
Abstract
Research in movement science aims at unravelling mechanisms and designing methods for restoring and maximizing human functional capacity, and many techniques provide access to neural adjustments (acute changes) or long-term adaptations (chronic changes) underlying changes in movement capabilities. First described by Paul Hoffmann over a century ago, when an electrical stimulus is applied to a peripheral nerve, this causes action potentials in afferent axons, primarily the Ia afferents of the muscle spindles, which recruit homonymous motor neurons, thereby causing an electromyographic response known as the Hoffmann (H) reflex. This technique is a valuable tool in the study of the neuromuscular function in humans and has provided relevant information in the neural control of movement. The large use of the H reflex in motor control research on humans relies in part to its relative simplicity. However, such simplicity masks subtleties that require rigorous experimental protocols and careful data interpretation. After highlighting basic properties and methodological aspects that should be considered for the correct use of the H-reflex technique, this brief narrative review discusses the purpose of the H reflex and emphasizes its use as a tool to assess the effectiveness of Ia afferents in discharging motor neurones. The review also aims to reconsider the link between H-reflex modulation and Ia presynaptic inhibition, the use of the H-reflex technique in motor control studies, and the effects of ageing. These aspects are summarized as recommendations for the use of the H reflex in motor control research on humans.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The abundant literature on the neural substrates of motor control in humans highlights a fascination for the beauty and complexity of the machinery that allows to perform an impressive range of movements. It also emphasises the tremendous effort made to understand the rules governing motor control in order to improve neuromuscular performance, to prevent the debilitating effects of age, and to counteract neuromuscular pathologies. Many techniques provide access to neural adjustments (acute changes) or long-term adaptations (chronic changes) underlying changes in movement capabilities. For example, electroencephalography, functional magnetic resonance imaging (fMRI) and magnetoencephalography (MEG) have provided fundamental information on brain activity and plasticity (Hallett et al. 2021). However, control strategies are specific to the task performed and experimental approaches must not constrain the movement of interest. Otherwise, the outcomes will have limited impact in our knowledge of how the human body works in everyday life, including occupational and recreational settings. This can also impede the development of interventions aimed at restoring and maximizing human functional capacity and well-being across the lifespan.
In this context, one valuable approach is the Hoffmann (H) reflex, which has a high sensitivity to posture (Baudry et al. 2015; Katz et al. 1988; Koceja et al. 1993), contraction modality (Burke et al. 1989; Duchateau and Baudry 2014; Gottlieb and Agarwal 1973; Stein et al. 2007), muscle stretching (Budini et al. 2018; Datoussaid et al. 2021) and massage (Behm et al. 2013), fatigability (Baudry et al. 2011; Duchateau and Hainaut 1993; Klass et al. 2008) and age (Baudry et al. 2010; Kido et al. 2004; Scaglioni et al. 2003). The relative simplicity of the H-reflex technique makes it an attractive tool for research. However, its simplicity masks subtleties that require rigorous experimental protocols and cautious data interpretation. Indeed, the apparent ease of the technique can lead to underestimate the underlying neurophysiology of the measurements and subsequently bias the outcomes. Several reviews have detailed the advantages and drawbacks of this technique (Burke 2016; Grosprêtre and Martin 2012; Knikou 2008; Pierrot-Deseilligny and Burke 2012; Pierrot-Deseilligny and Mazevet 2000; Schieppati 1987; Zehr 2002), with different perspectives ranging from purely methodological considerations to its use in clinical routine. Nonetheless, several aspects deserve additional attention to improve the use of the H-reflex technique in motor control research.
The main objective of this brief narrative review is to provide insights into several features of the H-reflex technique that should be considered for its appropriate use and interpretation. After an overview of the basic properties of the H reflex, we discuss its underlying neurophysiology in the context of motor control research and emphasize its use as a tool to assess the effectiveness of Ia afferents in evoking an action potential in motor neurones. Then, we debate the link between H-reflex modulation and Ia presynaptic inhibition, between the H-reflex modulation and motor behaviour, and adaptations with healthy ageing. To clarify the meaning of the main terms used in this brief review, a glossary is provided in Table 1.
The H-reflex technique
Basic properties of the H reflex
The H reflex is an electromyographic response triggered by the electrical stimulation of muscle spindle afferents, mainly Ia afferents, which recruit spinal motor neurones leading to a muscle activation. It was initially described by the German physiologist Paul Hoffmann more than one century ago (1918), and later coined the Hoffmann reflex by Magladery and McDougal (1950). The electrical stimulation of a nerve trunk with 1-ms pulse of sufficient current intensity can induce an action potential in several Ia afferent axons, which project onto α-motor neurones. If the spatial summation of the excitatory postsynaptic potentials (EPSPs) induced by the afferent volley reaches the firing threshold of a motor neuron, the action potential thus generated will be transmitted to the neuromuscular junction of the muscle fibres innervated by this neuron. This will excite the muscle fibres, and the resulting electromyographic response is the H reflex. To note, the greater excitability of Ia afferent axons compared with motor axons when using long pulse durations (1 ms) and low intensity is mainly attributable to a longer strength-duration time constant and lower rheobase, likely reflecting a greater numbers of persistent non-inactivating Na+ channels (Bostock and Rothwell 1997; Shimatani et al. 2018). The H reflex can be elicited in any muscle as long as its peripheral nerve is accessible to electrical stimulation. In practice, the soleus (SOL) and flexor carpi radialis (FCR) are the most studied muscles because the H reflex can be elicited at rest (Zehr 2002). In other muscles, the EPSPs induced by Ia afferent input are not sufficient to depolarise the motor neurone membrane to the firing threshold in the resting state. However, the H reflex can be evoked in these muscles during a voluntary contraction as the excitatory drive delivered by descending pathways increases the number of motor neurones within the subliminal fringe. This enhances the probability that the EPSPs evoked by Ia afferents input exceed the firing threshold in a greater number for motor neurones (Buller et al. 1980; Upton et al. 1971).
The recruitment curve
A classical approach when using the H reflex is to record the relation between the stimulus intensity (input) and the amplitude of the H reflex (output). This input–output relation is known as the H-reflex recruitment curve, which is described as an inverted U-shape with a peak value corresponding to the maximal amplitude of the H reflex (Hmax) (Fig. 1).

H-reflex and M-wave recruitment curves recorded in the soleus of one healthy adult at rest. A Consecutive traces represent the electromyographic responses evoked in soleus by the electrical stimuli (1-ms duration) applied to the tibial nerve at different current intensities. The pink and brown sections represent the ascending limb of the H-reflex and M-wave recruitment curve, respectively. The bold brown and pink traces represent Hmax and Mmax, respectively. B H-reflex (pink) and M-wave (brown) recruitment curves computed from traces illustrated in A. The amplitude (y axis) and stimulus intensity (x axis) are expressed as % of the maximal amplitude of the M wave (Mmax) and of the intensity associated with the Mmax, respectively
To understand the details of this relation, it is essential to consider the processes that link the stimulus intensity to H-reflex amplitude. It has been shown in cats that most Ia afferents project to each motor neurone of the homonymous muscle (Lucas et al. 1984), and that one motor neurone receives projections from most Ia afferents (Mendell and Henneman 1968). Such a distribution of Ia afferent projections increases the probability of spatial summation when stimulating afferent axons. According to the size principle based on the relation between the motor neurone input resistance and effective synaptic current (Henneman and Mendell 1981), and the possible greater density of Ia afferent synaptic inputs to small compared with large motor neurones (Burke 1968; Heckman and Binder 1988), excitatory inputs from Ia afferents evoke large EPSPs in low-threshold motor units and smaller EPSPs in higher threshold motor units (Burke et al. 1976; Burke 1968; Eccles et al. 1957). This implies that the motor units involved in the H reflex are recruited in order of increasing size (Aimonetti et al. 2000; Awiszus and Feistner 1993; Buller et al. 1980; Schmied et al. 1997), with small amplitude H-reflexes reflecting the recruitment of low-threshold (small) motor units, and larger amplitude H-reflexes corresponding to the recruitment of low- and higher threshold motor units. When the stimulus is increased, more afferents are activated, which recruits more motor neurones and, thereby, increases H-reflex amplitude (ascending limb of the H-reflex recruitment curve; Fig. 1B). The slope of the ascending limb of the recruitment curve reflects the rate of MN recruitment as a function of the increase in Ia input to MN pool, and is generally used to assess the reflex gain (Baudry et al. 2014; Funase et al. 1994; Penzer et al. 2015). However, the increase in the stimulus intensity leads to trigger action potentials in motor axons that emerge as a short-latency electromyography (EMG) response known as the compound muscle action potential (M wave). The action potential evoked in motor axons is also conveyed antidromically to the motor neurone with consequences for the H reflex. Importantly, H-reflex amplitude should be within the ascending limb of the recruitment curve to be correctly interpreted. This is because the sensitivity of the reflex to excitatory and inhibitory inputs depends on the number of motor neurones within the subliminal fringe and the last recruited motor neurones. Within the ascending limb, these motor neurones are likely not compromised by an antidromic volley. To note, the duration of the response can provide an indirect estimate of the temporal dispersion of action potentials reaching the motor neurone pool (Kiernan et al. 1996; Olney et al. 1987). Due to the range of axonal diameter and the relation between conduction velocity and axonal diameter (Waxman 1980), propagation velocities of action potentials vary across axons, causing action potentials to reach the motor neurone pool with different delays. In addition to influencing the summation of the EPSPs at the spinal level, it also increases the delay between the first and the last motor units activated (Burke et al. 1984). However, because the last recruited motor units have greater conduction velocity, the dispersion at the muscle level is minimal (1–2 ms) (Burke et al. 1984). Temporal dispersion increases with the length of the nerve path and demyelination, which may reduce the amplitude of the evoked potential (H reflex or M wave) (Kimura et al. 1988; Olney et al. 1987; Schulte-Mattler et al. 1999). However, H-reflex duration, as the M wave, is also influenced by the muscle fibre conduction velocity (Rodriguez-Falces et al. 2015). Therefore, differences or changes in H-reflex duration should be interpreted with cautious, depending on the experimental conditions and the population characteristics.
