2365–2384
Chen X-Y, Kim J-Y (2004) Callose synthesis in higher plants. Plant Signal Behav 4:489–492
Colmenero-Flores JM, Moreno LP, Smith CE, Covarrubias AA (1999) Pvlea-18, a member of a new late-embryogenesis-abundant protein family that accumulates during water stress and in the growing regions of well-irrigated bean seedlings. Plant Physiol 120:93–104
de Souza TC, Magalhães PC, de Castro EM, Carneiro NP, Padilha FA, Júnior CCG (2014) ABA application to maize hybrids contrasting for drought tolerance: changes in water parameters and in antioxidant enzyme activity. Plant Growth Regul 73:205–217
Delhaize E, Ryan PR (1995) Aluminum toxicity and tolerance in plants. Plant Physiol 107:315–321
Forster B, Ellis R, Moir J, Talame V, Sanguineti M, Tuberosa R, This D, Teulat-Merah B, Ahmed I, Mariy S (2004) Genotype and phenotype associations with drought tolerance in barley tested in North Africa. Ann Appl Biol 144:157–168
Furukawa J, Yamaji N, Wang H, Mitani N, Murata Y, Sato K, Katsuhara M, Takeda K, Ma JF (2007) An aluminum-activated citrate transporter in barley. Plant Cell Physiol 48:1081–1091
Goldman I, Carter T, Patterson R (1989) A detrimental interaction of subsoil aluminum and drought stress on the leaf water status of soybean. Agron J 81:461–463
Hartung W, Sauter A, Hose E (2002) Abscisic acid in the xylem: where does it come from, where does it go to? J Exp Bot 53:27–32
Horst WJ, Püschel A-K, Schmohl N (1997) Induction of callose formation is a sensitive marker for genotypic aluminium sensitivity in maize. Plant Soil 192:23–30
Ishikawa H, Evans ML (1993) The role of the distal elongation zone in the response of maize roots to auxin and gravity. Plant Physiol 102:1203–1210
Iuchi S, Kobayashi M, Taji T, Naramoto M, Seki M, Kato T, Tabata S, Kakubari Y, Yamaguchi-Shinozaki K, Shinozaki K (2001) Regulation of drought tolerance by gene manipulation of 9-cis-epoxycarotenoid dioxygenase, a key enzyme in abscisic acid biosynthesis in Arabidopsis. Plant J 27:325–333
Jiang M, Zhang J (2002) Water stress-induced abscisic acid accumulation triggers the increased generation of reactive oxygen species and up-regulates the activities of antioxidant enzymes in maize leaves. J Exp Bot 53:2401–2410
Kochian LV (1995) Cellular mechanisms of aluminum toxicity and resistance in plants. Annu Rev Plant Physiol Plant Mol Biol 46:237–260
Kochian LV, Hoekenga OA, Piñeros MA (2004) How do crop plants tolerate acid soils? Mechanisms of aluminum tolerance and phosphorous efficiency. Annu Rev Plant Biol 55:459–493
Köhle H, Jeblick W, Poten F, Blaschek W, Kauss H (1985) Chitosan-elicited callose synthesis in soybean cells as a Ca2+-dependent process. Plant Physiol 77:544–551
Kollmeier M, Felle HH, Horst WJ (2013) Genotypical differences in aluminum resistance of maize are expressed in the distal part of the transition zone. Is reduced basipetal auxin flow involved in inhibition of root elongation by aluminum? Plant Physiol 122:945–956
Liu X, Hua X, Guo J, Qi D, Wang L, Liu Z, Jin Z, Chen S, Liu G (2013) Enhanced tolerance to drought stress in transgenic tobacco plants overexpressing VTE1 for increased tocopherol production from Arabidopsis thaliana. Biotech Lett 30:1275–1280
Livak KJ, Schmittgen TD (2001) Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 25:402–408
Ma JF, Ryan PR, Delhaize E (2001) Aluminium tolerance in plants and the complexing role of organic acids. Trends Plant Sci 6:273–278
Massot N, Poschenrieder C, Barcelo J (1994) Aluminium-induced increase of zeatin riboside and dihydrozeatin riboside in Phaseolus vulgaris L. cultivars. J Plant Nutr 17:255–265
Mittler R (2006) Abiotic stress, the field environment and stress combination. Trends Plant Sci 11:15–19