After the H reflex reaches its maximal amplitude, an increase in current intensity is accompanied by a decrease in the H-reflex amplitude (descending limb of the H-reflex recruitment curve; Fig. 1B) and an increase in M-wave amplitude up to its maximal value (Mmax). The descending limb of the H-reflex recruitment curve reflects, in part, an increase in the antidromic volley that blocks the orthodromic propagation of action potentials in motor axons evoked by Ia afferent inputs (Pierrot-Deseilligny and Mazevet 2000). At low intensities, low-threshold motor units are reflexively recruited while the stimulus could only depolarise, if any, large motor axons. Consequently, a collision between orthodromic and antidromic volleys should not occur as the axons conveying the orthodromic volley (H reflex) are not those conducting an antidromic volley. At greater current intensities, the motor neurones recruited by the afferent volley can have their axons also directly depolarised by the stimulus, which reduces the contribution of these motor units to the H reflex due to collision between orthodromic and antidromic volleys. Accordingly, the Hmax may not reflect the maximal number of motor units recruited by the afferent volley, but the number of motor units recruited which are free of any antidromic volley. At the intensity associated with the Mmax, most motor axons are depolarised and no H reflex can be recorded due to a complete cancellation of the orthodromic volley by the antidromic one.
One issue that is often neglected in the assessment of the H-reflex recruitment curve is the associated activation of Ib afferents (from Golgi tendon organs). The Ib afferents project on inhibitory interneurones (non-reciprocal group I inhibitory interneurones) that induce postsynaptic inhibitory potentials in the motor neurones. Due to the longer synaptic delay (two synapses) in the Ib pathway, the evoked inhibitory postsynaptic potentials (IPSP) begin 0.5–1.0 ms after the onset of the Ia excitation (Pierrot-Deseilligny et al. 1981), and can therefore truncate the latter part of the Ia excitatory input. Although the group I volley (Ia and Ib) induced by the stimulation is dominated by the monosynaptic Ia volley, the Ib pathway likely limits the amplitude of the H-reflex (Marchand-Pauvert et al. 2002). Accordingly, the H reflex reflects a balance between Ia excitatory and Ib inhibitory inputs. Figure 2 summarises the main variables that can be measured from the H-reflex recruitment curve and their meaning.

Main variables used to characterise the H reflex. The top panel illustrates a typical recording of a H reflex in the soleus in resting condition with the indications to measure the H-reflex variables. The horizontal grey dotted line indicates the baseline of the EMG signal. The first deflection on the left (*) illustrates the stimulation artefact (truncated for the figure) indicating the time of the stimulation. Hmax maximal amplitude of the H reflex, AP action potential
Methodological considerations
To take full advantage of the information provided by the H reflex, it is necessary to consider how to record it. Some requirements have been detailed elsewhere and will not be developed here (Pierrot-Deseilligny and Mazevet 2000; Schieppati 1987), whereas others deserve additional attention.
Consistency of the stimulus intensity and normalisation
It is important to ensure that the position of the recording electrodes does not change and that the effectiveness of the stimulus remains constant. The gold standard procedure is to record the entire H-reflex and M-wave recruitment curves as they provide complete information on changes in H reflex and M wave (Baudry et al. 2015; Katz et al. 1988; Koceja et al. 1995). However, this requires time and conditions that are not always available. An alternative is to record the H reflex with a specific intensity, usually set to obtain an H-reflex amplitude of 50% Hmax. The rationale for this choice is that this amplitude allows observing a decrease or increase of the H reflex. However, a drawback of this approach is that, in many cases, a small M-wave could not be obtained for this H-reflex amplitude, making it impossible to control for the consistency of the stimulus intensity. It is therefore recommended to set a stimulus intensity that evokes a submaximal M wave preceding the H reflex, provided that this intensity evokes an H reflex on the ascending limb of the recruitment curve (Capaday and Stein 1986, 1987b; Datoussaid et al. 2021). The adjustment of the current intensity across experimental conditions to obtain a constant M-wave amplitude (typically around 5 or 10% of the Mmax amplitude) relies on the fact that the M-wave only depends on peripheral factors (numbers of motor axons recruited, position of the electrodes, muscle architecture, etc.). Consequently, a constant M-wave amplitude should ensure that the nerve trunk is stimulated identically across conditions, and that changes in H-reflex amplitude from one condition to another reflect modulation at the spinal level. However, changes in the propagation of action potentials from the point of stimulation onto the motor axon to the transverse tubules of muscle fibres may occur during the time course of an experiment (Crone et al. 1999). Because the Mmax, Hmax and the submaximal responses appear to be influenced similarly (Crone et al. 1999), a potential change in signal propagation should not limit the use of the M wave preceding the H reflex as a proxy for the consistency of the stimulus intensity. Nonetheless, adjusting the current intensity to maintain a constant M-wave amplitude across the experiment may involve small variations in the recruitment of Ia afferents. Another possible concern is differences in the change of the biophysical properties of the sensory (Ia afferents) and motor axons when investigating maximal voluntary contractions lasting 15 s or more as sensory afferents could be less sensitive to activity-dependent hyperpolarisation (Vagg et al. 1998). To circumvent this possible issue, an Mmax should also be recorded in similar conditions and timing to that of the H-reflex, with the current intensity adjusted when required to avoid corruption of the normalisation procedure. Additionally, as the Mmax provides an estimate of the response of the entire motor neurone pool, normalizing H-reflex amplitude to M-wave amplitude crudely reflects the proportion of the motor neurone pool that contributes to the H reflex. Noteworthy, the H reflex/Mmax ratio increases the reliability of the measure as shown by the robust inter-session reliability (ICC: 0.98) of this ratio reported for the soleus (Hoch and Krause 2009).
Background EMG
The H-reflex amplitude increases with the level of excitation of the motor neurone pool. For example, Funase and Miles (1999) reported a strong dependency of H-reflex amplitude to the level of background EMG activity at the time the afferent volley reaches the soleus motor neurone pool. The excitatory drive onto the motor neurone pool increases the number of motor neurones within the subliminal fringe (Capaday and Stein 1987a, b), and thereby enhances the probability that motor neurones discharge an action potential in response to the Ia afferent input. The same rationale applies when comparing voluntary contractions performed at different intensities. In other words, a given Ia afferent input causes motor neurones to discharge action potential in proportion to the size of the subliminal fringe, which increases with the level of motor neurone pool activity. In addition, the antidromic volley conveyed on some motor axons may be abolished during a voluntary contraction by the descending drive and thereby permit their involvement in the H-reflex response. Discharge rate and the trajectory of the after-hyperpolarisation potential can also influence the probability that motor neurones discharge action potentials in response to Ia afferent input (Jones and Bawa 1997). It is, therefore, necessary to control for the motor neurone pool activity when recording the H reflex. This is often done by measuring the background EMG activity of the homonymous muscle that precedes the stimulus (100- to 500-ms window), with the rationale that a constant value should reflect a similar excitability of the motor neurone pool to the afferent inputs.
H reflex variability
The H-reflex amplitude varies from trial-to-trial even when procedures used to control the stimulus intensity and the background EMG activity are applied (Funase and Miles 1999; Schieppati 1987). Based on recordings of the monosynaptic reflex in ventral root filaments, experiments in decerebrated and spinalised cats showed that the variability in the amplitude of the monosynaptic reflex is largely influenced by fluctuations in the membrane potential of the motor neurones (Gossard et al. 1994). As a consequence, one single H reflex is not sufficient to provide a reliable estimate of its amplitude. Instead, an average must be obtained from several H reflexes for each participant within the same experimental condition. Once averaged, the H-reflex amplitude (submaximal or maximal amplitude) is highly reliable in soleus and quadriceps at rest (Christie et al. 2004; Hopkins and Wagie 2003). The number of responses that should be included in average ranges from 3 to 10 (Christie et al. 2004; Hugon 1973; Jaberzadeh et al. 2004; Knikou 2008; Tucker et al. 2005); the fewer the number of required responses, the shorter the duration of the recording. However, in some experimental conditions, such as upright standing during which H-reflex amplitude varies with the direction of postural sway, a greater number of responses should be included in the average. In our experience (Abdelmoula et al. 2019; Baudry and Duchateau 2012, 2020; Johannsson et al. 2017) and that of others (Christie et al. 2004; Hopkins and Wagie 2003), 5–10 responses, are required to obtain a good estimate of H-reflex amplitude in different experimental conditions and muscles. For example, when recording H reflexes in the soleus of young adults during upright standing with a stimulus intensity that elicited an M-wave amplitude of 10% Mmax, the coefficient of variation for H-reflex amplitude decreased from 14 to 8% when 3 and 10 responses were recorded, respectively, without a change in the mean value (39% Mmax). Noteworthy, increasing the number of responses included in the average to 20 did not further decrease the coefficient of variation (9%).
If at least 10 s is required between successive stimuli to avoid homosynaptic depression for recordings in a resting muscle (Hultborn et al. 1996; Stein et al. 2007), the interval between successive stimuli can be reduced to 2 s without influencing H-reflex amplitude during voluntary contraction in the soleus (Stein et al. 2007). The decrease in homosynaptic depression may reflect the high discharge rates of Ia afferents during voluntary contractions (Macefield et al. 1993; Prochazka et al. 1977). As a consequence, the addition of one or two discharges induced by the stimulation should have little effect on the response of the motor neurone (Stein et al. 2007). Even when using a frequency of 3 Hz, no homosynaptic depression was observed in the tibialis anterior (Burke et al. 1989; Klass et al. 2011) and abductor pollicis brevis muscles (Abdelmoula et al. 2019; Burke et al. 1989). Moreover, Stein and colleagues (Stein et al. 2007) reported an absence of homosynaptic depression in the soleus during upright standing, suggesting that shorter inter-stimulus interval can be used to average several H reflexes. It is, therefore, easier to record 5–10 responses to obtain a reliable estimate of the H-reflex amplitude during voluntary contractions.
Evolving interpretation of the H reflex
The H reflex has been used for several decades in motor control studies focusing on spinal mechanisms. Although the understanding of the H-reflex has evolved with time, some aspects of the H-reflex technique remain underrated requiring additional refinement of its interpretation.
Previous and current understanding
Historically, the H reflex was regarded as a measure of the excitability of the motor neurone pool. However, after Frank and Fuortes (1957) reported the existence of presynaptic inhibition on the monosynaptic reflex (Frank and Fuortes 1957)—later confirmed by the exhaustive work of Eccles (for a review Willis 2006)—the H reflex cannot be used anymore as a tool to assess motor neurone excitability, despite this lingering misinterpretation.
The H reflex as a surrogate for the stretch reflex
The stretch reflex involves the activation of mechanoreceptors located in the muscle (muscle spindles) and in the muscle–tendon junction (tendon organs) in response to a brief stretch of the muscle–tendon unit. The afferent volleys are conveyed along the axons to activate the motor neurones and spinal interneurones through mono- and polysynaptic pathways. However, several aspects differ between the stretch reflex and the H reflex. The distinction is evident by the decrease in H-reflex amplitude in soleus during voluntary contraction when the antagonist muscle (tibialis anterior) is simultaneously activated (Nielsen and Kagamihara 1993), whereas the stretch reflex amplitude (short-latency components) does not change (Nielsen et al. 1993). Similarly, H-reflex amplitude is depressed during the stance phase of walking (Capaday and Stein 1986, 1987b), but not the stretch reflex (Sinkjaer et al. 1996).
As summarised by Burke (2016), the difference between the H and stretch reflexes includes more than the absence of muscle spindle involvement in the H reflex. The electrical stimulus used to evoke the H reflex induces a single action potential in the Ia afferents recruited, whereas the brief stretch of muscle produces high-frequency repetitive discharges in individual afferents with large temporal dispersion (Burke et al. 1983; Morita et al. 1998). Accordingly, there is more time for polysynaptic inputs to reach motor neurones during the stretch than the H reflex (Burke et al. 1984). In contrast, the EPSP rise time in single motor units for the H reflex appears to be brief, probably less than 2 ms, limiting the impact of polysynaptic pathways (disynaptic pathway from Ib afferents). The more synchronous Ia afferent discharge in response to the electrical stimulation can increase its sensitivity to distinct inputs, as shown for the Ia presynaptic inhibition (Morita et al. 1998). The interpretation of the stretch reflex is further complexified by its dependence to fusimotor activity (Llewellyn et al. 1990). Accordingly, the H reflex cannot be used as a surrogate for the stretch reflex, and is more suitable than the stretch reflex to assess neural modulation at the spinal level. A combination of the two types of reflex has been used to dissociate peripheral (muscle spindle sensitivity) vs. spinal mechanisms involved in stretch reflex changes. The results of such comparisons, however, should be interpreted with caution (due to the differences detailed above) and confirmed by complementary approaches.
Current interpretation
The H reflex is mainly used as a tool to assess the spinal excitability (Grosprêtre and Martin 2012; Kim et al. 2019) or the efficacy of the synaptic transmission between Ia afferents and motor neurones (Casabona et al. 1990; Johannsson et al. 2017; Palmieri et al. 2004). These definitions, however, are partly incomplete. The issue with the use of “spinal excitability” is the lack of precision in terms of the structures that are involved. The H reflex depends on both motor neurone excitability and the amount of neurotransmitter released by Ia terminals, which can vary with changes in Ia presynaptic inhibition or in response to repetitive activation of Ia afferents. Furthermore, the repetitive activation (frequency greater than 0.1 Hz) of Ia afferents at rest depresses the neurotransmitter release (homosynaptic depression), thereby decreasing H-reflex amplitude (Hultborn et al. 1996). This decrease is difficult to reconcile with the absence of change in excitability of neural components of the H-reflex pathway (axons, motor neurones and interneurones). The concept of the efficacy of the synaptic transmission suggests that modulations in the H-reflex amplitude reflects changes in the synaptic transmission between Ia afferent terminals and motor neurones. However, H-reflex amplitude can change due to an increase or decrease of other excitatory and inhibitory inputs received by motor neurones, without any changes in synaptic transmission between Ia afferents and motor neurones.
Revisited interpretation
To circumvent these limitations, the H reflex should be considered as a tool to assess the effectiveness of activated Ia afferents in evoking an action potential in motor neurones. This definition encompasses modifications of neurotransmitter release (Ia presynaptic inhibition and homosynaptic depression) and changes in the excitability of the motor neurone pool (postsynaptic mechanisms). Regardless of the loci of the adjustments underlying the change in H-reflex amplitude, there is a change in the probability that the Ia afferent input causes the motor neurones to discharge action potential. For instance, reciprocal inhibition—induced via interneurones activated by Ia afferents of the antagonist muscles (Ia inhibitory interneurones) (Crone et al. 1987)—hyperpolarises the motor neurone membrane and reduces the likelihood that it generates an action potential (Baldissera et al. 1981). This decrease in motor neurone excitability reduces the effectiveness of Ia afferent input to cause motor neurones to discharge action potential, thereby reducing H-reflex amplitude (Crone et al. 1987). However, an increase in Ia presynaptic inhibition, which decreases neurotransmitter release from Ia afferent terminals onto motor neurones (Rudomin 1990), depresses the amplitude and the summation of the EPSPs onto the motor neurone membrane. This reduces the effectiveness of the Ia afferent volley to bring the motor neurone membrane potential to the firing threshold, and thereby the H-reflex amplitude.
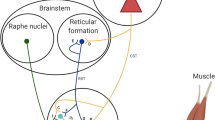
In addition to documenting modulation of H-reflex amplitude, it is important to determine the functional interpretation of the change to highlights part of the control strategy (Fig. 3). For example, a decrease in H-reflex amplitude as the result of an increase in Ia presynaptic inhibition decreases the contribution of Ia afferents to the activity of the homonymous motor neurone pool. In contrast, a decrease in H-reflex amplitude due to an increase in postsynaptic inhibitory mechanisms should be interpreted as a decrease in the excitability of the motor neurone pool without changes in the contribution of Ia afferents to the excitatory input. Further modulation of the motor neurone excitability during voluntary contraction can arise from persistent inward currents (PICs), which are generated by voltage-sensitive Na+ and Ca2+ currents via monoaminergic input from the brainstem (Heckman and Enoka 2012). PICs may amplify synaptic inputs from Ia afferents and increase the effectiveness of the Ia afferent input in cause motor neurones to discharge action potential, although they can have more diverse effects (Heckman et al. 2009; Rekling et al. 2000).

Schematic illustration of the main contributors to the modulation of the effectiveness of activated Ia afferents in causing an action potential in motor neurones (white text), and how they can be interpreted (green text). The arrows represent the main effect of input with ↑ and ↓ indicating an increase or decrease, respectively. MN motor neurone, IN interneurons, PAD primary afferent depolarisation, HD homosynaptic depression
These subtilities in the interpretation of the H reflex and their consequence in the understanding of the control strategy should be considered in future studies to improve the use of this technique and the valuable information it provides. Because the H reflex by itself cannot identify the underlying source of the modulation in its amplitude, additional techniques (D1 inhibition, heteronymous facilitation, post-stimulus time histogram of single motor units, etc.) should be used to unravel this information (Knikou 2008; Pierrot-Deseilligny and Burke 2012).
Perspectives for motor control studies
H-reflex modulation and Ia presynaptic inhibition
Changes in H-reflex amplitude between different experimental conditions have often been accompanied by changes in Ia presynaptic inhibition (Faist et al. 1996; Hultborn et al. 1987; Meunier and Pierrot-Deseilligny 1989; Nielsen and Kagamihara 1993). Modulating the Ia afferent input at a presynaptic level permits the selective gating of Ia input onto α-motor neurones without compromising either the recruitment of motor neurones by other excitatory sources (Rudomin 1990) or the propagation of the proprioceptive information to the supraspinal structures (Llewellyn et al. 1990; Morita et al. 1998). This inhibitory pathway can, therefore, decrease the contribution of Ia afferents to motor neurone activity in favour of other excitatory sources as the descending drive, although this latter possibility may be debated (Baudry and Duchateau 2014). As such, a modulation of Ia presynaptic inhibition could be part of a central set to face a specific situation or be used to modulate the effectiveness of Ia afferents to discharge motor neurones within the time course of a motor action. It should be noted that a decrease in Ia input does not have the same influence as postsynaptic inhibition, because it reduces the number of motor neurones with the subliminal fringe without changing the membrane conductance of the motor neurone (decreased motor neurone excitability). However, a decrease in H-reflex amplitude may not always reflect an increase in Ia presynaptic inhibition even in experimental conditions when a change in Ia presynaptic inhibition may appear the obvious candidate to explain the H-reflex modulation.