Müller M, Munné-Bosch S (2011) Rapid and sensitive hormonal profiling of complex plant samples by liquid chromatography coupled to electrospray ionization tandem mass spectrometry. Plant Methods 7:37
Nayyar H, Walia D (2003) Water stress induced proline accumulation in contrasting wheat genotypes as affected by calcium and abscisic acid. Biol Plant 46:275–279
Nian H, Yang Z, Huang H, Yan X, Matsumoto H (2005) Combined effect of short-term water deficit stress and aluminum toxicity on citrate secretion from soybean roots. J Plant Nutr 27:1281–1293
Nishimura MT, Stein M, Hou B-H, Vogel JP, Edwards H, Somerville SC (2003) Loss of a callose synthase results in salicylic acid-dependent disease resistance. Science 301:969–972
Qin X, Zeevaart JA (2002) Overexpression of a 9-cis-epoxycarotenoid dioxygenase gene in Nicotiana plumbaginifolia increases abscisic acid and phaseic acid levels and enhances drought tolerance. Plant Physiol 128:544–551
Rama Devi S, Prasad M (1998) Copper toxicity in Ceratophyllum demersum L. (Coontail), a free floating macrophyte: response of antioxidant enzymes and antioxidants. Plant Sci 138:157–165
Růžička K, Ljung K, Vanneste S, Podhorská R, Beeckman T, Friml J, Benková E (2007) Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 19:2197–2212
Ryan PR, Ditomaso JM, Kochian LV (1993) Aluminium toxicity in roots: an investigation of spatial sensitivity and the role of the root cap. J Exp Bot 44:437–446
Saab IN, Sharp RE, Pritchard J (1992) Effect of inhibition of abscisic acid accumulation on the spatial distribution of elongation in the primary root and mesocotyl of maize at low water potentials. Plant Physiol 99:26–33
Saftner RA, Wyse RE (1984) Effect of plant hormones on sucrose uptake by sugar beet root tissue discs. Plant Physiol 74:951–955
Serraj R, Sinclair T (2002) Osmolyte accumulation can it really help increase crop yield under drought conditions? Plant, Cell Environ 25:333–341
Sharp R (2002) Interaction with ethylene: changing views on the role of abscisic acid in root and shoot growth responses to water stress. Plant, Cell Environ 25:211–222
Sharp RE, Poroyko V, Hejlek LG, Spollen WG, Springer GK, Bohnert HJ, Nguyen HT (2004) Root growth maintenance during water deficits: physiology to functional genomics. J Exp Bot 55:2343–2351
Shimazaki Y, Ookawa T, Hirasawa T (2005) The root tip and accelerating region suppress elongation of the decelerating region without any effects on cell turgor in primary roots of maize under water stress. Plant Physiol 139:458–465
Silva S, Pinto-Carnide O, Martins-Lopes P, Matos M, Guedes-Pinto H, Santos C (2010) Differential aluminium changes on nutrient accumulation and root differentiation in an Al sensitive vs. tolerant wheat. Environ Exp Bot 68:91–98
Silva S, Santos C, Matos M, Pinto-Carnide O (2011) Al toxicity mechanisms in tolerant and sensitive rye genotypes. Environ Exp Bot 75:89–97
Souter M, Topping J, Pullen M, Friml J, Palme K, Hackett R, Grierson D, Lindsey K (2002) hydra mutants of Arabidopsis are defective in sterol profiles and auxin and ethylene signaling. Plant Cell 14:1017–1031
Staß A, Horst W (2009) Callose in abiotic stress. In: Bacic A, Fincher GB, Stone BA (eds) Chemistry, biochemistry and biology of (1 → 3)-β-glucans and related polysaccharides. Academic Press, New York, pp 499–524
Wang HL, Lee PD, Chen WL, Huang DJ, Su JC (2000) Osmotic stress-induced changes of sucrose metabolism in cultured sweet potato cells. J Exp Bot 51:1991–1999
Wang J, Raman H, Zhou M, Ryan PR, Delhaize E, Hebb DM, Coombes N, Mendham N (2007) High-resolution mapping of the Alp locus and identification of a candidate gene HvMATE controlling aluminium tolerance in barley (Hordeum vulgare L.). Theor Appl Genet 115:265–276