A first example underscores a dissociation in the modulation of H-reflex amplitude and Ia presynaptic inhibition during sustained isometric contractions. H-reflex amplitude during sustained fatiguing contractions has been shown to increase (Löscher et al. 1996; Patikas et al. 2006), decrease (Duchateau and Hainaut 1993; Klass et al. 2008), or remain unchanged (Cresswell and Löscher 2000; Martin et al. 2007). These divergent results likely reflect the influence of the experimental conditions (muscle studied, amplitude of the initial H reflex, resting or active muscle, and intensity of the contraction) on the neural adjustments, although Ia presynaptic inhibition has often been suggested to be responsible for such modulation of H-reflex amplitude. In this context, the duration an individual can sustain a contraction while supporting an inertial load (position task) is briefer compared with applying a similar net muscle torque against a rigid restraint for as long as possible (Cresswell and Löscher 2000; Mottram et al. 2006). When performed with the elbow flexors, the briefer time to task failure for the position task was accompanied by a faster and greater decrease in H-reflex amplitude recorded in the biceps brachii (Klass et al. 2008). Such a difference in H-reflex modulation between the two tasks sustained to failure may involve Ia presynaptic inhibition. In support, Ia presynaptic inhibition was shown to be lesser in position than force task when not sustained to failure (Baudry et al. 2010; Baudry and Enoka 2009; Magalhães et al. 2015). During fatiguing contractions, changes in Ia presynaptic inhibition was investigated by recording H reflexes in the extensor carpi radialis during the two types of tasks performed to failure at 20% of participant’s maximal force (Baudry et al. 2011). The H reflexes were evoked with and without a conditioning stimulus used to assess Ia presynaptic inhibition by means of D1 inhibition and heteronymous facilitation (Table 1) (Hultborn et al. 1987). The results indicated an absence of significant changes in unconditioned H reflex during the two tasks. However, the amount of Ia presynaptic inhibition initially decreased but subsequently returned to baseline values during the course of the position task. In contrast, Ia presynaptic inhibition was decreased at the end of the sustained contraction during the force task. The absence of a change in amplitude of the test H reflex while Ia presynaptic inhibition was modulated differently during the two tasks suggests that the nervous system adjusted the descending and peripheral inputs in a task- and time-dependent manner. Different adjustments may compensate themselves to control the net synaptic input (balance between excitatory and inhibitory input) received by the motor neuron pool. Although these results do not preclude an association between H-reflex amplitude and Ia presynaptic inhibition during sustained contractions performed in other experimental conditions or muscles, they suggest caution in inferring changes in presynaptic inhibition based only on H-reflex amplitude.
A second example of a dissociation between the modulation of the H-reflex amplitude and Ia presynaptic inhibition was reported when young and older adults stood in an upright posture (Baudry and Duchateau 2012). In this experiment, the soleus Hmax was recorded while participants stood upright with eyes open and closed. No modulation of the Hmax amplitude was observed between the two visual conditions for either age group. Similarly, the soleus background EMG activity did not differ across vision conditions. However, measures of D1 inhibition (Table 1) and post-stimulus discharge probability of motor unit clearly showed an increase in Ia presynaptic inhibition when standing with eyes closed compared with eyes open. This finding partially contrasts with the results from a previous study (Katz et al. 1988) in which there was a decrease in soleus H reflex accompanied by an increase in Ia presynaptic inhibition when participants went from standing with back support to standing without back support. In two of the six participants, however, the soleus H-reflex amplitude did not decrease when standing without support, whereas there was an increase in Ia presynaptic inhibition in these individuals. These results indicate the possible dissociation between the modulation of the H-reflex amplitude and Ia presynaptic inhibition during upright standing.
A third example involves the modulation of soleus H reflex with the position and direction of postural sway (Baudry and Duchateau 2012; Tokuno et al. 2008). H-reflex amplitude increases when participants lean forward (position) or sway forward (direction) compared with backward position and direction. It was suggested that the sway-related modulation of H-reflex amplitude involved Ia presynaptic inhibition (Tokuno et al. 2008). To confirm this hypothesis, the modulation of Ia presynaptic inhibition was investigated during natural and exaggerated sways in upright standing (Johannsson et al. 2015). The results confirmed the sway-related modulation of the H reflex, but Ia presynaptic inhibition, assessed by two H-reflex conditioning techniques (D1 inhibition and heteronymous facilitation) (Table 1), did not change with the position and the direction of the centre of pressure. These results suggest that, in contrast with H-reflex modulation, Ia presynaptic inhibition is not modulated with sway position and direction.
Along with others (Souron et al. 2019), these three examples illustrate that the involvement of Ia presynaptic inhibition in the modulation of the H-reflex pathway may not be as straightforward as expected. The dissociation between the H-reflex modulation and Ia presynaptic inhibition could reflect opposite changes in postsynaptic mechanisms that can mask the effect of Ia presynaptic inhibition on the H reflex. Regardless of the exact pathways involved, these examples show that caution is necessary when interpreting H-reflex changes. The primary interpretation of a change in H-reflex amplitude should be that spinal adjustments alter the effectiveness of activated Ia afferents in causing an action potential in motor neurones. Whether these modulations occur presynaptically or postsynaptically should be investigated with more refined techniques (D1 inhibition, heteronymous facilitation, post-stimulus time histogram of single motor units).
H-reflex and motor function: the example of postural control
Despite the limitations discussed in the previous sections, the H-reflex technique remains an appropriate method to assess the effectiveness of the activated Ia afferents to evoke action potential in motor neurones in various experimental conditions when methodological parameters are strictly controlled (see “Methodological considerations”). To illustrate this point, a few examples of how the H-reflex method can provide information on the neural control of movement followed by suggestions on how to enhance the outcomes. This section focuses on postural control as this research topic has provided numerous examples of H-reflex modulation.
The soleus H-reflex amplitude decreases during standing compared with sitting or lying position (Angulo-Kinzler et al. 1998; Baudry et al. 2015; Koceja et al. 1995), when standing on one leg compared with standing on two legs (Alizadehsaravi et al. 2022b; Pinar et al. 2010), and when standing on a foam mat compared with standing on a rigid surface (Earles et al. 2000). A decrease in H-reflex amplitude has also been reported when standing at the top of a staircase compared with standing at the bottom (Johannsson et al. 2017). Moreover, there is a decrease in H-reflex amplitude when going from standing with a support to standing without support, requiring thereby to control body sway (Baudry et al. 2014; Katz et al. 1988; Tokuno et al. 2008). Modulation of the H-reflex pathway during unsupported upright standing likely reflects a central set that adjusts sensorimotor pathways to the specificity of the upright posture adopted (Horak et al. 1989).
To evaluate the functional role of this modulation, however, it is necessary to associate changes in H reflex with some balance parameters. To address this issue, Koceja et al. (1995) investigated H-reflex modulation in young and older adults when they went from a prone to a standing position. Although H-reflex amplitude was depressed in young adults during standing, there was no significant correlation between the H-reflex variation and the centre of pressure excursion. In contrast, there was no statistically significant change in H-reflex amplitude when older adults went from prone to a standing position. Still, the H-reflex variations were correlated with the centre of pressure excursions; greater depression of H-reflex amplitude was correlated with less postural instability.
More recently, Alizadehsaravi et al. (2022a) investigated the adaptations underlying improved balance after one and ten sessions of balance training in older adults. H reflex was measured in soleus during bipedal and unipedal upright standing before and after one session of perturbed one-leg standing and ten sessions of unperturbed one-leg standing exercises. The change in H-reflex amplitude between two-leg and one-leg stance, which decreased in one-leg stance (Alizadehsaravi et al. 2022a; Pinar et al. 2010), was assessed. The decrease in H-reflex amplitude during one-leg stance was correlated with better balance performances in this situation. However, the improvement in balance performance in this same postural situation after one or ten sessions was not correlated with H-reflex amplitude. The conclusion was that the decrease in H-reflex amplitude (decreased effectiveness of the activated Ia afferents to discharge motor neurones) may contribute to control balance in one-leg stance, but that improvement in balance after one or several balance training sessions may not require specific modulation of the H-reflex.
Together, these studies illustrate two important points when using the H-reflex technique to identify changes associated with motor function. First, a change in H-reflex amplitude may not be associated with movement control per se, but may reflect a central set used to adjust sensorimotor pathways to the context of the task. Second, searching for the functional role of H-reflex modulation in motor performance can generate complementary information.
Even though the H-reflex pathway uses part of the proprioceptive circuitry (Henry and Baudry 2019), modulation of the H reflex can only provide a limited view of the change in the contribution of proprioception in motor control. Indeed, the H reflex cannot indicate how the information from the proprioceptors is integrated and used to control movement. As mentioned above, the H reflex reflects the effectiveness of activated Ia afferents to discharge motor neurones and its modulation does not tell us more than a change in the recruitment of motor units by the Ia input due to one or several mechanisms. Thus, it is not straightforward to consider discussing the contribution of proprioception in motor control based only on the H-reflex technique.
H reflex and chronic neuromuscular changes: the example of ageing
A large body of the literature documents a decrease in H-reflex amplitude and an increase in H-reflex latency with ageing (Baudry et al. 2015; Kido et al. 2004; Klass et al. 2011; Sabbahi and Sedgwick 1982; Scaglioni et al. 2003), although some studies report no age-related changes (Angulo-Kinzler et al. 1998; Earles et al. 2001). In two cross-sectional studies, there was a linear decrease in the soleus H-reflex amplitude with age for samples of 23 (Kido et al. 2004) and 142 participants (Baudry 2016) aged between 20 and 82 years old. It is noteworthy that these two studies also reported large variability in the H-reflex amplitude, regardless of age. For example, H-reflex amplitude ranged from 30 to 80% Mmax in adults aged between 20 and 30 years and from 10 to 50% Mmax in adults aged between 70 and 80 years (Baudry 2016). Similar variability was observed whether participants were tested seated or standing (Baudry 2016), and was also reported by others (Schieppati 1987; Táboríková and Sax 1968). Importantly, this variability can confound the effect of ageing on the H reflex as participants sampling can lead to similar or different H-reflex amplitude between young and older adults.
One source of variability to consider involves the habitual physical activity of the participants. For example, there was an increase in H-reflex amplitude in soleus during a low-force contraction (10% of the participant’s maximal force) after young adults perform a short-term (3 weeks) training program to increase cardiorespiratory capacities (endurance training), whereas there was no change after a strength-training programme (Vila-Chã et al. 2012); longer strength-training programmes can, however, increase in H-reflex amplitude (Aagaard et al. 2002). Balance training also influences the H-reflex pathway, as illustrated by a decrease in H-reflex amplitude after a training programme (Gruber et al. 2007). In agreement, Nielsen and collaborators reported that the H-reflex amplitude was lesser in professional ballet dancers than in untrained participants (Nielsen et al. 1993). Although the effects of strength and balance training on the H-reflex pathway in older adults appears to be more mitigated (Penzer et al. 2015; Ruffieux et al. 2017; Scaglioni et al. 2002), these results suggest that habitual levels of physical activity or inactivity should be taken into account when investigating the effect of ageing on H-reflex pathway.
However, the age-related reduction in H-reflex amplitude has been suggested to reflect an increase in Ia presynaptic inhibition, although the influence of ageing on this pathway is unclear. Morita et al. (1995) and Koceja and Mynark (2000) found greater Ia presynaptic inhibition in older than young participants in soleus. In contrast, using tendon vibration and D1 inhibition, Butchart et al. (1993) and Earles et al. (2001) reported a decrease in Ia presynaptic inhibition in older adults for the soleus. However, others found no difference in Ia presynaptic inhibition between young and older adults in upper and lower limb muscles (Baudry et al. 2010; Baudry and Duchateau 2012; Filho et al. 2021). These divergent results do not support Ia presynaptic inhibition as a primary contributor to the lesser H-reflex amplitude in older adults. Furthermore, previous work indicates that ageing may not influence the modulation of Ia presynaptic inhibition (Baudry and Duchateau 2012; Filho et al. 2021).
More likely explanations include a loss of afferent fibres, nerve demyelination, changes within the Ia afferent-motor neurone synapse, and an increase in the collision between the afferent and antidromic volleys elicited in motor axons (for review, Henry and Baudry 2019). The increase in H-reflex latency but not M-wave latency and the greater M-wave amplitude associated with Hmax reported in older adults also suggest earlier changes in the sensory than the motor part of the H-reflex pathway (Scaglioni et al. 2003). To further investigate this possibility, more studies are required, notably on changes in biophysical properties in afferent and motor axons with ageing.
Recommendations
This brief narrative review revisits specific aspects of the H-reflex technique to establish its appropriate use and interpretation, as summarized in Fig. 4. The main outcomes of this review are provided as the following recommendations.

Schematic illustration of the basic components of the Hoffmann (H) reflex (A) and a conceptual diagram for the interpretation of the modulation of the H-reflex amplitude (B). Ia Ia afferents from muscle spindles, Ib Ib afferents from Golgi tendon organ, α motor axon from α motor neurone, MN motor neurone, M wave compound muscle action potential, EMG electromyography, Δ variation
Procedure for recording H reflex
-
Use an intensity that elicits a preceding M-wave amplitude of 5–10% of the Mmax;
-
Normalising the H-reflex amplitude to the Mmax recorded in similar conditions;
-
Ensure a similar background EMG activity preceding the stimulation between conditions;
-
Average 5–10 H reflexes for each given stimulus intensity.
Definition and interpretations
-
The H reflex is a tool to assess the effectiveness of activated Ia afferents to discharge motor neurones.
-
The functional interpretation of changes in H-reflex amplitude should consider the origin of the adjustment. A modulation at a presynaptic level reflects a change in the contribution of Ia afferents to the activity of homonymous motor neurones. A modulation in H-reflex amplitude due to a change in postsynaptic excitatory or inhibitory mechanisms should, in contrast, indicates a modification in the excitability of the motor neurone pool, without compromising the excitatory input delivered by the Ia afferents to the pool.
-
Changes in H-reflex amplitude may not reflect changes in Ia presynaptic inhibition. To ensure the involvement of Ia presynaptic inhibition, additional techniques must be used.
-
Searching for the functional role of H-reflex modulation in motor performance can generate relevant information control strategies.
-
Comparisons between groups or populations must consider the level and type of physical activity, as well as the experimental conditions.
Conclusion
Movements of our everyday life, including occupational and recreational activities, involve complex and specific sensorimotor adjustments that should be studied with appropriate techniques. The H reflex is a relatively simple technique that can be used to test various hypothesis in humans during motor tasks that mimic real-life activities. Investigations with the H reflex and its modulation across experimental conditions, tasks, and participants have provided useful insight for our understanding of the neural control of movement. Nonetheless, the applied method must ensure appropriate recordings and caution should be used when interpreting the data. Our brief narrative review proposes that the H reflex should be used as a tool to assess the effectiveness of the Ia afferents to discharge motor neurones in a given experimental condition. This definition expands on previous definitions and emphasizes the need to test the origin of the adjustments (presynaptic or postsynaptic mechanisms) to provide relevant information on control strategies. We also emphasize the importance of developing appropriate approaches to link H reflex and motor function, including methodological recommendations. When the requirements are met, the H-reflex technique provides useful information on the function of the neuromuscular system.