Willekens H, Chamnongpol S, Davey M, Schraudner M, Langebartels C, Van Montagu M, Inzé D, Van Camp W (1997) Catalase is a sink for H2O2 and is indispensable for stress defence in C3 plants. EMBO J 16:4806–4816
Wu FB, Zhang GP, Dominy P (2003) Four barley genotypes respond differently to cadmium: lipid peroxidation and activities of antioxidant capacity. Environ Exp Bot 50:67–78
Yamaguchi M, Sharp RE (2010) Complexity and coordination of root growth at low water potentials: recent advances from transcriptomic and proteomic analyses. Plant, Cell Environ 33:590–603
Yang Z-B, Eticha D, Rao IM, Horst WJ (2010) Alteration of cell-wall porosity is involved in osmotic stress-induced enhancement of aluminium resistance in common bean (Phaseolus vulgaris L.). J Exp Bot 61:3245–3258
Yang ZB, Eticha D, Rotter B, Rao IM, Horst WJ (2011) Physiological and molecular analysis of polyethylene glycol-induced reduction of aluminium accumulation in the root tips of common bean (Phaseolus vulgaris L.). New Phytol 192:99–113
Yang Z-B, Eticha D, Albacete A, Rao IM, Roitsch T, Horst WJ (2012) Physiological and molecular analysis of the interaction between aluminium toxicity and drought stress in common bean (Phaseolus vulgaris L.). J Exp Bot 63:3109–3125
Yang ZB, Rao IM, Horst WJ (2013) Interaction of aluminium and drought stress on root growth and crop yield on acid soils. Plant Soil 372:3–25
Zeng F, Wu X, Qiu B, Wu FB, Jiang L, Zhang GP (2014) Physiological and proteomic alterations in rice (Oryza sativa L.) seedlings under hexavalent chromium stress. Planta 239:91–108
Zhao Z, Ma JF, Sato K, Takeda K (2003) Differential Al resistance and citrate secretion in barley (Hordeum vulgare L.). Planta 217:794–800
Zhao J, Sun H, Dai H, Zhang GP, Wu FB (2010) Difference in response to drought stress among Tibet wild barley genotypes. Euphytica 172:395–403
Zheng SJ, Yang JL, He YF, Yu XH, Zhang L, You JF, Shen RF, Matsumoto H (2005) Immobilization of aluminum with phosphorus in roots is associated with high aluminum resistance in buckwheat. Plant Physiol 138:297–303
Acknowledgments
The project was supported by National Natural Science Foundation of China (31171488), National 863 Program (2012AA101105), Jiangsu Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, China, and the Key Research Foundation of Science and Technology Department of Zhejiang Province of China (2012C12902-2), and the 56th China Postdoctoral Scientific Research Foundation (517000-X91408). We appreciate Ms Mei Li from the Analysis Center of Agrobiology and Environmental Sciences of Zhejiang University, for her kind help with the experiment.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no competing interests.
Electronic supplementary material
Below is the link to the electronic supplementary material.
425_2015_2442_MOESM2_ESM.tif
Supplemental Fig. S1 Histochemical detection of H2O2 (a) and \(\text{O}_{2}^{\cdot -}\) (b) in barley roots. Barley seedlings were exposed to pH, PEG and Al alone and combined (pH 4.0 + PEG + Al) stresses for 7 days. For details of experimental procedures, see Materials and methods. Scale bar 1 mm (TIFF 2115 kb)
Rights and permissions
About this article

Cite this article
Ahmed, I.M., Nadira, U.A., Cao, F. et al. Physiological and molecular analysis on root growth associated with the tolerance to aluminum and drought individual and combined in Tibetan wild and cultivated barley. Planta 243, 973–985 (2016). https://doi.org/10.1007/s00425-015-2442-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00425-015-2442-x