Abbreviations
- EMG:
-
Electromyography
- EPSP:
-
Excitatory postsynaptic potential
- H reflex:
-
Hoffmann reflex
- H max :
-
Maximal amplitude of the Hoffmann reflex
- IPSP:
-
Inhibitory postsynaptic potential
- MN:
-
Motor neurone
- M wave:
-
Compound muscle action potential
- M max :
-
Maximal amplitude of the compound muscle action potential
- PAD:
-
Primary afferent depolarisation interneurones
- PICs:
-
Persistent inward currents
References
Aagaard P, Simonsen EB, Andersen JL, Magnusson P, Dyhre-Poulsen P (2002) Neural adaptation to resistance training: changes in evoked V-wave and H-reflex responses. J Appl Physiol 92(6):2309–2318. https://doi.org/10.1152/japplphysiol.01185.2001
Abdelmoula A, Baudry S, Duchateau J (2019) Anodal transcranial direct current stimulation does not influence the neural adjustments associated with fatiguing contractions in a hand muscle. Eur J Appl Physiol 119(3):597–609. https://doi.org/10.1007/s00421-018-4027-4
Aimonetti JM, Vedel JP, Schmied A, Pagni S (2000) Distribution of presynaptic inhibition on type-identified motoneurones in the extensor carpi radialis pool in man. J Physiol 522(Pt 1):125–135. https://doi.org/10.1111/j.1469-7793.2000.t01-1-00125.xm
Alizadehsaravi L, Bruijn SM, Muijres W, Koster RAJ, van Dieën JH (2022a) Improvement in gait stability in older adults after ten sessions of standing balance training. PLoS One 17(7):e0242115. https://doi.org/10.1371/journal.pone.0242115
Alizadehsaravi L, Koster RAJ, Muijres W, Maas H, Bruijn SM, van Dieën JH (2022b) The underlying mechanisms of improved balance after one and ten sessions of balance training in older adults. Hum Mov Sci 81:102910. https://doi.org/10.1016/j.humov.2021.102910
Angulo-Kinzler RM, Mynark RG, Koceja DM (1998) Soleus H-reflex gain in elderly and young adults: modulation due to body pos. J Gerontol A Biol Sci Med Sci 53A(2):120–125
Awiszus F, Feistner H (1993) The relationship between estimates of Ia-EPSP amplitude and conduction velocity in human soleus motoneurons. Exp Brain Res 95(2):365–370. https://doi.org/10.1007/BF00229795
Baldissera F, Hultborn H, Illert M (1981) Integration in spinal neuronal systems. Comprehensive physiology. Wiley, New York, pp 509–595. https://doi.org/10.1002/cphy.cp010212
Baudry S (2016) Aging changes the contribution of spinal and corticospinal pathways to control balance. Exerc Sport Sci Rev 44(3):104–109. https://doi.org/10.1249/JES.0000000000000080
Baudry S, Duchateau J (2012) Age-related influence of vision and proprioception on Ia presynaptic inhibition in soleus muscle during upright stance: ageing and Ia presynaptic inhibition during balance. J Physiol 590(21):5541–5554. https://doi.org/10.1113/jphysiol.2012.228932
Baudry S, Duchateau J (2014) Independent modulation of corticospinal and group I afferents pathways during upright standing. Neuroscience 275:162–169. https://doi.org/10.1016/j.neuroscience.2014.06.021
Baudry S, Duchateau J (2020) Aftereffects of prolonged Achilles tendon vibration on postural control are reduced in older adults. Exp Gerontol 131:110822. https://doi.org/10.1016/j.exger.2019.110822
Baudry S, Enoka RM (2009) Influence of load type on presynaptic modulation of Ia afferent input onto two synergist muscles. Exp Brain Res 199(1):83–88. https://doi.org/10.1007/s00221-009-1951-x
Baudry S, Maerz AH, Enoka RM (2010) Presynaptic modulation of Ia afferents in young and old adults when performing force and position control. J Neurophysiol 103(2):623–631. https://doi.org/10.1152/jn.00839.2009
Baudry S, Maerz AH, Gould JR, Enoka RM (2011) Task- and time-dependent modulation of Ia presynaptic inhibition during fatiguing contractions performed by humans. J Neurophysiol 106(1):265–273. https://doi.org/10.1152/jn.00954.2010
Baudry S, Penzer F, Duchateau J (2014) Input-output characteristics of soleus homonymous Ia afferents and corticospinal pathways during upright standing differ between young and elderly adults. Acta Physiol 210(3):667–677. https://doi.org/10.1111/apha.12233
Baudry S, Collignon S, Duchateau J (2015) Influence of age and posture on spinal and corticospinal excitability. Exp Gerontol 69:62–69. https://doi.org/10.1016/j.exger.2015.06.006
Behm DG, Peach A, Maddigan M, Aboodarda SJ, DiSanto MC, Button DC, Maffiuletti NA (2013) Massage and stretching reduce spinal reflex excitability without affecting twitch contractile properties. J Electromyogr Kinesiol 23(5):1215–1221. https://doi.org/10.1016/j.jelekin.2013.05.002
Bostock H, Rothwell JC (1997) Latent addition in motor and sensory fibres of human peripheral nerve. J Physiol 498(1):277–294. https://doi.org/10.1113/jphysiol.1997.sp021857
Budini F, Christova M, Gallasch E, Rafolt D, Tilp M (2018) Soleus H-reflex inhibition decreases during 30 s static stretching of plantar flexors, showing two recovery steps. Front Physiol 9:935. https://doi.org/10.3389/fphys.2018.00935
Buller NP, Garnett R, Stephens JA (1980) The reflex responses of single motor units in human hand muscles following muscle afferent stimulation. J Physiol 303:337–349. https://doi.org/10.1113/jphysiol.1980.sp013289
Burke RE (1968) Group Ia synaptic input to fast and slow twitch motor units of cat triceps surae. J Physiol 196(3):605–630. https://doi.org/10.1113/jphysiol.1968.sp008526
Burke D (2016) Clinical uses of H reflexes of upper and lower limb muscles. Clin Neurophysiol Pract 1:9–17. https://doi.org/10.1016/j.cnp.2016.02.003
Burke D, Hagbarth KE, Löfstedt L, Wallin BG (1976) The responses of human muscle spindle endings to vibration of non-contracting muscles. J Physiol 261(3):673–693. https://doi.org/10.1113/jphysiol.1976.sp011580
Burke D, Gandevia SC, McKeon B (1983) The afferent volleys responsible for spinal proprioceptive reflexes in man. J Physiol 339:535–552
Burke D, Gandevia SC, McKeon B (1984) Monosynaptic and oligosynaptic contributions to human ankle jerk and H-reflex. J Neurophysiol 52(3):435–448. https://doi.org/10.1152/jn.1984.52.3.435
Burke D, Adams RW, Skuse NF (1989) The effects of voluntary contraction on the H reflex of human limb muscles. Brain J Neurol 112(Pt 2):417–433. https://doi.org/10.1093/brain/112.2.417
Butchart P, Farquhar R, Part N, Roberts R (1993) The effect of age and voluntary contraction on presynaptic inhibition of soleus muscle Ia afferent terminals in man. Exp Physiol 78(2):235–242. https://doi.org/10.1113/expphysiol.1993.sp003683
Capaday C, Stein R (1986) Amplitude modulation of the soleus H-reflex in the human during walking and standing. J Neurosci 6(5):1308–1313. https://doi.org/10.1523/JNEUROSCI.06-05-01308.1986
Capaday C, Stein RB (1987a) A method for simulating the reflex output of a motoneuron pool. J Neurosci Methods 21(2–4):91–104. https://doi.org/10.1016/0165-0270(87)90107-5
Capaday C, Stein RB (1987b) Difference in the amplitude of the human soleus H reflex during walking and running. J Physiol 392(1):513–522. https://doi.org/10.1113/jphysiol.1987.sp016794
Casabona A, Polizzi MC, Perciavalle V (1990) Differences in H-reflex between athletes trained for explosive contractions and non-trained subjects. Eur J Appl Physiol 61(1–2):26–32. https://doi.org/10.1007/BF00236689
Christie A, Lester S, LaPierre D, Gabriel DA (2004) Reliability of a new measure of H-reflex excitability. Clin Neurophysiol 115(1):116–123. https://doi.org/10.1016/S1388-2457(03)00306-7
Cresswell AG, Löscher WN (2000) Significance of peripheral afferent input to the α-motoneurone pool for enhancement of tremor during an isometric fatiguing contraction. Eur J Appl Physiol 82(1):129–136. https://doi.org/10.1007/s004210050662
Crone C, Hultborn H, Jespersen B, Nielsen J (1987) Reciprocal Ia inhibition between ankle flexors and extensors in man. J Physiol 389(1):163–185. https://doi.org/10.1113/jphysiol.1987.sp016652
Crone C, Johnsen LL, Hultborn H, Ørsnes GB (1999) Amplitude of the maximum motor response (Mmax) in human muscles typically decreases during the course of an experiment. Exp Brain Res 124(2):265–270. https://doi.org/10.1007/s002210050621
Datoussaid M, El Khalouqi H, Dahm C, Guissard N, Baudry S (2021) Passive torque influences the Hoffmann reflex pathway during the loading and unloading phases of plantar flexor muscles stretching. Physiol Rep. https://doi.org/10.14814/phy2.14834
Duchateau J, Baudry S (2014) Insights into the neural control of eccentric contractions. J Appl Physiol 116(11):1418–1425. https://doi.org/10.1152/japplphysiol.00002.2013
Duchateau J, Hainaut K (1993) Behaviour of short and long latency reflexes in fatigued human muscles. J Physiol 471(1):787–799. https://doi.org/10.1113/jphysiol.1993.sp019928
Earles DR, Koceja DM, Shively CW (2000) Environmental changes in soleus H-reflex excitability in young and elderly subjects. Int J Neurosci 105(1–4):1–13. https://doi.org/10.3109/00207450009003261
Earles D, Vardaxis V, Koceja D (2001) Regulation of motor output between young and elderly subjects. Clin Neurophysiol 112(7):1273–1279. https://doi.org/10.1016/S1388-2457(01)00571-5
Eccles JC, Eccles RM, Lundberg A (1957) The convergence of monosynaptic excitatory afferents on to many different species of alpha motoneurones. J Physiol 137(1):22–50. https://doi.org/10.1113/jphysiol.1957.sp005794
Faist M, Dietz V, Pierrot-Deseilligny E (1996) Modulation, probably presynaptic in origin, of monosynaptic Ia excitation during human gait. Exp Brain Res. https://doi.org/10.1007/BF00229628
Filho SS, Coelho DB, Ugrinowitsch C, de Souza CR, Magalhães FH, de Lima-Pardini AC, de Oliveira ÉMB, Mattos E, Teixeira LA, Silva-Batista C (2021) Age-related changes in presynaptic inhibition during gait initiation. J Gerontol Ser A 76(4):568–575. https://doi.org/10.1093/gerona/glab010
Frank K, Fuortes M (1957) Presynaptic and postsynaptic inhibition of monosynaptic reflexes, vol 16, pp 39–40
Funase K, Miles TS (1999) Observations on the variability of the H reflex in human soleus. Muscle Nerve 22(3):341–346. https://doi.org/10.1002/(SICI)1097-4598(199903)22:3%3c341::AID-MUS6%3e3.0.CO;2-R
Funase K, Imanaka K, Nishihira Y (1994) Excitability of the soleus motoneuron pool revealed by the developmental slope of the H-reflex as reflex gain. Electromyogr Clin Neurophysiol 34(8):477–489
Gossard J, Floeter M, Kawai Y, Burke R, Chang T, Schiff S (1994) Fluctuations of excitability in the monosynaptic reflex pathway to lumbar motoneurons in the cat. J Neurophysiol 72:1227–1239. https://doi.org/10.1152/jn.1994.72.3.1227
Gottlieb GL, Agarwal GC (1973) Modulation of postural reflexes by voluntary movement. J Neurol Neurosurg Psychiatry 36(4):529–539
Grosprêtre S, Martin A (2012) H reflex and spinal excitability: methodological considerations. J Neurophysiol 107(6):1649–1654. https://doi.org/10.1152/jn.00611.2011
Gruber M, Taube W, Gollhofer A, Beck S, Amtage F, Schubert M (2007) Training-specific adaptations of H- and stretch reflexes in human soleus muscle. J Mot Behav 39(1):68–78. https://doi.org/10.3200/JMBR.39.1.68-78
Hallett M, DelRosso LM, Elble R, Ferri R, Horak FB, Lehericy S, Mancini M, Matsuhashi M, Matsumoto R, Muthuraman M, Raethjen J, Shibasaki H (2021) Evaluation of movement and brain activity. Clin Neurophysiol off J Int Feder Clin Neurophysiol 132(10):2608–2638. https://doi.org/10.1016/j.clinph.2021.04.023
Heckman CJ, Binder MD (1988) Analysis of effective synaptic currents generated by homonymous Ia afferent fibers in motoneurons of the cat. J Neurophysiol 60(6):1946–1966. https://doi.org/10.1152/jn.1988.60.6.1946
Heckman CJ, Enoka RM (2012) Motor unit. In: Terjung R (ed) Comprehensive physiology, 1st edn. Wiley, New York, pp 2629–2682. https://doi.org/10.1002/cphy.c100087
Heckman CJ, Mottram C, Quinlan K, Theiss R, Schuster J (2009) Motoneuron excitability: the importance of neuromodulatory inputs. Clin Neurophysiol 120(12):2040–2054. https://doi.org/10.1016/j.clinph.2009.08.009
Henneman E, Mendell LM (1981) Functional organization of motoneuron pool and its inputs. In: Terjung R (ed) Comprehensive physiology, 1st edn. Wiley, New York, pp 423–507. https://doi.org/10.1002/cphy.cp010211
Henry M, Baudry S (2019) Age-related changes in leg proprioception: implications for postural control. J Neurophysiol 122(2):525–538. https://doi.org/10.1152/jn.00067.2019
Hoch MC, Krause BA (2009) Intersession reliability of H:M ratio is greater than the H-reflex at a percentage of M-max. Int J Neurosci 119(3):345–352. https://doi.org/10.1080/00207450802480309
Hopkins JT, Wagie NC (2003) Intrasession and intersession reliability of the quadriceps Hoffmann reflex. Electromyogr Clin Neurophysiol 43(2):85–89
Horak F, Diener H, Nashner L (1989) Influence of central set on human postural responses. J Neurophysiol 62:841–853. https://doi.org/10.1152/jn.1989.62.4.841
Hugon M (1973) Methodology of the Hoffmann reflex in man. Hum Reflexes Pathophysiol Motor Syst Methodol Hum Reflexes 3:277–293. https://doi.org/10.1159/000394143
Hultborn H, Meunier S, Morin C, Pierrot-Deseilligny E (1987) Assessing changes in presynaptic inhibition of I a fibres: a study in man and the cat. J Physiol 389(1):729–756. https://doi.org/10.1113/jphysiol.1987.sp016680
Hultborn H, Illert M, Nielsen J, Paul A, Ballegaard M, Wiese H (1996) On the mechanism of the post-activation depression of the H-reflex in human subjects. Exp Brain Res. https://doi.org/10.1007/BF00227268
Jaberzadeh S, Scutter S, Warden-Flood A, Nazeran H (2004) Between-days reliability of H-reflexes in human flexor carpi radialis11No commercial party having a direct financial interest in the results of the research supporting this article has or will confer a benefit on the author(s) or on any organization with which the author(s) is/are associated. Arch Phys Med Rehabil 85(7):1168–1173. https://doi.org/10.1016/j.apmr.2003.09.009
Johannsson J, Duchateau J, Baudry S (2015) Presynaptic inhibition of soleus Ia afferents does not vary with center of pressure displacements during upright standing. Neuroscience 298:63–73. https://doi.org/10.1016/j.neuroscience.2015.04.010
Johannsson J, Duchateau J, Baudry S (2017) Spinal and corticospinal pathways are differently modulated when standing at the bottom and the top of a three-step staircase in young and older adults. Eur J Appl Physiol 117(6):1165–1174. https://doi.org/10.1007/s00421-017-3603-3
Jones KE, Bawa P (1997) Computer simulation of the responses of human motoneurons to composite 1A EPSPS: effects of background firing rate. J Neurophysiol 77(1):405–420. https://doi.org/10.1152/jn.1997.77.1.405
Katz R, Meunier S, Pierrot-Deseilligny E (1988) Changes in presynaptic inhibition of Ia fibres in man while standing. Brain J Neurol 111(Pt 2):417–437. https://doi.org/10.1093/brain/111.2.417
Kido A, Tanaka N, Stein RB (2004) Spinal reciprocal inhibition in human locomotion. J Appl Physiol 96:9
Kiernan MC, Mogyoros I, Miller TA, Burke D (1996) The significance of temporal dispersion in the measurement of compound nerve action potentials. Electroencephalogr Clin Neurophysiol 3(98):31P
Kim K-M, Kim J-S, Cruz-Díaz D, Ryu S, Kang M, Taube W (2019) Changes in spinal and corticospinal excitability in patients with chronic ankle instability: a systematic review with meta-analysis. J Clin Med 8(7):E1037. https://doi.org/10.3390/jcm8071037
Kimura J, Sakimura Y, Machida M, Fuchigami Y, Ishida T, Claus D, Kameyama S, Nakazumi Y, Wang J, Yamada T (1988) Effect of desynchronized inputs on compound sensory and muscle action potentials. Muscle Nerve 11(7):694–702. https://doi.org/10.1002/mus.880110705
Klass M, Lévénez M, Enoka RM, Duchateau J (2008) Spinal mechanisms contribute to differences in the time to failure of submaximal fatiguing contractions performed with different loads. J Neurophysiol 99(3):1096–1104. https://doi.org/10.1152/jn.01252.2007
Klass M, Baudry S, Duchateau J (2011) Modulation of reflex responses in activated ankle dorsiflexors differs in healthy young and elderly subjects. Eur J Appl Physiol 111(8):1909–1916. https://doi.org/10.1007/s00421-010-1815-x
Knikou M (2008) The H-reflex as a probe: pathways and pitfalls. J Neurosci Methods 171(1):1–12. https://doi.org/10.1016/j.jneumeth.2008.02.012
Koceja DM, Mynark RG (2000) Comparison of heteronymous monosynaptic Ia facilitation in young and elderly subjects in supine and standing positions. Int J Neurosci 103(1–4):1–17. https://doi.org/10.3109/00207450009035005
Koceja DM, Trimble MH, Earles DR (1993) Inhibition of the soleus H-reflex in standing man. Brain Res 629(1):155–158. https://doi.org/10.1016/0006-8993(93)90495-9
Koceja DM, Markus CA, Trimble MH (1995) Postural modulation of the soleus H reflex in young and old subjects. Electroencephalogr Clin Neurophysiol Electromyogr Motor Control 97(6):387–393. https://doi.org/10.1016/0924-980X(95)00163-F
Llewellyn M, Yang JF, Prochazka A (1990) Human H-reflexes are smaller in difficult beam walking than in normal treadmill walking. Exp Brain Res 83(1):22–28. https://doi.org/10.1007/BF00232189
Löscher WN, Cresswell AG, Thorstensson A (1996) Excitatory drive to the alpha-motoneuron pool during a fatiguing submaximal contraction in man. J Physiol 491(Pt 1):271–280
Lucas SM, Cope TC, Binder MD (1984) Analysis of individual Ia-afferent EPSPs in a homonymous motoneuron pool with respect to muscle topography. J Neurophysiol 51(1):64–74. https://doi.org/10.1152/jn.1984.51.1.64
Macefield VG, Gandevia SC, Bigland-Ritchie B, Gorman RB, Burke D (1993) The firing rates of human motoneurones voluntarily activated in the absence of muscle afferent feedback. J Physiol 471(1):429–443. https://doi.org/10.1113/jphysiol.1993.sp019908
Magalhães F, Elias L, Silva C, Fava de Lima F, Toledo D, Kohn A (2015) D1 and d2 inhibitions of the soleus H-reflex are differentially modulated during plantarflexion force and position tasks. PLoS One 10:e0143862. https://doi.org/10.1371/journal.pone.0143862
Magladery JW, McDougal DB (1950) Electrophysiological studies of nerve and reflex activity in normal man. I. Identification of certain reflexes in the electromyogram and the conduction velocity of peripheral nerve fibers. Bull Johns Hopkins Hosp 86(5):265–290
Marchand-Pauvert V, Nicolas G, Burke D, Pierrot-Deseilligny E (2002) Suppression of the H reflex in humans by disynaptic autogenetic inhibitory pathways activated by the test volley. J Physiol 542(3):963–976. https://doi.org/10.1113/jphysiol.2002.021683
Martin PG, Gandevia SC, Taylor JL (2007) Muscle fatigue changes cutaneous suppression of propriospinal drive to human upper limb muscles. J Physiol 580(Pt 1):211–223. https://doi.org/10.1113/jphysiol.2006.125997
Mendell LM, Henneman E (1968) Terminals of single Ia fibers: distribution within a pool of 300 homonymous motor neurons. Science 160(3823):96–98
Meunier S, Pierrot-Deseilligny E (1989) Gating of the afferent volley of the monosynaptic stretch reflex during movement in man. J Physiol 419(1):753–763. https://doi.org/10.1113/jphysiol.1989.sp017896
Morita H, Shindo M, Yanagawa S, Yoshida T, Momoi H, Yanagisawa N (1995) Progressive decrease in heteronymous monosynaptic Ia facilitation with human ageing. Exp Brain Res 104(1):167–170. https://doi.org/10.1007/BF00229867
Morita H, Petersen N, Christensen LOD, Sinkjær T, Nielsen J (1998) Sensitivity of H-reflexes and stretch reflexes to presynaptic inhibition in humans. J Neurophysiol 80(2):610–620. https://doi.org/10.1152/jn.1998.80.2.610
Mottram CJ, Maluf KS, Stephenson JL, Anderson MK, Enoka RM (2006) Prolonged vibration of the biceps brachii tendon reduces time to failure when maintaining arm position with a submaximal load. J Neurophysiol 95(2):1185–1193. https://doi.org/10.1152/jn.00807.2005
Nielsen J, Kagamihara Y (1993) The regulation of presynaptic inhibition during co-contraction of antagonistic muscles in man. J Physiol 464(1):575–593. https://doi.org/10.1113/jphysiol.1993.sp019652
Nielsen J, Crone C, Hultborn H (1993) H-reflexes are smaller in dancers from The Royal Danish Ballet than in well-trained athletes. Eur J Appl Physiol 66(2):116–121. https://doi.org/10.1007/BF01427051
Olney RK, Budingen HJ, Miller RG (1987) The effect of temporal dispersion on compound action potential area in human peripheral nerve. Muscle Nerve 10(8):728–733. https://doi.org/10.1002/mus.880100809
Palmieri RM, Ingersoll CD, Hoffman MA (2004) The Hoffmann reflex : methodologic considerations and applications for use in sports medicine and athletic training research. J Athl Train 39(3):268–277
Patikas DA, Bassa H, Kotzamanidis C (2006) Changes in the reflex excitability during and after a sustained, low-intensity muscle contraction. Int J Sports Med 27(02):124–130. https://doi.org/10.1055/s-2005-837490
Penzer F, Duchateau J, Baudry S (2015) Effects of short-term training combining strength and balance exercises on maximal strength and upright standing steadiness in elderly adults. Exp Gerontol 61:38–46. https://doi.org/10.1016/j.exger.2014.11.013
Pierrot-Deseilligny E, Burke D (2012) The circuitry of the human spinal cord : spinal and corticospinal mechanisms of movement. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9781139026727
Pierrot-Deseilligny E, Mazevet D (2000) The monosynaptic reflex: a tool to investigate motor control in humans. Interest and limits. Neurophysiol Clin Clin Neurophysiol 30(2):67–80. https://doi.org/10.1016/S0987-7053(00)00062-9
Pierrot-Deseilligny E, Morin C, Bergego C, Tankov N (1981) Pattern of group I fibre projections from ankle flexor and extensor muscles in man. Exp Brain Res 42(3):337–350. https://doi.org/10.1007/BF00237499
Pinar S, Kitano K, Koceja DM (2010) Role of vision and task complexity on soleus H-reflex gain. J Electromyogr Kinesiol 20(2):354–358. https://doi.org/10.1016/j.jelekin.2009.03.002
Prochazka A, Westerman RA, Ziccone SP (1977) Ia afferent activity during a variety of voluntary movements in the cat. J Physiol 268(2):423–448. https://doi.org/10.1113/jphysiol.1977.sp011864
Rekling JC, Funk GD, Bayliss DA, Dong X-W, Feldman JL (2000) Synaptic control of motoneuronal excitability. Physiol Rev 80(2):767–852
Rodriguez-Falces J, Duchateau J, Muraoka Y, Baudry S (2015) M-wave potentiation after voluntary contractions of different durations and intensities in the tibialis anterior. J Appl Physiol 118(8):953–964. https://doi.org/10.1152/japplphysiol.01144.2014
Rudomin P (1990) Presynaptic inhibition of muscle spindle and tendon organ afferents in the mammalian spinal cord. Trends Neurosci 13(12):499–505. https://doi.org/10.1016/0166-2236(90)90084-N
Ruffieux J, Mouthon A, Keller M, Wälchli M, Taube W (2017) Behavioral and neural adaptations in response to five weeks of balance training in older adults: a randomized controlled trial. J Negat Results Biomed 16(1):11. https://doi.org/10.1186/s12952-017-0076-1
Sabbahi MA, Sedgwick EM (1982) Age-related changes in monosynaptic reflex excitability. J Gerontol 37(1):24–32. https://doi.org/10.1093/geronj/37.1.24
Scaglioni G, Ferri A, Minetti AE, Martin A, Van Hoecke J, Capodaglio P, Sartorio A, Narici MV (2002) Plantar flexor activation capacity and H reflex in older adults: adaptations to strength training. J Appl Physiol 92(6):2292–2302. https://doi.org/10.1152/japplphysiol.00367.2001
Scaglioni G, Narici MV, Maffiuletti NA, Pensini M, Martin A (2003) Effect of ageing on the electrical and mechanical properties of human soleus motor units activated by the H reflex and M wave. J Physiol 548(2):649–661. https://doi.org/10.1113/jphysiol.2002.032763
Schieppati M (1987) The Hoffmann reflex: a means of assessing spinal reflex excitability and its descending control in man. Prog Neurobiol 28(4):345–376. https://doi.org/10.1016/0301-0082(87)90007-4
Schmied A, Morin D, Vedel JP, Pagni S (1997) The « size principle » and synaptic effectiveness of muscle afferent projections to human extensor carpi radialis motoneurones during wrist extension. Exp Brain Res 113(2):214–229. https://doi.org/10.1007/BF02450320
Schulte-Mattler WJ, Jakob M, Zierz S (1999) Assessment of temporal dispersion in motor nerves with normal conduction velocity. Clin Neurophysiol 110(4):740–747. https://doi.org/10.1016/S1388-2457(98)00068-6
Shimatani Y, Matamala JM, Howells J, Kiernan MC (2018) 3. Characterization of persistent Na+ current in human motor and sensory axons: exploring the activity at different target levels. Clin Neurophysiol 129(4):e2. https://doi.org/10.1016/j.clinph.2017.12.016
Sinkjaer T, Andersen JB, Larsen B (1996) Soleus stretch reflex modulation during gait in humans. J Neurophysiol 76:1112–1120
Souron R, Baudry S, Millet GY, Lapole T (2019) Vibration-induced depression in spinal loop excitability revisited. J Physiol 597(21):5179–5193. https://doi.org/10.1113/JP278469
Stein RB, Estabrooks KL, McGie S, Roth MJ, Jones KE (2007) Quantifying the effects of voluntary contraction and inter-stimulus interval on the human soleus H-reflex. Exp Brain Res 182(3):309–319. https://doi.org/10.1007/s00221-007-0989-x
Táboríková H, Sax DS (1968) Motoneurone pool and the H-reflex. J Neurol Neurosurg Psychiatry 31(4):354–361. https://doi.org/10.1136/jnnp.31.4.354
Tokuno CD, Garland SJ, Carpenter MG, Thorstensson A, Cresswell AG (2008) Sway-dependent modulation of the triceps surae H-reflex during standing. J Appl Physiol 104(5):1359–1365. https://doi.org/10.1152/japplphysiol.00857.2007
Tucker KJ, Tuncer M, Türker KS (2005) A review of the H-reflex and M-wave in the human triceps surae. Hum Mov Sci 24(5):667–688. https://doi.org/10.1016/j.humov.2005.09.010
Upton AR, McComas AJ, Sica RE (1971) Potentiation of « late » responses evoked in muscles during effort. J Neurol Neurosurg Psychiatry 34(6):699–711. https://doi.org/10.1136/jnnp.34.6.699
Vagg R, Mogyoros I, Kiernan MC, Burke D (1998) Activity-dependent hyperpolarization of human motor axons produced by natural activity. J Physiol 507(3):919–925. https://doi.org/10.1111/j.1469-7793.1998.919bs.x
Vila-Chã C, Falla D, Correia MV, Farina D (2012) Changes in H reflex and V wave following short-term endurance and strength training. J Appl Physiol (bethesda, Md.: 1985) 112(1):54–63. https://doi.org/10.1152/japplphysiol.00802.2011
Waxman SG (1980) Determinants of conduction velocity in myelinated nerve fibers. Muscle Nerve 3(2):141–150. https://doi.org/10.1002/mus.880030207
Willis WD (2006) John Eccles’ studies of spinal cord presynaptic inhibition. Prog Neurobiol 78(3):189–214. https://doi.org/10.1016/j.pneurobio.2006.02.007
Zehr EP (2002) Considerations for use of the Hoffmann reflex in exercise studies. Eur J Appl Physiol 86(6):455–468. https://doi.org/10.1007/s00421-002-0577-5
Funding
Funding was provided by Fonds De La Recherche Scientifique—FNRS, ERA-Net Neuron.
Author information
Authors and Affiliations
Contributions
All authors made substantial contributions to the conception or design of the work, drafted the work or revised it critically for important intellectual content, approved the version to be published, and agree to be accountable for all aspects of the work in ensuring that questions related to the accuracy or integrity of any part of the work are appropriately investigated and resolved.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Additional information
Communicated by Michael I Lindinger.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Theodosiadou, A., Henry, M., Duchateau, J. et al. Revisiting the use of Hoffmann reflex in motor control research on humans. Eur J Appl Physiol 123, 695–710 (2023). https://doi.org/10.1007/s00421-022-05119-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-022-05119-7